

Vitamin D, Gut Microbiota, and Cardiometabolic Diseases—A Possible Three-Way Axis

Abstract
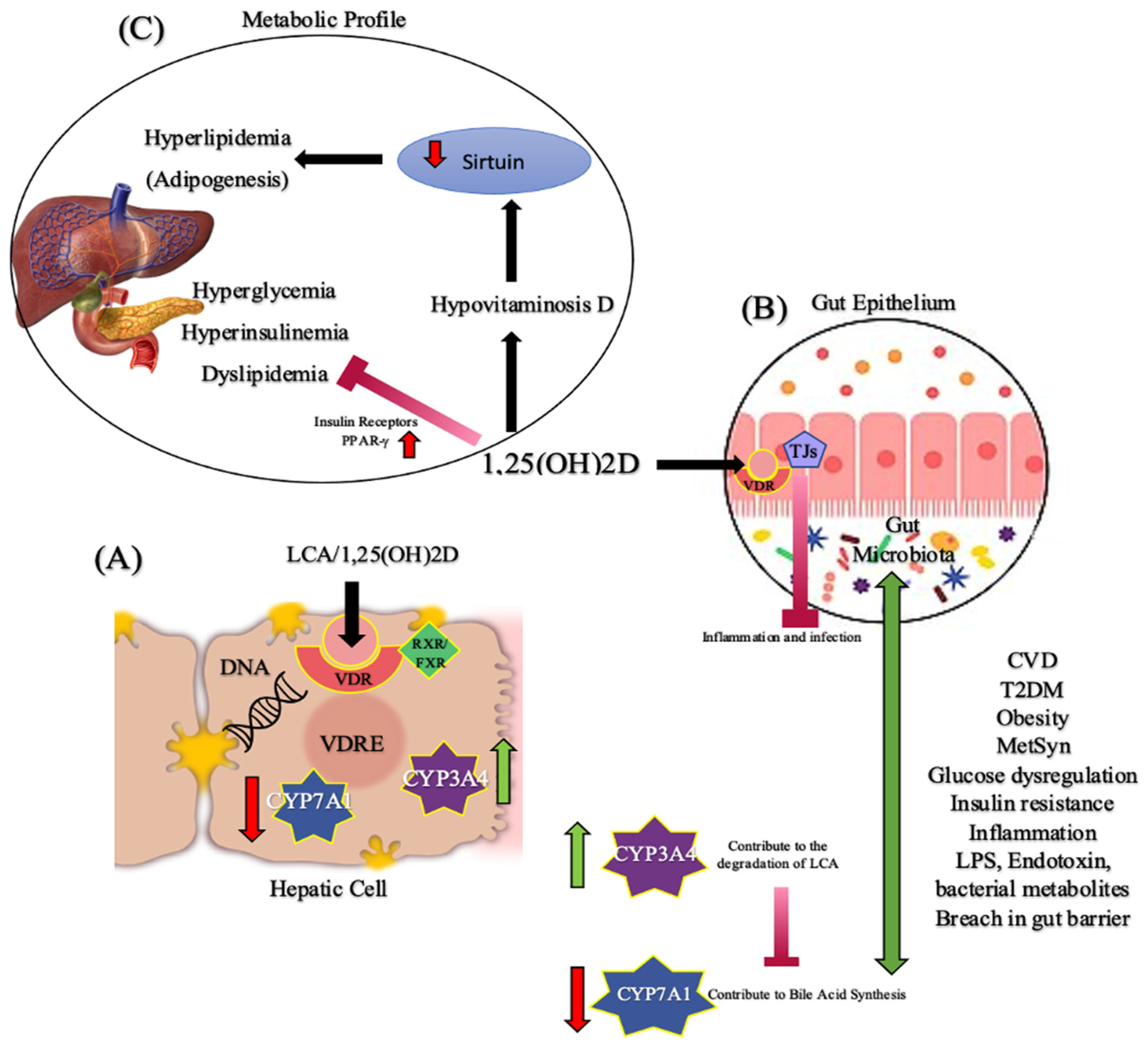
:1. Introduction
2. Literature Search Strategy
3. Vitamin D and Cardiometabolic Diseases: Proposed Mechanism of Action
3.1. Vitamin D and Cardiometabolic Diseases: Evidence from Interventional Studies
3.2. Vitamin D and Cardiometabolic Diseases: Evidence from Epidemiological Studies, Systematic Reviews, and Meta-Analyses
4. Microbiota and Cardiometabolic Diseases
5. Proposed Mechanism of Action for Microbiota and Cardiometabolic Diseases
6. Vitamin D and Microbiota in Cardiometabolic Diseases
7. Limitations
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cashman, K.D.; Kiely, M. Recommended dietary intakes for vitamin D: Where do they come from, what do they achieve and how can we meet them? J. Hum. Nutr. Diet. 2014, 27, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.W.; Lee, H.C. Vitamin D and health—The missing vitamin in humans. Pediatr. Neonatol. 2019, 60, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Valle, H.B.; Yaktine, A.L.; Taylor, C.L.; Ross, A.C. Dietary Reference Intakes for Calcium and Vitamin D; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Day, C. Metabolic syndrome, or What you will: Definitions and epidemiology. Diabetes Vasc. Dis. Res. 2007, 4, 32–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, J. A comprehensive review on metabolic syndrome. Cardiol. Res. Pract. 2014, 2014, 943162. [Google Scholar] [CrossRef]
- Christakos, S.; Li, S.; De La Cruz, J.; Bikle, D.D. New developments in our understanding of vitamin metabolism, action and treatment. Metabolism 2019, 98, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, W.; Li, D.; Yin, X.; Zhang, X.; Olsen, N.; Zheng, S.G. Vitamin D and Chronic Diseases. Aging Dis. 2017, 8, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Cojic, M.; Kocic, R.; Klisic, A.; Kocic, G. The Effects of Vitamin D Supplementation on Metabolic and Oxidative Stress Markers in Patients with Type 2 Diabetes: A 6-Month Follow Up Randomized Controlled Study. Front. Endocrinol. 2021, 12, 610893. [Google Scholar] [CrossRef]
- Ganji, V.; Tangpricha, V.; Zhang, X. Serum Vitamin D Concentration ≥75 nmol/L Is Related to Decreased Cardiometabolic and Inflammatory Biomarkers, Metabolic Syndrome, and Diabetes; and Increased Cardiorespiratory Fitness in US Adults. Nutrients 2020, 12, 730. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Cao, Z.; Lu, F.; Liu, Y.; Lv, Y.; Qu, Y.; Gu, H.; Li, C.; Cai, J.; Ji, S.; et al. Vitamin D deficiency and metabolic syndrome in elderly Chinese individuals: Evidence from CLHLS. Nutr. Metab. 2020, 17, 58. [Google Scholar] [CrossRef]
- Raposo, L.; Martins, S.; Ferreira, D.; Guimarães, J.T.; Santos, A.C. Vitamin D, parathyroid hormone and metabolic syndrome—The PORMETSYN study. BMC Endocr. Disord. 2017, 17, 71. [Google Scholar] [CrossRef]
- Ahmad, A.F.; Dwivedi, G.; O’Gara, F.; Caparros-Martin, J.; Ward, N.C. The gut microbiome and cardiovascular disease: Current knowledge and clinical potential. Am. J. Physiol. Heart Circ. Physiol. 2019, 317, H923–H938. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Tripathi, P. Gut microbiome and type 2 diabetes: Where we are and where to go? J. Nutr. Biochem. 2019, 63, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Carrelli, A.; Bucovsky, M.; Horst, R.; Cremers, S.; Zhang, C.; Bessler, M.; Schrope, B.; Evanko, J.; Blanco, J.; Silverberg, S.J.; et al. Vitamin D Storage in Adipose Tissue of Obese and Normal Weight Women. J. Bone Miner. Res. 2017, 32, 237–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pott-Junior, H.; Nascimento, C.M.C.; Costa-Guarisco, L.P.; Gomes, G.A.D.O.; Gramani-Say, K.; Orlandi, F.D.S.; Gratão, A.C.M.; Orlandi, A.A.D.S.; Pavarini, S.C.I.; Vasilceac, F.A.; et al. Vitamin D Deficient Older Adults Are More Prone to Have Metabolic Syndrome, but Not to a Greater Number of Metabolic Syndrome Parameters. Nutrients 2020, 12, 748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlAnouti, F.; Abboud, M.; Papandreou, D.; Mahboub, N.; Haidar, S.; Rizk, R. Effects of Vitamin D Supplementation on Lipid Profile in Adults with the Metabolic Syndrome: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2020, 12, 3352. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ballantyne, C.M. Metabolic Inflammation and Insulin Resistance in Obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, J.; DeLuca, H.F. Where is the vitamin D receptor? Arch. Biochem. Biophys. 2012, 523, 123–133. [Google Scholar] [CrossRef]
- Zeitz, U.; Weber, K.; Soegiarto, D.W.; Wolf, E.; Balling, R.; Erben, R.G. Impaired insulin secretory capacity in mice lacking a functional vitamin D receptor. FASEB J. 2003, 17, 509–511. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Ashraf, A. Role of vitamin d in insulin secretion and insulin sensitivity for glucose homeostasis. Int. J. Endocrinol. 2010, 2010, 351385. [Google Scholar] [CrossRef] [Green Version]
- Teegarden, D.; Donkin, S.S. Vitamin D: Emerging new roles in insulin sensitivity. Nutr. Res. Rev. 2009, 22, 82–92. [Google Scholar] [CrossRef]
- Sisley, S.R.; Arble, D.M.; Chambers, A.P.; Gutierrez-Aguilar, R.; He, Y.; Xu, Y.; Gardner, D.; Moore, D.D.; Seeley, R.J.; Sandoval, D.A. Hypothalamic Vitamin D Improves Glucose Homeostasis and Reduces Weight. Diabetes 2016, 65, 2732–2741. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, T.W.; Väisänen, S.; Frank, C.; Molnár, F.; Sinkkonen, L.; Carlberg, C. The human peroxisome proliferator-activated receptor delta gene is a primary target of 1alpha,25-dihydroxyvitamin D3 and its nuclear receptor. J. Mol. Biol. 2005, 349, 248–260. [Google Scholar] [CrossRef]
- Han, L.; Shen, W.J.; Bittner, S.; Kraemer, F.B.; Azhar, S. PPARs: Regulators of metabolism and as therapeutic targets in cardiovascular disease. Part II: PPAR-β/δ and PPAR-γ. Future Cardiol. 2017, 13, 279–296. [Google Scholar] [CrossRef]
- Lee, K.; Kim, J. Serum vitamin D status and metabolic syndrome: A systematic review and dose-response meta-analysis. Nutr. Res. Pract. 2021, 15, 329–345. [Google Scholar] [CrossRef]
- Jamka, M.; Woźniewicz, M.; Jeszka, J.; Mardas, M.; Bogdański, P.; Stelmach-Mardas, M. The effect of vitamin D supplementation on insulin and glucose metabolism in overweight and obese individuals: Systematic review with meta-analysis. Sci. Rep. 2015, 5, 16142. [Google Scholar] [CrossRef] [Green Version]
- Poolsup, N.; Suksomboon, N.; Plordplong, N. Effect of vitamin D supplementation on insulin resistance and glycaemic control in prediabetes: A systematic review and meta-analysis. Diabet. Med. 2016, 33, 290–299. [Google Scholar] [CrossRef]
- Chandler, P.D.; Wang, L.; Zhang, X.; Sesso, H.D.; Moorthy, M.V.; Obi, O.; Lewis, J.; Prince, R.L.; Danik, J.S.; Manson, J.E.; et al. Effect of vitamin D supplementation alone or with calcium on adiposity measures: A systematic review and meta-analysis of randomized controlled trials. Nutr. Rev. 2015, 73, 577–593. [Google Scholar] [CrossRef]
- Jorde, R.; Sollid, S.T.; Svartberg, J.; Schirmer, H.; Joakimsen, R.M.; Njølstad, I.; Fuskevåg, O.M.; Figenschau, Y.; Hutchinson, M.Y. Vitamin D 20 000 IU per week for five years does not prevent progression from prediabetes to diabetes. J. Clin. Endocrinol. Metab. 2016, 101, 1647–1655. [Google Scholar] [CrossRef] [Green Version]
- Gulseth, H.L.; Wium, C.; Angel, K.; Eriksen, E.F.; Birkeland, K.I. Effects of Vitamin D Supplementation on Insulin Sensitivity and Insulin Secretion in Subjects with Type 2 Diabetes and Vitamin D Deficiency: A Randomized Controlled Trial. Diabetes Care 2017, 40, 872–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemieux, P.; Weisnagel, J.S.; Caron, A.Z.; Julien, A.S.; Morisset, A.S.; Carreau, A.M.; Poirier, J.; Tchernof, A.; Robitaille, J.; Bergeron, J.; et al. Effects of 6-month vitamin D supplementation on insulin sensitivity and secretion: A randomized, placebo-controlled trial. Eur. J. Endocrinol. 2019, 181, 287–299. [Google Scholar] [CrossRef]
- Moreira-Lucas, T.S.; Duncan, A.M.; Rabasa-Lhoret, R.; Vieth, R.; Gibbs, A.L.; Badawi, A.; Wolever, T.M. Effect of vitamin D supplementation on oral glucose tolerance in individuals with low vitamin D status and increased risk for developing type 2 diabetes (EVIDENCE): A double-blind, randomized, placebo-controlled clinical trial. Diabetes Obes. Metab. 2016, 19, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Zhang, L.; Li, C.; Gai, Z.; Li, Y. Vitamin D and Vitamin D Receptor: New Insights in the Treatment of Hypertension. Curr. Protein Pept. Sci. 2019, 20, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Watts, S.W.; Ng, M.; Chen, S.; Glenn, D.J.; Gardner, D.G. Elimination of vitamin D receptor in vascular endothelial cells alters vascular function. Hypertension 2014, 64, 1290–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.C.; Qiao, G.; Uskokovic, M.; Xiang, W.; Zheng, W.; Kong, J. Vitamin D: A negative endocrine regulator of the renin-angiotensin system and blood pressure. J. Steroid Biochem. Mol. Biol. 2004, 89, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Kheiri, B.; Abdalla, A.; Osman, M.; Ahmed, S.; Hassan, M.; Bachuwa, M. Vitamin D deficiency and risk of cardiovascular diseases: A narrative review. Clin. Hypertens. 2018, 24, 9. [Google Scholar] [CrossRef]
- Ghobadi, S.; Rostami, Z.H.; Marzijarani, M.S.; Faghih, S. Association of Vitamin D Status and Metabolic Syndrome Components in Iranian Children. Int. J. Prev. Med. 2019, 10, 77. [Google Scholar]
- Javed, Z.; Papageorgiou, M.; Deshmukh, H.; Kilpatrick, E.S.; Mann, V.; Corless, L.; Abouda, G.; Rigby, A.S.; Atkin, S.L.; Sathyapalan, T. A Randomized, Controlled Trial of Vitamin D Supplementation on Cardiovascular Risk Factors, Hormones, and Liver Markers in Women with Polycystic Ovary Syndrome. Nutrients 2019, 11, 188. [Google Scholar] [CrossRef] [Green Version]
- Krivoy, A.; Onn, R.; Vilner, Y.; Hochman, E.; Weizman, S.; Paz, A.; Hess, S.; Sagy, R.; Kimhi-Nesher, S.; Kalter, E.; et al. Vitamin D Supplementation in Chronic Schizophrenia Patients Treated with Clozapine: A Randomized, Double-Blind, Placebo-controlled Clinical Trial. EBioMedicine 2017, 26, 138–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Yan, L.; Lu, Y.; Jiang, Q.; Pu, Y.; Sun, Q. Correction of hypovitaminosis D does not improve the metabolic syndrome risk profile in a Chinese population: A randomized controlled trial for 1 year. Asia Pac. J. Clin. Nutr. 2016, 25, 71–77. [Google Scholar]
- Talaei, A.; Mohamadi, M.; Adgi, Z. The effect of vitamin D on insulin resistance in patients with type 2 diabetes. Diabetol. Metab. Syndr. 2013, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Salekzamani, S.; Mehralizadeh, H.; Ghezel, A.; Salekzamani, Y.; Jafarabadi, M.A.; Bavil, A.S.; Gargari, B.P. Effect of high-dose vitamin D supplementation on cardiometabolic risk factors in subjects with metabolic syndrome: A randomized controlled double-blind clinical trial. J. Endocrinol. Investig. 2016, 39, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Bennouar, S.; Cherif, A.B.; Kessira, A.; Bennouar, D.E.; Abdi, S. Association and interaction between vitamin D level and metabolic syndrome for non-alcoholic fatty liver disease. J. Diabetes Metab. Disord. 2021, 20, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Yoo, T.; Choi, W.; Hong, J.H.; Lee, J.Y.; Kim, J.M.; Shin, I.S.; Yang, S.J.; Amminger, P.; Berk, M.; Yoon, J.S.; et al. Association Between Vitamin D Insufficiency and Metabolic Syndrome in Patients with Psychotic Disorders. Psychiatry Investig. 2018, 15, 396–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, C.; Reynolds, J.A.; Lertratanakul, A.; Wu, P.; Urowitz, M.; Gladman, D.D.; Fortin, P.R.; Bae, S.C.; Gordon, C.; Clarke, A.E.; et al. Lower vitamin D is associated with metabolic syndrome and insulin resistance in systemic lupus: Data from an international inception cohort. Rheumatology 2021, 60, 4737–4747. [Google Scholar] [CrossRef] [PubMed]
- Mirhosseini, N.; Rainsbury, J.; Kimball, S.M. Vitamin D Supplementation, Serum 25(OH)D Concentrations and Cardiovascular Disease Risk Factors: A Systematic Review and Meta-Analysis. Front. Cardiovasc. Med. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, S.; Ohno, H. Gut microbiome and metabolic diseases. Semin. Immunopathol. 2014, 36, 103–114. [Google Scholar] [CrossRef]
- Festi, D.; Schiumerini, R.; Eusebi, L.H.; Marasco, G.; Taddia, M.; Colecchia, A. Gut microbiota and metabolic syndrome. World J. Gastroenterol. 2014, 20, 16079–16094. [Google Scholar] [CrossRef]
- Belizário, J.E.; Faintuch, J. Microbiome and Gut Dysbiosis. Exp. Suppl. 2018, 109, 459–476. [Google Scholar]
- Kaushal, D.; Kalsi, G. Dietary intervention of prebiotics and vitamins on gut health of children. Nutr. Food Sci. 2022, 52, 1045–1054. [Google Scholar] [CrossRef]
- Malesza, I.J.; Malesza, M.; Walkowiak, J.; Mussin, N.; Walkowiak, D.; Aringazina, R.; Bartkowiak-Wieczorek, J.; Mądry, E. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells 2021, 10, 3164. [Google Scholar] [CrossRef]
- Backhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, N.; Zhao, L. An opportunistic pathogen isolated from the gut of an obese human causes obesity in germfree mice. ISME J. 2013, 7, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef] [PubMed]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga–Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916.e7, Correction in Gastroenterology 2013, 144, 250. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Duncan, S.H.; Lobley, G.E.; Holtrop, G.; Ince, J.; Johnstone, A.M.; Louis, P.; Flint, H.J. Human colonic microbiota associated with diet, obesity and weight loss. Int. J. Obes. 2008, 32, 1720–1724. [Google Scholar] [CrossRef] [Green Version]
- Tomas, J.; Mulet, C.; Saffarian, A.; Cavin, J.B.; Ducroc, R.; Regnault, B.; Kun Tan, C.; Duszka, K.; Burcelin, R.; Wahli, W.; et al. High-fat diet modifies the PPAR-gamma pathway leading to disruption of microbial and physiological ecosystem in murine small intestine. Proc. Natl. Acad. Sci. USA 2016, 113, E5934–E5943. [Google Scholar] [CrossRef] [Green Version]
- Pendyala, S.; Walker, J.M.; Holt, P.R. A high-fat diet is associated with endotoxemia that originates from the gut. Gastroenterology 2012, 142, 1100–1101.e2. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Creely, S.J.; McTernan, P.G.; Kusminski, C.M.; Fisher, F.M.; Da Silva, N.F.; Khanolkar, M.; Evans, M.; Harte, A.L.; Kumar, S. Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E740–E747. [Google Scholar] [CrossRef] [Green Version]
- Lau, W.L.; Tran, T.; Rhee, C.M.; Kalantar-Zadeh, K.; Vaziri, N.D. Diabetes and the Gut Microbiome. Semin. Nephrol. 2021, 41, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Rawat, A.; Saadaoui, M.; Elhag, D.; Tomei, S.; Elanbari, M.; Akobeng, A.K.; Mustafa, A.; Abdelgadir, I.; Udassi, S.; et al. Tipping the Balance: Vitamin D Inadequacy in Children Impacts the Major Gut Bacterial Phyla. Biomedicines 2022, 10, 278. [Google Scholar] [CrossRef] [PubMed]
- Amar, J.; Serino, M.; Lange, C.; Chabo, C.; Iacovoni, J.; Mondot, S.; Lepage, P.; Klopp, C.; Mariette, J.; Bouchez, O.; et al. Involvement of tissue bacteria in the onset of diabetes in humans: Evidence for a concept. Diabetologia 2011, 54, 3055–3061. [Google Scholar] [CrossRef]
- Yoshida, N.; Emoto, T.; Yamashita, T.; Watanabe, H.; Hayashi, T.; Tabata, T.; Hoshi, N.; Hatano, N.; Ozawa, G.; Sasaki, N.; et al. Bacteroides vulgatus and Bacteroides dorei Reduce Gut Microbial Lipopolysaccharide Production and Inhibit Atherosclerosis. Circulation 2018, 138, 2486–2498. [Google Scholar] [CrossRef]
- Koh, A.; Molinaro, A.; Ståhlman, M.; Khan, M.T.; Schmidt, C.; Mannerås-Holm, L.; Wu, H.; Carreras, A.; Jeong, H.; Olofsson, L.E.; et al. Microbially Produced Imidazole Propionate Impairs Insulin Signaling through mTORC1. Cell 2018, 175, 947–961.e17. [Google Scholar] [CrossRef] [Green Version]
- Caesar, R.; Reigstad, C.S.; Bäckhed, H.K.; Reinhardt, C.; Ketonen, M.; Lundén, G.Ö.; Cani, P.D.; Bäckhed, F. Gut-derived lipopolysaccharide augments adipose macrophage accumulation but is not essential for impaired glucose or insulin tolerance in mice. Gut 2012, 61, 1701–1707. [Google Scholar] [CrossRef] [Green Version]
- Rajagopala, S.V.; Vashee, S.; Oldfield, L.M.; Suzuki, Y.; Venter, J.C.; Telenti, A.; Nelson, K.E. The Human Microbiome and Cancer. Cancer Prev. Res. 2017, 10, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Lach, G.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. Anxiety, Depression, and the Microbiome: A Role for Gut Peptides. Neurotherapeutics 2018, 15, 36–59. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Zogg, H.; Wei, L.; Bartlett, A.; Ghoshal, U.C.; Rajender, S.; Ro, S. Gut Microbial Dysbiosis in the Pathogenesis of Gastrointestinal Dysmotility and Metabolic Disorders. J. Neurogastroenterol. Motil. 2021, 27, 19–34. [Google Scholar] [CrossRef]
- Bakke, D.; Chatterjee, I.; Agrawal, A.; Dai, Y.; Sun, J. Regulation of Microbiota by Vitamin D Receptor: A Nuclear Weapon in Metabolic Diseases. Nucl. Recept. Res. 2018, 5, 101377. [Google Scholar] [CrossRef] [Green Version]
- Su, D.; Nie, Y.; Zhu, A.; Chen, Z.; Wu, P.; Zhang, L.; Luo, M.; Sun, Q.; Cai, L.; Lai, Y.; et al. Vitamin D Signaling through Induction of Paneth Cell Defensins Maintains Gut Microbiota and Improves Metabolic Disorders and Hepatic Steatosis in Animal Models. Front. Physiol. 2016, 7, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.; Wu, S.; Zhang, Y.G.; Lu, R.; Xia, Y.; Dong, H.; Sun, J. Lack of Vitamin D Receptor Causes Dysbiosis and Changes the Functions of the Murine Intestinal Microbiome. Clin. Ther. 2015, 37, 996–1009.e7. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Xia, Y.; Sun, J. Imbalance of the intestinal virome and altered viral-bacterial interactions caused by a conditional deletion of the vitamin D receptor. Gut Microbes 2021, 13, 1957408. [Google Scholar] [CrossRef]
- Luthold, R.V.; Fernandes, G.R.; Franco-de-Moraes, A.C.; Folchetti, L.G.; Ferreira, S.R.G. Gut microbiota interactions with the immunomodulatory role of vitamin D in normal individuals. Metabolism 2017, 69, 76–86. [Google Scholar] [CrossRef]
- Hussein, H.M.; Elyamany, M.F.; Rashed, L.A.; Sallam, N.A. Vitamin D mitigates diabetes-associated metabolic and cognitive dysfunction by modulating gut microbiota and colonic cannabinoid receptor 1. Eur. J. Pharm. Sci. 2022, 170, 106105. [Google Scholar] [CrossRef]
- Voland, L.; Le Roy, T.; Debédat, J.; Clément, K. Gut microbiota and vitamin status in persons with obesity: A key interplay. Obes. Rev. 2022, 23, e13377. [Google Scholar] [CrossRef]
- Singh, P.; Rawat, A.; Alwakeel, M.; Sharif, E.; Al Khodor, S. The potential role of vitamin D supplementation as a gut microbiota modifier in healthy individuals. Sci. Rep. 2020, 10, 21641. [Google Scholar] [CrossRef]


{kind=link}
| Reference | Study Design | Study Sample & I/C | Dose and Duration of Vitamin D Supplementation | Outcome Measurements | Findings Related to Cardiometabolic Diseases 2 | Other Findings 2 |
|---|---|---|---|---|---|---|
| Cojic et al. [8] | RCT | Subjects with T2DM n = 130 (65/65) | Vitamin D deficient: 50,000 IU/week vitamin D, first 3 months and 14,000 IU/week, next 3 months Vitamin D sufficient: 14,000 IU/week, 6 months | FBG, HDL-C TG, WC, BP, HbA1c | Effect of time on FBG and BP was significant ↓HbA1c | |
| Javed et al. [38] | Double-blind RCT | Women with PCOS n = 37 (18/19) | 3200 IU/day vitamin D, 3 months | BP, FBG, TG, HDL-C, CRP | ↓HOMA-IR, ALT, E LF score, Hyaluronic acid | |
| Krivoy et al. [39] | Double-blind RCT | Subjects with schizophrenia n = 47 (24/23) | 14,000 IU/week, 8 weeks | WC, HDL-C, TG, FBG, SBP | No significant difference between treatment and placebo groups | ↑total MoCA score (cognitive performance) |
| Yin et al. [40] | RCT | Vitamin D deficient n = 123 (61/62) | 700 IU/day vitamin D, 12 months | FBG, TG, HDL-C, WC, BP | No significant difference between treatment and placebo groups | ↓serum PTH |
| Talaei et al. [41] | Single arm study | Subjects with T2DM n = 100 | 50,000 IU/week vitamin D, 8 weeks | FBG, HDL-C, insulin, HOMA-IR | ↓FBG, insulin, HOMA-IR | - |
| Salekzamani et al. [42] | Double-blind RCT | Subjects with MetSyn n = 71 (35/36) | 50,000 IU/week vitamin D, 16 weeks | FBG, HDL-C, TG, WC, BP | ↓TG | - |
| Reference | Study Design | Study Sample | Vitamin D Assessment | Outcome Measurements: Biomarker of Cardiometabolic Diseases | Findings Related to Cardiometabolic Diseases 2 |
|---|---|---|---|---|---|
| Ganji et al. [9] | Cross-sectional | NHANES n = 8241 | Serum 25(OH)D | FBG, HDL-C, TG, WC, BP, MetSyn prevalence | ↓DBP, TG, WC, HbA1c, serum insulin, C-peptide, CRP, tHcy, HOMA-IR, MetSyn prevalence ↑HDL-C |
| Liu et al. [10] | Cross-sectional | Elderly n = 2493 | Serum 25(OH)D | FBG, HDL-C TG, WC, BP, MetSyn prevalence | ↓TG, MetSyn prevalence ↑HDL-C |
| Raposo et al. [11] | Cross-sectional | Adults n = 4095 | Serum 25(OH)D | FBG, HDL-C TG, WC, BP, MetSyn Prevalence | ↓TG, BP |
| Pott-Junior et al. [15] | Cross-sectional | Older adults n = 265 | Serum 25(OH)D | FBG, HDL-C, TG, WC, BP, MetSyn prevalence | ↓MetSyn prevalence, IR |
| Bennouar et al. [43] | Cross-sectional | Adults with and without NAFLD n = 874 | Serum 25(OH)D | FBG, HDL-C, TG, WC, BP, MetSyn prevalence | ↓MetSyn prevalence |
| Yoo et al. [44] | Cross-sectional | Subjects with psychotic disorders n = 302 | Serum 25(OH)D | FBG, HDL-C, TG, WC, BP, MetSyn prevalence | ↓MetSyn prevalence, BP |
| Chew et al. [45] | Multi-center cross-sectional | Patients with lupus erythematosus n = 1163 | Serum 25(OH)D | FBG, HDL-C, TG WC, BP, MetSyn prevalence | ↓BP, TG, HOMA-IR, MetSyn prevalence ↑HDL |
| Reference | Study Design | Study Sample Characteristics | Interventions | Measurements | Findings 2 (Association with Microbiota) |
|---|---|---|---|---|---|
| Backhed et al. [52] | Experimental | Germ-free mice and conventionally raised mice | Conventionalization with microbiota from CONV-R | Body composition, IR | ↑60% body fat, IR, hepatic TG synthesis |
| Fei and Zhao [53] | Experimental | Germ-free mice | LPS (endotoxin) transfer | Obesity Insulin resistance | ↑Obesity, IR |
| Vrieze et al. [55] | Experimental | Lean (n = 9) and individuals with MetSyn (n = 9) (6 week) | Intestinal microbiota transfer | Insulin Sensitivity Large and small intestine gut microbiota composition | ↑butyrate-producing intestinal microbiota, insulin sensitivity |
| Cani et al. [60] | Experimental | Mice | High fat diet (4 week) Subcutaneous infusion of LPS | Insulin sensitivity Oral glucose tolerance tests LPS tolerance test | ↑LPS containing microbiota in the gut, circulating LPS, IR, hyperglycemia, hyperinsulinemia, weight gain |
| Amar et al. [64] | Longitudinal | Without diabetes or obesity at baseline (9 years of follow-up) n = 3280 | - | Diabetes, 16S rDNA concentration | ↑16S rDNA (with diabetes) |
| Pendyala et al. [59] | Cohort | n = 8 | High-fat Western-style diets (1 month) | Gastrointestinal barrier function, Microbiota composition | ↑Plasma endotoxins |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sukik, A.; Alalwani, J.; Ganji, V. Vitamin D, Gut Microbiota, and Cardiometabolic Diseases—A Possible Three-Way Axis. Int. J. Mol. Sci. 2023, 24, 940. https://doi.org/10.3390/ijms24020940
Sukik A, Alalwani J, Ganji V. Vitamin D, Gut Microbiota, and Cardiometabolic Diseases—A Possible Three-Way Axis. International Journal of Molecular Sciences. 2023; 24(2):940. https://doi.org/10.3390/ijms24020940
Chicago/Turabian StyleSukik, Ayah, Joud Alalwani, and Vijay Ganji. 2023. "Vitamin D, Gut Microbiota, and Cardiometabolic Diseases—A Possible Three-Way Axis" International Journal of Molecular Sciences 24, no. 2: 940. https://doi.org/10.3390/ijms24020940
APA StyleSukik, A., Alalwani, J., & Ganji, V. (2023). Vitamin D, Gut Microbiota, and Cardiometabolic Diseases—A Possible Three-Way Axis. International Journal of Molecular Sciences, 24(2), 940. https://doi.org/10.3390/ijms24020940