
The Pressurized Skin: A Review on the Pathological Effect of Mechanical Pressure on the Skin from the Cellular Perspective



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
3. Results
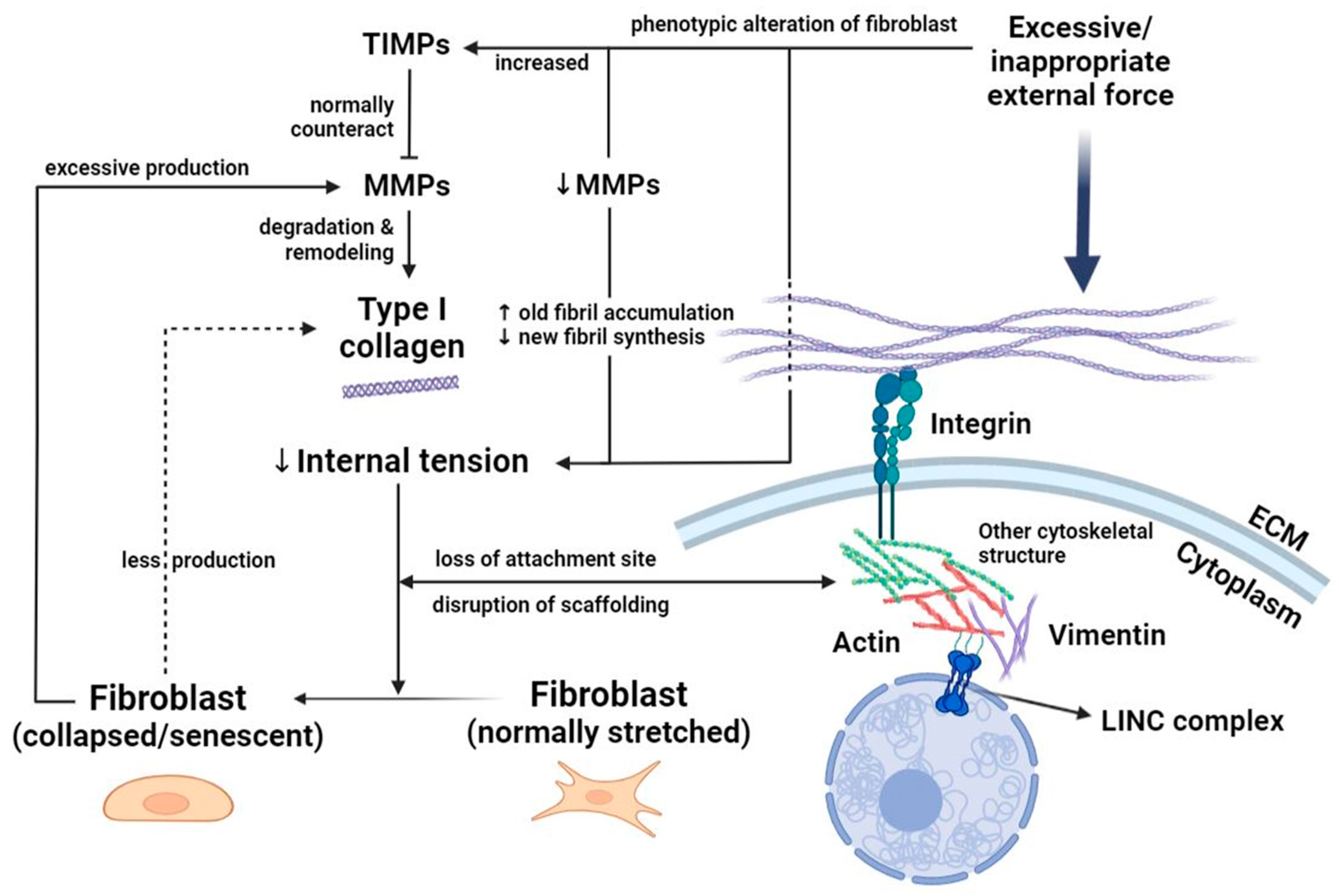
3.1. Fibroblasts: Heterogeneous Responders to Mechanical Load
3.2. Keratinocytes: The Vanguard against Pressure
3.3. Mast Cells: Degranulation Facilitates Neuronal Mechanotransduction
3.4. Melanocytes: Cellular and Molecular Crosstalk Expedites Mechanosignaling
3.5. Adipocytes and Stem Cells: Versatile Navigators of Cell Fate upon Pressure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hsu, C.K.; Lin, H.H.; Harn, H.I.; Hughes, M.W.; Tang, M.J.; Yang, C.C. Mechanical forces in skin disorders. J. Dermatol. Sci. 2018, 90, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Devillers, C.; Pierard-Franchimont, C.; Schreder, A.; Docquier, V.; Pierard, G.E. High resolution skin colorimetry, strain mapping and mechanobiology. Int. J. Cosmet. Sci. 2010, 32, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Hajhosseini, B.; Longaker, M.T.; Gurtner, G.C. Pressure injury. Ann. Surg. 2020, 271, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Aragona, M.; Sifrim, A.; Malfait, M.; Song, Y.; Van Herck, J.; Dekoninck, S.; Gargouri, S.; Lapouge, G.; Swedlund, B.; Dubois, C.; et al. Mechanisms of stretch-mediated skin expansion at single-cell resolution. Nature 2020, 584, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Bouten, C.V.; Oomens, C.W.; Baaijens, F.P.; Bader, D.L. The etiology of pressure ulcers: Skin deep or muscle bound? Arch. Phys. Med. Rehabil. 2003, 84, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Biggs, L.C.; Kim, C.S.; Miroshnikova, Y.A.; Wickström, S.A. Mechanical forces in the skin: Roles in tissue architecture, stability, and function. J. Investig. Dermatol. 2020, 140, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Lichtenberger, B.M.; Hoste, E.; Kretzschmar, K.; Simons, B.D.; Charalambous, M.; Ferron, S.R.; Herault, Y.; Pavlovic, G.; Ferguson-Smith, A.C.; et al. Distinct fibroblast lineages determine dermal architecture in skin development and repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Talbott, H.E.; Mascharak, S.; Griffin, M.; Wan, D.C.; Longaker, M.T. Wound healing, fibroblast heterogeneity, and fibrosis. Cell Stem Cell 2022, 29, 1161–1180. [Google Scholar] [CrossRef]
- Tan, P.C.; Zhou, S.B.; Ou, M.Y.; He, J.Z.; Zhang, P.Q.; Zhang, X.J.; Xie, Y.; Gao, Y.M.; Zhang, T.Y.; Li, Q.F. Mechanical stretching can modify the papillary dermis pattern and papillary fibroblast characteristics during skin regeneration. J. Investig. Dermatol. 2022, 142, 2384–2394.e8. [Google Scholar] [CrossRef]
- Sawant, M.; Hinz, B.; Schönborn, K.; Zeinert, I.; Eckes, B.; Krieg, T.; Schuster, R. A story of fibers and stress: Matrix-embedded signals for fibroblast activation in the skin. Wound Repair Regen. 2021, 29, 515–530. [Google Scholar] [CrossRef]
- Karakesisoglou, I.; Mroß, C.; Noegel, A.A. Functional analysis of LINC complexes in the skin. Methods Mol. Biol. 2018, 1840, 295–306. [Google Scholar] [PubMed]
- Gupta, R.; Misri, R.; Gupta, A.; Chowdhary, M.; Singh, A. Genome-wide profiling reveals pervasive transcriptional alterations in fibroblasts derived from lesional skin in vitiligo including a reduced potential to proliferate. Exp. Dermatol. 2022, 32, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Varani, J.; Voorhees, J.J. Looking older: Fibroblast collapse and therapeutic implications. Arch. Dermatol. 2008, 144, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Silver, F.H.; Kelkar, N.; Deshmukh, T. Molecular basis for mechanical properties of ECMs: Proposed role of fibrillar collagen and proteoglycans in tissue biomechanics. Biomolecules 2021, 11, 1018. [Google Scholar] [CrossRef] [PubMed]
- Brun, C.; Jean-Louis, F.; Oddos, T.; Bagot, M.; Bensussan, A.; Michel, L. Phenotypic and functional changes in dermal primary fibroblasts isolated from intrinsically aged human skin. Exp. Dermatol. 2016, 25, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Quan, T.; Purohit, T.; Shao, Y.; Cho, M.K.; He, T.; Varani, J.; Kang, S.; Voorhees, J.J. Collagen fragmentation promotes oxidative stress and elevates matrix metalloproteinase-1 in fibroblasts in aged human skin. Am. J. Pathol. 2009, 174, 101–114. [Google Scholar] [CrossRef] [PubMed]
- de Bengy, A.F.; Lamartine, J.; Sigaudo-Roussel, D.; Fromy, B. Newborn and elderly skin: Two fragile skins at higher risk of pressure injury. Biol. Rev. Camb. Philos. Soc. 2022, 97, 874–895. [Google Scholar] [CrossRef]
- Fisher, G.J.; Wang, B.; Cui, Y.; Shi, M.; Zhao, Y.; Quan, T.; Voorhees, J.J. Skin aging from the perspective of dermal fibroblasts: The interplay between the adaptation to the extracellular matrix microenvironment and cell autonomous processes. J. Cell Commun. Signal. 2023, 17, 523–529. [Google Scholar] [CrossRef]
- Riethmuller, C. Assessing the skin barrier via corneocyte morphometry. Exp. Dermatol. 2018, 27, 923–930. [Google Scholar] [CrossRef]
- Wang, G.; Sweren, E.; Andrews, W.; Li, Y.; Chen, J.; Xue, Y.; Wier, E.; Alphonse, M.P.; Luo, L.; Miao, Y.; et al. Commensal microbiome promotes hair follicle regeneration by inducing keratinocyte HIF-1α signaling and glutamine metabolism. Sci. Adv. 2023, 9, eabo7555. [Google Scholar] [CrossRef]
- Evora, A.S.; Adams, M.J.; Johnson, S.A.; Zhang, Z. Corneocytes: Relationship between structural and biomechanical properties. Skin Pharmacol. Physiol. 2021, 34, 146–161. [Google Scholar] [CrossRef] [PubMed]
- Moehring, F.; Halder, P.; Seal, R.P.; Stucky, C.L. Uncovering the cells and circuits of touch in normal and pathological settings. Neuron 2018, 100, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Denda, M.; Nakanishi, S. Do epidermal keratinocytes have sensory and information processing systems? Exp. Dermatol. 2022, 31, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.J.; Thomson, C.; Lunny, D.P.; Dopping-Hepenstal, P.J.; McGrath, J.A.; Smith, F.J.D.; Irwin McLean, W.H.; Leslie Pedrioli, D.M. Keratin 9 is required for the structural integrity and terminal differentiation of the palmoplantar epidermis. J. Investig. Dermatol. 2014, 134, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Maruthappu, T.; Chikh, A.; Fell, B.; Delaney, P.J.; Brooke, M.A.; Levet, C.; Moncada-Pazos, A.; Ishida-Yamamoto, A.; Blaydon, D.; Waseem, A.; et al. Rhomboid family member 2 regulates cytoskeletal stress-associated keratin 16. Nat. Commun. 2017, 8, 14174. [Google Scholar] [CrossRef] [PubMed]
- Zieman, A.G.; Poll, B.G.; Ma, J.; Coulombe, P.A. Altered keratinocyte differentiation is an early driver of keratin mutation-based palmoplantar keratoderma. Hum. Mol. Genet. 2019, 28, 2255–2270. [Google Scholar] [CrossRef] [PubMed]
- Moehring, F.; Cowie, A.M.; Menzel, A.D.; Weyer, A.D.; Grzybowski, M.; Arzua, T.; Geurts, A.M.; Palygin, O.; Stucky, C.L. Keratinocytes mediate innocuous and noxious touch via ATP-P2X4 signaling. eLife 2018, 7, e31684. [Google Scholar] [CrossRef] [PubMed]
- Mikesell, A.R.; Isaeva, O.; Moehring, F.; Sadler, K.E.; Menzel, A.D.; Stucky, C.L. Keratinocyte PIEZO1 modulates cutaneous mechanosensation. eLife 2022, 11, e65987. [Google Scholar] [CrossRef]
- Huang, X.; Liang, X.; Zhou, Y.; Li, H.; Du, H.; Suo, Y.; Liu, W.; Jin, R.; Chai, B.; Duan, R.; et al. CDH1 is identified as a therapeutic target for skin regeneration after mechanical loading. Int. J. Biol. Sci. 2021, 17, 353–367. [Google Scholar] [CrossRef]
- Corley, S.M.; Mendoza-Reinoso, V.; Giles, N.; Singer, E.S.; Common, J.E.; Wilkins, M.R.; Beverdam, A. Plau and Tgfbr3 are YAP-regulated genes that promote keratinocyte proliferation. Cell Death Dis. 2018, 9, 1106. [Google Scholar] [CrossRef]
- Tay, S.S.; Roediger, B.; Tong, P.L.; Tikoo, S.; Weninger, W. The skin-resident immune network. Curr. Dermatol. Rep. 2013, 3, 13–22. [Google Scholar] [CrossRef]
- Zhang, S.; Sumpter, T.L.; Kaplan, D.H. Neuron-mast cell cross-talk in the skin. J. Investig. Dermatol. 2022, 142, 841–848. [Google Scholar] [CrossRef]
- Zuberbier, T.; Aberer, W.; Asero, R.; Abdul Latiff, A.H.; Baker, D.; Ballmer-Weber, B.; Bernstein, J.A.; Bindslev-Jensen, C.; Brzoza, Z.; Buense Bedrikow, R.; et al. The EAACI/GA2LEN/EDF/WAO guideline for the definition, classification, diagnosis and management of urticaria. Allergy 2018, 73, 1393–1414. [Google Scholar] [CrossRef] [PubMed]
- Green, D.P.; Limjunyawong, N.; Gour, N.; Pundir, P.; Dong, X. A mast-cell-specific receptor mediates neurogenic inflammation and pain. Neuron 2019, 101, 412–420.e3. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Yang, H.; Yin, N.; Ding, G. Mast cell-nerve cell interaction at acupoint: Modeling mechanotransduction pathway induced by acupuncture. Int. J. Biol. Sci. 2014, 10, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.W.; Liu, X.Y.; Shen, Z.F.; Yao, W.; Gong, X.B.; Huang, H.X.; Ding, G.H. An investigation of the distribution and location of mast cells affected by the stiffness of substrates as a mechanical niche. Int. J. Biol. Sci. 2018, 14, 1142–1152. [Google Scholar] [CrossRef] [PubMed]
- Church, M.K.; Kolkhir, P.; Metz, M.; Maurer, M. The role and relevance of mast cells in urticaria. Immunol. Rev. 2018, 282, 232–247. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.R.; Ansary, T.M.; Komine, M.; Ohtsuki, M. Diversified stimuli-induced inflammatory pathways cause skin pigmentation. Int. J. Mol. Sci. 2021, 22, 3970. [Google Scholar] [CrossRef]
- Domingues, L.; Hurbain, I.; Gilles-Marsens, F.; Sirés-Campos, J.; André, N.; Dewulf, M.; Romao, M.; Viaris de Lesegno, C.; Macé, A.S.; Blouin, C.; et al. Coupling of melanocyte signaling and mechanics by caveolae is required for human skin pigmentation. Nat. Commun. 2020, 11, 2988. [Google Scholar] [CrossRef] [PubMed]
- Takamura, N.; Yamaguchi, Y. Involvement of caveolin-1 in skin diseases. Front. Immunol. 2022, 13, 1035451. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Scherer, P.E. Caveolin-1 as a pathophysiological factor and target in psoriasis. NPJ Aging Mech. Dis. 2019, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Kim, H.; Min, K.I.; Kim, C.; Kwon, Y.; Zheng, Z.; Kim, Y.; Park, H.S.; Ju, Y.S.; Roh, M.R.; et al. Weight-bearing activity impairs nuclear membrane and genome integrity via YAP activation in plantar melanoma. Nat. Commun. 2022, 13, 2214. [Google Scholar] [CrossRef] [PubMed]
- Panciera, T.; Azzolin, L.; Cordenonsi, M.; Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nat. Rev. Mol. Cell. Biol. 2017, 18, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Su, J.; Liu, J.; Yi, X.; Zhou, F.; Zhang, J.; Wang, J.; Meng, X.; Si, L.; Wu, C. YAP activation in promoting negative durotaxis and acral melanoma progression. Cells 2022, 11, 3543. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, K.; Hemmersbach, R. Guanylyl cyclase-cGMP signaling pathway in melanocytes: Differential effects of altered gravity in non-metastatic and metastatic cells. Int. J. Mol. Sci. 2020, 21, 1139. [Google Scholar] [CrossRef] [PubMed]
- Svedman, P. Mechanical homeostasis regulating adipose tissue volume. Head Face Med. 2007, 3, 34. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Scherer, P.E. General theory of skin reinforcement. PLoS ONE 2017, 12, e0182865. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, H.K.; Kim, K.S. Intrinsic and extrinsic mechanical properties related to the differentiation of mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2016, 473, 752–757. [Google Scholar] [CrossRef]
- Xing, Y.; Naik, S. Under pressure: Stem cell-niche interactions coordinate tissue adaptation to inflammation. Curr. Opin. Cell. Biol. 2020, 67, 64–70. [Google Scholar] [CrossRef]
- Doucet, Y.S.; Owens, D.M. Isolation and functional assessment of cutaneous stem cells. Methods Mol. Biol. 2015, 1235, 147–164. [Google Scholar]
- Liang, X.; Huang, X.; Zhou, Y.; Jin, R.; Li, Q. Mechanical stretching promotes skin tissue regeneration via enhancing mesenchymal stem cell homing and transdifferentiation. Stem Cells Transl. Med. 2016, 5, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Spector, M. In vitro response of the bone marrow-derived mesenchymal stem cells seeded in a type-I collagen-glycosaminoglycan scaffold for skin wound repair under the mechanical loading condition. Mol. Cell. Biomech. 2009, 6, 217–227. [Google Scholar] [PubMed]
- Piroli, M.E.; Jabbarzadeh, E. Matrix stiffness modulates mesenchymal stem cell sensitivity to geometric asymmetry signals. Ann. Biomed. Eng. 2018, 46, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.; Almeida, F.V.; Connelly, J.T. Biophysical signals controlling cell fate decisions: How do stem cells really feel? Int. J. Biochem. Cell Biol. 2012, 44, 2233–2237. [Google Scholar] [CrossRef] [PubMed]
- Shojaei, S.; Tafazzoli-Shahdpour, M.; Shokrgozar, M.A.; Haghighipour, N. Effects of mechanical and chemical stimuli on differentiation of human adipose-derived stem cells into endothelial cells. Int. J. Artif. Organs. 2013, 36, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Panayi, A.; Fan, J.; Mayer, H.F.; Daya, M.; Khouri, R.K.; Gurtner, G.C.; Ogawa, R.; Orgill, D.P. Mechanotransduction in wound healing: From the cellular and molecular level to the clinic. Adv. Skin Wound Care 2021, 34, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Bravo, B.; García de Durango, C.; González, Á.; Gortázar, A.R.; Santos, X.; Forteza-Vila, J.; Vidal-Vanaclocha, F. Opposite effects of mechanical action of fluid flow on proangiogenic factor secretion from human adipose-derived stem cells with and without oxidative stress. J. Cell Physiol. 2017, 232, 2158–2167. [Google Scholar] [CrossRef]
- Shoham, N.; Gottlieb, R.; Sharabani-Yosef, O.; Zaretsky, U.; Benayahu, D.; Gefen, A. Static mechanical stretching accelerates lipid production in 3T3-L1 adipocytes by activating the MEK signaling pathway. Am. J. Physiol. Cell Physiol. 2012, 302, C429–C441. [Google Scholar] [CrossRef]
- Tanabe, Y.; Koga, M.; Saito, M.; Matsunaga, Y.; Nakayama, K. Inhibition of adipocyte differentiation by mechanical stretching through ERK-mediated downregulation of PPARgamma2. J. Cell Sci. 2004, 117, 3605–3614. [Google Scholar] [CrossRef]
- Liao, Y.; Zeng, Z.; Lu, F.; Dong, Z.; Chang, Q.; Gao, J. In vivo dedifferentiation of adult adipose cells. PLoS ONE 2015, 10, e0125254. [Google Scholar] [CrossRef]
- Ma, J.; Xia, M.D.J.; Gao, J.; Lu, F.; Liao, Y. Mechanical signals induce dedifferentiation of mature adipocytes and increase the retention rate of fat grafts. Plast. Reconstr. Surg. 2019, 144, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Côté, J.A.; Ostinelli, G.; Gauthier, M.F.; Lacasse, A.; Tchernof, A. Focus on dedifferentiated adipocytes: Characteristics, mechanisms, and possible applications. Cell Tissue Res. 2019, 378, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.; Gefen, A. An integrated experimental-computational study of the microclimate under dressings applied to intact weight-bearing skin. Int. Wound J. 2020, 17, 562–577. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Minematsu, T.; Yoshida, M.; Nishijima, Y.; Noguchi, H.; Ohta, Y.; Nakagami, G.; Mori, T.; Sanada, H. Compression-induced HIF-1 enhances thrombosis and PAI-1 expression in mouse skin. Wound Repair Regen. 2015, 23, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cao, T.; Li, Q.; Zhang, J.; Du, J.; Chen, J.; Bai, Y.; Hao, J.; Zhu, Z.; Qiao, H.; et al. Cross-disease characterization of fibroblast heterogeneities and their pathogenic roles in skin inflammation. Clin. Immunol. 2023, 255, 109742. [Google Scholar] [CrossRef] [PubMed]
- Cassano, N.; Mastrandrea, V.; Vestita, M.; Vena, G.A. An overview of delayed pressure urticaria with special emphasis on pathogenesis and treatment. Dermatol. Ther. 2009, 22, S22–S26. [Google Scholar] [CrossRef] [PubMed]
- Aragona, M.; Panciera, T.; Manfrin, A.; Giulitti, S.; Michielin, F.; Elvassore, N.; Dupont, S.; Piccolo, S. A mechanical checkpoint controls multicellular growth through YAP/TAZ regulation by actin-processing factors. Cell 2013, 154, 1047–1059. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef]
- Totaro, A.; Castellan, M.; Battilana, G.; Zanconato, F.; Azzolin, L.; Giulitti, S.; Cordenonsi, M.; Piccolo, S. YAP/TAZ link cell mechanics to Notch signalling to control epidermal stem cell fate. Nat. Commun. 2017, 8, 15206. [Google Scholar] [CrossRef]
- Akladios, B.; Mendoza-Reinoso, V.; Samuel, M.S.; Hardeman, E.C.; Khosrotehrani, K.; Key, B.; Beverdam, A. Epidermal YAP2-5SA-ΔC drives β-catenin activation to promote keratinocyte proliferation in mouse skin in vivo. J. Investig. Dermatol. 2017, 137, 716–726. [Google Scholar] [CrossRef]
- Huang, S.; Chen, Z.; Hou, X.; Han, K.; Xu, B.; Zhang, M.; Ding, S.; Wang, Y.; Yang, Y. Promotion of melanoma cell proliferation by cyclic straining through regulatory morphogenesis. Int. J. Mol. Sci. 2022, 23, 11884. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; He, L.; Yang, Z. Recent advances in the role of Yes-associated protein in dermatosis. Skin Res. Technol. 2023, 29, e13285. [Google Scholar] [CrossRef] [PubMed]
- Brewer, C.M.; Nelson, B.R.; Wakenight, P.; Collins, S.J.; Okamura, D.M.; Dong, X.R.; Mahoney, W.M., Jr.; McKenna, A.; Shendure, J.; Timms, A.; et al. Adaptations in Hippo-Yap signaling and myofibroblast fate underlie scar-free ear appendage wound healing in spiny mice. Dev. Cell. 2021, 56, 2722–2740.e6. [Google Scholar] [CrossRef] [PubMed]
- McCluggage, W.G.; Smith, J.H. Reactive fibroblastic and myofibroblastic proliferation of the vulva (cyclist’s nodule): A hitherto poorly described vulval lesion occurring in cyclists. Am. J. Surg. Pathol. 2011, 35, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Padmanaban, D.; Rangasami, R.; Chandrasekharan, A.; Swaminathan, R. Decubital ischemic fasciitis presenting in an unusual location. Radiol. Case Rep. 2015, 6, 590. [Google Scholar] [CrossRef] [PubMed]
- Bensa, T.; Tekkela, S.; Rognoni, E. Skin fibroblast functional heterogeneity in health and disease. J. Pathol. 2023, 260, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.W.; Fernández-Yagüe, M.A.; Holland, E.N.; García, A.F.; Castro, N.S.; O’Neill, E.B.; Eyckmans, J.; Chen, C.S.; Fu, J.; Schlaepfer, D.D.; et al. Force-FAK signaling coupling at individual focal adhesions coordinates mechanosensing and microtissue repair. Nat. Commun. 2021, 12, 2359. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Hossain, M.Z.; Nieves, A.; Gu, L.; Ratliff, T.S.; Mi Oh, S.; Park, A.; Han, S.; Yang, N.B.; Qi, J.; et al. To control site-specific skin gene expression, autocrine mimics paracrine canonical Wnt signaling and is activated ectopically in skin disease. Am. J. Pathol. 2016, 186, 1140–1150. [Google Scholar] [CrossRef]
- Yang, L.; Rong, G.C.; Wu, Q.N. Diabetic foot ulcer: Challenges and future. World J. Diabetes 2022, 13, 1014–1034. [Google Scholar] [CrossRef]
- Thomas, D.R. Does pressure cause pressure ulcers? An inquiry into the etiology of pressure ulcers. J. Am. Med. Dir. Assoc. 2010, 11, 397–405. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chien, W.-C.; Tsai, T.-F. The Pressurized Skin: A Review on the Pathological Effect of Mechanical Pressure on the Skin from the Cellular Perspective. Int. J. Mol. Sci. 2023, 24, 15207. https://doi.org/10.3390/ijms242015207
Chien W-C, Tsai T-F. The Pressurized Skin: A Review on the Pathological Effect of Mechanical Pressure on the Skin from the Cellular Perspective. International Journal of Molecular Sciences. 2023; 24(20):15207. https://doi.org/10.3390/ijms242015207
Chicago/Turabian StyleChien, Wei-Chen, and Tsen-Fang Tsai. 2023. "The Pressurized Skin: A Review on the Pathological Effect of Mechanical Pressure on the Skin from the Cellular Perspective" International Journal of Molecular Sciences 24, no. 20: 15207. https://doi.org/10.3390/ijms242015207
APA StyleChien, W. -C., & Tsai, T. -F. (2023). The Pressurized Skin: A Review on the Pathological Effect of Mechanical Pressure on the Skin from the Cellular Perspective. International Journal of Molecular Sciences, 24(20), 15207. https://doi.org/10.3390/ijms242015207