
Effects of Exercise and Omega-3-Supplemented, High-Protein Diet on Inflammatory Markers in Serum, on Gene Expression Levels in PBMC, and after Ex Vivo Whole-Blood LPS Stimulation in Old Adults

 , ,
, , .png) and
and
Abstract
:1. Introduction
2. Results
2.1. Baseline Characteristics
2.2. Adherence to Interventions and Changes in Body Composition
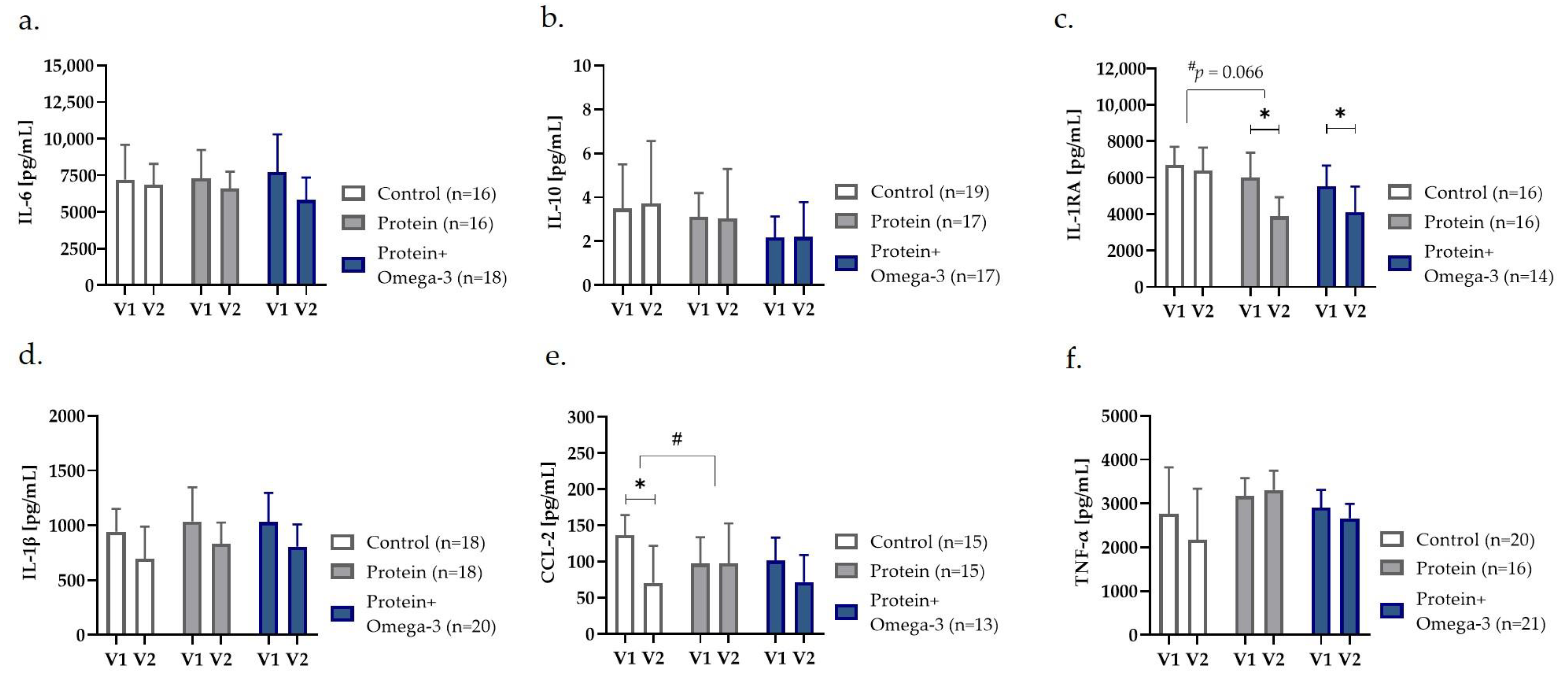
2.3. Omega-3 Supplementation Decreased Circulating Inflammatory Markers
2.4. Gene Expression Levels of Inflammatory Markers in PBMC Are Reduced with Both Protein-Enriched Dietary Interventions
2.5. Reduction in Ex Vivo LPS-Stimulated Cytokine/Chemokine Release after Exercise and Dietary Interventions
3. Discussion
3.1. Isolated Effects of Exercise Intervention on LPS-Induced Chemokine Release
3.2. Decreased Inflammatory Markers in Serum after High-Protein, Omega-3 Enriched Diet
3.3. Gene Expression Levels of Inflammatory Markers Reduced with Protein-Enriched Diets
3.4. Attenuated or no Change in LPS-Induced Immune Response with Dietary Interventions
3.5. Sex-Related Differences
3.6. Strength and Limitations
4. Materials and Methods
4.1. Study Design and Population Sample
4.2. Anthropometric Measurements
4.3. Dietary Assessment
4.4. Laboratory Assessments
4.5. PBMC Isolation, RNA Extraction, and Gene Expression with qPCR
4.6. Ex Vivo Whole-Blood LPS Stimulation
4.7. Whole-Body Vibration Training and Home-Based Resistance Exercise
4.8. Dietary Intervention
4.9. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 95% CI | 95% confidence interval |
| β/B | beta |
| B2M | beta-2-microglobulin |
| BMI | body mass index |
| CCL-2/CCL2 | c-c motif chemokine ligand-2 |
| DHA | docosahexaenoic acid |
| ELISA | enzyme-linked immunosorbent assays |
| EPA | eicosapentaenoic acid |
| FMI | fat mass index |
| HMGB-1 | high-mobility group box-1 |
| IL | interleukin |
| IQR | interquartile range |
| LPS | lipopolysaccharide |
| PBMC | peripheral blood mononuclear cells |
| qPCR | quantitative real-time polymerase chain reaction |
| RA/RN | receptor antagonist |
| RNA | ribonucleic acid |
| TNF-α/TNFA | tumor necrosis factor-α |
References
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A new immune-metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 2018, 14, 576–590. [Google Scholar] [CrossRef]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and Oxidative Stress in Human Diseases: From Molecular Mechanisms to Novel Treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef] [Green Version]
- Boirie, Y. Physiopathological mechanism of sarcopenia. J. Nutr. Health Aging 2009, 13, 717–723. [Google Scholar] [CrossRef]
- Teissier, T.; Boulanger, E.; Cox, L.S. Interconnections between Inflammageing and Immunosenescence during Ageing. Cells 2022, 11, 359. [Google Scholar] [CrossRef]
- Flynn, M.G.; Markofski, M.M.; Carrillo, A.E. Elevated Inflammatory Status and Increased Risk of Chronic Disease in Chronological Aging: Inflamm-aging or Inflamm-inactivity? Aging Dis. 2019, 10, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Monteiro-Junior, R.S.; de Tarso Maciel-Pinheiro, P.; da Matta Mello Portugal, E.; da Silva Figueiredo, L.F.; Terra, R.; Carneiro, L.S.F.; Rodrigues, V.D.; Nascimento, O.J.M.; Deslandes, A.C.; Laks, J. Effect of Exercise on Inflammatory Profile of Older Persons: Systematic Review and Meta-Analyses. J. Phys. Act. Health 2018, 15, 64–71. [Google Scholar] [CrossRef]
- Zhao, H.; He, Z.; Yun, H.; Wang, R.; Liu, C. A Meta-Analysis of the Effects of Different Exercise Modes on Inflammatory Response in the Elderly. Int. J. Environ. Res. Public Health 2022, 19, 451. [Google Scholar] [CrossRef]
- Haß, U.; Herpich, C.; Kochlik, B.; Weber, D.; Grune, T.; Norman, K. Dietary Inflammatory Index and Cross-Sectional Associations with Inflammation, Muscle Mass and Function in Healthy Old Adults. J. Nutr. Health Aging 2022, 26, 346–351. [Google Scholar] [CrossRef]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Kavyani, Z.; Musazadeh, V.; Fathi, S.; Hossein Faghfouri, A.; Dehghan, P.; Sarmadi, B. Efficacy of the omega-3 fatty acids supplementation on inflammatory biomarkers: An umbrella meta-analysis. Int. Immunopharmacol. 2022, 111, 109104. [Google Scholar] [CrossRef]
- Rangel-Huerta, O.D.; Aguilera, C.M.; Mesa, M.D.; Gil, A. Omega-3 long-chain polyunsaturated fatty acids supplementation on inflammatory biomakers: A systematic review of randomised clinical trials. Br. J. Nutr. 2012, 107 (Suppl. S2), S159–S170. [Google Scholar] [CrossRef] [Green Version]
- Draganidis, D.; Karagounis, L.G.; Athanailidis, I.; Chatzinikolaou, A.; Jamurtas, A.Z.; Fatouros, I.G. Inflammaging and Skeletal Muscle: Can Protein Intake Make a Difference? J. Nutr. 2016, 146, 1940–1952. [Google Scholar] [CrossRef] [Green Version]
- Markova, M.; Koelman, L.; Hornemann, S.; Pivovarova, O.; Sucher, S.; Machann, J.; Rudovich, N.; Thomann, R.; Schneeweiss, R.; Rohn, S.; et al. Effects of plant and animal high protein diets on immune-inflammatory biomarkers: A 6-week intervention trial. Clin. Nutr. 2020, 39, 862–869. [Google Scholar] [CrossRef]
- Hashemilar, M.; Khalili, M.; Rezaeimanesh, N.; Sadeghi Hokmabadi, E.; Rasulzade, S.; Shamshirgaran, S.M.; Taheraghdam, A.; Farhoudi, M.; Shaafi, S.; Shakouri, S.K.; et al. Effect of Whey Protein Supplementation on Inflammatory and Antioxidant Markers, and Clinical Prognosis in Acute Ischemic Stroke (TNS Trial): A Randomized, Double Blind, Controlled, Clinical Trial. Adv. Pharm. Bull. 2020, 10, 135–140. [Google Scholar] [CrossRef]
- Liberman, K.; Njemini, R.; Luiking, Y.; Forti, L.N.; Verlaan, S.; Bauer, J.M.; Memelink, R.; Brandt, K.; Donini, L.M.; Maggio, M.; et al. Thirteen weeks of supplementation of vitamin D and leucine-enriched whey protein nutritional supplement attenuates chronic low-grade inflammation in sarcopenic older adults: The PROVIDE study. Aging Clin. Exp. Res. 2019, 31, 845–854. [Google Scholar] [CrossRef] [Green Version]
- Haß, U.; Kochlik, B.; Herpich, C.; Rudloff, S.; Norman, K. Effects of an Omega-3 Supplemented, High-Protein Diet in Combination with Vibration and Resistance Exercise on Muscle Power and Inflammation in Old Adults: A Pilot Randomized Controlled Trial. Nutrients 2022, 14, 4274. [Google Scholar] [CrossRef]
- Varghese, M.; Song, J.; Singer, K. Age and Sex: Impact on adipose tissue metabolism and inflammation. Mech. Ageing Dev. 2021, 199, 111563. [Google Scholar] [CrossRef]
- Cristi, C.; Collado, P.S.; Márquez, S.; Garatachea, N.; Cuevas, M.J. Whole-body vibration training increases physical fitness measures without alteration of inflammatory markers in older adults. Eur. J. Sport Sci. 2014, 14, 611–619. [Google Scholar] [CrossRef]
- de Lourdes Nahhas Rodacki, C.; Rodacki, A.L.; Coelho, I.; Pequito, D.; Krause, M.; Bonatto, S.; Naliwaiko, K.; Fernandes, L.C. Influence of fish oil supplementation and strength training on some functional aspects of immune cells in healthy elderly women. Br. J. Nutr. 2015, 114, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Chaabene, H.; Prieske, O.; Herz, M.; Moran, J.; Höhne, J.; Kliegl, R.; Ramirez-Campillo, R.; Behm, D.G.; Hortobágyi, T.; Granacher, U. Home-based exercise programmes improve physical fitness of healthy older adults: A PRISMA-compliant systematic review and meta-analysis with relevance for COVID-19. Ageing Res. Rev. 2021, 67, 101265. [Google Scholar] [CrossRef]
- Stupin, M.; Kibel, A.; Stupin, A.; Selthofer-Relatić, K.; Matić, A.; Mihalj, M.; Mihaljević, Z.; Jukić, I.; Drenjančević, I. The Physiological Effect of n-3 Polyunsaturated Fatty Acids (n-3 PUFAs) Intake and Exercise on Hemorheology, Microvascular Function, and Physical Performance in Health and Cardiovascular Diseases; Is There an Interaction of Exercise and Dietary n-3 PUFA Intake? Front. Physiol. 2019, 10, 1129. [Google Scholar] [CrossRef] [Green Version]
- Markova, M.; Pivovarova, O.; Hornemann, S.; Sucher, S.; Frahnow, T.; Wegner, K.; Machann, J.; Petzke, K.J.; Hierholzer, J.; Lichtinghagen, R.; et al. Isocaloric Diets High in Animal or Plant Protein Reduce Liver Fat and Inflammation in Individuals With Type 2 Diabetes. Gastroenterology 2017, 152, 571–585.e8. [Google Scholar] [CrossRef] [Green Version]
- Nieman, K.M.; Anderson, B.D.; Cifelli, C.J. The Effects of Dairy Product and Dairy Protein Intake on Inflammation: A Systematic Review of the Literature. J. Am. Coll. Nutr. 2021, 40, 571–582. [Google Scholar] [CrossRef]
- Bruunsgaard, H.; Pedersen, A.N.; Schroll, M.; Skinhoj, P.; Pedersen, B.K. Impaired production of proinflammatory cytokines in response to lipopolysaccharide (LPS) stimulation in elderly humans. Clin. Exp. Immunol. 1999, 118, 235–241. [Google Scholar] [CrossRef]
- Vedin, I.; Cederholm, T.; Freund Levi, Y.; Basun, H.; Garlind, A.; Faxén Irving, G.; Jönhagen, M.E.; Vessby, B.; Wahlund, L.O.; Palmblad, J. Effects of docosahexaenoic acid-rich n-3 fatty acid supplementation on cytokine release from blood mononuclear leukocytes: The OmegAD study. Am. J. Clin. Nutr. 2008, 87, 1616–1622. [Google Scholar] [CrossRef] [Green Version]
- Thies, F.; Miles, E.A.; Nebe-von-Caron, G.; Powell, J.R.; Hurst, T.L.; Newsholme, E.A.; Calder, P.C. Influence of dietary supplementation with long-chain n-3 or n-6 polyunsaturated fatty acids on blood inflammatory cell populations and functions and on plasma soluble adhesion molecules in healthy adults. Lipids 2001, 36, 1183–1193. [Google Scholar] [CrossRef]
- Ellulu, M.S.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef]
- Jannasch, F.; Bedu-Addo, G.; Schulze, M.B.; Mockenhaupt, F.P.; Danquah, I. Serum phospholipid fatty acids, dietary patterns and type 2 diabetes among urban Ghanaians. Nutr. J. 2017, 16, 63. [Google Scholar] [CrossRef] [Green Version]
- Kessler, K.; Hornemann, S.; Petzke, K.J.; Kemper, M.; Markova, M.; Rudovich, N.; Grune, T.; Kramer, A.; Pfeiffer, A.F.H.; Pivovarova-Ramich, O. Diurnal distribution of carbohydrates and fat affects substrate oxidation and adipokine secretion in humans. Am. J. Clin. Nutr. 2018, 108, 1209–1219. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Control (n = 20) | Protein (n = 20) | Protein + Omega-3 (n = 21) | |
|---|---|---|---|
| Sex [female/male] | 10/10 | 11/9 | 11/10 |
| Age [years] | 69.9 ± 4.5 | 71.5 ± 4.6 | 70.4 ± 5.1 |
| Medication [n] | 3 ± 2 | 2 ± 2 | 2 ± 2 |
| Waist/height ratio | 0.58 ± 0.06 | 0.60 ± 0.05 | 0.59 ± 0.06 |
| Body mass index [kg/m2] | 26.9 ± 2.7 | 28.2 ± 2.3 | 27.8 ± 2.7 |
| Fat mass index [kg/m2] | 8.75 ± 2.38 | 9.44 ± 2.28 | 9.37 ± 1.98 |
| Serum concentrations | |||
| IL-6 (pg/mL) | 2.97 (1.34) | 2.62 (1.65) | 3.01 (2.10) |
| IL-10 (pg/mL) | 8.21 (4.28) | 7.84 (3.55) | 9.43 (4.21) |
| IL-6/IL-10 ratio | 0.37 (0.38) | 0.34 (0.25) | 0.34 (0.23) |
| IL-1RA (pg/mL) | 923 ± 366 | 790 ± 432 | 1228 ± 661 a |
| CCL-2 (pg/mL) | 214 (119) | 229 (118) | 251 (62) |
| HMGB-1 (ng/mL) | 0.38 (1.29) | 0.29 (0.50) | 0.25 (0.64) |
| Gene expression levels in PBMC | |||
| IL6 | 0.03 (0.04) | 0.05 (0.07) | 0.05 (0.05) |
| IL10 | 0.19 (0.20) | 0.17 (0.28) | 0.16 (0.21) |
| IL1RN | 0.05 (0.03) | 0.05 (0.01) | 0.05 (0.01) |
| IL1B | 0.04 (0.03) | 0.04 (0.02) | 0.04 (0.01) |
| CCL2 | 0.004 (0.007) | 0.005 (0.005) | 0.005 (0.003) |
| TNFA | 0.56 (0.28) | 0.66 (0.24) | 0.57 (0.19) |
| LPS-induced concentrations in whole-blood cultures | |||
| IL-6 (pg/mL) | 7618 (3598) | 6407 (5659) | 7059 (5004) |
| IL-10 (pg/mL) | 2.48 (7.12) | 3.41 (4.32) | 2.66 (2.82) |
| IL-1RA (pg/mL) | 6194 (2640) | 5116 (4605) | 4913 (4310) |
| IL-1β (pg/mL) | 850 (587) | 954 (660) | 879 (440) |
| CCL-2 (pg/mL) | 122 (100) | 109 (131) | 101 (82) |
| TNF-α (pg/mL) | 3245 ± 1488 | 2787 ± 1394 | 3117 ± 1151 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haß, U.; Heider, S.; Kochlik, B.; Herpich, C.; Pivovarova-Ramich, O.; Norman, K. Effects of Exercise and Omega-3-Supplemented, High-Protein Diet on Inflammatory Markers in Serum, on Gene Expression Levels in PBMC, and after Ex Vivo Whole-Blood LPS Stimulation in Old Adults. Int. J. Mol. Sci. 2023, 24, 928. https://doi.org/10.3390/ijms24020928
Haß U, Heider S, Kochlik B, Herpich C, Pivovarova-Ramich O, Norman K. Effects of Exercise and Omega-3-Supplemented, High-Protein Diet on Inflammatory Markers in Serum, on Gene Expression Levels in PBMC, and after Ex Vivo Whole-Blood LPS Stimulation in Old Adults. International Journal of Molecular Sciences. 2023; 24(2):928. https://doi.org/10.3390/ijms24020928
Chicago/Turabian StyleHaß, Ulrike, Sarah Heider, Bastian Kochlik, Catrin Herpich, Olga Pivovarova-Ramich, and Kristina Norman. 2023. "Effects of Exercise and Omega-3-Supplemented, High-Protein Diet on Inflammatory Markers in Serum, on Gene Expression Levels in PBMC, and after Ex Vivo Whole-Blood LPS Stimulation in Old Adults" International Journal of Molecular Sciences 24, no. 2: 928. https://doi.org/10.3390/ijms24020928
APA StyleHaß, U., Heider, S., Kochlik, B., Herpich, C., Pivovarova-Ramich, O., & Norman, K. (2023). Effects of Exercise and Omega-3-Supplemented, High-Protein Diet on Inflammatory Markers in Serum, on Gene Expression Levels in PBMC, and after Ex Vivo Whole-Blood LPS Stimulation in Old Adults. International Journal of Molecular Sciences, 24(2), 928. https://doi.org/10.3390/ijms24020928