
Effect of Hydralazine on Angiotensin II-Induced Abdominal Aortic Aneurysm in Apolipoprotein E-Deficient Mice

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
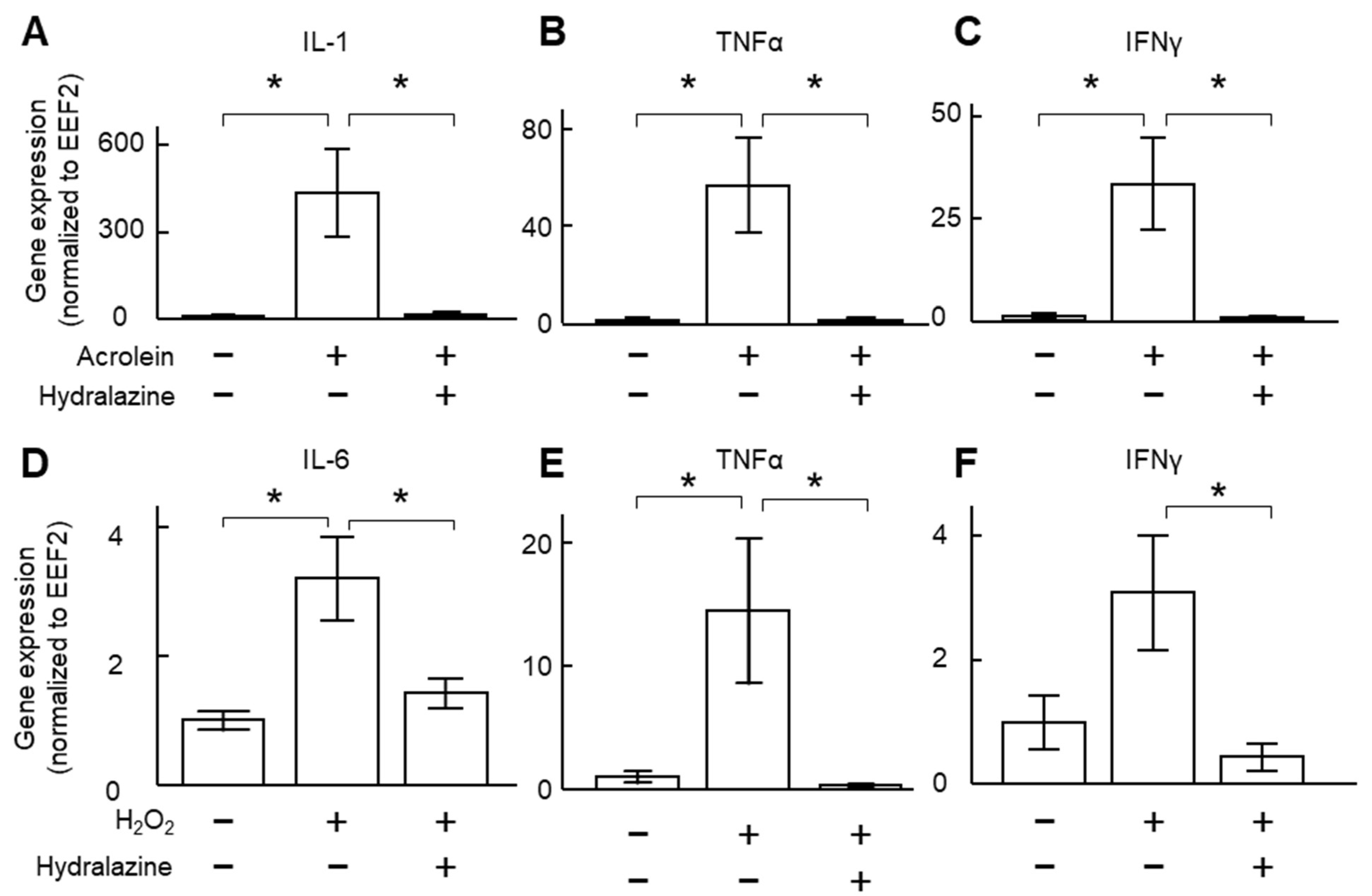
2.1. Hydralazine Decreased Inflammatory Cytokine Expression in Cultured VSMCs
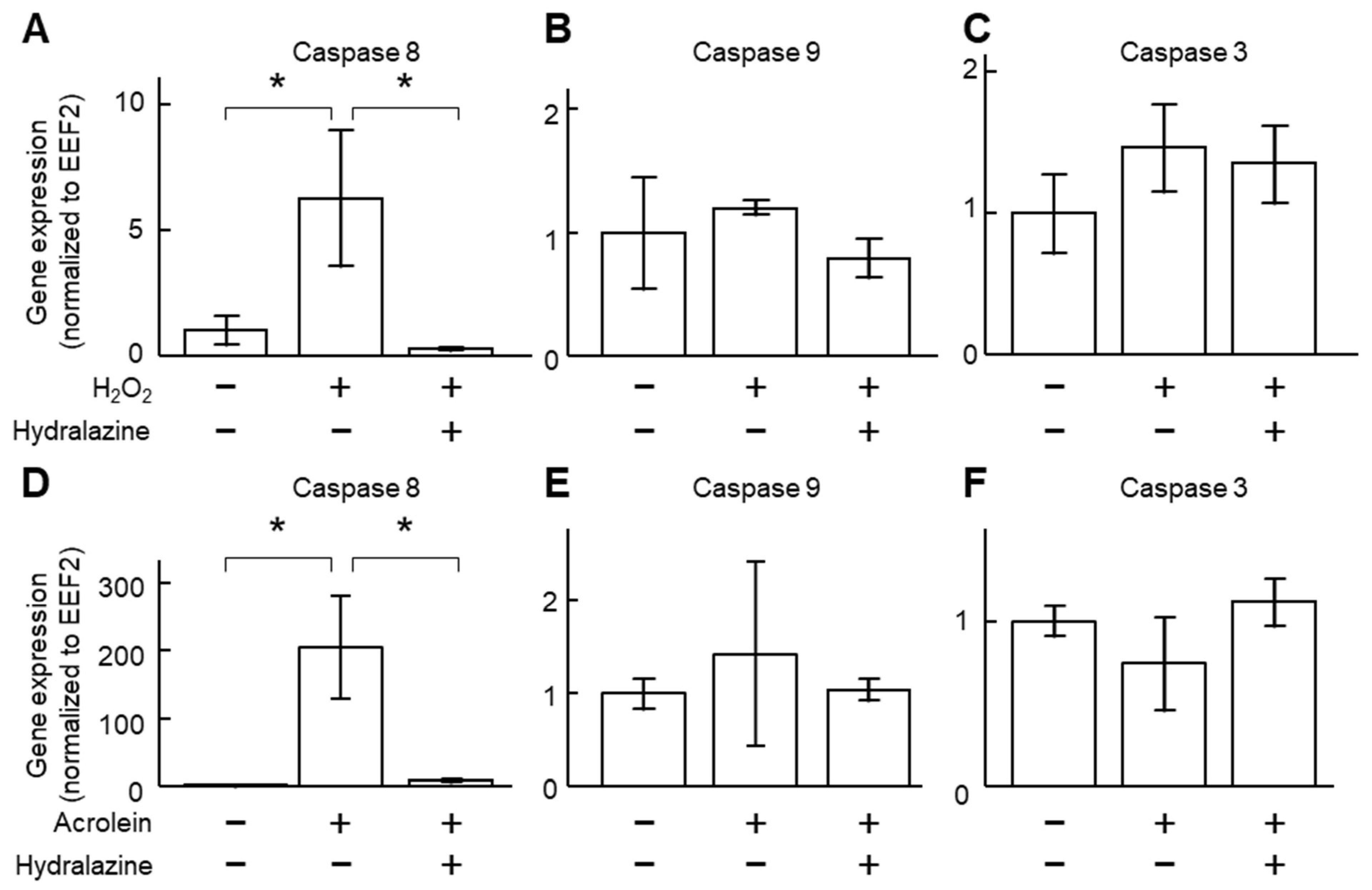
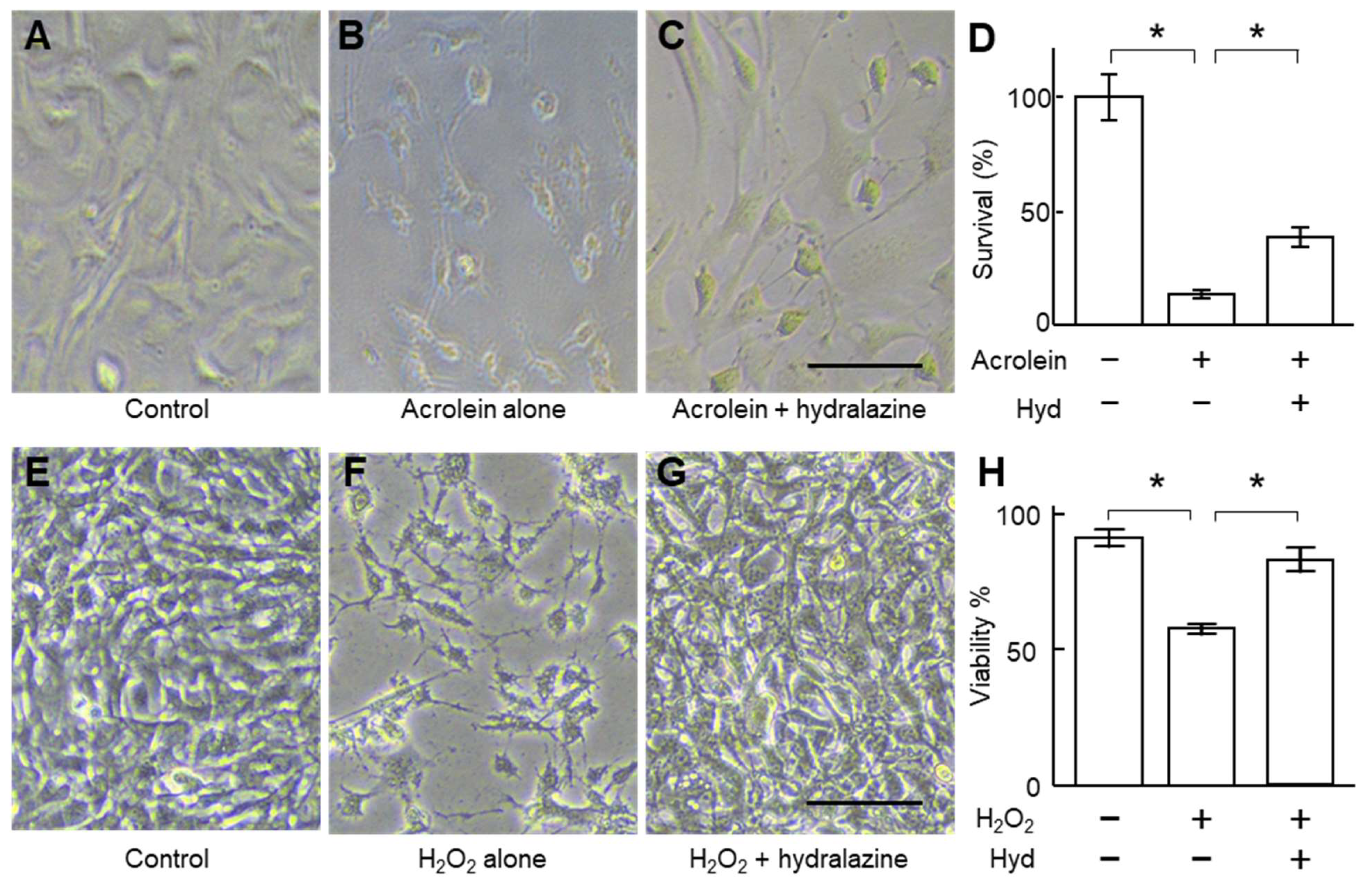
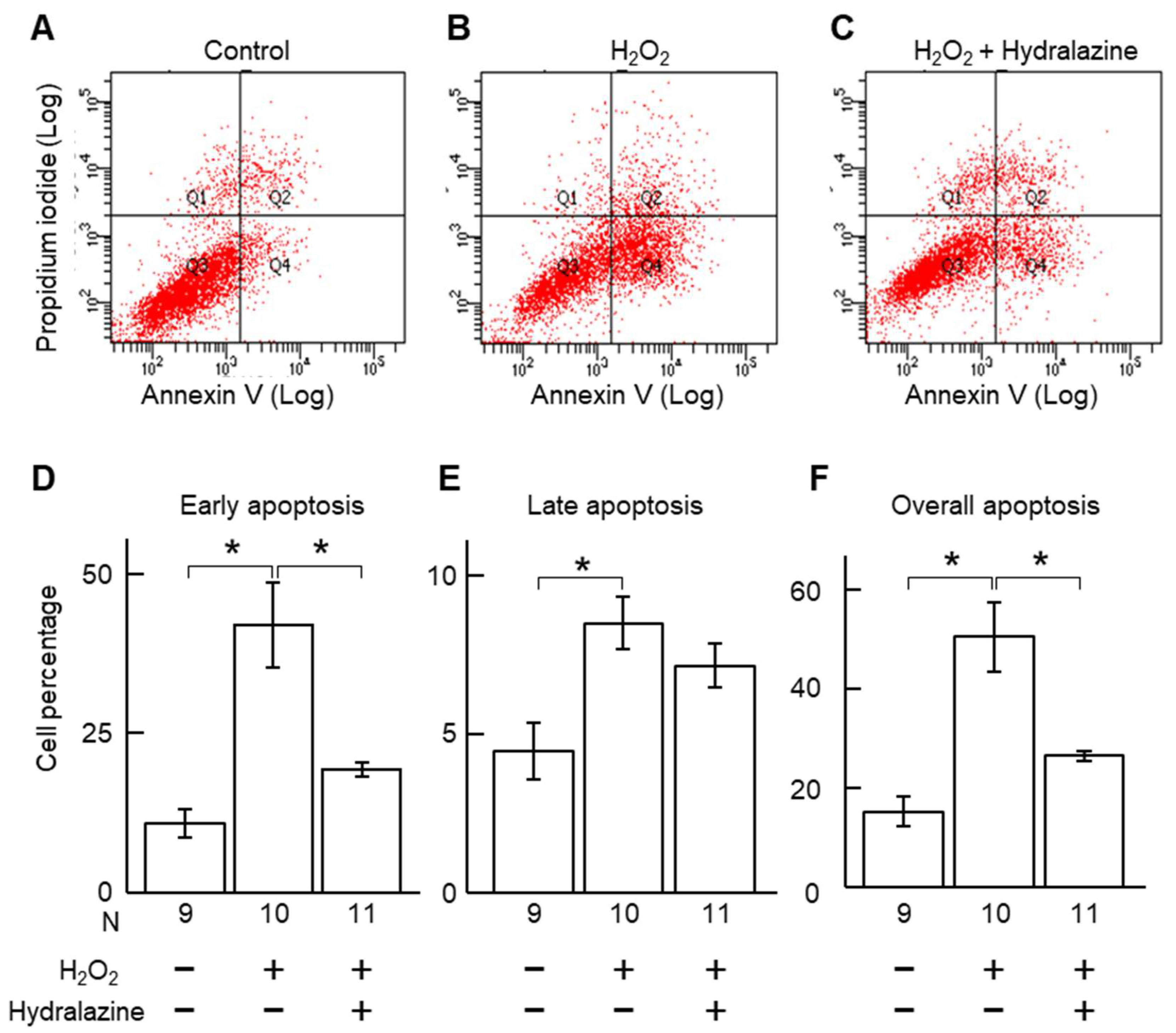
2.2. Hydralazine Decreased Acrolein- and H2O2-Induced Cell Death in Cultured VSMCs
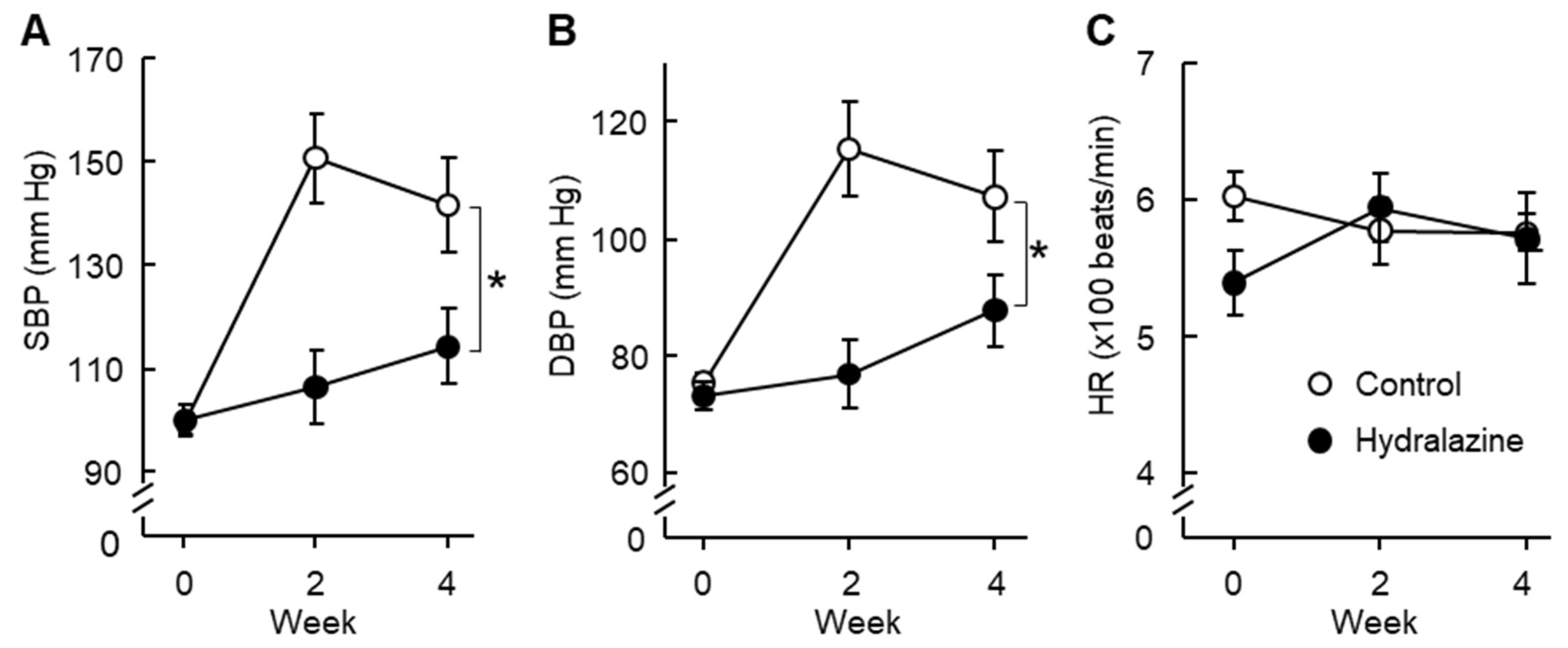
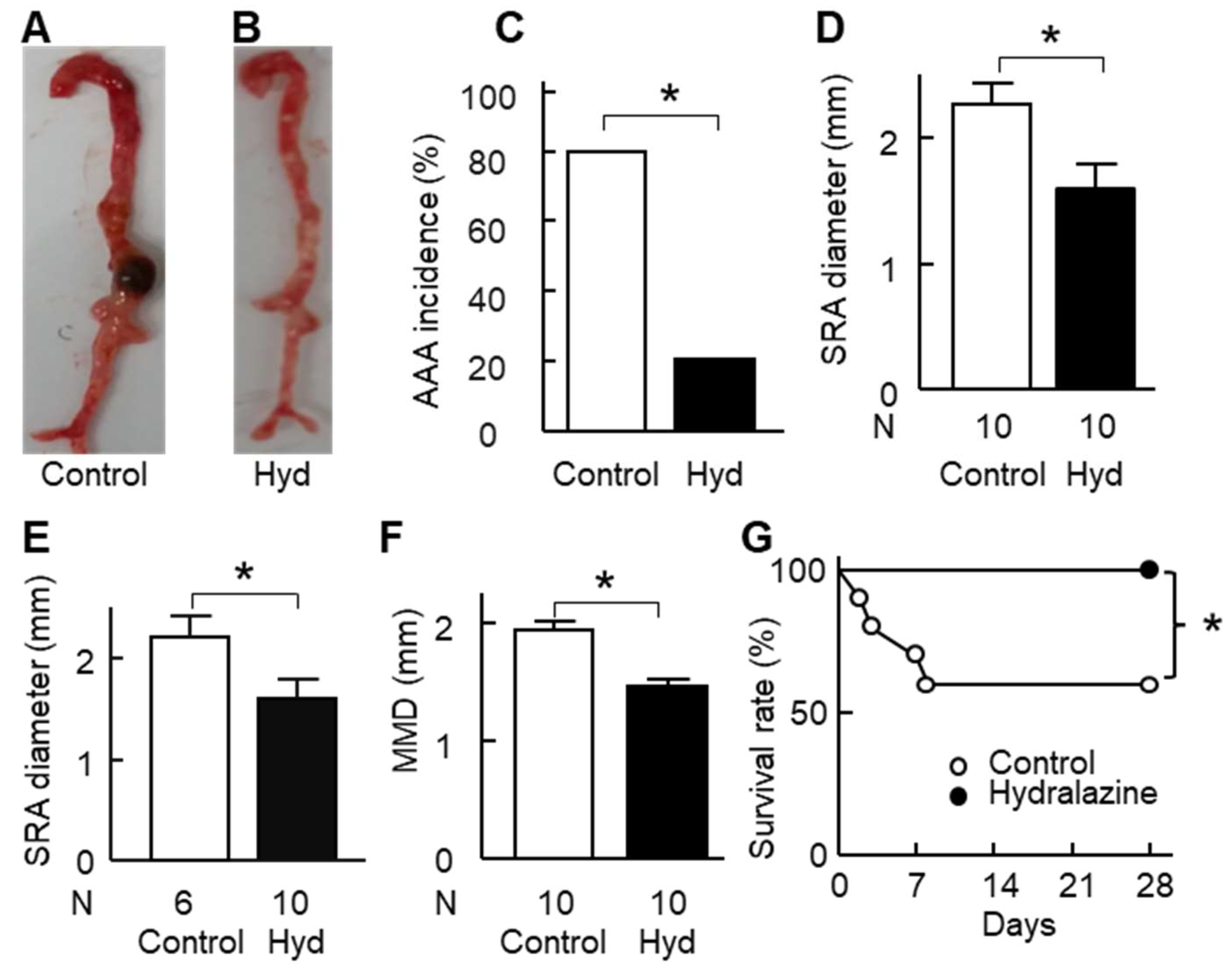
2.3. Hydralazine Treatment Protected Apolipoprotein E-Deficient (ApoE−/−) Mice against AAA
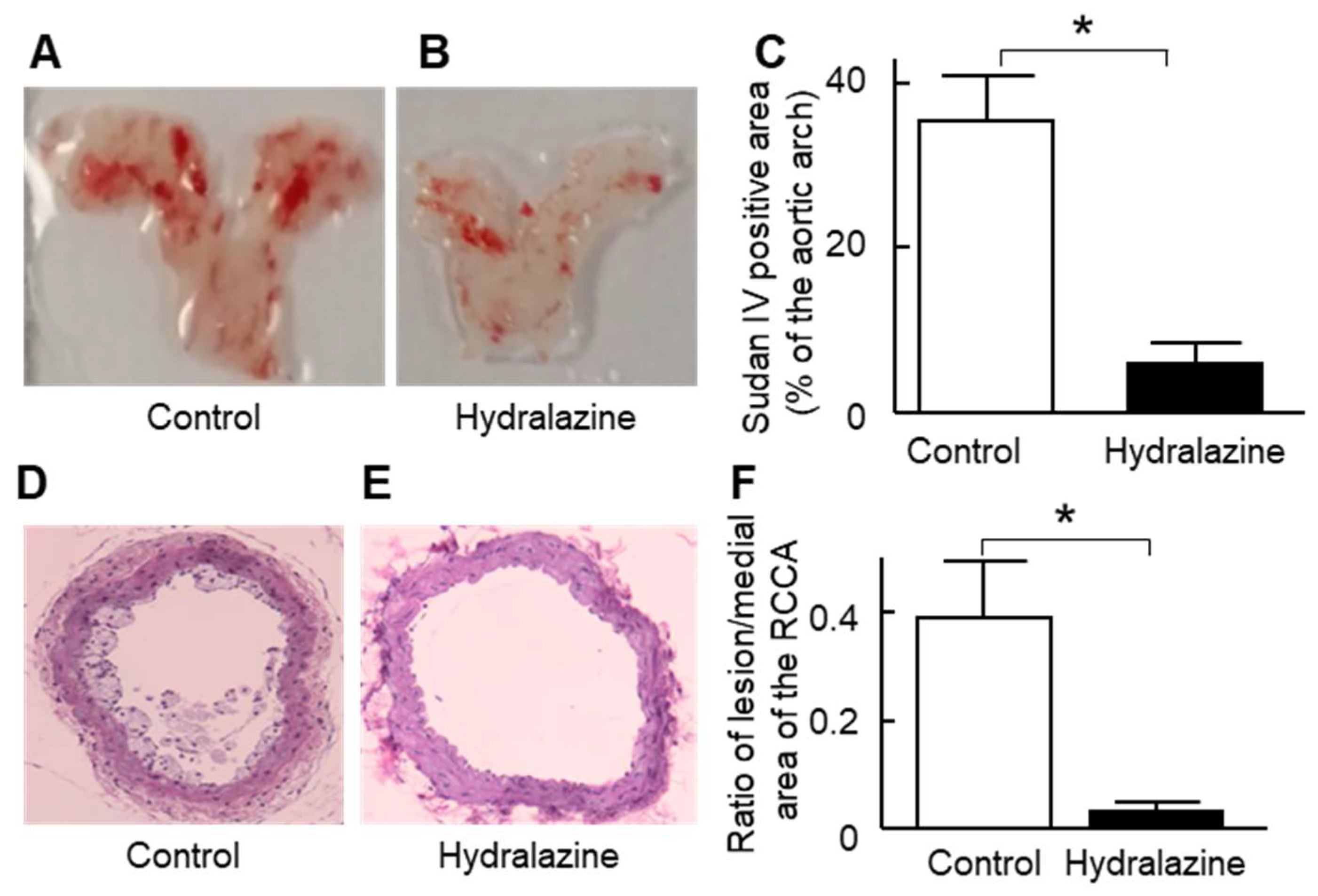
2.4. Hydralazine Treatment Protected Mice against Atherosclerosis and Cardiac Hypertrophy
3. Discussion
4. Methods
4.1. Animals
4.2. Experimental Protocol
4.3. Morphometry Measurement of Aortic Diameter
4.4. Quantification of the Atherosclerotic Lesion Area
4.5. Cell Width of Cardiomyocytes
4.6. Cell Culture
4.7. MTS Assay
4.8. Trypan Blue Assay
4.9. Gene Expression Analysis
4.10. Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. Flow Cytometry Assay
4.12. Terminal Deoxynucleotidyl Transferase Mediated dUTP Nick-End Labeling (TUNEL) Assay
4.13. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sampson, U.K.; Norman, P.E.; Fowkes, F.G.; Aboyans, V.; Yanna, S.; Harrell, F.E., Jr.; Forouzanfar, M.H.; Naghavi, M.; Denenberg, J.O.; McDermott, M.M.; et al. Global and regional burden of aortic dissection and aneurysms: Mortality trends in 21 world regions, 1990 to 2010. Global Heart 2014, 9, 171–180.e110. [Google Scholar] [CrossRef]
- Golledge, J.; Krishna, S.M.; Wang, Y. Mouse models for abdominal aortic aneurysm. Br. J. Pharmacol. 2022, 179, 792–810. [Google Scholar] [CrossRef] [PubMed]
- Landry, G.J.; Liem, T.K.; Abraham, C.Z.; Jung, E.; Moneta, G.L. Predictors of perioperative morbidity and mortality in open abdominal aortic aneurysm repair. Am. J. Surg. 2019, 217, 943–947. [Google Scholar] [CrossRef]
- Stather, P.W.; Sidloff, D.; Dattani, N.; Choke, E.; Bown, M.J.; Sayers, R.D. Systematic review and meta-analysis of the early and late outcomes of open and endovascular repair of abdominal aortic aneurysm. Br. J. Surg. 2013, 100, 863–872. [Google Scholar] [CrossRef]
- Brewster, D.C.; Cronenwett, J.L.; Hallett, J.W., Jr.; Johnston, K.W.; Krupski, W.C.; Matsumura, J.S. Guidelines for the treatment of abdominal aortic aneurysms. Report of a subcommittee of the Joint Council of the American Association for Vascular Surgery and Society for Vascular Surgery. J. Vasc. Surg. 2003, 37, 1106–1117. [Google Scholar] [CrossRef]
- Su, Z.; Guo, J.; Gu, Y. Pharmacotherapy in Clinical Trials for Abdominal Aortic Aneurysms: A Systematic Review and Meta-Analysis. Clin. Appl. Thromb. Hemost. 2022, 28, 10760296221120423. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.M.; Loftin, C.D.; Zhan, C.G. Development of pharmacotherapies for abdominal aortic aneurysms. Biomed. Pharmacother. 2022, 153, 113340. [Google Scholar] [CrossRef]
- Kesavan, S.K.; Bhat, S.; Golegaonkar, S.B.; Jagadeeshaprasad, M.G.; Deshmukh, A.B.; Patil, H.S.; Bhosale, S.D.; Shaikh, M.L.; Thulasiram, H.V.; Boppana, R.; et al. Proteome wide reduction in AGE modification in streptozotocin induced diabetic mice by hydralazine mediated transglycation. Sci. Rep. 2013, 3, 2941. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, F.; Azuma, K.; Ogihara, T.; Toyofuku, Y.; Otsuka, A.; Mita, T.; Hirose, T.; Tanaka, Y.; Kawamori, R.; Watada, H. Angiotensin II type 1 receptor blocker reduces monocyte adhesion to endothelial cells in spontaneously hypertensive rats. Endocr. J. 2007, 54, 605–612. [Google Scholar] [CrossRef]
- Quan, X.; Ma, T.; Guo, K.; Wang, H.; Yu, C.Y.; Qi, C.C.; Song, B.Q. Hydralazine Promotes Central Nervous System Recovery after Spinal Cord Injury by Suppressing Oxidative Stress and Inflammation through Macrophage Regulation. Curr. Med. Sci. 2023, 43, 749–758. [Google Scholar] [CrossRef]
- Chiang, C.H.; Chen, C.; Fang, S.Y.; Lin, S.C.; Chen, J.W.; Chang, T.T. Xanthine oxidase/NADPH oxidase inhibition by hydralazine attenuates acute kidney injury and prevents the transition of acute kidney injury to chronic kidney disease. Life Sci. 2023, 327, 121863. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.M.D.; Da Silva, E.A.P.; Oliveira, J.Y.S.; Marinho, Y.Y.M.; Santana, I.R.; Heimfarth, L.; Pereira, E.W.M.; Júnior, L.J.Q.; Assreuy, J.; Menezes, I.A.C.; et al. The Therapeutic Value of Hydralazine in Reducing Inflammatory Response, Oxidative Stress, and Mortality in Animal Sepsis: Involvement of the PI3K/AKT Pathway. Shock 2021, 56, 782–792. [Google Scholar] [CrossRef]
- Li, C.; Su, Z.; Ge, L.; Chen, Y.; Chen, X.; Li, Y. Cardioprotection of hydralazine against myocardial ischemia/reperfusion injury in rats. Eur. J. Pharmacol. 2020, 869, 172850. [Google Scholar] [CrossRef]
- Jia, L.; Liu, Z.; Sun, L.; Miller, S.S.; Ames, B.N.; Cotman, C.W.; Liu, J. Acrolein, a toxicant in cigarette smoke, causes oxidative damage and mitochondrial dysfunction in RPE cells: Protection by (R)-alpha-lipoic acid. Investig. Ophthalmol. Vis. Sci. 2007, 48, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Hikisz, P.; Jacenik, D. The Tobacco Smoke Component, Acrolein, as a Major Culprit in Lung Diseases and Respiratory Cancers: Molecular Mechanisms of Acrolein Cytotoxic Activity. Cells 2023, 12, 879. [Google Scholar] [CrossRef]
- Clark, J.B. Role of Hydrogen Peroxide in the Development of Abdominal Aortic Aneurysms. Master’s Thesis, Emory University, Atlanta, GA, USA, 2011. Available online: https://etd.library.emory.edu/concern/etds/cn69m4331?locale=en (accessed on 16 March 2023).
- Wang, Y.; Nguyen, D.T.; Anesi, J.; Alramahi, A.; Witting, P.K.; Chai, Z.; Khan, A.W.; Kelly, J.; Denton, K.M.; Golledge, J. Moxonidine Increases Uptake of Oxidised Low-Density Lipoprotein in Cultured Vascular Smooth Muscle Cells and Inhibits Atherosclerosis in Apolipoprotein E-Deficient Mice. Int. J. Mol. Sci. 2023, 24, 3857. [Google Scholar] [CrossRef]
- Tsuruda, T.; Sekita-Hatakeyama, Y.; Hao, Y.; Sakamoto, S.; Kurogi, S.; Nakamura, M.; Udagawa, N.; Funamoto, T.; Sekimoto, T.; Hatakeyama, K.; et al. Angiotensin II Stimulation of Cardiac Hypertrophy and Functional Decompensation in Osteoprotegerin-Deficient Mice. Hypertension 2016, 67, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, E.; Sasaki, S.; Kinoshita, H.; Kito, T.; Ohta, H.; Konishi, M.; Kuwahara, K.; Nakao, K.; Itoh, N. Angiotensin II-induced cardiac hypertrophy and fibrosis are promoted in mice lacking Fgf16. Genes Cells 2013, 18, 544–553. [Google Scholar] [CrossRef]
- Shao, D.; Gao, Z.; Zhao, Y.; Fan, M.; Zhao, X.; Wei, Q.; Pan, M.; Ma, B. Sulforaphane Suppresses H2O2-Induced Oxidative Stress and Apoptosis via the Activation of AMPK/NFE2L2 Signaling Pathway in Goat Mammary Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 1070. [Google Scholar] [CrossRef]
- Wang, H.T.; Chen, T.Y.; Weng, C.W.; Yang, C.H.; Tang, M.S. Acrolein preferentially damages nucleolus eliciting ribosomal stress and apoptosis in human cancer cells. Oncotarget 2016, 7, 80450–80464. [Google Scholar] [CrossRef]
- Kimura, N.; Shimizu, H.; Eldawoody, H.; Nakayama, T.; Saito, A.; Tominaga, T.; Takahashi, A. Effect of olmesartan and pravastatin on experimental cerebral aneurysms in rats. Brain Res. 2010, 1322, 144–152. [Google Scholar] [CrossRef]
- Tada, Y.; Wada, K.; Shimada, K.; Makino, H.; Liang, E.I.; Murakami, S.; Kudo, M.; Kitazato, K.T.; Nagahiro, S.; Hashimoto, T. Roles of Hypertension in the Rupture of Intracranial Aneurysms. Stroke 2014, 45, 579–586. [Google Scholar] [CrossRef]
- Cassis, L.A.; Gupte, M.; Thayer, S.; Zhang, X.; Charnigo, R.; Howatt, D.A.; Rateri, D.L.; Daugherty, A. ANG II infusion promotes abdominal aortic aneurysms independent of increased blood pressure in hypercholesterolemic mice. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1660–H1665. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, S.; Loftin, C.D. Cyclooxygenase-2 inhibition attenuates abdominal aortic aneurysm progression in hyperlipidemic mice. PLoS ONE 2012, 7, e44369. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.M.; Li, J.; Wang, Y.; Moran, C.S.; Trollope, A.; Huynh, P.; Jose, R.; Biros, E.; Ma, J.; Golledge, J. Kallistatin limits abdominal aortic aneurysm by attenuating generation of reactive oxygen species and apoptosis. Sci. Rep. 2021, 11, 17451. [Google Scholar] [CrossRef] [PubMed]
- Paige, E.; Clément, M.; Lareyre, F.; Sweeting, M.; Raffort, J.; Grenier, C.; Finigan, A.; Harrison, J.; Peters, J.E.; Sun, B.B.; et al. Interleukin-6 Receptor Signaling and Abdominal Aortic Aneurysm Growth Rates. Circ. Genom. Precis. Med. 2019, 12, e002413. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Wang, H.X.; Tian, C.; Ren, H.L.; Li, F.D.; Wang, C.Y.; Li, H.H.; Zheng, Y.H. Chemokine (C-X-C motif) receptor 2 blockade by SB265610 inhibited angiotensin II-induced abdominal aortic aneurysm in Apo E(-/-) mice. Heart Vessel. 2019, 34, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Sun, J.; Liang, W.; Chang, Z.; Rom, O.; Zhao, Y.; Zhao, G.; Xiong, W.; Wang, H.; Zhu, T.; et al. Cyclodextrin Prevents Abdominal Aortic Aneurysm via Activation of Vascular Smooth Muscle Cell Transcription Factor EB. Circulation 2020, 142, 483–498. [Google Scholar] [CrossRef]
- Cooper, H.A.; Cicalese, S.; Preston, K.J.; Kawai, T.; Okuno, K.; Choi, E.T.; Kasahara, S.; Uchida, H.A.; Otaka, N.; Scalia, R.; et al. Targeting mitochondrial fission as a potential therapeutic for abdominal aortic aneurysm. Cardiovasc. Res. 2021, 117, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Kobeissi, E.; Hibino, M.; Pan, H.; Aune, D. Blood pressure, hypertension and the risk of abdominal aortic aneurysms: A systematic review and meta-analysis of cohort studies. Eur. J. Epidemiol. 2019, 34, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Siordia, J.A. Beta-Blockers and Abdominal Aortic Aneurysm Growth: A Systematic Review and Meta-Analysis. Curr. Cardiol. Rev. 2021, 17, e230421187502. [Google Scholar] [CrossRef] [PubMed]
- Inanaga, K.; Ichiki, T.; Matsuura, H.; Miyazaki, R.; Hashimoto, T.; Takeda, K.; Sunagawa, K. Resveratrol attenuates angiotensin II-induced interleukin-6 expression and perivascular fibrosis. Hypertens. Res. 2009, 32, 466–471. [Google Scholar] [CrossRef]
- Kai, T.; Kino, H.; Ishikawa, K. Role of the renin-angiotensin system in cardiac hypertrophy and renal glomerular sclerosis in transgenic hypertensive mice carrying both human renin and angiotensinogen genes. Hypertens. Res. 1998, 21, 39–46. [Google Scholar] [CrossRef]
- Custodis, F.; Baumhäkel, M.; Schlimmer, N.; List, F.; Gensch, C.; Böhm, M.; Laufs, U. Heart rate reduction by ivabradine reduces oxidative stress, improves endothelial function, and prevents atherosclerosis in apolipoprotein E-deficient mice. Circulation 2008, 117, 2377–2387. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Chen, L.; Mizel, D.; Huang, Y.G.; Briggs, J.P.; Schnermann, J. Low plasma renin and reduced renin secretory responses to acute stimuli in conscious COX-2-deficient mice. Am. J. Physiol. Renal Physiol. 2007, 292, F415–F422. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.; Taylor, W.R. Deoxycorticosterone acetate salt hypertension in apolipoprotein E-/- mice results in accelerated atherosclerosis: The role of angiotensin II. Hypertension 2008, 51, 218–224. [Google Scholar] [CrossRef]
- Galvani, S.; Coatrieux, C.; Elbaz, M.; Grazide, M.H.; Thiers, J.C.; Parini, A.; Uchida, K.; Kamar, N.; Rostaing, L.; Baltas, M.; et al. Carbonyl scavenger and antiatherogenic effects of hydrazine derivatives. Free Radic. Biol. Med. 2008, 45, 1457–1467. [Google Scholar] [CrossRef]
- Alexander, R.W. Hypertension and the Pathogenesis of Atherosclerosis. Hypertension 1995, 25, 155–161. [Google Scholar] [CrossRef]
- Kosmas, C.E.; Silverio, D.; Sourlas, A.; Montan, P.D.; Guzman, E.; Garcia, M.J. Anti-inflammatory therapy for cardiovascular disease. Ann. Transl. Med. 2019, 7, 147. [Google Scholar] [CrossRef] [PubMed]
- Arnett Donna, K.; Blumenthal Roger, S.; Albert Michelle, A.; Buroker Andrew, B.; Goldberger Zachary, D.; Hahn Ellen, J.; Himmelfarb Cheryl, D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA Guideline on the Primary Prevention of Cardiovascular Disease: Executive Summary. J. Am. Coll. Cardiol. 2019, 74, 1376–1414. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Cheng, X.W.; Hu, L.; Hao, C.N.; Hayashi, M.; Takeshita, K.; Hamrah, M.S.; Shi, G.P.; Kuzuya, M.; Murohara, T. Renin inhibition reduces atherosclerotic plaque neovessel formation and regresses advanced atherosclerotic plaques. Atherosclerosis 2014, 237, 739–747. [Google Scholar] [CrossRef]
- Noda, K.; Hosoya, M.; Nakajima, S.; Ohashi, J.; Fukumoto, Y.; Shimokawa, H. Anti-atherogenic effects of the combination therapy with olmesartan and azelnidipine in diabetic apolipoprotein E-deficient mice. Tohoku J. Exp. Med. 2012, 228, 305–315. [Google Scholar] [CrossRef]
- Vial, J.H.; Yong, A.C.; Boyd, G.W. Structural change in the rat hindlimb during deoxycorticosterone acetate hypertension; its reversibility and prevention. J. Hypertens. 1989, 7, 143–150. [Google Scholar] [CrossRef]
- Aronow, W.S. Hypertension and left ventricular hypertrophy. Ann. Transl. Med. 2017, 5, 310. [Google Scholar] [CrossRef]
- Dahlöf, B.; Pennert, K.; Hansson, L. Reversal of left ventricular hypertrophy in hypertensive patients. A metaanalysis of 109 treatment studies. Am. J. Hypertens. 1992, 5, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Smeets, P.J.; Teunissen, B.E.; Planavila, A.; de Vogel-van den Bosch, H.; Willemsen, P.H.; van der Vusse, G.J.; van Bilsen, M. Inflammatory pathways are activated during cardiomyocyte hypertrophy and attenuated by peroxisome proliferator-activated receptors PPARalpha and PPARdelta. J. Biol. Chem. 2008, 283, 29109–29118. [Google Scholar] [CrossRef] [PubMed]
- Loufrani, L.; Henrion, D. Vasodilator treatment with hydralazine increases blood flow in mdx mice resistance arteries without vascular wall remodelling or endothelium function improvement. J. Hypertens. 2005, 23, 1855–1860. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Krishna, S.M.; Moxon, J.; Dinh, T.N.; Jose, R.J.; Yu, H.; Golledge, J. Influence of apolipoprotein E, age and aortic site on calcium phosphate induced abdominal aortic aneurysm in mice. Atherosclerosis 2014, 235, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nguyen, D.T.; Yang, G.; Anesi, J.; Chai, Z.; Charchar, F.; Golledge, J. An Improved 3-(4,5-Dimethylthiazol-2-yl)-5-(3-Carboxymethoxyphenyl)-2-(4-Sulfophenyl)-2H-Tetrazolium Proliferation Assay to Overcome the Interference of Hydralazine. Assay. Drug Dev. Technol. 2020, 18, 379–384. [Google Scholar] [CrossRef]
- Haefliger, I.; Pedrini, M.; Anderson, D.R. Relaxing effect of CEDO 8956 and hydralazine HCl in cultured smooth muscle cells versus pericytes: A preliminary study. Klin. Monbl. Augenheilkd. 2002, 219, 277–280. [Google Scholar] [CrossRef]
- Burcham, P.C.; Raso, A.; Kaminskas, L.M. Chaperone heat shock protein 90 mobilization and hydralazine cytoprotection against acrolein-induced carbonyl stress. Mol. Pharmacol. 2012, 82, 876–886. [Google Scholar] [CrossRef]
- Wang, Y.; Nguyen, D.T.; Yang, G.; Anesi, J.; Kelly, J.; Chai, Z.; Ahmady, F.; Charchar, F.; Golledge, J. A Modified MTS Proliferation Assay for Suspended Cells to Avoid the Interference by Hydralazine and β-Mercaptoethanol. Assay. Drug Dev. Technol. 2021, 19, 184–190. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dinh, T.N.; Nield, A.; Krishna, S.M.; Denton, K.; Golledge, J. Renal Denervation Promotes Atherosclerosis in Hypertensive Apolipoprotein E-Deficient Mice Infused with Angiotensin II. Front. Physiol. 2017, 8, 215. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, I.; Batra, S.K. Protocol for Apoptosis Assay by Flow Cytometry Using Annexin V Staining Method. Bio-Protocol 2013, 3, e374. [Google Scholar] [CrossRef]
- Kuystermans, D.; Avesh, M.; Al-Rubeai, M. Online flow cytometry for monitoring apoptosis in mammalian cell cultures as an application for process analytical technology. Cytotechnology 2016, 68, 399–408. [Google Scholar] [CrossRef]
- Liu, Y.; Nadeem, A.; Sebastian, S.; Olsson, M.A.; Wai, S.N.; Styring, E.; Engellau, J.; Isaksson, H.; Tägil, M.; Lidgren, L.; et al. Bone mineral: A trojan horse for bone cancers. Efficient mitochondria targeted delivery and tumor eradication with nano hydroxyapatite containing doxorubicin. Mater. Today Bio 2022, 14, 100227. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Definition, prevalence, and risk factors of low sex hormone-binding globulin in US adults. J. Clin. Endocrinol. Metab. 2021, 106, e3946–e3956. [Google Scholar] [CrossRef]
- Qian, T.; Sun, H.; Xu, Q.; Hou, X.; Hu, W.; Zhang, G.; Drummond, G.R.; Sobey, C.G.; Charchar, F.J.; Golledge, J.; et al. Hyperuricemia is independently associated with hypertension in men under 60 years in a general Chinese population. J. Hum. Hypertens. 2021, 35, 1020–1028. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Sargisson, O.; Nguyen, D.T.; Parker, K.; Pyke, S.J.R.; Alramahi, A.; Thihlum, L.; Fang, Y.; Wallace, M.E.; Berzins, S.P.; et al. Effect of Hydralazine on Angiotensin II-Induced Abdominal Aortic Aneurysm in Apolipoprotein E-Deficient Mice. Int. J. Mol. Sci. 2023, 24, 15955. https://doi.org/10.3390/ijms242115955
Wang Y, Sargisson O, Nguyen DT, Parker K, Pyke SJR, Alramahi A, Thihlum L, Fang Y, Wallace ME, Berzins SP, et al. Effect of Hydralazine on Angiotensin II-Induced Abdominal Aortic Aneurysm in Apolipoprotein E-Deficient Mice. International Journal of Molecular Sciences. 2023; 24(21):15955. https://doi.org/10.3390/ijms242115955
Chicago/Turabian StyleWang, Yutang, Owen Sargisson, Dinh Tam Nguyen, Ketura Parker, Stephan J. R. Pyke, Ahmed Alramahi, Liam Thihlum, Yan Fang, Morgan E. Wallace, Stuart P. Berzins, and et al. 2023. "Effect of Hydralazine on Angiotensin II-Induced Abdominal Aortic Aneurysm in Apolipoprotein E-Deficient Mice" International Journal of Molecular Sciences 24, no. 21: 15955. https://doi.org/10.3390/ijms242115955
APA StyleWang, Y., Sargisson, O., Nguyen, D. T., Parker, K., Pyke, S. J. R., Alramahi, A., Thihlum, L., Fang, Y., Wallace, M. E., Berzins, S. P., Oqueli, E., Magliano, D. J., & Golledge, J. (2023). Effect of Hydralazine on Angiotensin II-Induced Abdominal Aortic Aneurysm in Apolipoprotein E-Deficient Mice. International Journal of Molecular Sciences, 24(21), 15955. https://doi.org/10.3390/ijms242115955