
Dose- and Segment-Dependent Disturbance of Rat Gut by Ionizing Radiation: Impact of Tight Junction Proteins

 and
and 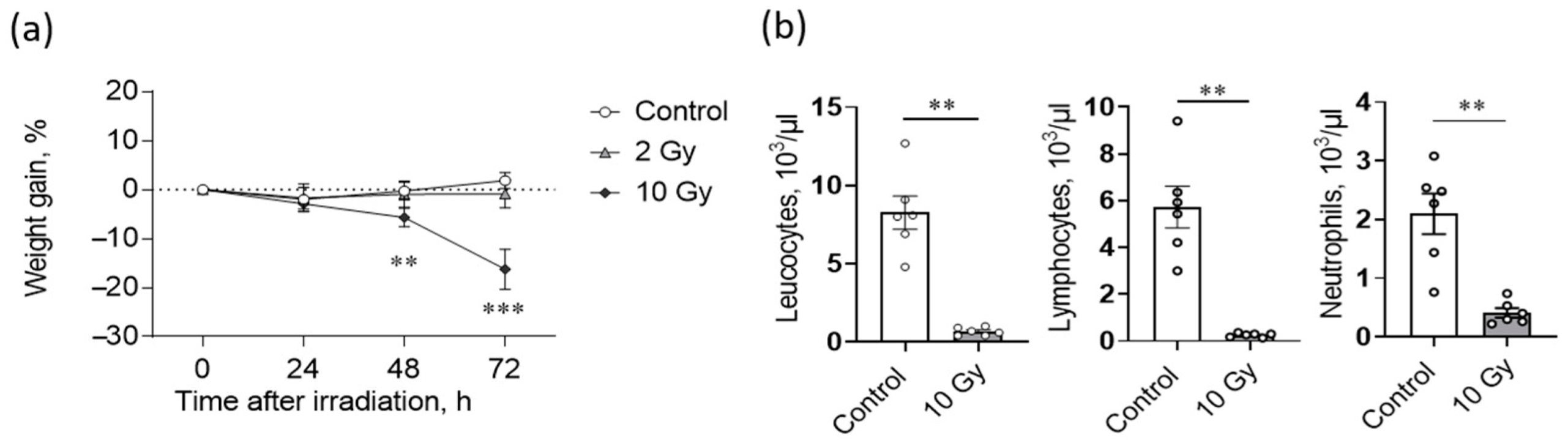{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. High Dose IR Exposure Affects Weight Gain and Leukocyte Counts in Rats
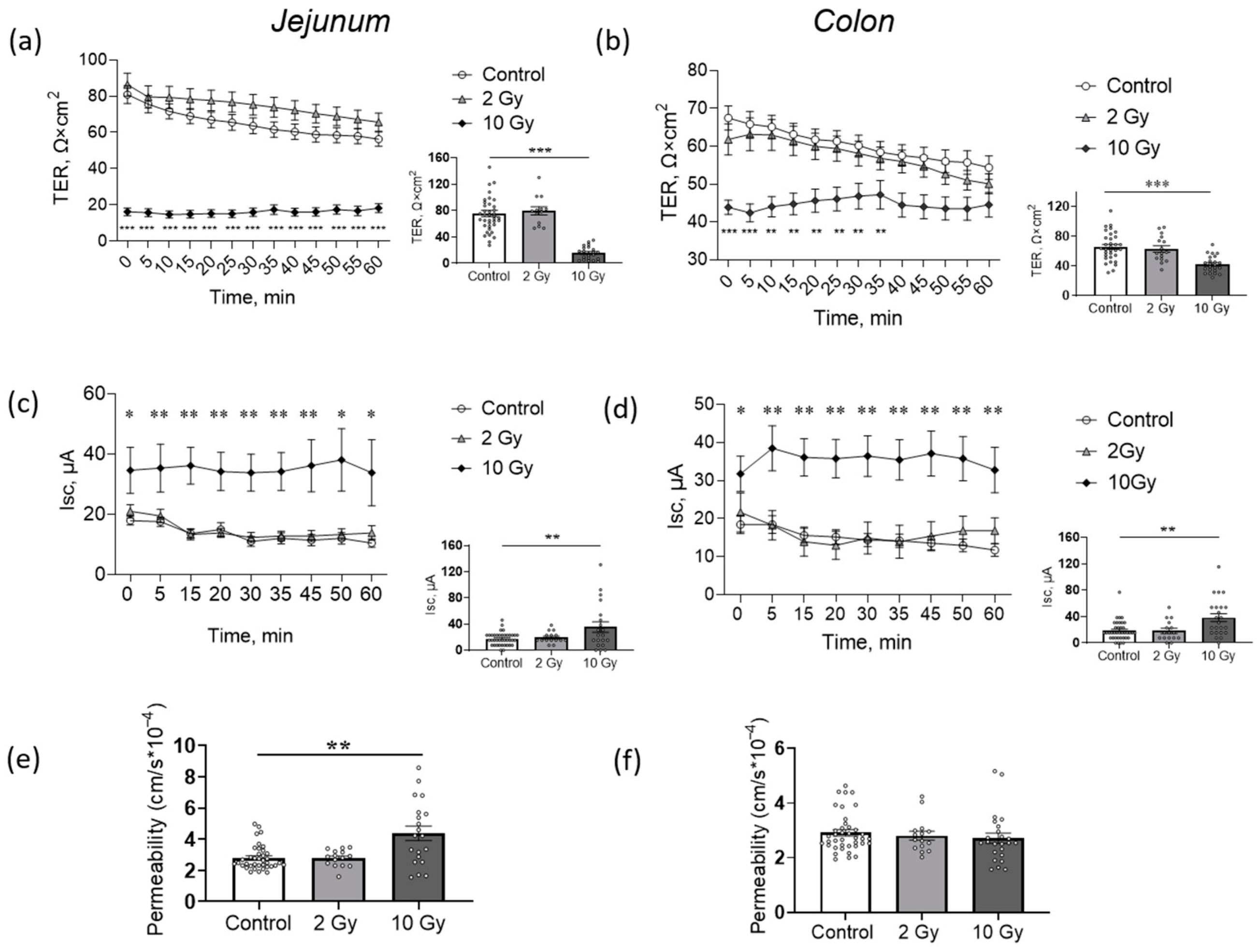
2.2. High-Dose IR Disturbs Transepithelial Resistence (TER) and Short-Circuit Current (Isc) of Jejunum and Colon
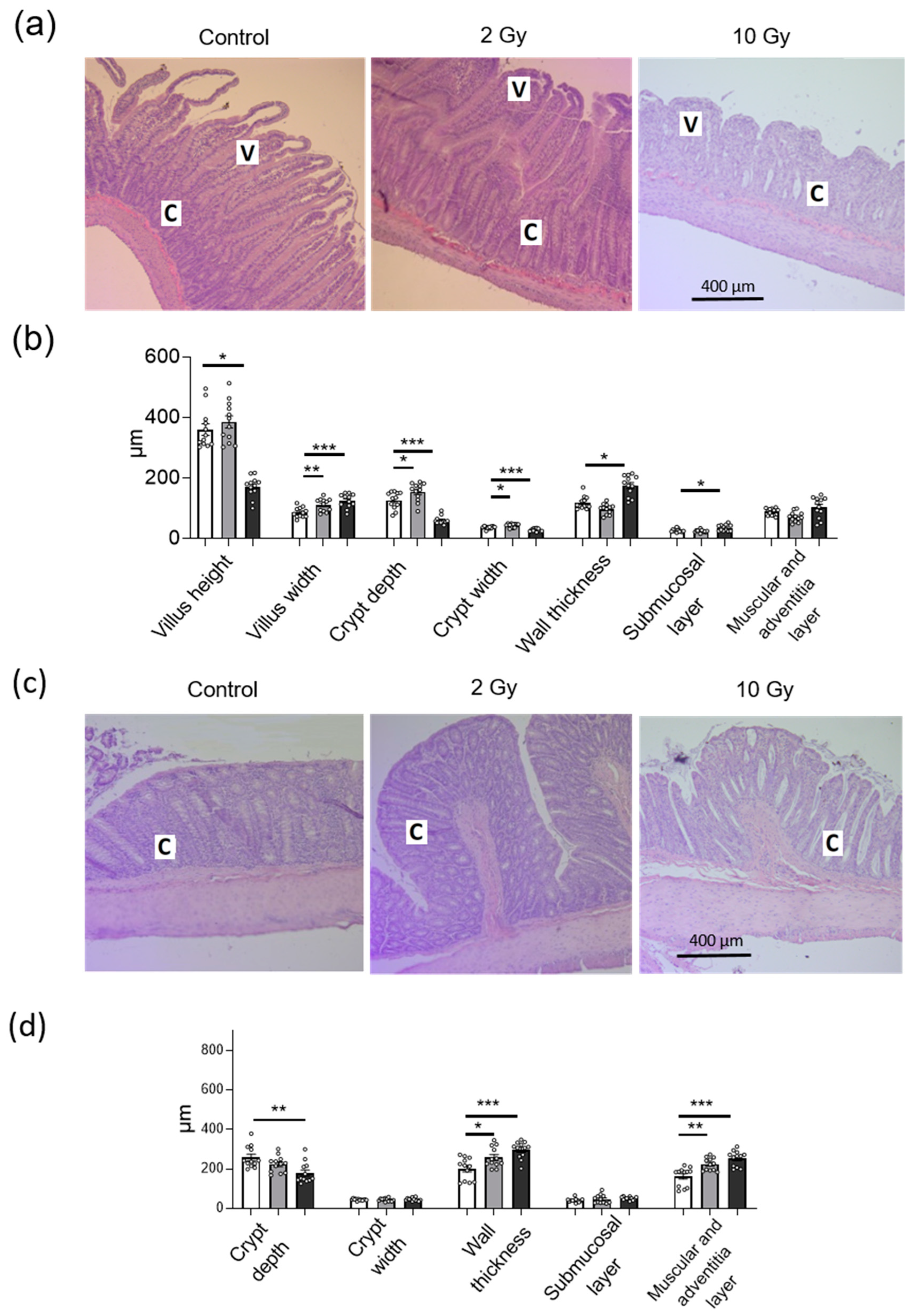
2.3. IR in Low and High Doses Disturbs Histological Structure of Jejunum and Colon Tissue
2.4. IR Disturbs Tight Junction (TJ) Proteins in Jejunum and Colon in Dose- and Segment-Specific Manners
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Hematological Analysis
4.3. Registration of Electrophysiological Parameters in the Ussing Chamber
4.4. Permeability for Sodium Fluorescein
4.5. Histological Analysis
4.6. Western Blot
4.7. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kiang, J.G.; Olabisi, A.O. Radiation: A poly-traumatic hit leading to multi-organ injury. Cell. Biosci. 2019, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Dubois, A.; Walker, R.I. Prospects for management of gastrointestinal injury associated with the acute radiation syndrome. Gastroenterology 1988, 95, 500–507. [Google Scholar] [CrossRef]
- Garau, M.M.; Calduch, A.L.; López, E.C. Radiobiology of the acute radiation syndrome. Rep. Pract. Oncol. Radiother. 2011, 16, 123–130. [Google Scholar] [CrossRef] [Green Version]
- MacVittie, T.J.; Bennett, A.; Booth, C.; Garofalo, M.; Tudor, G.; Ward, A.; Shea-Donohue, T.; Gelfond, D.; McFarland, E.; Jackson, W., 3rd; et al. The prolonged gastrointestinal syndrome in rhesus macaques: The relationship between gastrointestinal, hematopoietic, and delayed multi-organ sequelae following acute, potentially lethal, partial-body irradiation. Health Phys. 2012, 103, 427–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacon, C.G.; Giovanucci, E.; Testa, M.; Kawachi, I. The impact of cancer treatment on quality of life outcomes for patients with localized prostate cancer. J. Urol. 2001, 166, 1804–1810. [Google Scholar] [CrossRef] [PubMed]
- Sher, D.J. Cost-effectiveness studies in radiation therapy. Expert. Rev. Pharmacoecon. Outcomes 2010, 10, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.; Schwartz, A.; Sultan, S.; Schaefer, I.M.; Hermann, R.; Rave-Fränk, M.; Hess, C.F.; Christiansen, H.; Ramadori, G. Radiation-induced damage in different segments of the rat intestine after external beam irradiation of the liver. Exp. Mol. Pathol. 2012, 92, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Driák, D.; Osterreicher, J.; Vávrová, J.; Řeháková, Z.; Vilasová, Z. Morphological changes of rat jejunum after whole body gamma-irradiation and their impact in biodosimetry. Physiol. Res. 2008, 57, 475–479. [Google Scholar] [CrossRef]
- Marshman, E.; Ottewell, P.D.; Potten, C.S.; Watson, A.J. Caspase activation during spontaneous and radiation-induced apoptosis in the murine intestine. J. Pathol. 2001, 195, 285–292. [Google Scholar] [CrossRef]
- Mercantepe, F.; Topcu, A.; Rakici, S.; Tumkaya, L.; Yilmaz, A. The effects of N-acetylcysteine on radiotherapy-induced small intestinal damage in rats. Exp. Biol. Med. 2019, 244, 372–379. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Kruglova, N.; Razgovorova, I.; Amasheh, S.; Markov, A. Accumulation of milk increases the width of tight junctions in the epithelium of mouse mammary alveoli. Biol. Commun. 2020, 65, 277–280. [Google Scholar] [CrossRef]
- Markov, A.G.; Aschenbach, J.R.; Amasheh, S. Claudin clusters as determinants of epithelial barrier function. IUBMB Life 2015, 67, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Ikenouchi, J.; Furuse, M.; Furuse, K.; Sasaki, H.; Tsukita, S.; Tsukita, S. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J. Cell. Biol. 2005, 171, 939–945. [Google Scholar] [CrossRef]
- Günzel, D.; Fromm, M. Claudins and other tight junction proteins. Compr. Physiol. 2012, 2, 1819–1852. [Google Scholar] [CrossRef]
- Lu, Z.; Ding, L.; Lu, Q.; Chen, Y.H. Claudins in intestines: Distribution and functional significance in health and diseases. Tissue Barriers 2013, 1, e24978. [Google Scholar] [CrossRef] [Green Version]
- Markov, A.G.; Veshnyakova, A.; Fromm, M.; Amasheh, M.; Amasheh, S. Segmental expression of claudin proteins correlates with tight junction barrier properties in rat intestine. J. Comp. Physiol. B 2010, 180, 591–598. [Google Scholar] [CrossRef]
- Zhu, L.; Han, J.; Li, L.; Wang, Y.; Li, Y.; Zhang, S. Claudin family participates in the pathogenesis of inflammatory bowel diseases and colitis-associated colorectal cancer. Front. Immunol. 2019, 10, 1441. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.A.; Uppada, S.; Achkar, I.W.; Hashem, S.; Yadav, S.K.; Shanmugakonar, M.; Al-Naemi, H.A.; Haris, M.; Uddin, S. Tight Junction Proteins and Signaling Pathways in Cancer and Inflammation: A Functional Crosstalk. Front. Physiol. 2019, 9, 1942. [Google Scholar] [CrossRef] [Green Version]
- Shim, S.; Lee, J.G.; Bae, C.H.; Lee, S.B.; Jang, W.S.; Lee, S.J.; Lee, S.S.; Park, S. Claudin-3 expression in radiation-exposed rat models: A potential marker for radiation-induced intestinal barrier failure. Biochem. Biophys. Res. Commun. 2015, 456, 351–354. [Google Scholar] [CrossRef]
- Shukla, P.K.; Gangwar, R.; Manda, B.; Meena, A.S.; Yadav, N.; Szabo, E.; Balogh, A.; Lee, S.C.; Tigyi, G.; Rao, R. Rapid disruption of intestinal epithelial tight junction and barrier dysfunction by ionizing radiateon in mouse colon in vivo: Protection by N-acetyl-l-cysteine. Am. J. Physiol. Gastrointest. Liver. Physiol. 2016, 310, G705–G715. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Yu, J.; Jones, J.W.; Carter, C.L.; Pierzchalski, K.; Tudor, G.; Booth, C.; MacVittie, T.J.; Kane, M.A. Proteomic Evaluation of the Acute Radiation Syndrome of the Gastrointestinal Tract in a Murine Total-body Irradiation Model. Health Phys. 2019, 116, 516–528. [Google Scholar] [CrossRef]
- Howarth, G.S.; Fraser, R.; Frisby, C.L.; Schirmer, M.B.; Yeoh, E.K. Effects of insulin-like growth factor-I administration on radiation enteritis in rats. Scand. J. Gastroenterol. 1997, 32, 1118–1124. [Google Scholar] [CrossRef]
- Gu, J.; Chen, Y.Z.; Zhang, Z.X.; Yang, Z.X.; Duan, G.X.; Qin, L.Q.; Zhao, L.; Xu, J.Y. At What Dose Can Total Body and Whole Abdominal Irradiation Cause Lethal Intestinal Injury Among C57BL/6J Mice? Dose Response. 2020, 18, 1559325820956783. [Google Scholar] [CrossRef] [PubMed]
- International Atomic Energy Agency. Manual on Radiation Haematology; International Atomic Energy Agency: Vienna, Austria, 1971; pp. 1–450. [Google Scholar]
- Mihandoost, E.; Shirazi, A.; Mahdavi, S.R.; Aliasgharzadeh, A. Consequences of lethal-whole-body gamma radiation and possible ameliorative role of melatonin. Sci. World J. 2014, 2014, 621570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livanova, A.A.; Fedorova, A.A.; Zavirsky, A.V.; Bikmurzina, A.E.; Krivoi, I.I.; Markov, A.G. Dose and time dependence of functional impairments in rat jejunum following ionizing radiation exposure. Physiol. Rep. 2021, 9, e14960. [Google Scholar] [CrossRef]
- Molenda, N.; Urbanova, K.; Weiser, N.; Kusche-Vihrog, K.; Günzel, D.; Schillers, H. Paracellular transport through healthy and cystic fibrosis bronchial epithelial cell lines--do we have a proper model? PLoS ONE 2014, 9, e100621. [Google Scholar] [CrossRef] [Green Version]
- Markov, A.G.; Fedorova, A.A.; Kravtsova, V.V.; Bikmurzina, A.E.; Okorokova, L.S.; Matchkov, V.V.; Cornelius, V.; Amasheh, S.; Krivoi, I.I. Circulating Ouabain Modulates Expression of Claudins in Rat Intestine and Cerebral Blood Vessels. Int. J. Mol. Sci. 2020, 21, 5067. [Google Scholar] [CrossRef]
- Freeman, S.L.; Hossain, M.; MacNaughton, W.K. Radiation-induced acute intestinal inflammation differs following total-body versus abdominopelvic irradiation in the ferret. Int. J. Radiat. Biol. 2001, 77, 389–395. [Google Scholar] [CrossRef]
- Potten, C.S.; Grant, H.K. The relationship between ionizing radiation induced apoptosis and stem cells in the small and large intestine. Br. J. Cancer 1998, 78, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Kiang, J.G.; Smith, J.T.; Anderson, M.N.; Elliott, T.B.; Gupta, P.; Balakathiresan, N.S.; Maheshwari, R.K.; Knollmann-Ritschel, B. Hemorrhage enhancescytokine, complement component 3, and caspase-3, and regulates microRNAs associated with intestinal damage after whole-body gamma-irradiation in combined injury. PLoS ONE 2017, 12, e0184393. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, L.W.; Al-Sadi, R.; Ma, T.Y. IL-1β and the Intestinal Epithelial Tight Junction Barrier. Front. Immunol. 2021, 12, 767456. [Google Scholar] [CrossRef]
- Romero, E.S.; Cotoner, C.A.; Camacho, C.P.; Bedmar, M.C.; Vicario, M. The Intestinal Barrier Function and its Involvement in Digestive Disease. Rev. Esp. Enferm. Dig. 2015, 107, 686–696. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Guo, C.; Zhang, D.; Zhang, J.; Wang, X.; Geng, C. The altered tight junctions: An important gateway of bacterial translocation in cachexia patients with advanced gastric cancer. J. Interferon Cytokine Res. 2014, 34, 518–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, F.; Loucas, B.D.; Ito, I.; Asai, A.; Suzuki, S.; Kobayashi, M. Survival of Mice with Gastrointestinal Acute Radiation Syndrome through Control of Bacterial Translocation. J. Immunol. 2018, 201, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Challapalli, S.; Kumar, A.; Rai, R.; Kini, J.; Kumarchandra, R. Standardization of mean lethal dose (LD 50/30) of X-rays using linear accelerator (LINAC) in Albino Wistar rat model based on survival analysis studies and hematological parameters. Res. J. Pharm. Biol. Chem. Sci. 2015, 5, 1215–1219. [Google Scholar]
- Markov, A.G.; Falchuk, E.L.; Kruglova, N.M.; Radloff, J.; Amasheh, S. Claudin expression in follicle-associated epithelium of rat Peyer’s patches defines a major restriction of the paracellular pathway. Acta Physiol. 2016, 216, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Overgaard, C.E.; Daugherty, B.L.; Mitchell, L.A.; Koval, M. Claudins: Control of barrier function and regulation in response to oxidant stress. Antioxid. Redox. Signal 2011, 15, 1179–1193. [Google Scholar] [CrossRef] [Green Version]
- Weber, C.R.; Nalle, S.C.; Tretiakova, M.; Rubin, D.T.; Turner, J.R. Claudin-1 and claudin-2 expression is elevated in inflammatory bowel disease and may contribute to early neoplastic transformation. Lab. Investig. 2008, 88, 1110–1120. [Google Scholar] [CrossRef] [Green Version]
- Oshima, T.; Miwa, H.; Joh, T. Changes in the expression of claudins in active ulcerative colitis. J. Gastroenterol. Hepatol. 2008, 23, S146–S150. [Google Scholar] [CrossRef]
- Garcia-Hernandez, V.; Quiros, M.; Nusrat, A. Intestinal epithelial claudins: Expression and regulation in homeostasis and inflammation. Ann. N. Y. Acad. Sci. 2017, 1397, 66–79. [Google Scholar] [CrossRef]
- Saeedi, B.J.; Kao, D.J.; Kitzenberg, D.A.; Dobrinskikh, E.; Schwisow, K.D.; Masterson, J.C.; Kendrick, A.A.; Kelly, C.J.; Bayless, A.J.; Kominsky, D.J.; et al. HIF-dependent regulation of claudin-1 is central to intestinal epithelial tight junction integrity. Mol. Biol. Cell 2015, 26, 2252–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Chaturvedi, R.; Olivares-Villagómez, D.; Habib, T.; Asim, M.; Shivesh, P.; Polk, D.B.; Wilson, K.T.; Washington, M.K.; Van Kaer, L.; et al. Targeted colonic claudin-2 expression renders resistance to epithelial injury, induces immune suppression, and protects from colitis. Mucosal Immunol. 2014, 7, 1340–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, K.; Katahira, J.; Horiguchi, Y.; Sonoda, N.; Furuse, M.; Tsukita, S. Clostridium perfringens enterotoxin binds to the second extracellular loop of claudin-3, a tight junction integral membrane protein. FEBS Lett. 2000, 476, 258–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Yin, W.; Ma, H.; Elshoura, I.; Wang, L. Targeting claudin-3 suppresses stem cell-like phenotype in nonsquamous non-small-cell lung carcinoma. Lung Cancer Manag. 2019, 8, LMT04. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredriksson, K.; Van Itallie, C.M.; Aponte, A.; Gucek, M.; Tietgens, A.J.; Anderson, J.M. Proteomic analysis of proteins surrounding occludin and claudin-4 reveals their proximity to signaling and trafficking networks. PLoS ONE 2015, 10, e0117074. [Google Scholar] [CrossRef]
- Michl, P.; Barth, C.; Buchholz, M.; Lerch, M.M.; Rolke, M.; Holzmann, K.H.; Menke, A.; Fensterer, H.; Giehl, K.; Löhr, M.; et al. Claudin-4 expression decreases invasiveness and metastatic potential of pancreatic cancer. Cancer Res. 2003, 63, 6265–6271. [Google Scholar]
- Hicks, D.A.; Galimanis, C.E.; Webb, P.G.; Spillman, M.A.; Behbakht, K.; Neville, M.C.; Baumgartner, H.K. Claudin-4 activity in ovarian tumor cell apoptosis resistance and migration. BMC Cancer 2016, 16, 788. [Google Scholar] [CrossRef] [Green Version]
- Yahyapour, R.; Motevaseli, E.; Rezaeyan, A.; Abdollahi, H.; Farhood, B.; Cheki, M.; Rezapoor, S.; Shabeeb, D.; Musa, A.E.; Najafi, M.; et al. Reduction-oxidation (redox) system in radiation-induced normal tissue injury: Molecular mechanisms and implications in radiation therapeutics. Clin. Transl. Oncol. 2018, 20, 975–988. [Google Scholar] [CrossRef] [PubMed]
- Elias, B.C.; Suzuki, T.; Seth, A.; Giorgianni, F.; Kale, G.; Shen, L.; Turner, J.R.; Naren, A.; Desiderio, D.M.; Rao, R. Phosphorylation of Tyr-398 and Tyr-402 in occludin prevents its interaction with ZO-1 and destabilizes its assembly at the tight junctions. J. Biol. Chem. 2009, 284, 1559–1569. [Google Scholar] [CrossRef] [Green Version]
- Kale, G.; Naren, A.P.; Sheth, P.; Rao, R.K. Tyrosine phosphorylation of occludin attenuates its interactions with ZO-1, ZO-2, and ZO-3. Biochem. Biophys. Res. Commun. 2003, 302, 324–329. [Google Scholar] [CrossRef]
- Baradaran-Ghahfarokhi, M.; Amouheidari, A.; Shahbazi-Gahrouei, D.; Baradaran-Ghahfarokhi, H.R.; Tanderup, K.; Dörr, W.; Shokrani, P. Evaluation of the Effects of Prostate Radiation Therapy on Occludin Expression and Ultrasonography Characteristics of the Bladder. Int. J. Radiat. Oncol. Biol. Phys. 2017, 99, 963–971. [Google Scholar] [CrossRef]
- Osanai, M.; Murata, M.; Nishikiori, N.; Chiba, H.; Kojima, T.; Sawada, N. Epigenetic silencing of occludin promotes tumorigenic and metastatic properties of cancer cells via modulations of unique sets of apoptosis-associated genes. Cancer Res. 2006, 66, 9125–9133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, J. Paracellular Water Channel. In The Paracellular Channel: Biology, Physiology, and Disease; Academic Press: Cambridge, MA, USA, 2019; pp. 83–92. [Google Scholar]
- Krug, S.M.; Amasheh, S.; Richter, J.F.; Milatz, S.; Günzel, D.; Westphal, J.K.; Huber, O.; Schulzke, J.D.; Fromm, M. Tricellulin forms a barrier to macromolecules in tricellular tight junctions without affecting ion permeability. Mol. Biol. Cell. 2009, 20, 3713–3724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kravtsova, V.V.; Fedorova, A.A.; Tishkova, M.V.; Livanova, A.A.; Vetrovoy, O.V.; Markov, A.G.; Matchkov, V.V.; Krivoi, I.I. Chronic Ouabain Prevents Radiation-Induced Reduction in the α2 Na, K-ATPase Function in the Rat Diaphragm Muscle. Int. J. Mol. Sci. 2022, 23, 10921. [Google Scholar] [CrossRef] [PubMed]
- Larre, I.; Lazaro, A.; Contreras, R.G.; Balda, M.S.; Matter, K.; Flores-Maldonado, C.; Ponce, A.; Flores-Benitez, D.; Rincon-Heredia, R.; Padilla-Benavides, T.; et al. Ouabain modulates epithelial cell tight junction. Proc. Natl. Acad. Sci. USA 2010, 107, 11387–11392. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011; pp. 1–246. [Google Scholar]
- Harvey, J.W.; Stevens, A.; Lowe, J.S.; Scott, I. Veterinary Hematology; WB Saunders: St. Louis, MO, USA, 2012; pp. 234–259. [Google Scholar]
- Thomson, A.; Smart, K.; Somerville, M.S.; Lauder, S.N.; Appanna, G.; Horwood, J.; Sunder Raj, L.; Srivastava, B.; Durai, D.; Scurr, M.J.; et al. The Ussing chamber system for measuring intestinal permeability in health and disease. BMC Gastroenterol. 2019, 19, 98. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Livanova, A.A.; Fedorova, A.A.; Zavirsky, A.V.; Krivoi, I.I.; Markov, A.G. Dose- and Segment-Dependent Disturbance of Rat Gut by Ionizing Radiation: Impact of Tight Junction Proteins. Int. J. Mol. Sci. 2023, 24, 1753. https://doi.org/10.3390/ijms24021753
Livanova AA, Fedorova AA, Zavirsky AV, Krivoi II, Markov AG. Dose- and Segment-Dependent Disturbance of Rat Gut by Ionizing Radiation: Impact of Tight Junction Proteins. International Journal of Molecular Sciences. 2023; 24(2):1753. https://doi.org/10.3390/ijms24021753
Chicago/Turabian StyleLivanova, Alexandra A., Arina A. Fedorova, Alexander V. Zavirsky, Igor I. Krivoi, and Alexander G. Markov. 2023. "Dose- and Segment-Dependent Disturbance of Rat Gut by Ionizing Radiation: Impact of Tight Junction Proteins" International Journal of Molecular Sciences 24, no. 2: 1753. https://doi.org/10.3390/ijms24021753
APA StyleLivanova, A. A., Fedorova, A. A., Zavirsky, A. V., Krivoi, I. I., & Markov, A. G. (2023). Dose- and Segment-Dependent Disturbance of Rat Gut by Ionizing Radiation: Impact of Tight Junction Proteins. International Journal of Molecular Sciences, 24(2), 1753. https://doi.org/10.3390/ijms24021753