
Enhanced Cytotoxic Activity of PEGylated Curcumin Derivatives: Synthesis, Structure–Activity Evaluation, and Biological Activity
 , , , , ,
, , , , , 

Abstract
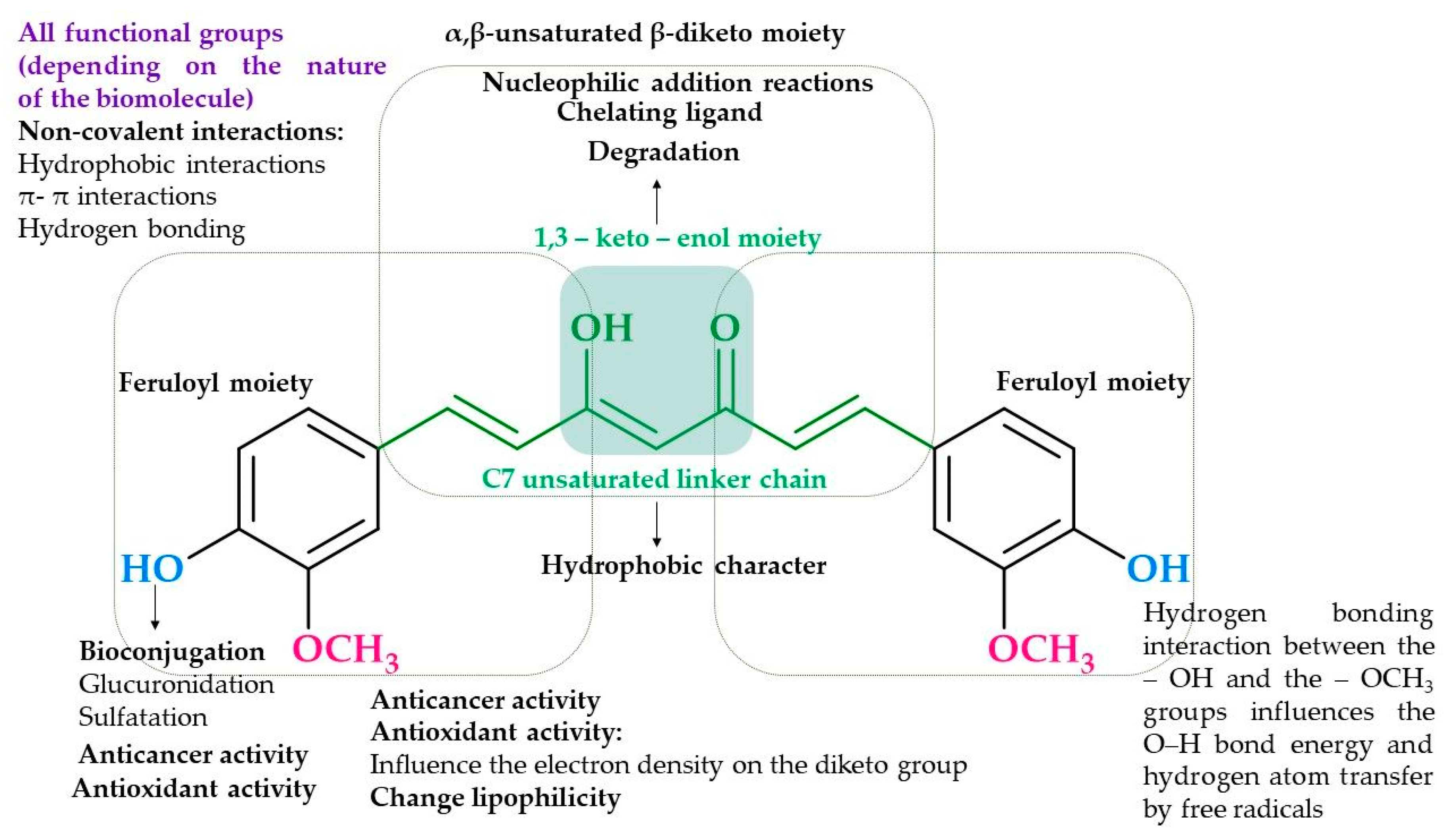
:1. Introduction
2. Results and Discussion
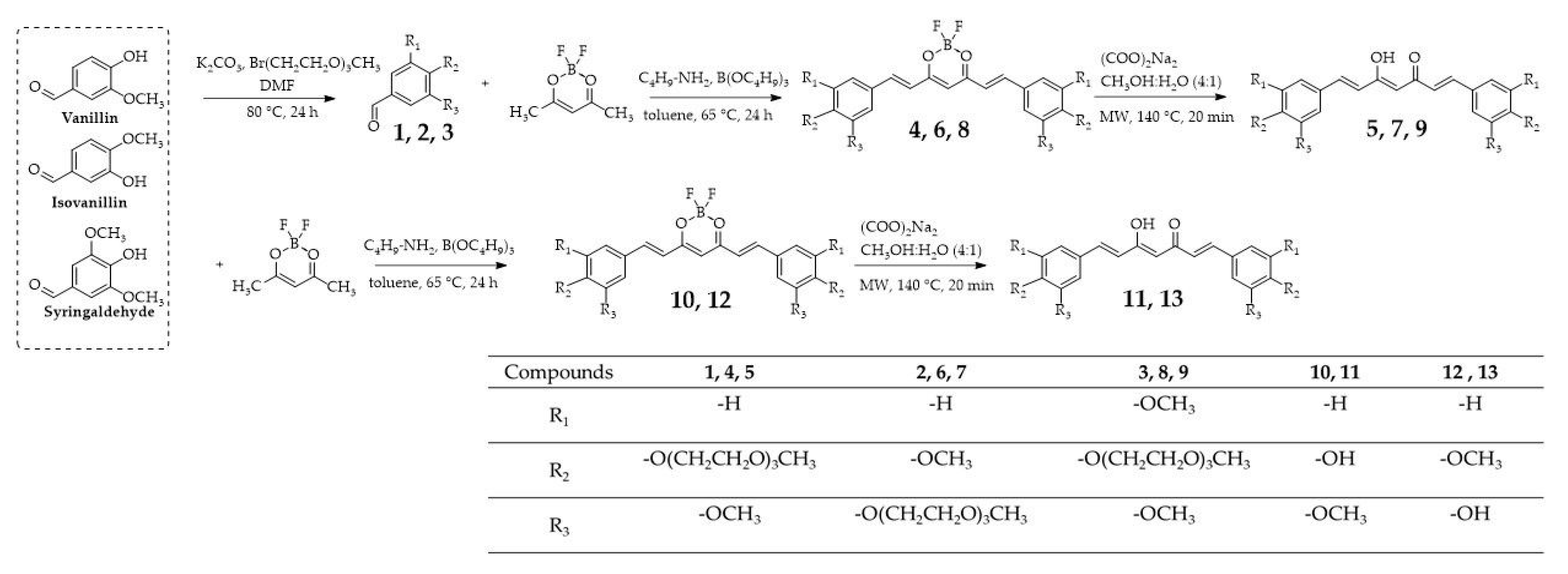
2.1. Synthesis
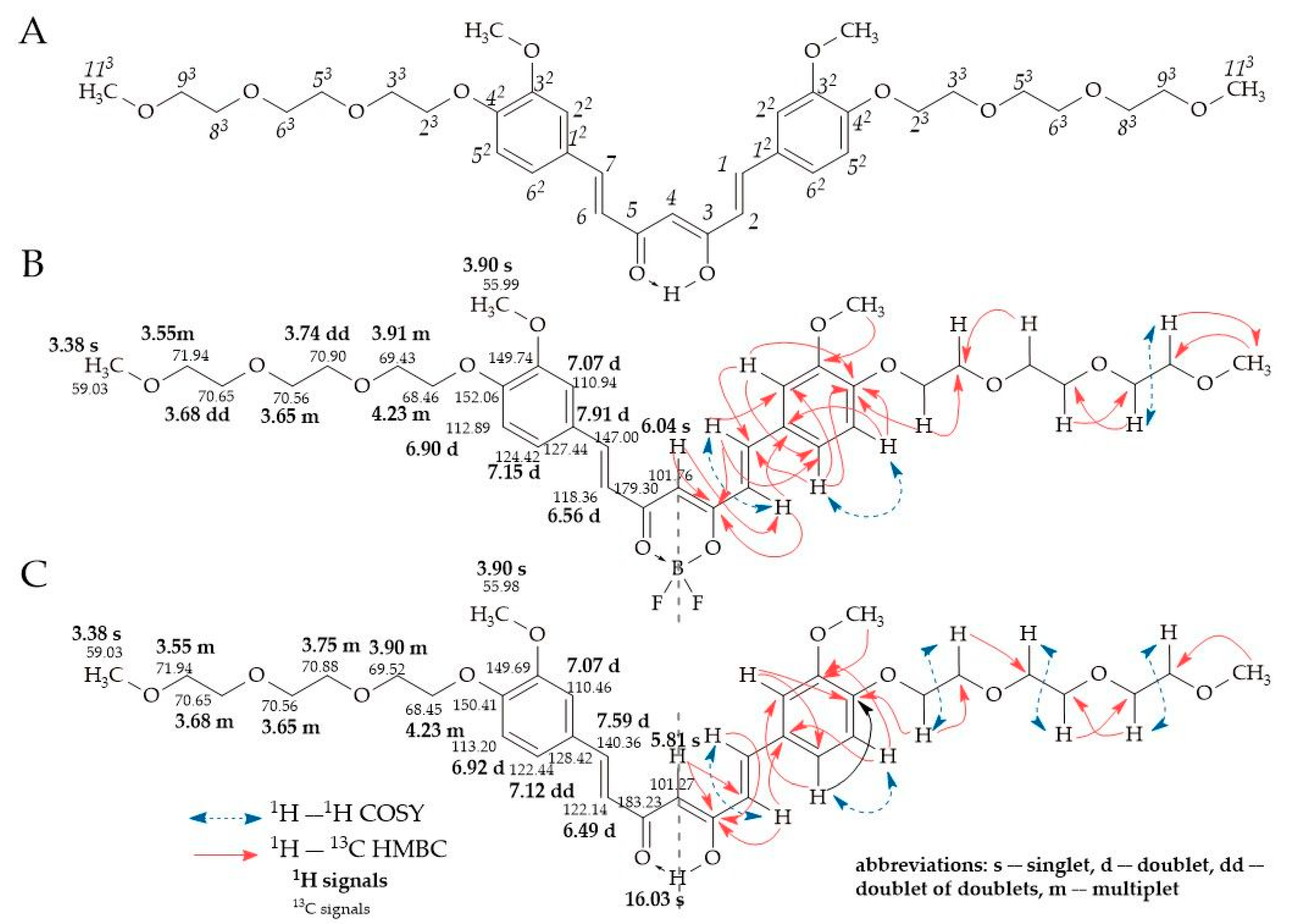
2.2. NMR Study
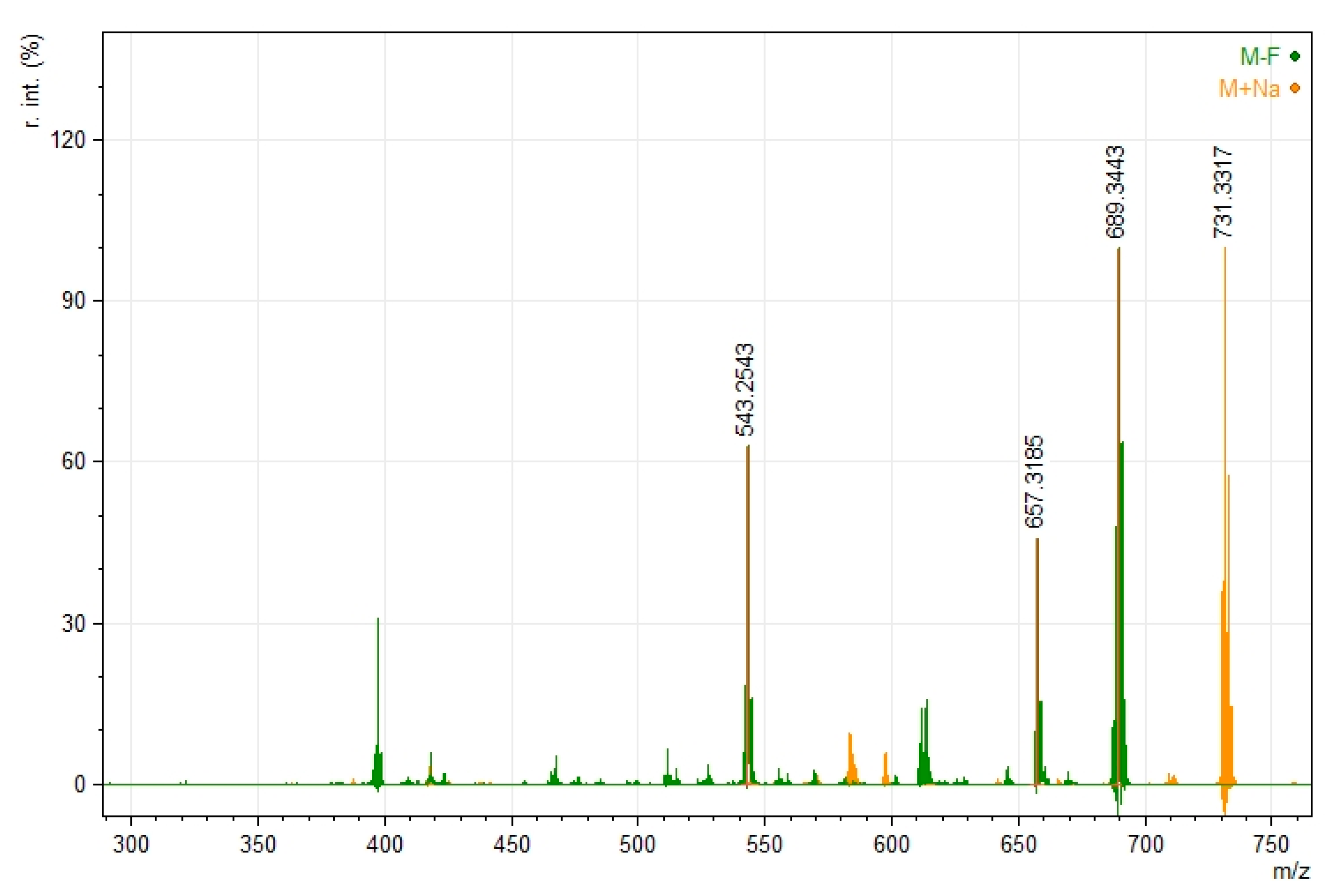
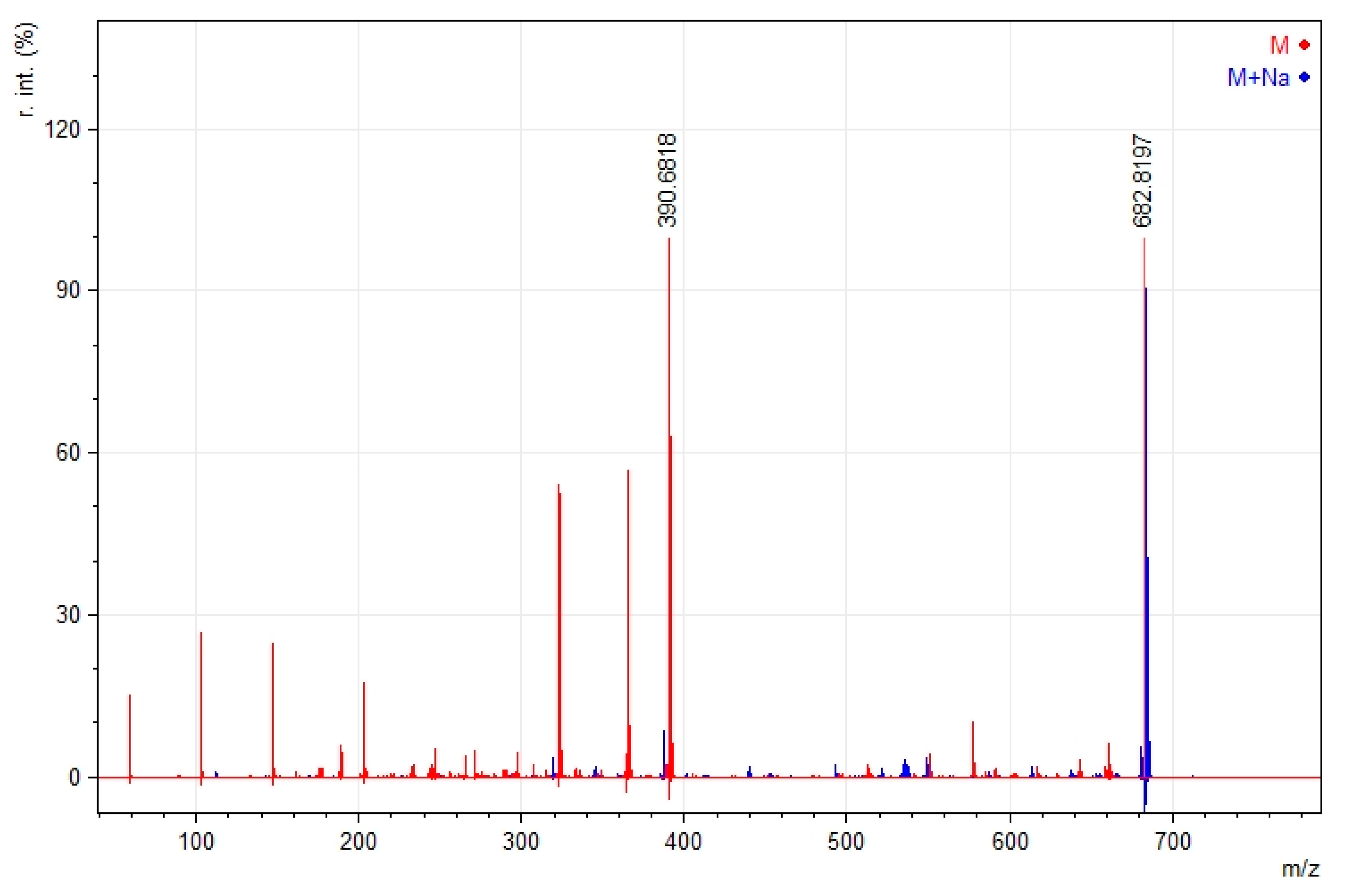
2.3. Mass Spectroscopy
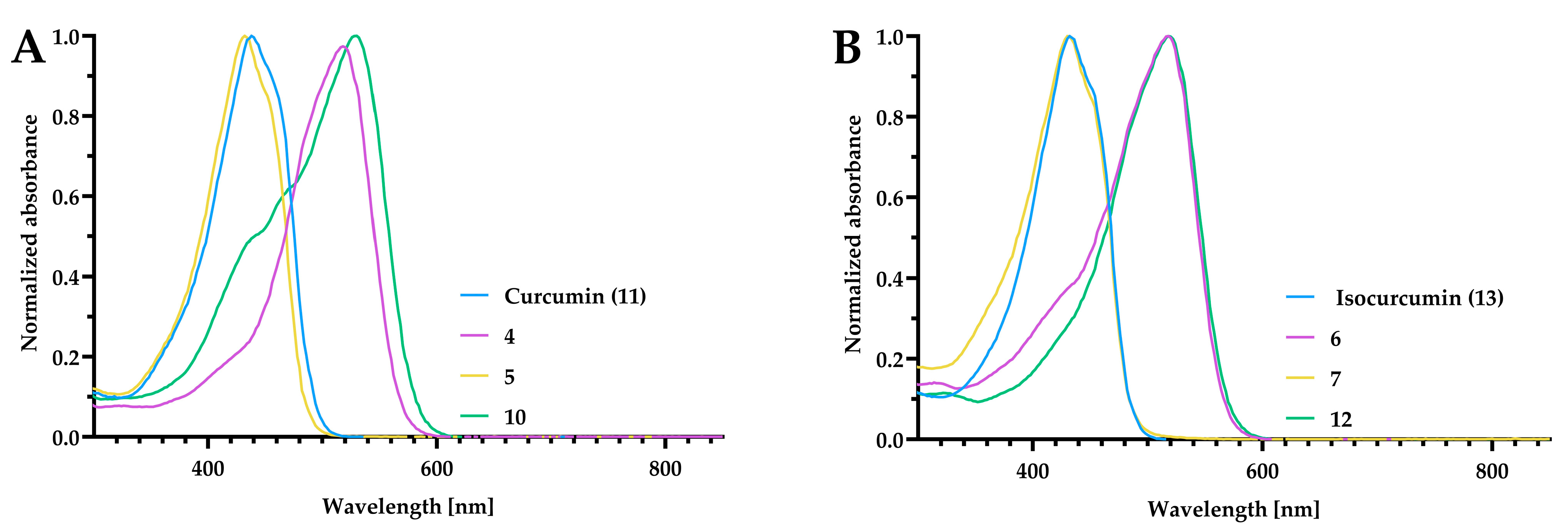
2.4. UV–Vis Study
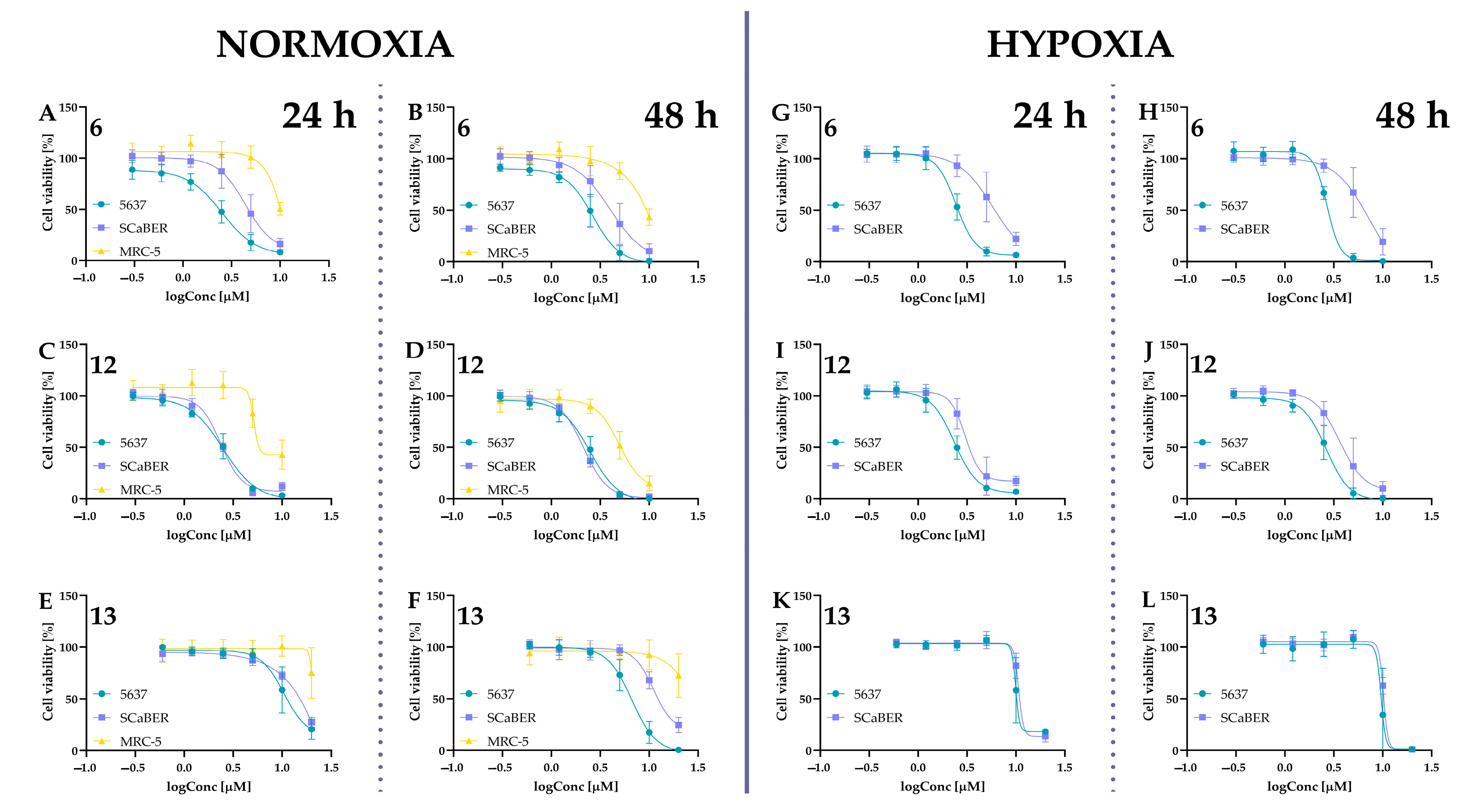
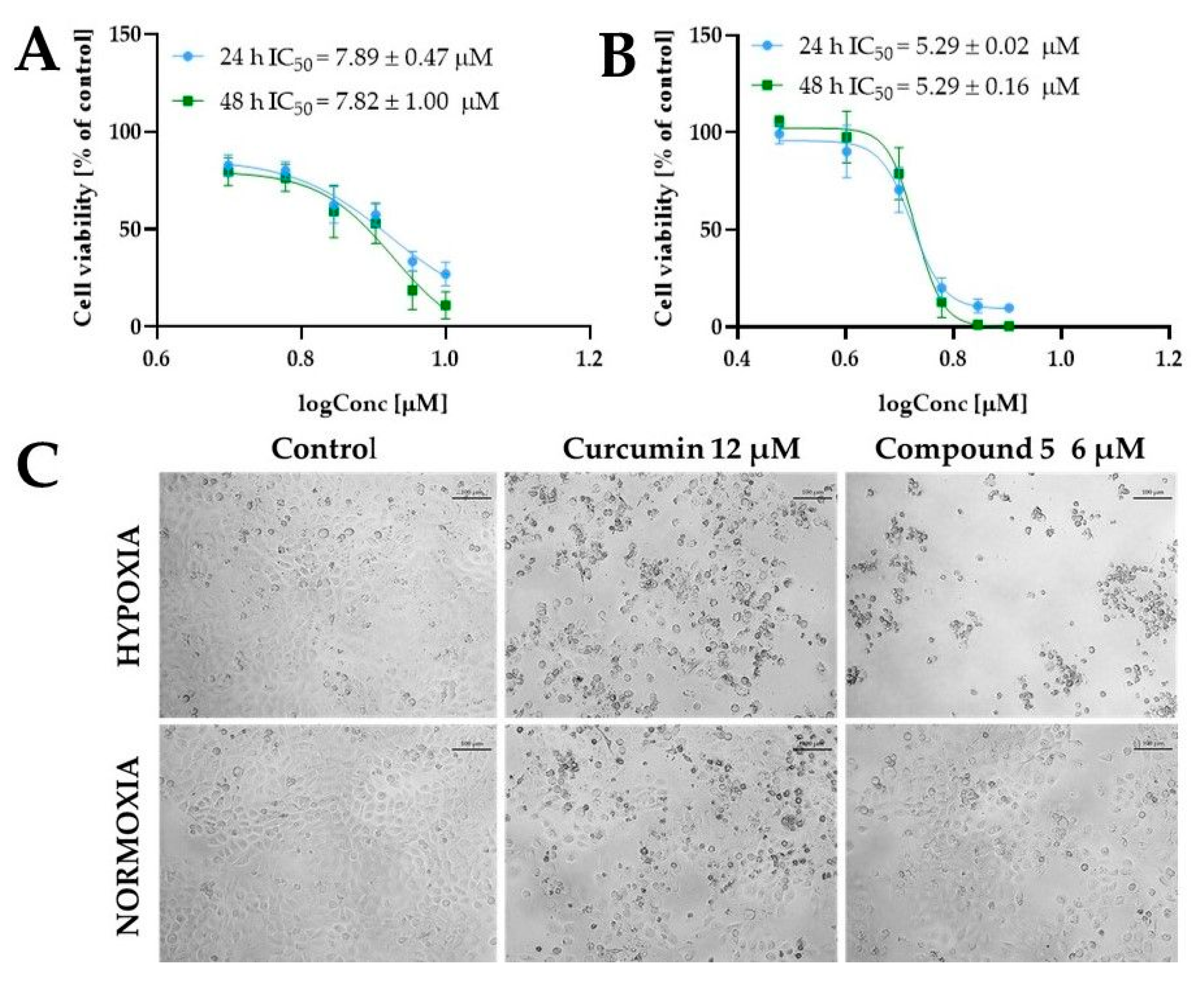
2.5. Biological Activity of Tested Compounds
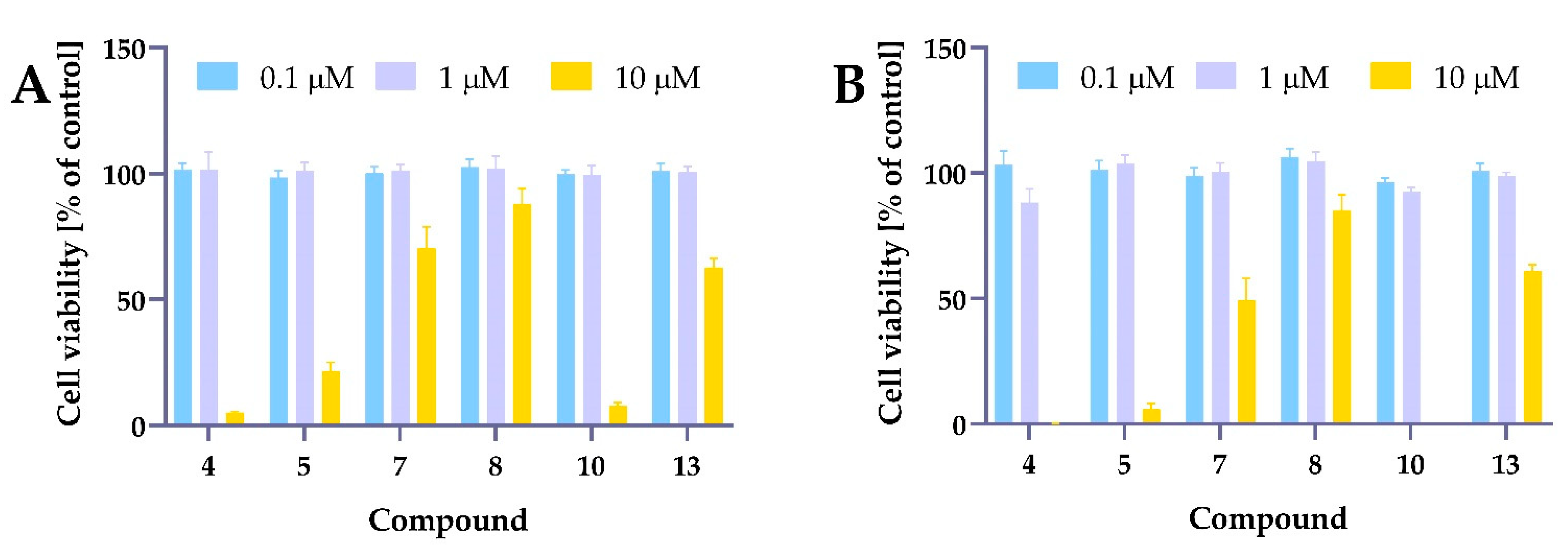
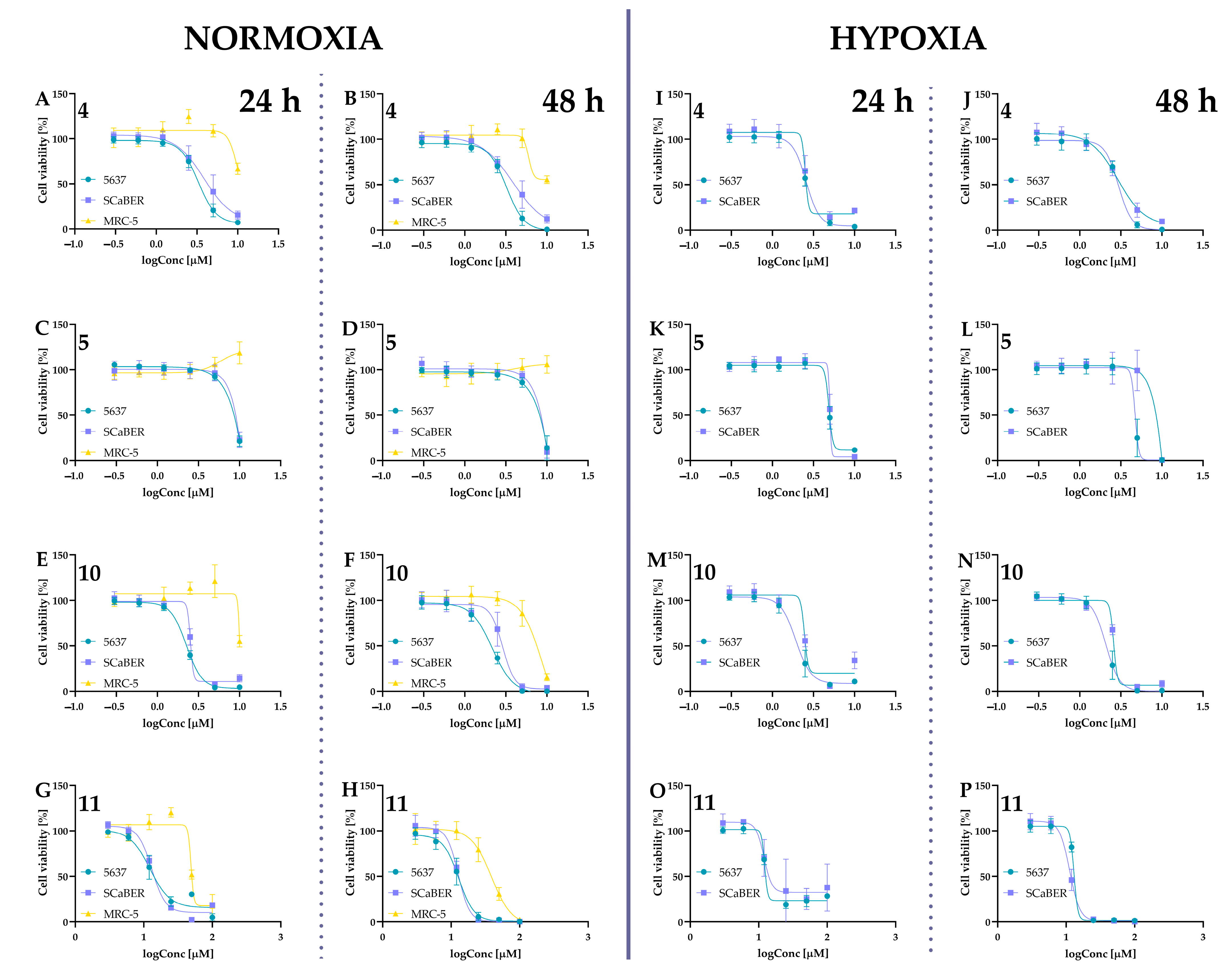
2.5.1. Cytotoxic Activity of Tested Compounds
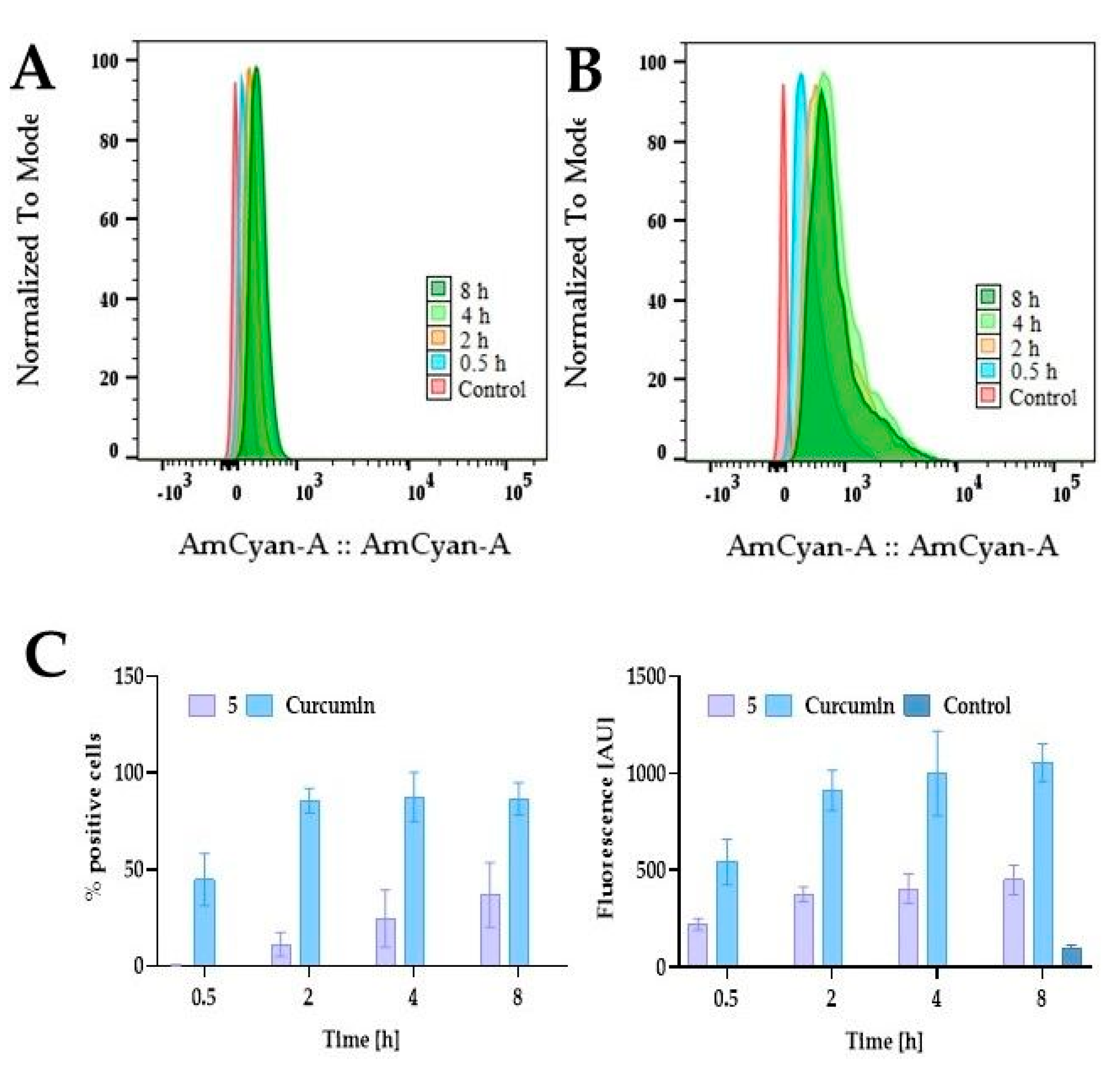
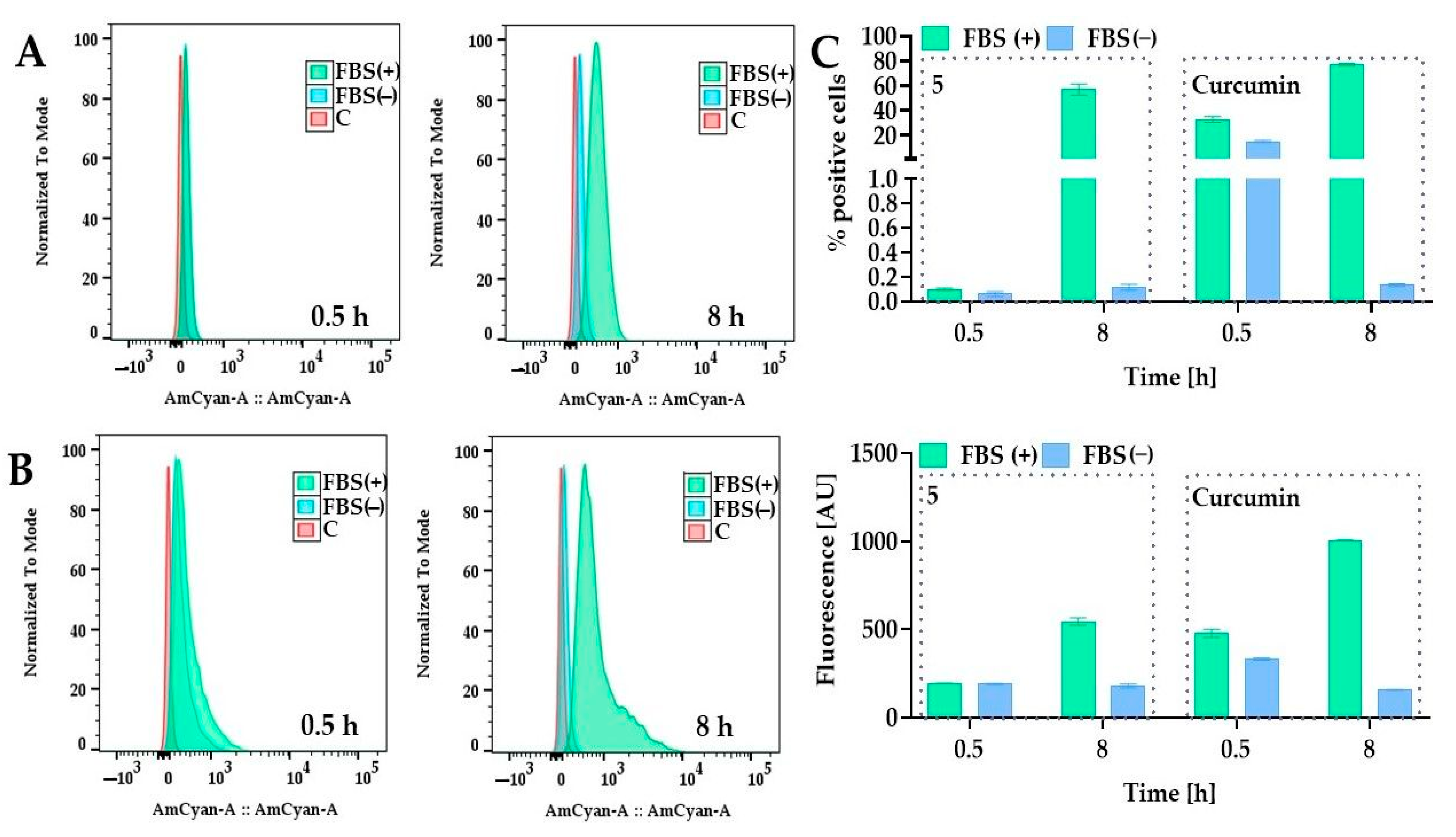
2.5.2. Cellular Uptake of PEGylated Curcumin
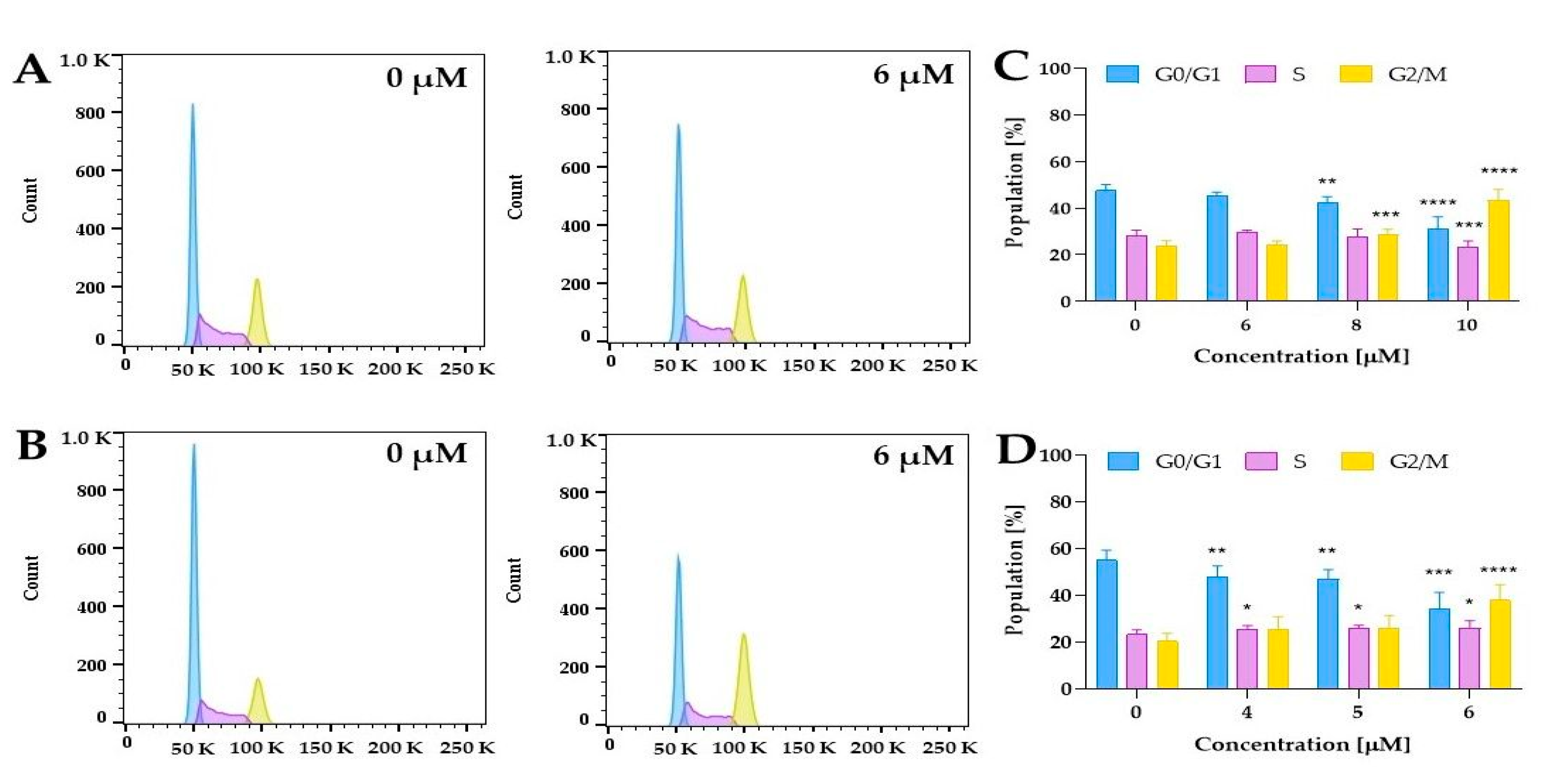
2.5.3. Cell Cycle Distribution
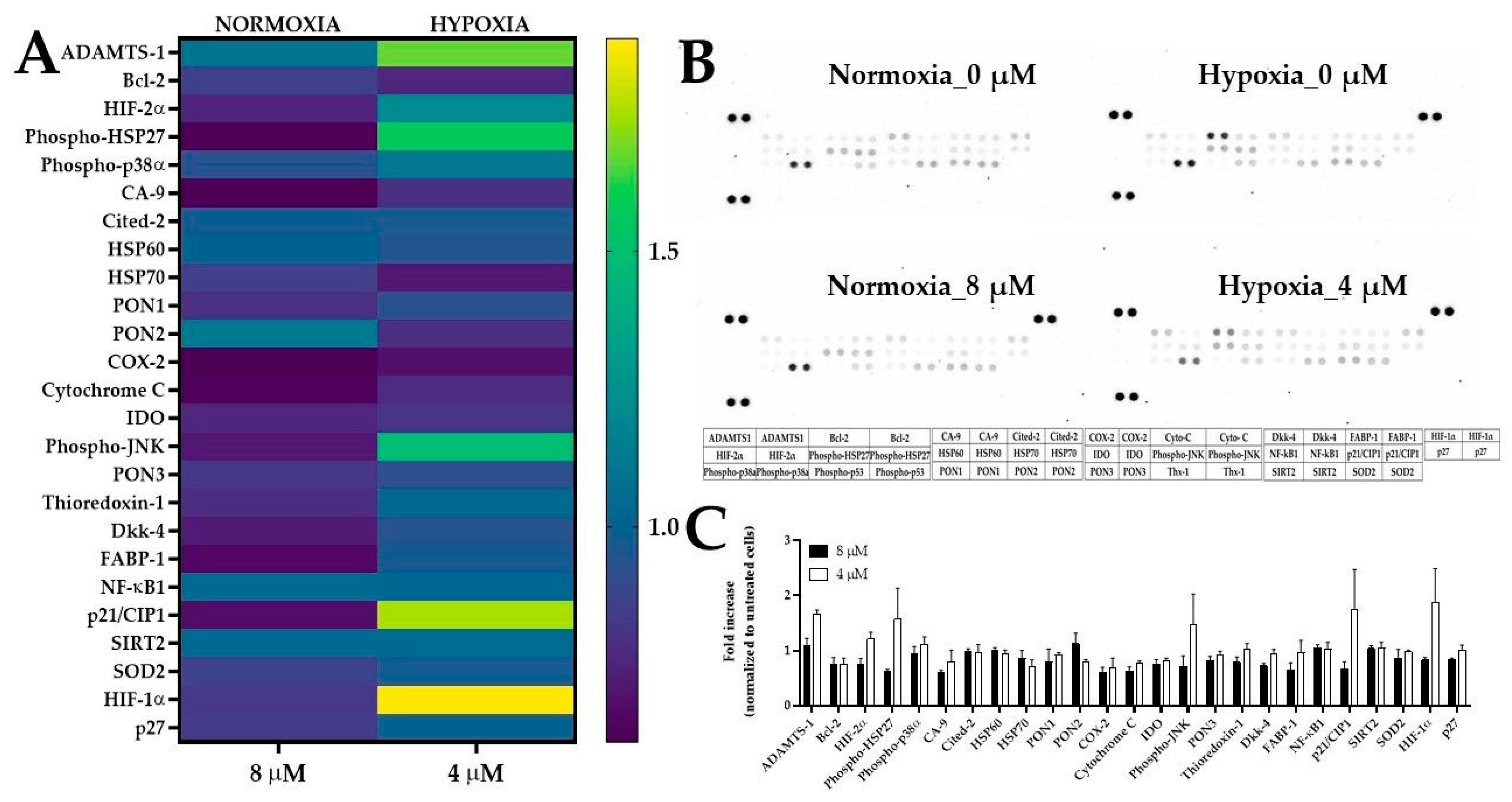
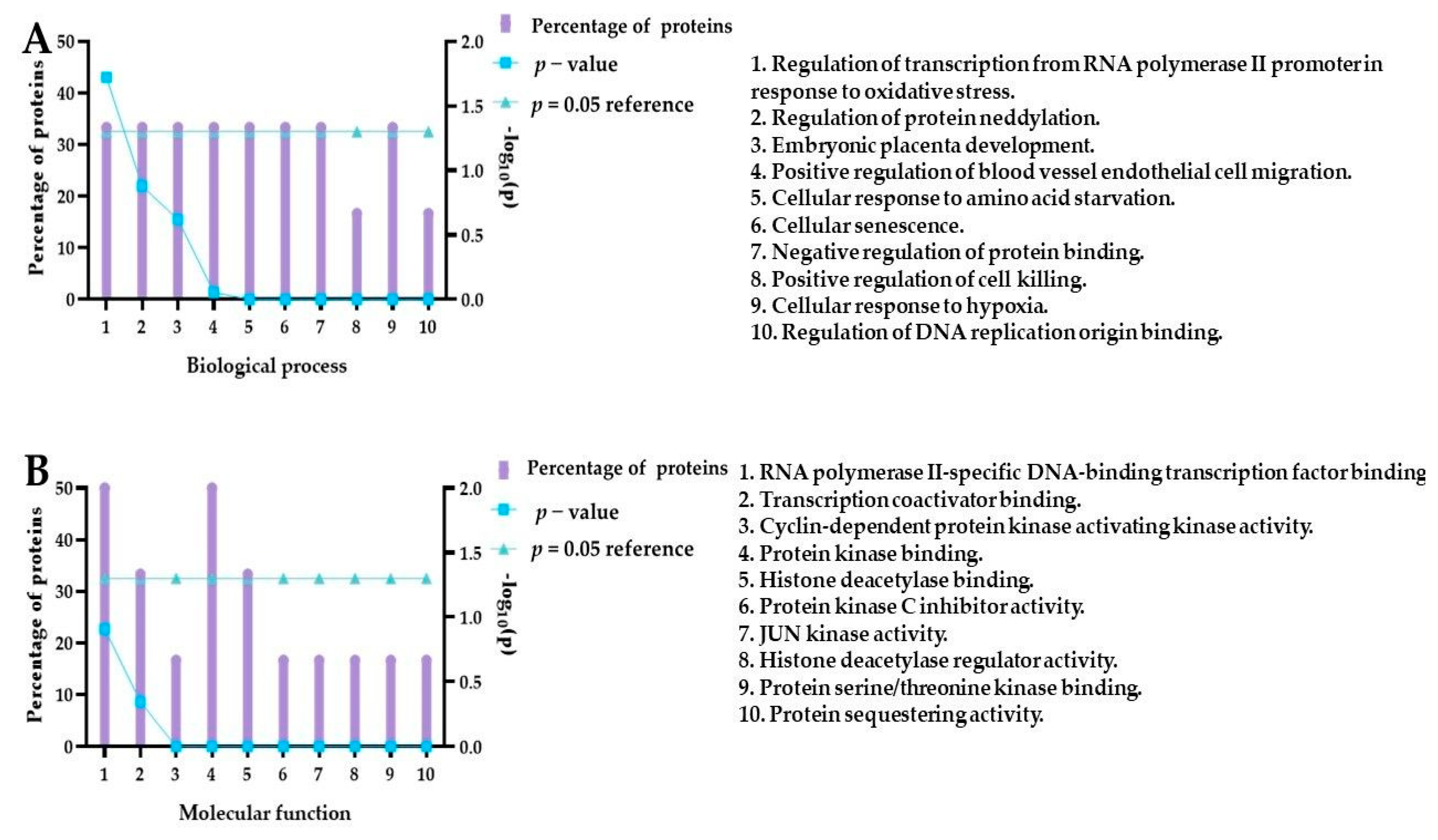
2.5.4. The Expression of Stress-Related Proteins after Treatment of 5637 Cells with PEGylated Curcumin
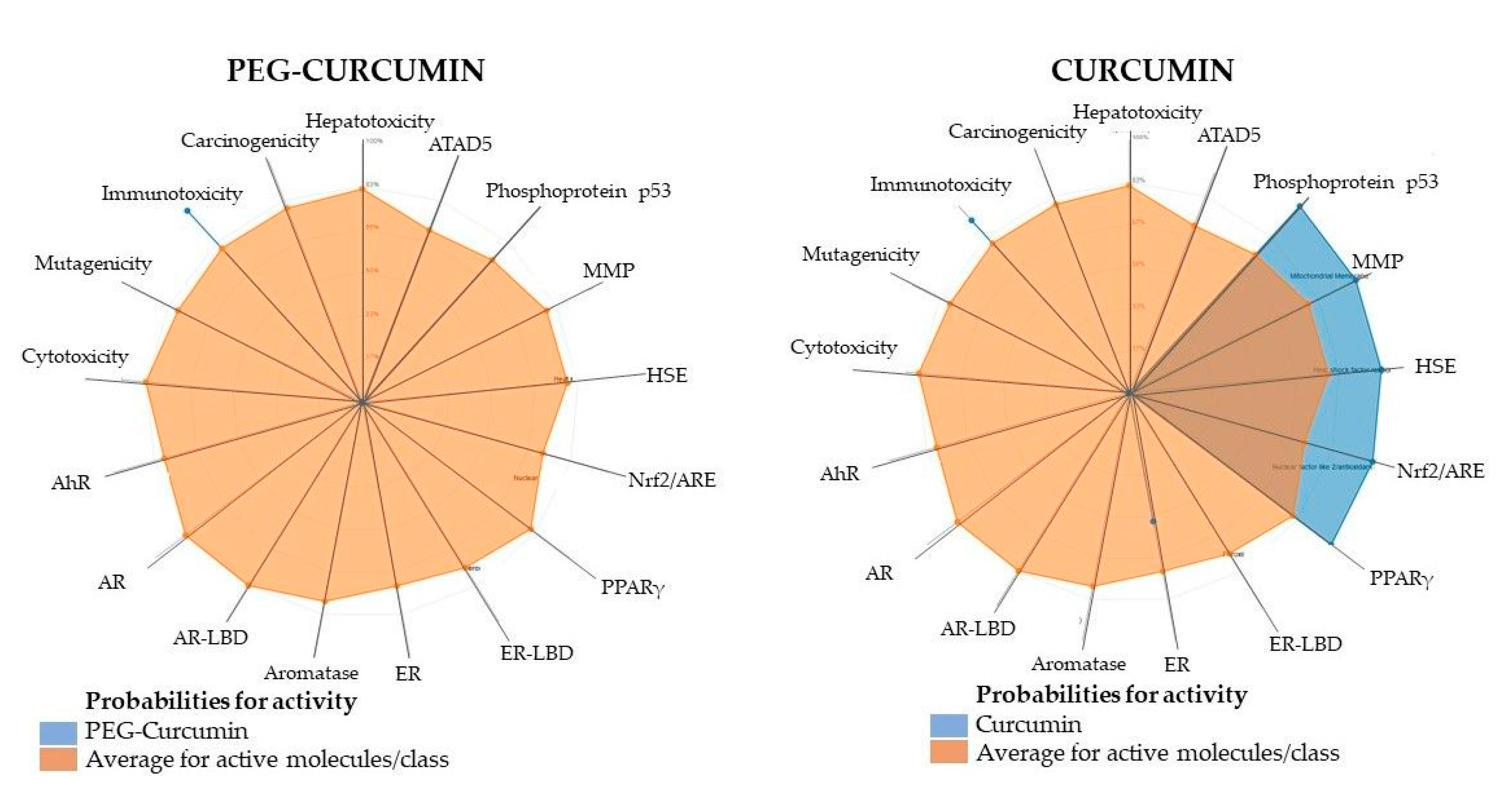
2.6. Computational Modeling to Predict ADMET Properties
2.7. Study Limitations and Future Directions
3. Materials and Methods
3.1. Materials
3.2. Synthesis
3.2.1. PEGylated Aldehyde Synthesis—General Procedure
3.2.2. Curcuminoids Synthesis
3.2.3. HPLC Purity
3.3. Cell Culture Conditions
3.4. In Vitro Anticancer Activity
3.5. Cellular Uptake
3.6. Cell Cycle Analysis
3.7. Proteome Profiler Array
3.8. Functional Enrichment Analysis
3.9. ADMET Properties Analysis Using the Computational Method
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, A.W.; Farooq, M.; Haseeb, M.; Choi, S. Role of Plant-Derived Active Constituents in Cancer Treatment and Their Mechanisms of Action. Cells 2022, 11, 1326. [Google Scholar] [CrossRef] [PubMed]
- Amalraj, A.; Pius, A.; Gopi, S.; Gopi, S. Biological Activities of Curcuminoids, Other Biomolecules from Turmeric and Their Derivatives—A Review. J. Tradit. Complement. Med. 2017, 7, 205–233. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Ji, X.; Zhang, Q.; Wei, Y. Curcumin Combined with Photodynamic Therapy, Promising Therapies for the Treatment of Cancer. Biomed. Pharmacother. 2022, 146, 112567. [Google Scholar] [CrossRef] [PubMed]
- Sandur, S.K.; Ichikawa, H.; Pandey, M.K.; Kunnumakkara, A.B.; Sung, B.; Sethi, G.; Aggarwal, B.B. Role of Pro-Oxidants and Antioxidants in the Anti-Inflammatory and Apoptotic Effects of Curcumin (Diferuloylmethane). Free Radic. Biol. Med. 2007, 43, 568–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doello, K.; Ortiz, R.; Alvarez, P.J.; Melguizo, C.; Cabeza, L.; Prados, J. Latest in Vitro and in Vivo Assay, Clinical Trials and Patents in Cancer Treatment Using Curcumin: A Literature Review. Nutr. Cancer 2018, 70, 569–578. [Google Scholar] [CrossRef]
- Xue, B.; Huang, J.; Zhang, H.; Li, B.; Xu, M.; Zhang, Y.; Xie, M.; Li, X. Micronized Curcumin Fabricated by Supercritical CO2 to Improve Antibacterial Activity against Pseudomonas Aeruginosa. Artif. Cells Nanomedicine Biotechnol. 2020, 48, 1135–1143. [Google Scholar] [CrossRef]
- Kamat, A.M.; Sethi, G.; Aggarwal, B.B. Curcumin Potentiates the Apoptotic Effects of Chemotherapeutic Agents and Cytokines through Down-Regulation of Nuclear Factor-KappaB and Nuclear Factor-KappaB-Regulated Gene Products in IFN-Alpha-Sensitive and IFN-Alpha-Resistant Human Bladder Cancer Cells. Mol. Cancer Ther. 2007, 6, 1022–1030. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.-H.; Dai, H.-P.; Shen, Q.; Ji, O.; Zhang, Q.; Zhai, Y.-L. Curcumin Induces Apoptosis and Suppresses Invasion through MAPK and MMP Signaling in Human Monocytic Leukemia SHI-1 Cells. Pharm. Biol. 2016, 54, 1303–1311. [Google Scholar] [CrossRef]
- Li, Y.; Sun, W.; Han, N.; Zou, Y.; Yin, D. Curcumin Inhibits Proliferation, Migration, Invasion and Promotes Apoptosis of Retinoblastoma Cell Lines through Modulation of MiR-99a and JAK/STAT Pathway. BMC Cancer 2018, 18, 1230. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Su, J.; Zhao, J.; Chen, J.; Cui, X.; Sun, M.; Zhang, X. Curcumin Inhibits Invasion and Metastasis of Human Hepatoma Cells through Bclaf1-Mediated Wnt/β-Catenin Signalling. Food Agric. Immunol. 2022, 33, 664–676. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, X.; Tan, J.; Tian, R.; Shen, P.; Cai, W.; Liao, H. Curcumin Suppresses the Stemness of Non-Small Cell Lung Cancer Cells via Promoting the Nuclear-Cytoplasm Translocation of TAZ. Environ. Toxicol. 2021, 36, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Sha, J.; Li, J.; Wang, W.; Pan, L.; Cheng, J.; Li, L.; Zhao, H.; Lin, W. Curcumin Induces G0/G1 Arrest and Apoptosis in Hormone Independent Prostate Cancer DU-145 Cells by down Regulating Notch Signaling. Biomed. Pharmacother. 2016, 84, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Borges, G.A.; Elias, S.T.; Amorim, B.; de Lima, C.L.; Coletta, R.D.; Castilho, R.M.; Squarize, C.H.; Guerra, E.N.S. Curcumin Downregulates the PI3K–AKT–MTOR Pathway and Inhibits Growth and Progression in Head and Neck Cancer Cells. Phytother. Res. 2020, 34, 3311–3324. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef] [PubMed]
- Peddi, P.F.; Hurvitz, S.A. Ado-Trastuzumab Emtansine (T-DM1) in Human Epidermal Growth Factor Receptor 2 (HER2)-Positive Metastatic Breast Cancer: Latest Evidence and Clinical Potential. Ther. Adv. Med. Oncol. 2014, 6, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Delgado, J.; Vleminckx, C.; Sarac, S.; Sosa, A.; Bergh, J.; Giuliani, R.; Enzmann, H.; Pignatti, F. The EMA Review of Trastuzumab Emtansine (T-DM1) for the Adjuvant Treatment of Adult Patients with HER2-Positive Early Breast Cancer. ESMO Open 2021, 6, 100074. [Google Scholar] [CrossRef]
- Priyadarsini, K.I. Chemical and Structural Features Influencing the Biological Activity of Curcumin. Curr. Pharm. Des. 2013, 19, 2093–2100. [Google Scholar] [CrossRef]
- Sanchez Armengol, E.; Unterweger, A.; Laffleur, F. PEGylated Drug Delivery Systems in the Pharmaceutical Field: Past, Present and Future Perspective. Drug Dev. Ind. Pharm. 2022, 48, 129–139. [Google Scholar] [CrossRef]
- Banerjee, S.S.; Aher, N.; Patil, R.; Khandare, J. Poly(Ethylene Glycol)-Prodrug Conjugates: Concept, Design, and Applications. J. Drug Deliv. 2012, 2012, 103973. [Google Scholar] [CrossRef] [Green Version]
- Freire Haddad, H.; Burke, J.A.; Scott, E.A.; Ameer, G.A. Clinical Relevance of Pre-Existing and Treatment-Induced Anti-Poly(Ethylene Glycol) Antibodies. Regen. Eng. Transl. Med. 2022, 8, 32–42. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, S.; Ge, L.; Wu, W.; Jiang, X. Translatable High Drug Loading Drug Delivery Systems Based on Biocompatible Polymer Nanocarriers. Biomacromolecules 2018, 19, 1732–1745. [Google Scholar] [CrossRef] [PubMed]
- Estapé Senti, M.; de Jongh, C.A.; Dijkxhoorn, K.; Verhoef, J.J.F.; Szebeni, J.; Storm, G.; Hack, C.E.; Schiffelers, R.M.; Fens, M.H.; Boross, P. Anti-PEG Antibodies Compromise the Integrity of PEGylated Lipid-Based Nanoparticles via Complement. J. Control. Release 2022, 341, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.; Ramadan, E.; Elsadek, N.E.; Emam, S.E.; Shimizu, T.; Ando, H.; Ishima, Y.; Elgarhy, O.H.; Sarhan, H.A.; Hussein, A.K.; et al. Polyethylene Glycol (PEG): The Nature, Immunogenicity, and Role in the Hypersensitivity of PEGylated Products. J. Control. Release 2022, 351, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Hardiansyah, A.; Yang, M.-C.; Liu, T.-Y.; Kuo, C.-Y.; Huang, L.-Y.; Chan, T.-Y. Hydrophobic Drug-Loaded PEGylated Magnetic Liposomes for Drug-Controlled Release. Nanoscale Res. Lett. 2017, 12, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sesarman, A.; Tefas, L.; Sylvester, B.; Licarete, E.; Rauca, V.; Luput, L.; Patras, L.; Porav, S.; Banciu, M.; Porfire, A. Co-Delivery of Curcumin and Doxorubicin in PEGylated Liposomes Favored the Antineoplastic C26 Murine Colon Carcinoma Microenvironment. Drug Deliv. Transl. Res. 2019, 9, 260–272. [Google Scholar] [CrossRef]
- Muddineti, O.S.; Vanaparthi, A.; Rompicharla, S.V.K.; Kumari, P.; Ghosh, B.; Biswas, S. Cholesterol and Vitamin E-Conjugated PEGylated Polymeric Micelles for Efficient Delivery and Enhanced Anticancer Activity of Curcumin: Evaluation in 2D Monolayers and 3D Spheroids. Artif. Cells Nanomedicine Biotechnol. 2018, 46, 773–786. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, D.; Zhong, W.; Kuang, S.; Luo, Q.; Song, L.; He, L.; Feng, X.; Tao, X. Evaluation of the PEG Density in the PEGylated Chitosan Nanoparticles as a Drug Carrier for Curcumin and Mitoxantrone. Nanomaterials 2018, 8, 486. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhan, P.; De Clercq, E.; Lou, H.; Liu, X. Current Drug Research on PEGylation with Small Molecular Agents. Prog. Polym. Sci. 2013, 38, 421–444. [Google Scholar] [CrossRef]
- Delgado, D.; Abonia, R. Synthetic Approaches for BF2-Containing Adducts of Outstanding Biological Potential. A Review. Arab. J. Chem. 2022, 15, 103528. [Google Scholar] [CrossRef]
- Bai, G.; Yu, C.; Cheng, C.; Hao, E.; Wei, Y.; Mu, X.; Jiao, L. Syntheses and Photophysical Properties of BF2 Complexes of Curcumin Analogues. Org. Biomol. Chem. 2014, 12, 1618–1626. [Google Scholar] [CrossRef]
- Gál, E.; Nagy, L.C. Photophysical Properties and Electronic Structure of Symmetrical Curcumin Analogues and Their BF2 Complexes, Including a Phenothiazine Substituted Derivative. Symmetry 2021, 13, 2299. [Google Scholar] [CrossRef]
- Laali, K.K.; Rathman, B.M.; Bunge, S.D.; Qi, X.; Borosky, G.L. Fluoro-Curcuminoids and Curcuminoid-BF2 Adducts: Synthesis, X-Ray Structures, Bioassay, and Computational/Docking Study. J. Fluor. Chem. 2016, 191, 29–41. [Google Scholar] [CrossRef]
- Liu, K.; Chen, J.; Chojnacki, J.; Zhang, S. BF3·OEt2-Promoted Concise Synthesis of Difluoroboron-Derivatized Curcumins from Aldehydes and 2,4-Pentanedione. Tetrahedron Lett. 2013, 54, 2070–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abonia, R.; Laali, K.K.; Raja Somu, D.; Bunge, S.D.; Wang, E.C. A Flexible Strategy for Modular Synthesis of Curcuminoid-BF2/Curcuminoid Pairs and Their Comparative Antiproliferative Activity in Human Cancer Cell Lines. ChemMedChem 2020, 15, 354–362. [Google Scholar] [CrossRef]
- Pabon, H.J.J. A Synthesis of Curcumin and Related Compounds. Recl. Trav. Chim. Pays-Bas 1964, 83, 379–386. [Google Scholar] [CrossRef]
- Cheng, F.; Chen, Y.; Zhan, Z.; Liu, Y.; Hu, P.; Ren, H.; Tang, H.; Peng, M. Curc-MPEG454, a PEGylated Curcumin Derivative, Improves Anti-Inflammatory and Antioxidant Activities: A Comparative Study. Inflammation 2018, 41, 579–594. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Yang, C.; Wang, P.; Oelschlager, D.K.; Zheng, Y.; Tian, D.-A.; Grizzle, W.E.; Buchsbaum, D.J.; Wan, M. Polyethylene Glycosylated Curcumin Conjugate Inhibits Pancreatic Cancer Cell Growth through Inactivation of Jab1. Mol. Pharmacol. 2009, 76, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Deters, M.; Knochenwefel, H.; Lindhorst, D.; Koal, T.; Meyer, H.H.; Hänsel, W.; Resch, K.; Kaever, V. Different Curcuminoids Inhibit T-Lymphocyte Proliferation Independently of Their Radical Scavenging Activities. Pharm. Res. 2008, 25, 1822–1827. [Google Scholar] [CrossRef]
- Nini, A.; Hoffmann, M.J.; Lampignano, R.; große Siemer, R.; van Dalum, G.; Szarvas, T.; Cotarelo, C.L.; Schulz, W.A.; Niederacher, D.; Neubauer, H.; et al. Evaluation of HER2 Expression in Urothelial Carcinoma Cells as a Biomarker for Circulating Tumor Cells. Cytometry B Clin. Cytom. 2020, 98, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Erez, Y.; Simkovitch, R.; Shomer, S.; Gepshtein, R.; Huppert, D. Effect of Acid on the Ultraviolet–Visible Absorption and Emission Properties of Curcumin. J. Phys. Chem. A 2014, 118, 872–884. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Pan, M.-H.; Cheng, A.-L.; Lin, L.-I.; Ho, Y.-S.; Hsieh, C.-Y.; Lin, J.-K. Stability of Curcumin in Buffer Solutions and Characterization of Its Degradation Products. J. Pharm. Biomed. Anal. 1997, 15, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Walkey, C.; Chan, W.C.W. Polyethylene Glycol Backfilling Mitigates the Negative Impact of the Protein Corona on Nanoparticle Cell Targeting. Angew. Chem. Int. Ed. 2014, 53, 5093–5096. [Google Scholar] [CrossRef] [PubMed]
- Verhoef, J.J.F.; Anchordoquy, T.J. Questioning the Use of PEGylation for Drug Delivery. Drug Deliv. Transl. Res. 2013, 3, 499–503. [Google Scholar] [CrossRef]
- Pozzi, D.; Caracciolo, G.; Digiacomo, L.; Colapicchioni, V.; Palchetti, S.; Capriotti, A.L.; Cavaliere, C.; Chiozzi, R.Z.; Puglisi, A.; Laganà, A. The Biomolecular Corona of Nanoparticles in Circulating Biological Media. Nanoscale 2015, 7, 13958–13966. [Google Scholar] [CrossRef]
- Pelaz, B.; del Pino, P.; Maffre, P.; Hartmann, R.; Gallego, M.; Rivera-Fernández, S.; de la Fuente, J.M.; Nienhaus, G.U.; Parak, W.J. Surface Functionalization of Nanoparticles with Polyethylene Glycol: Effects on Protein Adsorption and Cellular Uptake. ACS Nano 2015, 9, 6996–7008. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, J.; Zhao, M.; Tang, S.; Cheng, X.; Zhang, W.; Li, W.; Liu, X.; Peng, H.; Wang, Q. Effects of Polyethylene Glycol on the Surface of Nanoparticles for Targeted Drug Delivery. Nanoscale 2021, 13, 10748–10764. [Google Scholar] [CrossRef]
- Bekale, L.; Agudelo, D.; Tajmir-Riahi, H.A. The Role of Polymer Size and Hydrophobic End-Group in PEG–Protein Interaction. Colloids Surf. B Biointerfaces 2015, 130, 141–148. [Google Scholar] [CrossRef]
- Wang, T.; Guo, Y.; He, Y.; Ren, T.; Yin, L.; Fawcett, J.P.; Gu, J.; Sun, H. Impact of Molecular Weight on the Mechanism of Cellular Uptake of Polyethylene Glycols (PEGs) with Particular Reference to P-Glycoprotein. Acta Pharm. Sin. B 2020, 10, 2002–2009. [Google Scholar] [CrossRef]
- Hugger, E.D.; Novak, B.L.; Burton, P.S.; Audus, K.L.; Borchardt, R.T. A Comparison of Commonly Used Polyethoxylated Pharmaceutical Excipients on Their Ability to Inhibit P-glycoprotein Activity in Vitro. J. Pharm. Sci. 2002, 91, 1991–2002. [Google Scholar] [CrossRef]
- Shen, Q.; Lin, Y.; Handa, T.; Doi, M.; Sugie, M.; Wakayama, K.; Okada, N.; Fujita, T.; Yamamoto, A. Modulation of Intestinal P-Glycoprotein Function by Polyethylene Glycols and Their Derivatives by In Vitro Transport and In Situ Absorption Studies. Int. J. Pharm. 2006, 313, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Snyder, S.; Murundi, S.; Crawford, L.; Putnam, D. Enabling P-Glycoprotein Inhibition in Multidrug Resistant Cancer through the Reverse Targeting of a Quinidine-PEG Conjugate. J. Control. Release 2020, 317, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, G.; Zhang, R.; Dong, L.; Chen, H.; Bo, J.; Xue, W.; Huang, Y. Curcumin Inhibits Cell Proliferation and Motility via Suppression of TROP2 in Bladder Cancer Cells. Int. J. Oncol. 2018, 53, 515–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.; Kim, G.Y.; Kim, G.D.; Choi, B.T.; Park, Y.-M.; Choi, Y.H. Induction of G2/M Arrest and Inhibition of Cyclooxygenase-2 Activity by Curcumin in Human Bladder Cancer T24 Cells. Oncol. Rep. 2006, 15, 1225–1231. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.; Ho, J.-N.; Kook, H.R.; Lee, S.; Oh, J.J.; Hong, S.K.; Lee, S.E.; Byun, S.-S. Theracurmin® Efficiently Inhibits the Growth of Human Prostate and Bladder Cancer Cells via Induction of Apoptotic Cell Death and Cell Cycle Arrest. Oncol. Rep. 2016, 35, 1463–1472. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Wang, Z.; Zhao, Y.; Wang, D.; Li, Y.; Ma, L.; Li, X.; Li, J.; Xiao, N.; Tian, J.; et al. Effects of Curcumin on Bladder Cancer Cells and Development of Urothelial Tumors in a Rat Bladder Carcinogenesis Model. Cancer Lett. 2008, 264, 299–308. [Google Scholar] [CrossRef]
- Novak Kujundžić, R.; Grbeša, I.; Ivkić, M.; Katdare, M.; Gall-Trošelj, K. Curcumin Downregulates H19 Gene Transcription in Tumor Cells. J. Cell. Biochem. 2008, 104, 1781–1792. [Google Scholar] [CrossRef] [PubMed]
- Mondal, N.; Parvin, J.D. DNA Topoisomerase IIalpha Is Required for RNA Polymerase II Transcription on Chromatin Templates. Nature 2001, 413, 435–438. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, W.; Sun, Y.; Jia, L. Protein Neddylation and Its Alterations in Human Cancers for Targeted Therapy. Cell. Signal. 2018, 44, 92–102. [Google Scholar] [CrossRef]
- Lobato-Gil, S.; Heidelberger, J.B.; Maghames, C.; Bailly, A.; Brunello, L.; Rodriguez, M.S.; Beli, P.; Xirodimas, D.P. Proteome-Wide Identification of NEDD8 Modification Sites Reveals Distinct Proteomes for Canonical and Atypical NEDDylation. Cell Rep. 2021, 34, 108635. [Google Scholar] [CrossRef]
- Gai, W.; Peng, Z.; Liu, C.H.; Zhang, L.; Jiang, H. Advances in Cancer Treatment by Targeting the Neddylation Pathway. Front. Cell Dev. Biol. 2021, 9, 653882. [Google Scholar] [CrossRef]
- Yu, Q.; Jiang, Y.; Sun, Y. Anticancer Drug Discovery by Targeting Cullin Neddylation. Acta Pharm. Sin. B 2020, 10, 746–765. [Google Scholar] [CrossRef] [PubMed]
- Berse, M.; Bounpheng, M.; Huang, X.; Christy, B.; Pollmann, C.; Dubiel, W. Ubiquitin-Dependent Degradation of Id1 and Id3 Is Mediated by the COP9 Signalosome. J. Mol. Biol. 2004, 343, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Uhle, S.; Medalia, O.; Waldron, R.; Dumdey, R.; Henklein, P.; Bech-Otschir, D.; Huang, X.; Berse, M.; Sperling, J.; Schade, R.; et al. Protein Kinase CK2 and Protein Kinase D Are Associated with the COP9 Signalosome. EMBO J. 2003, 22, 1302–1312. [Google Scholar] [CrossRef] [Green Version]
- Füllbeck, M.; Huang, X.; Dumdey, R.; Frommel, C.; Dubiel, W.; Preissner, R. Novel Curcumin- and Emodin-Related Compounds Identified by in Silico 2D/3D Conformer Screening Induce Apoptosis in Tumor Cells. BMC Cancer 2005, 5, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Wang, M.; Bu, X.; Zuo, Y.; Wang, S.; Wang, D.; Liu, Q.; Su, B.; Xu, T.; Wang, C.; et al. Curcumin Analogue T83 Exhibits Potent Antitumor Activity and Induces Radiosensitivity through Inactivation of Jab1 in Nasopharyngeal Carcinoma. BMC Cancer 2013, 13, 323. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Wang, D.; Liu, G.; Pan, Y. Jab1/Cops5: A Promising Target for Cancer Diagnosis and Therapy. Int. J. Clin. Oncol. 2021, 26, 1159–1169. [Google Scholar] [CrossRef]
- Al Bitar, S.; Gali-Muhtasib, H. The Role of the Cyclin Dependent Kinase Inhibitor P21cip1/Waf1 in Targeting Cancer: Molecular Mechanisms and Novel Therapeutics. Cancers 2019, 11, 1475. [Google Scholar] [CrossRef] [Green Version]
- Aljabery, F.; Shabo, I.; Gimm, O.; Jahnson, S.; Olsson, H. The Expression Profile of P14, P53 and P21 in Tumour Cells Is Associated with Disease-Specific Survival and the Outcome of Postoperative Chemotherapy Treatment in Muscle-Invasive Bladder Cancer. Urol. Oncol. Semin. Orig. Investig. 2018, 36, 530.e7–530.e18. [Google Scholar] [CrossRef]
- Tang, K.; Wang, C.; Chen, Z.; Xu, H.; Ye, Z. Clinicopathologic and Prognostic Significance of P21 (Cip1/Waf1) Expression in Bladder Cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 4999–5007. [Google Scholar]
- Karimian, A.; Ahmadi, Y.; Yousefi, B. Multiple Functions of P21 in Cell Cycle, Apoptosis and Transcriptional Regulation after DNA Damage. DNA Repair 2016, 42, 63–71. [Google Scholar] [CrossRef]
- Koshiji, M.; Kageyama, Y.; Pete, E.A.; Horikawa, I.; Barrett, J.C.; Huang, L.E. HIF-1α Induces Cell Cycle Arrest by Functionally Counteracting Myc. EMBO J. 2004, 23, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Adachi, S.; Yasuda, I.; Yamauchi, T.; Kawaguchi, J.; Itani, M.; Yoshioka, T.; Matsushima-Nishiwaki, R.; Hirose, Y.; Kozawa, O.; et al. Phosphorylation Status of Heat Shock Protein 27 Plays a Key Role in Gemcitabine-Induced Apoptosis of Pancreatic Cancer Cells. Cancer Lett. 2011, 313, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.-H.; Huang, C.-Y.; Chou, C.-W.; Makondi, P.T.; Huang, M.-T.; Wei, P.-L.; Chang, Y.-J. Heat Shock Protein 27 Influences the Anti-Cancer Effect of Curcumin in Colon Cancer Cells through ROS Production and Autophagy Activation. Life Sci. 2018, 209, 43–51. [Google Scholar] [CrossRef] [PubMed]
- De Arao Tan, I.; Ricciardelli, C.; Russell, D.L. The Metalloproteinase ADAMTS1: A Comprehensive Review of Its Role in Tumorigenic and Metastatic Pathways. Int. J. Cancer 2013, 133, 2263–2276. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Khalil, R.A. A Disintegrin and Metalloproteinase (ADAM) and ADAM with Thrombospondin Motifs (ADAMTS) Family in Vascular Biology and Disease. Biochem. Pharmacol. 2019, 164, 188–204. [Google Scholar] [CrossRef]
- Fontanil, T.; Mohamedi, Y.; Cobo, T.; Cal, S.; Obaya, Á.J. Novel Associations Within the Tumor Microenvironment: Fibulins Meet ADAMTSs. Front. Oncol. 2019, 9, 796. [Google Scholar] [CrossRef]
- Freitas, V.M.; do Amaral, J.B.; Silva, T.A.; Santos, E.S.; Mangone, F.R.; Pinheiro, J.D.J.; Jaeger, R.G.; Nagai, M.A.; Machado-Santelli, G.M. Decreased Expression of ADAMTS-1 in Human Breast Tumors Stimulates Migration and Invasion. Mol. Cancer 2013, 12, 2. [Google Scholar] [CrossRef]
- Rocks, N.; Paulissen, G.; Quesada-Calvo, F.; Munaut, C.; Gonzalez, M.-L.A.; Gueders, M.; Hacha, J.; Gilles, C.; Foidart, J.-M.; Noel, A.; et al. ADAMTS-1 Metalloproteinase Promotes Tumor Development through the Induction of a Stromal Reaction in Vivo. Cancer Res. 2008, 68, 9541–9550. [Google Scholar] [CrossRef] [Green Version]
- De Assis Lima, M.; da Silva, S.V.; Serrano-Garrido, O.; Hülsemann, M.; Santos Neres, L.; Rodríguez-Manzaneque, J.C.; Hodgson, L.; Freitas, V.M. Metalloprotease ADAMTS-1 Decreases Cell Migration and Invasion Modulating the Spatiotemporal Dynamics of Cdc42 Activity. Cell. Signal. 2021, 77, 109827. [Google Scholar] [CrossRef]
- Wang, B.; Chen, S.; Zhao, J.-Q.; Xiang, B.-L.; Gu, X.; Zou, F.; Zhang, Z.-H. ADAMTS-1 Inhibits Angiogenesis via the PI3K/Akt-ENOS-VEGF Pathway in Lung Cancer Cells. Transl. Cancer Res. 2019, 8, 2725. [Google Scholar] [CrossRef]
- Obika, M.; Ogawa, H.; Takahashi, K.; Li, J.; Hatipoglu, O.F.; Cilek, M.Z.; Miyoshi, T.; Inagaki, J.; Ohtsuki, T.; Kusachi, S.; et al. Tumor Growth Inhibitory Effect of ADAMTS1 Is Accompanied by the Inhibition of Tumor Angiogenesis. Cancer Sci. 2012, 103, 1889–1897. [Google Scholar] [CrossRef] [PubMed]
- Hatipoglu, O.F.; Hirohata, S.; Cilek, M.Z.; Ogawa, H.; Miyoshi, T.; Obika, M.; Demircan, K.; Shinohata, R.; Kusachi, S.; Ninomiya, Y. ADAMTS1 Is a Unique Hypoxic Early Response Gene Expressed by Endothelial Cells. J. Biol. Chem. 2009, 284, 16325–16333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesário, J.M.S.; Brito, R.B.O.; Malta, C.S.; Silva, C.S.; Matos, Y.S.T.; Kunz, T.C.M.; Urbano, J.J.; Oliveira, L.V.F.; Dalboni, M.A.; Dellê, H. A Simple Method to Induce Hypoxia-Induced Vascular Endothelial Growth Factor-A (VEGF-A) Expression in T24 Human Bladder Cancer Cells. In Vitro Cell. Dev. Biol. Anim. 2017, 53, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.; Li, M.; Xu, J.; Howell, D.C.; Li, Z.; Chen, F.-E. Recent Development on COX-2 Inhibitors as Promising Anti-Inflammatory Agents: The Past 10 Years. Acta Pharm. Sin. B 2022, 12, 2790–2807. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kalra, H.; Akundi, R.S. Extracellular ATP Mediates Cancer Cell Migration and Invasion Through Increased Expression of Cyclooxygenase 2. Front. Pharmacol. 2021, 11, 617211. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.R.; Pelly, V.S.; Moeini, A.; Chiang, S.-C.; Flanagan, E.; Bromley, C.P.; Clark, C.; Earnshaw, C.H.; Koufaki, M.A.; Bonavita, E.; et al. Chemotherapy-Induced COX-2 Upregulation by Cancer Cells Defines Their Inflammatory Properties and Limits the Efficacy of Chemoimmunotherapy Combinations. Nat. Commun. 2022, 13, 2063. [Google Scholar] [CrossRef]
- Boström, P.J.; Aaltonen, V.; Söderström, K.O.; Uotila, P.; Laato, M. Expression of Cyclooxygenase-1 and -2 in Urinary Bladder Carcinomas in Vivo and in Vitro and Prostaglandin E2 Synthesis in Cultured Bladder Cancer Cells. Pathology (Phila.) 2001, 33, 469–474. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 2012, 64, 4–17. [Google Scholar] [CrossRef]
- Cheng, F.; Li, W.; Zhou, Y.; Shen, J.; Wu, Z.; Liu, G.; Lee, P.W.; Tang, Y. AdmetSAR: A Comprehensive Source and Free Tool for Assessment of Chemical ADMET Properties. J. Chem. Inf. Model. 2012, 52, 3099–3105. [Google Scholar] [CrossRef]
- Guan, L.; Yang, H.; Cai, Y.; Sun, L.; Di, P.; Li, W.; Liu, G.; Tang, Y. ADMET-Score-a Comprehensive Scoring Function for Evaluation of Chemical Drug-Likeness. MedChemComm 2019, 10, 148–157. [Google Scholar] [CrossRef]
- Venkatraman, V. FP-ADMET: A Compendium of Fingerprint-Based ADMET Prediction Models. J. Cheminformatics 2021, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; Eckert, A.O.; Schrey, A.K.; Preissner, R. ProTox-II: A Webserver for the Prediction of Toxicity of Chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grover, A. Drug Design: Principles and Applications; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 978-981-10-5187-6. [Google Scholar]
- Fonseka, P.; Pathan, M.; Chitti, S.V.; Kang, T.; Mathivanan, S. FunRich Enables Enrichment Analysis of OMICs Datasets. J. Mol. Biol. 2021, 433, 166747. [Google Scholar] [CrossRef] [PubMed]
- Pathan, M.; Keerthikumar, S.; Ang, C.-S.; Gangoda, L.; Quek, C.Y.J.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A.; et al. FunRich: An Open Access Standalone Functional Enrichment and Interaction Network Analysis Tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 [µM] | ||||||
|---|---|---|---|---|---|---|
| Compounds | Incubation Time [h] | Normoxia | Hypoxia | |||
| 5637 | SCaBER | MRC-5 | 5637 | SCaBER | ||
| 4 | 24 | 3.31 ± 0.34 | 4.18 ± 1.30 | >10 | 2.58 ± 0.11 | 2.42 ± 0.23 |
| 48 | 3.27 ± 0.45 | 4.57 ± 2.07 | >10 | 2.96 ± 0.23 | 2.86 ± 0.62 | |
| 5 | 24 | 8.40 ± 0.21 | 8.53 ± 0.16 | >10 | 6.43 ± 0.26 | 7.27 ± 0.20 |
| 48 | 7.91 ± 0.46 | 7.97 ± 0.40 | >10 | 5.02 ± 1.06 | 7.58 ± 0.91 | |
| 6 | 24 | 2.60 ± 0.99 | 4.11 ± 1.40 | >10 | 2.37 ± 0.19 | 5.85 ± 1.98 |
| 48 | 2.63 ± 0.77 | 4.35 ± 1.97 | 9.34 ± 0.76 | 2.66 ± 0.07 | 7.28 ± 1.81 | |
| 10 | 24 | 2.25 ± 0.08 | 2.59 ± 0.16 | >10 | 1.96 ± 0.47 | 2.35 ± 0.22 |
| 48 | 2.17 ± 0.24 | 2.71 ± 0.34 | 7.84 ± 0.15 | 2.12 ± 0.33 | 2.60 ± 0.03 | |
| Curcumin (11) | 24 | 12.65 ± 3.03 | 13.14 ± 0.68 | 70.35 ± 12.45 | 12.14 ± 0.16 | 11.60 ± 0.83 |
| 48 | 12.86 ± 2.35 | 12.57 ± 0.62 | 45.33 ± 3.18 | 13.29 ± 0.54 | 11.33 ± 0.19 | |
| 12 | 24 | 2.48 ± 0.55 | 2.28 ± 0.13 | 9.63 ± 2.61 | 2.32 ± 0.29 | 3.26 ± 1.05 |
| 48 | 2.40 ± 0.45 | 2.21 ± 0.17 | 5.80 ± 0.77 | 2.64 ± 0.54 | 4.25 ± 1.94 | |
| Isocurcumin (13) | 24 | 10.85 ± 3.46 | 14.66 ± 0.88 | >20 | 13.09 ± 2.89 | 14.17 ± 0.23 |
| 48 | 6.71 ± 1.70 | 14.26 ± 0.41 | >20 | 10.12 ± 3.12 | 11.94 ± 0.09 | |
| Compound | ||||
|---|---|---|---|---|
| Parameter | 4 | 5 | 10 | Curcumin |
| TPSA a,* | 118.6 | 129.6 | 85.22 | 93.06 |
| logP b,* | 4.94 | 4.56 | 4.24 | 3.37 |
| Subcellular Localization ** | Mitochondria | Mitochondria | Mitochondria | Mitochondria |
| Caco-2 permeability * LogPapp cm/s ** | 1.0738 | 1.4124 | 0.8938 | 1.2769 |
| BBB c,** | + | + | + | - |
| Carcinogenicity ** | - | - | + | - |
| AOT d,** | III | IV | III | III |
| P-glycoprotein Substrate ** | Substrate | Substrate | Substrate | Substrate |
| P-glycoprotein Inhibitor ** | Inhibitor | Inhibitor | Non-inhibitor | Non-inhibitor |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazewski, D.; Kucinska, M.; Potapskiy, E.; Kuzminska, J.; Popenda, L.; Tezyk, A.; Goslinski, T.; Wierzchowski, M.; Murias, M. Enhanced Cytotoxic Activity of PEGylated Curcumin Derivatives: Synthesis, Structure–Activity Evaluation, and Biological Activity. Int. J. Mol. Sci. 2023, 24, 1467. https://doi.org/10.3390/ijms24021467
Lazewski D, Kucinska M, Potapskiy E, Kuzminska J, Popenda L, Tezyk A, Goslinski T, Wierzchowski M, Murias M. Enhanced Cytotoxic Activity of PEGylated Curcumin Derivatives: Synthesis, Structure–Activity Evaluation, and Biological Activity. International Journal of Molecular Sciences. 2023; 24(2):1467. https://doi.org/10.3390/ijms24021467
Chicago/Turabian StyleLazewski, Dawid, Malgorzata Kucinska, Edward Potapskiy, Joanna Kuzminska, Lukasz Popenda, Artur Tezyk, Tomasz Goslinski, Marcin Wierzchowski, and Marek Murias. 2023. "Enhanced Cytotoxic Activity of PEGylated Curcumin Derivatives: Synthesis, Structure–Activity Evaluation, and Biological Activity" International Journal of Molecular Sciences 24, no. 2: 1467. https://doi.org/10.3390/ijms24021467
APA StyleLazewski, D., Kucinska, M., Potapskiy, E., Kuzminska, J., Popenda, L., Tezyk, A., Goslinski, T., Wierzchowski, M., & Murias, M. (2023). Enhanced Cytotoxic Activity of PEGylated Curcumin Derivatives: Synthesis, Structure–Activity Evaluation, and Biological Activity. International Journal of Molecular Sciences, 24(2), 1467. https://doi.org/10.3390/ijms24021467