
Carnosine, Zinc and Copper: A Menage a Trois in Bone and Cartilage Protection



Abstract
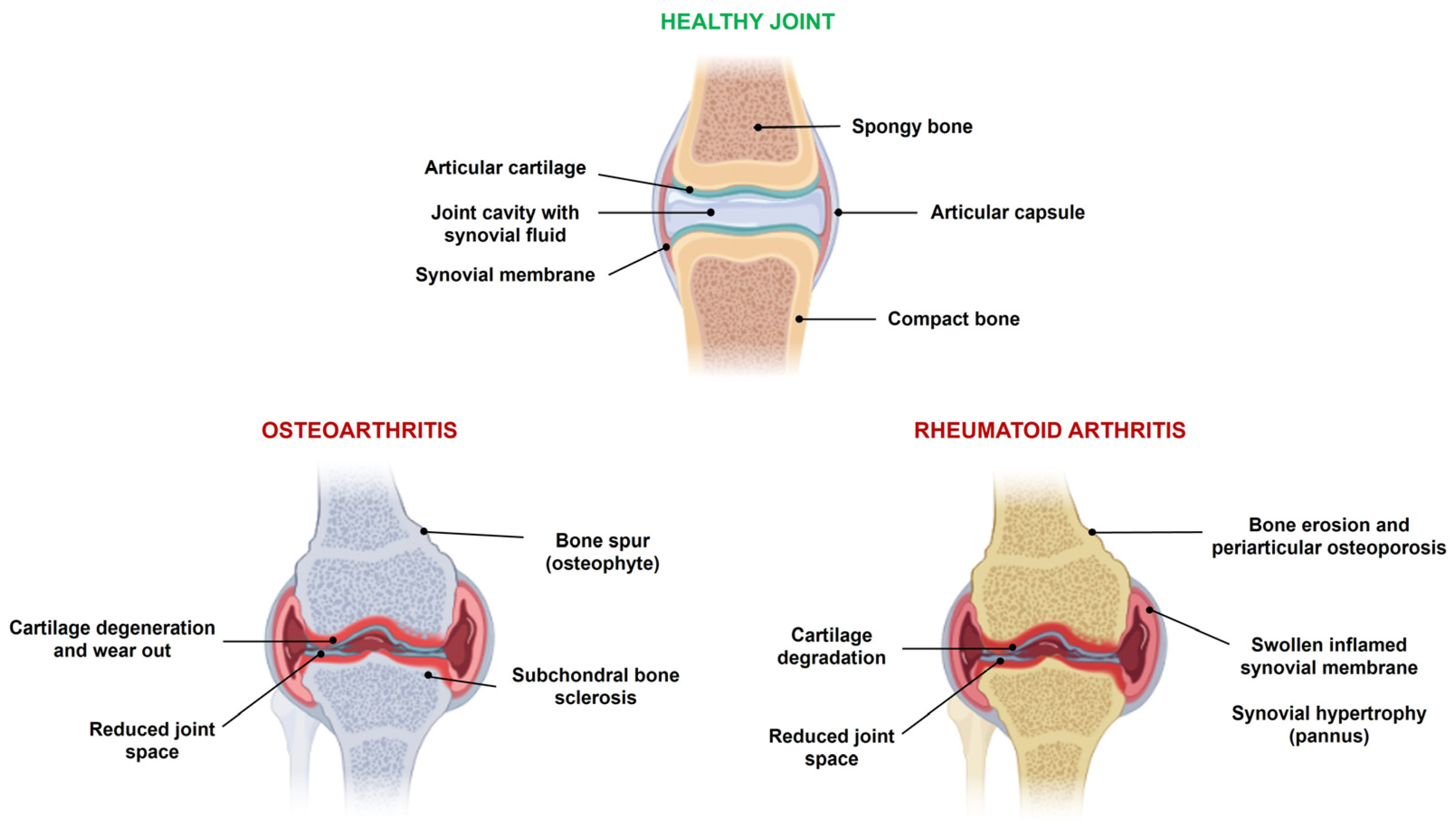
:1. Introduction
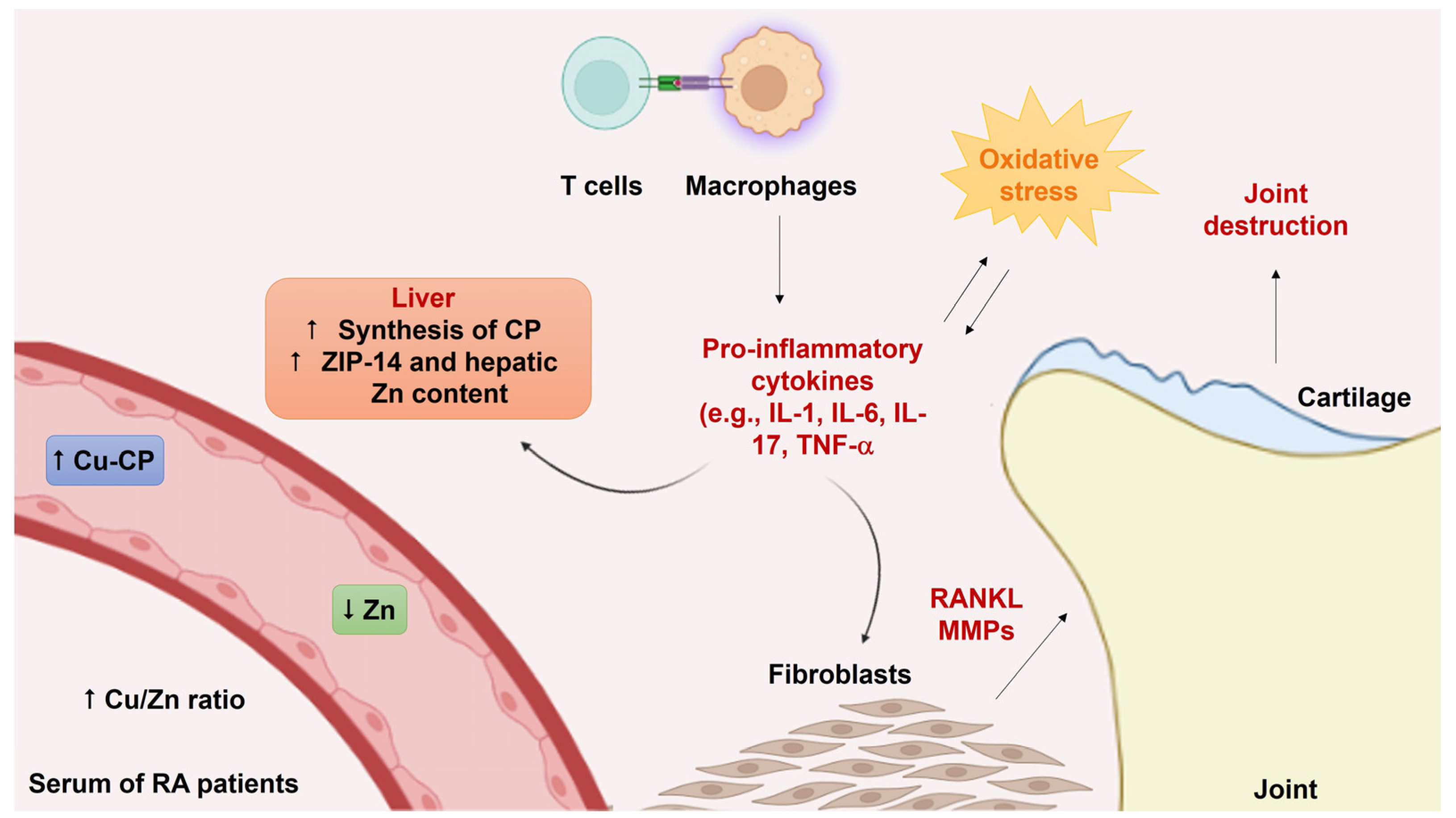
2. Copper and Zinc Dyshomeostasis in Arthritis
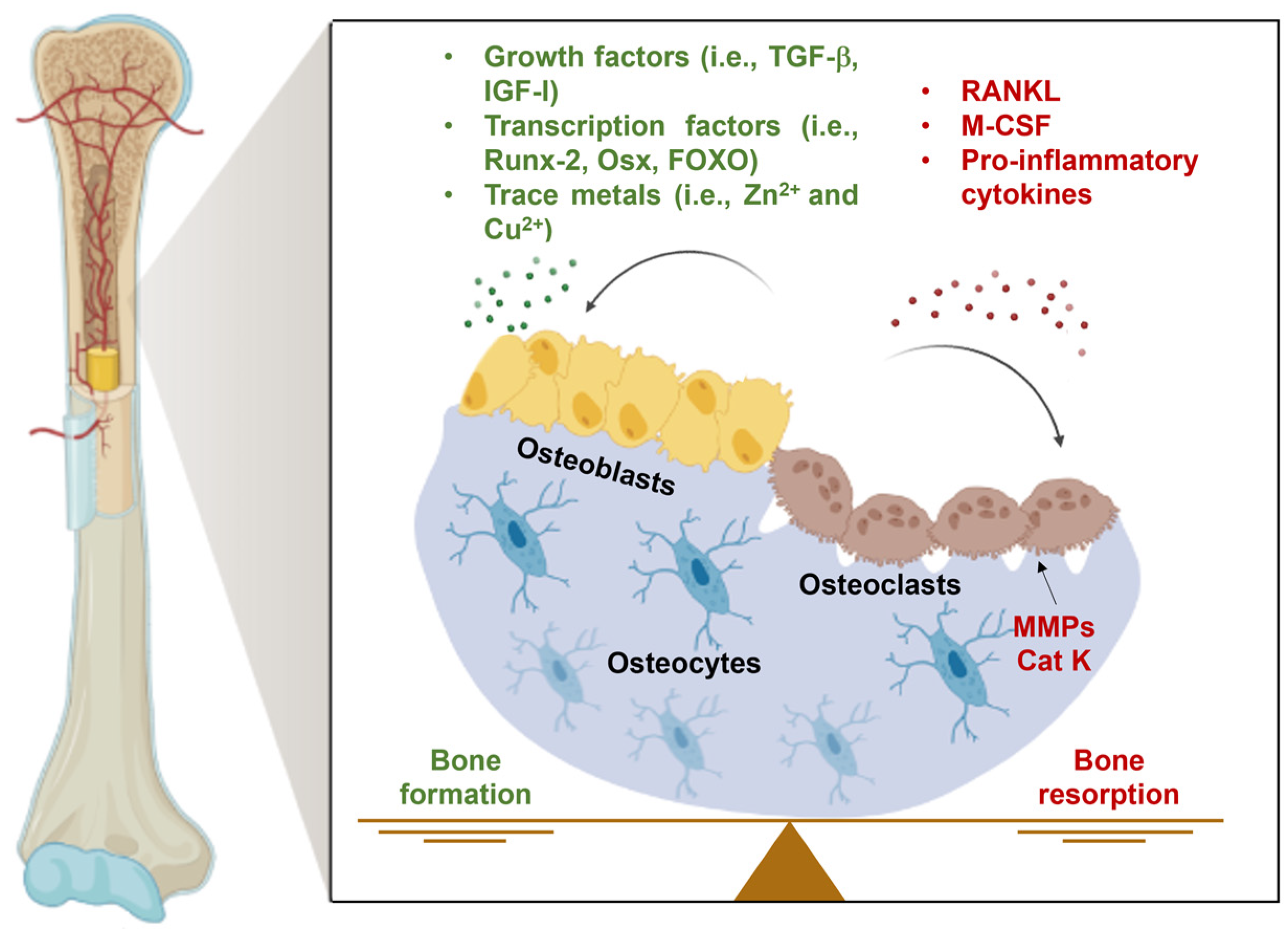
3. The Bioinorganic Chemistry of Zinc and Copper in Bone and Cartilage Tissues
3.1. Zinc
Effects of Zinc in Bone and Cartilage Tissues

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zn Source | Cell Type | Animal Model | Effects | Outcomes | Ref. |
|---|---|---|---|---|---|
| Zinc chloride | MC3T3-E1 | - | ↑ cell proliferation, ALP activity, collagen synthesis | ↑ bone formation | [119] |
| Zinc carbonate | - | diabetes-induced bone loss in rats | ↑ OCN expression, ALP activity | ↓ bone loss | [120] |
| Zinc acexamate | - | rats with fracture healing | ↑ IGF-1 and TGF-β | ↑ fracture healing | [121] |
| Zinc sulfate | MC3T3-E1 | - | ↑ Runx2, OPG, regucalcin mRNA | ↑ osteoblast differentiation | [125] |
| Zinc chloride | MC3T3-E1 | - | ↑ Runx2 expression via BMP-2 signaling | ↑ osteoblast differentiation | [126] |
| Zinc sulfate | ADSCs | - | ↑ β-catenin, Wnt1, Wnt3a, LRP5 and DKK1 mRNA ↑ ALP and PKA activity ↑ Ca2+ levels | ↑ osteoblast differentiation | [129] |
| Zinc sulfate | RAW264.7 | - | ↓ NF-KB activation | ↓ osteoclastogenesis | [133] |
| Zinc sulfate | mBMMs, RAW264.7 | - | ↓ NFATc1 signaling pathway | ↓ osteoclastogenesis | [134] |
| Zinc sulfate | - | ovariectomized and type 1 diabetic rats | ↓ RANKL/OPG ratio | ↓ bone alterations | [137] |
| Zn-containing tricalcium phosphate | osteoclasts isolated from rabbits | - | ↓ actin ring formation | ↑ osteoclast apoptosis | [138] |
| Zn-carbon dots | BMSCs | calvarial bone defect model in rats | ↑ ALP activity, area of calcified nodules | ↑ bone regeneration | [139] |
| Zn-PCL/gelatin nanofiber | MG-63 | - | ↑ ALP activity, Ca2+ content, ↑ Runx2 and COL1 expression | ↑ bone regeneration | [140] |
| Zn-implanted titanium | MC3T3-E1 | - | ↑ ALP activity, collagen secretion, mineralization, antibacterial activity towards E. coli and S. aureus | osteogenic, antibacterial effects | [141] |
| Zn-doped hydroxyapatite/ graphene nanocomposite | MSCs | - | ↑ cell proliferation, ALP activity, antibacterial function against E. coli and S. aureus | osteogenic, antibacterial effects | [142] |
| Zn silicate/nanohydroxyapatite/ collagen scaffolds | BMSCs | calvarial defect model in rats | ↑ BMP-2, Osx, VEGF-A, Cd31 expression | ↑ osteogenesis, angiogenesis | [143] |
| Zinc chloride | ATDC5 | - | ↑ VEGF-A, VEGF-B, SOX9, Runx1, collagen expression | ↑ chondrogenesis | [146] |
| Zinc sulfate | SW1353 | MIA-induced OA in rats | ↓ pro-inflammatory cytokines ↑ PI3K/Akt/Nrf-2 pathway | protective effects | [147] |
3.2. Copper
Effects of Copper in Bone and Cartilage Tissues
4. Beneficial Effects of Carnosine and Its Zinc Complex, Polaprezinc, on Bone and Cartilage
| Compound | Experimental Model | Effects | Outcomes | Ref |
|---|---|---|---|---|
| Car (10−5–10−4 M) | PDLSCs | ↑ Runx2/Cbfa1 and SOX9 mRNA ↑ BMP-2 and BMP-7 expression | ↑ osteoblast differentiation | [193] |
| Car (10−4 M, 0.1/0.3/0.9 g/kg/day for 8 weeks) | FLSs, diabetes-induced OA in rats | ↓ ROS, MMP-3 and MMP-13 expression ↓ NF-KB translocation | chondroprotective effects | [197] |
| Car (10−4 M, 0.5/1.0 g/kg/day for 12 weeks) | FLSs, OA model in rats | ↑ Nrf-2/HO-1 signaling pathway ↓ MMP-3 and MMP-13 mRNA expression | protection towards the synovium, ↓ pain | [13] |
| Car (10−5 M, 150 mg/kg for 28 days) | rat primary chondrocytes, rat adjuvant arthritis, model of carrageenan induced hind paw edema | ↓ IL-1α, markers of oxidative stress, intracellular oxidant levels | anti-inflammatory, antioxidant activity | [198] |
| Polaprezinc (10−7–10−5 M) | MC3T3-E1 | ↑ ALP activity and protein synthesis | ↑ osteoblast differentiation | [209] |
| Polaprezinc (10−5 M) | MC3T3-E1 | ↑ Runx2 and regucalcin mRNA expression | ↑ osteoblast differentiation | [125] |
| Polaprezinc (10−5–10−4 M) | HPDL | ↑ Runx2/Cbfa1 and SOX9, BMP-2, BMP-7 expression | ↑ osteoblast differentiation | [210] |
| Polaprezinc (10−5–10−4 M) | C2C12 | ↑ Cbfa1/Runx2, ALP, SOX9 and type X collagen mRNA expression | ↑ osteoblast and chondrocyte differentiation | [211] |
| Polaprezinc (2.75 mg Zn/kg/day) | weanling rats | ↑ Zn, DNA, calcium contents, and ALP activity | ↑ bone metabolism | [212] |
| Polaprezinc (10−5 M) | femoral diaphysis from rats | ↑ ALP activity | ↑ bone metabolism | [213] |
| Polaprezinc (10−7–10−5 M) | mouse marrow cells | ↓ osteoclast-like cell formation | ↓ bone resorption | [214] |
| Polaprezinc (10−6 M) | mouse marrow cells | ↓ PTH stimulatory effects | ↓ PTH-stimulated osteoclastogenesis | [215] |
| Polaprezinc (10−7–10−5 M) | mouse marrow cells | ↓ TGF-β stimulatory effects | ↓ osteoclastogenesis | [216] |
| Polaprezinc (5 × 10−5 M) | hBMSCs | ↑ ALP activity, ↑ Runx2, ALP, COL1A1, SPP1, IBSP, BGLAP mRNA levels | ↑ osteogenic differentiation | [17] |
| Polaprezinc (5 × 10−5 M) | mBMMs | ↑ YAP activity, ↑ NFATc1, Cat K, Dcstamp mRNA levels | ↑ osteoclastogenic differentiation | [17] |
| Polaprezinc (25 mg/kg) | mouse femoral fracture model | ↑ active bone homeostasis | ↑ bone remodeling | [17] |
| Polaprezinc (PG/0.2–0.8%PZ) | MC3T3-E1, cranial bone defects of osteoporotic rats | ↑ Nrf-2/HO-1/SOD1 pathway ↑ ALP activity ↑ osteogenesis | antioxidant effects ↑ osteogenesis ↑ bone regeneration | [218] |
5. Hypothesis on a Potential Exchange of Zinc for Copper by Polarezinc
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rabiei, M.; Kashanian, S.; Samavati, S.S.; Derakhshankhah, H.; Jamasb, S.; McInnes, S.J. Nanotechnology application in drug delivery to osteoarthritis (OA), rheumatoid arthritis (RA), and osteoporosis (OSP). J. Drug Deliv. Sci. Technol. 2020, 61, 102011. [Google Scholar] [CrossRef]
- Fan, X.-X.; Xu, M.-Z.; Leung, E.L.-H.; Jun, C.; Yuan, Z.; Liu, L. ROS-responsive berberine polymeric micelles effectively suppressed the inflammation of rheumatoid arthritis by targeting mitochondria. Nano-Micro Lett. 2020, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Holyoak, D.T.; Tian, Y.F.; van der Meulen, M.C.H.; Singh, A. Osteoarthritis: Pathology, mouse models, and nanoparticle injectable systems for targeted treatment. Ann. Biomed. Eng. 2016, 44, 2062–2075. [Google Scholar] [CrossRef] [PubMed]
- Dalisson, B.; Barralet, J. Bioinorganics and wound healing. Adv. Health Mater. 2019, 8, e1900764. [Google Scholar] [CrossRef] [PubMed]
- Ciosek, Ż.; Kot, K.; Rotter, I. Iron, zinc, copper, cadmium, mercury, and bone tissue. Int. J. Environ. Res. Public Health 2023, 20, 2197. [Google Scholar] [CrossRef]
- Héraud, F.; Savineau, C.; Harmand, M.-F. Copper modulation of extracellular matrix synthesis by human articular chondrocytes. Scand. J. Rheumatol. 2002, 31, 279–284. [Google Scholar] [CrossRef]
- Innella, K.; Levidy, M.F.; Kadkoy, Y.; Lin, A.; Selles, M.; Sanchez, A.; Weiner, A.; Greendyk, J.; Moriarty, B.; Lauritsen, K.; et al. Local zinc treatment enhances fracture callus properties in diabetic rats. J. Orthop Res. 2023, 41, 1494–1504. [Google Scholar] [CrossRef]
- Gaffney-Stomberg, E. The impact of trace minerals on bone metabolism. Biol. Trace Elem. Res. 2018, 188, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Hou, X.; Xing, L.; Tian, F. Carnosine and bone (Review). Mol. Med. Rep. 2022, 27, 12. [Google Scholar] [CrossRef]
- Frangos, T.; Maret, W. Zinc and cadmium in the aetiology and pathogenesis of osteoarthritis and rheumatoid arthritis. Nutrients 2020, 13, 53. [Google Scholar] [CrossRef]
- Xin, L.; Yang, X.; Cai, G.; Fan, D.; Xia, Q.; Liu, L.; Hu, Y.; Ding, N.; Xu, S.; Wang, L.; et al. Serum levels of copper and zinc in patients with rheumatoid arthritis: A Meta-analysis. Biol. Trace Elem. Res. 2015, 168, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Alabarse, P.V.G.; Silva, J.M.S.; Santo, R.C.E.; Oliveira, M.S.; Almeida, A.S.; de Oliveira, M.S.; Immig, M.L.; Freitas, E.C.; Teixeira, V.O.N.; Bathurst, C.L.; et al. Metabolomic biomarker candidates for skeletal muscle loss in the collagen-induced arthritis (CIA) model. J. Pers. Med. 2021, 11, 837. [Google Scholar] [CrossRef] [PubMed]
- Busa, P.; Lee, S.-O.; Huang, N.; Kuthati, Y.; Wong, C.-S. Carnosine alleviates knee osteoarthritis and promotes synoviocyte protection via activating the Nrf2/HO-1 signaling pathway: An in-vivo and in-vitro study. Antioxidants 2022, 11, 1209. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Siracusa, R.; Cordaro, M.; Peritore, A.F.; Gugliandolo, E.; D’amico, R.; Fusco, R.; Crupi, R.; Rizzarelli, E.; Cuzzocrea, S.; et al. Protective effect of a new hyaluronic acid -carnosine conjugate on the modulation of the inflammatory response in mice subjected to collagen-induced arthritis. Biomed. Pharmacother. 2020, 125, 110023. [Google Scholar] [CrossRef]
- Odawara, S.; Doi, H.; Shikata, T.; Kitajima, K.; Suzuki, H.; Niwa, Y.; Kosaka, K.; Tarutani, K.; Tsujimura, T.; Kamikonya, N.; et al. Polaprezinc protects normal intestinal epithelium against exposure to ionizing radiation in mice. Mol. Clin. Oncol. 2016, 5, 377–381. [Google Scholar] [CrossRef]
- Ooi, T.C.; Chan, K.M.; Sharif, R. Zinc Carnosine inhibits lipopolysaccharide-induced inflammatory mediators by suppressing NF-κb activation in Raw 264.7 macrophages, independent of the MAPKs signaling pathway. Biol. Trace Elem. Res. 2016, 172, 458–464. [Google Scholar] [CrossRef]
- Ko, E.A.; Park, Y.J.; Yoon, D.S.; Lee, K.-M.; Kim, J.; Jung, S.; Lee, J.W.; Park, K.H. Drug repositioning of polaprezinc for bone fracture healing. Commun. Biol. 2022, 5, 462. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Ozaki, K. A new zinc compound, β-alanyl-l-histidinato zinc, stimulates bone growth in weanling rats. Res. Exp. Med. 1990, 190, 105–110. [Google Scholar] [CrossRef]
- Bellia, F.; Lanza, V.; Naletova, I.; Tomasello, B.; Ciaffaglione, V.; Greco, V.; Sciuto, S.; Amico, P.; Inturri, R.; Vaccaro, S.; et al. Copper(II) complexes with carnosine conjugates of hyaluronic acids at different dipeptide loading percentages behave as multiple SOD mimics and stimulate Nrf2 translocation and antioxidant response in in vitro inflammatory model. Antioxidants 2023, 12, 1632. [Google Scholar] [CrossRef]
- Sorenson, J.R.J.; Hangarter, W. Treatment of rheumatoid and degenerative diseases with copper complexes. Inflammation 1977, 2, 217–238. [Google Scholar] [CrossRef]
- Scuderi, P. Differential effects of copper and zinc on human peripheral blood monocyte cytokine secretion. Cell. Immunol. 1990, 126, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Vinci, C.; Caltabiano, V.; Santoro, A.M.; Rabuazzo, A.M.; Buscema, M.; Purrello, R.; Rizzarelli, E.; Vigneri, R.; Purrello, F. Copper addition prevents the inhibitory effects of interleukin 1-β on rat pancreatic islets. Diabetologia 1995, 38, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Pasqualicchio, M.; Gasperini, R.; Velo, G.P.; Davies, M.E. Effects of copper and zinc on proteoglycan metabolism in articular cartilage. Mediat. Inflamm. 1996, 5, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Katz, J.N.; Arant, K.R.; Loeser, R.F. Diagnosis and treatment of hip and knee osteoarthritis: A Review. JAMA 2021, 325, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: Pathogenic signaling pathways and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.-P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2010, 7, 33–42. [Google Scholar] [CrossRef]
- Molnar, V.; Matišić, V.; Kodvanj, I.; Bjelica, R.; Jeleč, Ž.; Hudetz, D.; Rod, E.; Čukelj, F.; Vrdoljak, T.; Vidović, D.; et al. Cytokines and chemokines involved in osteoarthritis pathogenesis. Int. J. Mol. Sci. 2021, 22, 9208. [Google Scholar] [CrossRef]
- Roman-Blas, J.; Jimenez, S. NF-κB as a potential therapeutic target in osteoarthritis and rheumatoid arthritis. Osteoarthr. Cartil. 2006, 14, 839–848. [Google Scholar] [CrossRef]
- Hu, Q.; Ecker, M. Overview of MMP-13 as a promising target for the treatment of osteoarthritis. Int. J. Mol. Sci. 2021, 22, 1742. [Google Scholar] [CrossRef]
- Li, T.; Peng, J.; Li, Q.; Shu, Y.; Zhu, P.; Hao, L. The mechanism and role of ADAMTS protein family in osteoarthritis. Biomolecules 2022, 12, 959. [Google Scholar] [CrossRef]
- Guo, Y.; Cui, S.; Tian, Y.; Zhao, N.; Zhang, Y.; Gan, Y.; Zhou, Y.; Wang, X. Chondrocyte apoptosis in temporomandibular joint osteoarthritis promotes bone resorption by enhancing chemotaxis of osteoclast precursors. Osteoarthr. Cartil. 2022, 30, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Zhao, T.; Dai, Y.; Shi, M.; Huang, X.; Wei, Y.; Shen, J.; Zhang, X.; Xie, Z.; Wang, Q.; et al. New insights into the interplay between autophagy and cartilage degeneration in osteoarthritis. Front. Cell Dev. Biol. 2022, 10, 1089668. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Sun, K.; Yu, S.; Luo, J.; Guo, J.; Lin, J.; Wang, G.; Guo, Z.; Ye, Y.; Guo, F. Chondrocyte ferroptosis contribute to the progression of osteoarthritis. J. Orthop. Transl. 2020, 27, 33–43. [Google Scholar] [CrossRef]
- Yang, J.; Hu, S.; Bian, Y.; Yao, J.; Wang, D.; Liu, X.; Guo, Z.; Zhang, S.; Peng, L. Targeting cell death: Pyroptosis, ferroptosis, apoptosis and necroptosis in osteoarthritis. Front. Cell Dev. Biol. 2022, 9, 789948. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xing, R.; Huang, Z.; Zhang, N.; Li, X.; Wang, P. Inhibition of synovial macrophage pyroptosis alleviates synovitis and fibrosis in knee osteoarthritis. Mediat. Inflamm. 2019, 2019, 2165918. [Google Scholar] [CrossRef]
- Finckh, A.; Gilbert, B.; Hodkinson, B.; Bae, S.-C.; Thomas, R.; Deane, K.D.; Alpizar-Rodriguez, D.; Lauper, K. Global epidemiology of rheumatoid arthritis. Nat. Rev. Rheumatol. 2022, 18, 591–602. [Google Scholar] [CrossRef]
- Alivernini, S.; MacDonald, L.; Elmesmari, A.; Finlay, S.; Tolusso, B.; Gigante, M.R.; Petricca, L.; Di Mario, C.; Bui, L.; Perniola, S.; et al. Distinct synovial tissue macrophage subsets regulate inflammation and remission in rheumatoid arthritis. Nat. Med. 2020, 26, 1295–1306. [Google Scholar] [CrossRef]
- Lin, Y.-J.; Anzaghe, M.; Schülke, S. Update on the pathomechanism, diagnosis, and treatment options for rheumatoid arthritis. Cells 2020, 9, 880. [Google Scholar] [CrossRef]
- Nygaard, G.; Firestein, G.S. Restoring synovial homeostasis in rheumatoid arthritis by targeting fibroblast-like synoviocytes. Nat. Rev. Rheumatol. 2020, 16, 316–333. [Google Scholar] [CrossRef]
- da Fonseca, L.J.S.; Nunes-Souza, V.; Goulart, M.O.F.; Rabelo, L.A. Oxidative stress in rheumatoid arthritis: What the future might hold regarding novel biomarkers and add-on therapies. Oxidative Med. Cell. Longev. 2019, 2019, 7536805. [Google Scholar] [CrossRef]
- Ansari, M.Y.; Ahmad, N.; Haqqi, T.M. Oxidative stress and inflammation in osteoarthritis pathogenesis: Role of polyphenols. Biomed. Pharmacother. 2020, 129, 110452. [Google Scholar] [CrossRef] [PubMed]
- Phull, A.-R.; Nasir, B.; Haq, I.U.; Kim, S.J. Oxidative stress, consequences and ROS mediated cellular signaling in rheumatoid arthritis. Chem. Interac. 2018, 281, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, X.; Fan, D.; Xia, Q.; Wang, M.; Pan, F. Common trace metals in rheumatoid arthritis: A systematic review and meta-analysis. J. Trace Elem. Med. Biol. 2019, 56, 81–89. [Google Scholar] [CrossRef]
- Jomova, K.; Makova, M.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Rhodes, C.J.; Valko, M. Essential metals in health and disease. Chem. Interac. 2022, 367, 110173. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, R.; Shen, J.; Jin, Y.; Chang, C.; Hong, M.; Guo, S.; He, D. Circulating level of blood iron and copper associated with inflammation and disease activity of rheumatoid arthritis. Biol. Trace Elem. Res. 2022, 201, 90–97. [Google Scholar] [CrossRef]
- Ballou, E.R.; Wilson, D. The roles of zinc and copper sensing in fungal pathogenesis. Curr. Opin. Microbiol. 2016, 32, 128–134. [Google Scholar] [CrossRef]
- Malavolta, M.; Giacconi, R.; Piacenza, F.; Santarelli, L.; Cipriano, C.; Costarelli, L.; Tesei, S.; Pierpaoli, S.; Basso, A.; Galeazzi, R.; et al. Plasma copper/zinc ratio: An inflammatory/nutritional biomarker as predictor of all-cause mortality in elderly population. Biogerontology 2009, 11, 309–319. [Google Scholar] [CrossRef]
- Khadim, R.M.; Al-Fartusie, F.S. Evaluation of some trace elements and antioxidants in sera of patients with rheumatoid arthritis: A case–control study. Clin. Rheumatol. 2022, 42, 55–65. [Google Scholar] [CrossRef]
- Sahebari, M.; Ayati, R.; Mirzaei, H.; Sahebkar, A.; Hejazi, S.; Saghafi, M.; Saadati, N.; Ferns, G.A.; Ghayour-Mobarhan, M. Serum trace element concentrations in rheumatoid arthritis. Biol. Trace Elem. Res. 2015, 171, 237–245. [Google Scholar] [CrossRef]
- Valko, M.; Morris, H.; Cronin, M.T.D. Metals, toxicity and oxidative stress. Curr. Med. Chem. 2005, 12, 1161–1208. [Google Scholar] [CrossRef]
- Culotta, V.C.; Klomp, L.W.J.; Strain, J.; Casareno, R.L.B.; Krems, B.; Gitlin, J.D. The copper chaperone for superoxide dismutase. J. Biol. Chem. 1997, 272, 23469–23472. [Google Scholar] [CrossRef] [PubMed]
- Pufahl, R.A.; Singer, C.P.; Peariso, K.L.; Lin, S.-J.; Schmidt, P.J.; Fahrni, C.J.; Culotta, V.C.; Penner-Hahn, J.E.; O’Halloran, T.V. Metal ion chaperone function of the soluble Cu(I) receptor Atx1. Science 1997, 278, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Hatori, Y.; Lutsenko, S. The role of copper chaperone atox1 in coupling redox homeostasis to intracellular copper distribution. Antioxidants 2016, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Wee, N.K.; Weinstein, D.C.; Fraser, S.T.; Assinder, S.J. The mammalian copper transporters CTR1 and CTR2 and their roles in development and disease. Int. J. Biochem. Cell Biol. 2013, 45, 960–963. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-Y.; Liebig, J.K.; Tsigelny, I.F.; Howell, S.B. The copper transporter 1 (CTR1) is required to maintain the stability of copper transporter 2 (CTR2). Metallomics 2015, 7, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.C. Ceruloplasmin and other copper binding components of blood plasma and their functions: An update. Metallomics 2016, 8, 887–905. [Google Scholar] [CrossRef]
- Woimant, F.; Djebrani-Oussedik, N.; Poujois, A. New tools for Wilson’s disease diagnosis: Exchangeable copper fraction. Ann. Transl. Med. 2019, 7, S70. [Google Scholar] [CrossRef]
- Kirsipuu, T.; Zadorožnaja, A.; Smirnova, J.; Friedemann, M.; Plitz, T.; Tõugu, V.; Palumaa, P. Copper(II)-binding equilibria in human blood. Sci. Rep. 2020, 10, 5686. [Google Scholar] [CrossRef]
- Malavolta, M.; Piacenza, F.; Basso, A.; Giacconi, R.; Costarelli, L.; Mocchegiani, E. Serum copper to zinc ratio: Relationship with aging and health status. Mech. Ageing Dev. 2015, 151, 93–100. [Google Scholar] [CrossRef]
- Sidhu, A.; Miller, P.J.; Hollenbach, A.D. FOXO1 stimulates ceruloplasmin promoter activity in human hepatoma cells treated with IL-6. Biochem. Biophys. Res. Commun. 2011, 404, 963–967. [Google Scholar] [CrossRef]
- Aldabbagh, K.A.O.; Al-Bustany, D.A. Relationship of serum copper and HLADR4 tissue typing to disease activity and severity in patients with rheumatoid arthritis: A cross sectional study. Ann. Med. Surg. 2021, 73, 103193. [Google Scholar] [CrossRef] [PubMed]
- Schweigel-Röntgen, M. The families of zinc (SLC30 and SLC39) and copper (SLC31) transporters. In Current Topics in Membranes; Elsevier: Amsterdam, The Netherlands, 2014; Volume 73, pp. 321–355. [Google Scholar]
- Aydemir, T.B.; Chang, S.-M.; Guthrie, G.J.; Maki, A.B.; Ryu, M.-S.; Karabiyik, A.; Cousins, R.J. Zinc transporter ZIP14 functions in hepatic zinc, iron and glucose homeostasis during the innate immune response (endotoxemia). PLoS ONE 2012, 7, e48679. [Google Scholar] [CrossRef]
- Milanino, R.; Marrella, M.; Gasperini, R.; Pasqualicchio, M.; Velo, G. Copper and zinc body levels in inflammation: An overview of the data obtained from animal and human studies. Inflamm. Res. 1993, 39, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Simkin, P. Oral zinc sulphate in rheumatoid arthritis. Lancet 1976, 308, 539–542. [Google Scholar] [CrossRef]
- Li, G.; Cheng, T.; Yu, X. The impact of trace elements on osteoarthritis. Front. Med. 2021, 8, 771297. [Google Scholar] [CrossRef]
- Yazar, M.; Sarban, S.; Kocyigit, A.; Isikan, U.E. Synovial fluid and plasma selenium, copper, zinc, and iron concentrations in patients with rheumatoid arthritis and osteoarthritis. Biol. Trace Elem. Res. 2005, 106, 123–132. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, C.; Sun, Y.; Francis, M.; Ryu, M.; Grider, A.; Ye, K. Genetically predicted circulating levels of copper and zinc are associated with osteoarthritis but not with rheumatoid arthritis. Osteoarthr. Cartil. 2021, 29, 1029–1035. [Google Scholar] [CrossRef]
- Al-Bari, A.A.; Al Mamun, A. Current advances in regulation of bone homeostasis. FASEB BioAdv. 2020, 2, 668–679. [Google Scholar] [CrossRef]
- Kim, J.-M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.-H. Osteoblast-osteoclast communication and bone homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Halloran, D.; Durbano, H.W.; Nohe, A. Bone morphogenetic protein-2 in development and bone homeostasis. J. Dev. Biol. 2020, 8, 19. [Google Scholar] [CrossRef]
- Komori, T. Signaling networks in RUNX2-dependent bone development. J. Cell. Biochem. 2011, 112, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Gong, Y.; Xu, L.; Zhou, M.; Li, J.; Song, J. Bidirectional regulation of osteogenic differentiation by the FOXO subfamily of Forkhead transcription factors in mammalian MSCs. Cell Prolif. 2019, 52, e12540. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xu, Z.; Duan, C.; Liu, W.; Sun, J.; Han, B. Role of TCF/LEF transcription factors in bone development and osteogenesis. Int. J. Med. Sci. 2018, 15, 1415–1422. [Google Scholar] [CrossRef]
- Amarasekara, D.S.; Yun, H.; Kim, S.; Lee, N.; Kim, H.; Rho, J. Regulation of osteoclast differentiation by cytokine networks. Immune Netw. 2018, 18, e8. [Google Scholar] [CrossRef]
- Choi, J.U.A.; Kijas, A.W.; Lauko, J.; Rowan, A.E. The mechanosensory role of osteocytes and implications for bone health and disease states. Front. Cell Dev. Biol. 2022, 9, 770143. [Google Scholar] [CrossRef]
- Zhu, S.; Pang, Y.; Xu, J.; Chen, X.; Zhang, C.; Wu, B.; Gao, J. Endocrine regulation on bone by thyroid. Front. Endocrinol. 2022, 13, 873820. [Google Scholar] [CrossRef]
- Yao, D.; Huang, L.; Ke, J.; Zhang, M.; Xiao, Q.; Zhu, X. Bone metabolism regulation: Implications for the treatment of bone diseases. BioMedicine 2020, 129, 110494. [Google Scholar] [CrossRef]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The basic science of articular cartilage: Structure, composition, and function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Carballo, C.B.; Nakagawa, Y.; Sekiya, I.; Rodeo, S.A. Basic science of articular cartilage. Clin. Sports Med. 2017, 36, 413–425. [Google Scholar] [CrossRef]
- Zhao, Z.; Fan, C.; Chen, F.; Sun, Y.; Xia, Y.; Ji, A.; Wang, D. Progress in articular cartilage tissue engineering: A review on therapeutic cells and macromolecular scaffolds. Macromol. Biosci. 2019, 20, e1900278. [Google Scholar] [CrossRef]
- Nishimura, R.; Hata, K.; Nakamura, E.; Murakami, T.; Takahata, Y. Transcriptional network systems in cartilage development and disease. Histochem. Cell Biol. 2018, 149, 353–363. [Google Scholar] [CrossRef]
- Song, H.; Park, K.-H. Regulation and function of SOX9 during cartilage development and regeneration. Semin. Cancer Biol. 2020, 67, 12–23. [Google Scholar] [CrossRef]
- Karuppal, R. Current concepts in the articular cartilage repair and regeneration. J. Orthop. 2017, 14, A1–A3. [Google Scholar] [CrossRef]
- Makris, E.A.; Gomoll, A.H.; Malizos, K.N.; Hu, J.C.; Athanasiou, K.A. Repair and tissue engineering techniques for articular cartilage. Nat. Rev. Rheumatol. 2014, 11, 21–34. [Google Scholar] [CrossRef]
- Qi, J.; Yu, T.; Hu, B.; Wu, H.; Ouyang, H. Current biomaterial-based bone tissue engineering and translational medicine. Int. J. Mol. Sci. 2021, 22, 10233. [Google Scholar] [CrossRef] [PubMed]
- Shibli, J.A.; Nagay, B.E.; Suárez, L.J.; Hung, C.U.; Bertolini, M.; Barão, V.A.; Souza, J.G.S. Bone tissue engineering using osteogenic cells: From the bench to the clinical application. Tissue Eng. Part C Methods 2022, 28, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Liang, Z.; Yang, L.; Du, W.; Yu, T.; Tang, H.; Li, C.; Qiu, H. The role of natural polymers in bone tissue engineering. J. Control. Release 2021, 338, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.B.; Lalzawmliana, V.; Saidivya, M.; Kumar, V.; Roy, M.; Nandi, S.K. Magnesium phosphate bioceramics for bone tissue engineering. Chem. Rec. 2022, 22, e202200136. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, X.; Zhao, Q.; Tan, B.; Chen, X.; Liao, J. Role of hydrogels in bone tissue engineering: How properties shape regeneration. J. Biomed. Nanotechnol. 2020, 16, 1667–1686. [Google Scholar] [CrossRef]
- Rout, P.K.; Roy, S.; Ganguly, S.; Rathore, D.K. A review on properties of magnesium-based alloys for biomedical applications. Biomed. Phys. Eng. Express 2022, 8, 042002. [Google Scholar] [CrossRef] [PubMed]
- Glenske, K.; Donkiewicz, P.; Köwitsch, A.; Milosevic-Oljaca, N.; Rider, P.; Rofall, S.; Franke, J.; Jung, O.; Smeets, R.; Schnettler, R.; et al. Applications of metals for bone regeneration. Int. J. Mol. Sci. 2018, 19, 826. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, V.; Sumathi, S.; Narayanasamy, A.N.R. Tricomponent composite containing copper–hydroxyapatite/chitosan/polyvinyl pyrrolidone for bone tissue engineering. J. Biomed. Mater. Res. Part A 2020, 108, 1867–1880. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Huang, D.; Zhu, T.; Yu, X.; Xu, K.; Li, H.; Qu, H.; Zhou, Z.; Cheng, K.; Wen, W.; et al. Sustained zinc release in cooperation with CaP scaffold promoted bone regeneration via directing stem cell fate and triggering a pro-healing immune stimuli. J. Nanobiotechnology 2021, 19, 207. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, M.; Gu, W.; Shen, Z.; Ma, X.; Lu, F.; Yang, X.; Zheng, Y.; Gou, Z. Zinc-/copper-substituted dicalcium silicate cement: Advanced biomaterials with enhanced osteogenesis and long-term antibacterial properties. J. Mater. Chem. B 2020, 8, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cui, Y.; Liu, H.; Tian, Y.; Wang, G.; Fan, Y.; Wang, J.; Wu, D.; Wang, Y. Application of bioactive metal ions in the treatment of bone defects. J. Mater. Chem. B 2022, 10, 9369–9388. [Google Scholar] [CrossRef]
- Habibovic, P.; Barralet, J. Bioinorganics and biomaterials: Bone repair. Acta Biomater. 2011, 7, 3013–3026. [Google Scholar] [CrossRef]
- Costa, M.I.; Sarmento-Ribeiro, A.B.; Gonçalves, A.C. Zinc: From biological functions to therapeutic potential. Int. J. Mol. Sci. 2023, 24, 4822. [Google Scholar] [CrossRef]
- Molenda, M.; Kolmas, J. The role of zinc in bone tissue health and regeneration—A review. Biol. Trace Elem. Res. 2023, 201, 5640–5651. [Google Scholar] [CrossRef]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N.; Nakagawa, M.; Tsujimura, N.; Miyazaki, S.; Kizu, K.; Goto, T.; Komatsu, Y.; et al. The physiological, biochemical, and molecular roles of zinc transporters in zinc homeostasis and metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. Int. Rev. J. 2013, 4, 82–91. [Google Scholar] [CrossRef]
- Hou, R.; He, Y.; Yan, G.; Hou, S.; Xie, Z.; Liao, C. Zinc enzymes in medicinal chemistry. Eur. J. Med. Chem. 2021, 226, 113877. [Google Scholar] [CrossRef]
- Singh, J.K.; van Attikum, H. DNA double-strand break repair: Putting zinc fingers on the sore spot. Semin. Cell Dev. Biol. 2020, 113, 65–74. [Google Scholar] [CrossRef]
- Jarosz, M.; Olbert, M.; Wyszogrodzka, G.; Młyniec, K.; Librowski, T. Antioxidant and anti-inflammatory effects of zinc. Zinc-dependent NF-κB signaling. Inflammopharmacology 2017, 25, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Olechnowicz, J.; Tinkov, A.; Skalny, A.; Suliburska, J. Zinc status is associated with inflammation, oxidative stress, lipid, and glucose metabolism. J. Physiol. Sci. 2017, 68, 19–31. [Google Scholar] [CrossRef]
- Foster, M.; Samman, S. Zinc and regulation of inflammatory cytokines: Implications for cardiometabolic disease. Nutrients 2012, 4, 676–694. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.-W.; Fan, J.; Bai, S.-L.; Hou, W.-J.; Li, X.; Tong, H. Zinc prevents abdominal aortic aneurysm formation by induction of A20-mediated suppression of NF-κB pathway. PLoS ONE 2016, 11, e0148536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Dischler, A.; Glover, K.; Qin, Y. Neuronal signalling of zinc: From detection and modulation to function. Open Biol. 2022, 12, 220188. [Google Scholar] [CrossRef]
- Yamaguchi, M. Role of nutritional zinc in the prevention of osteoporosis. Mol. Cell. Biochem. 2009, 338, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S.; Miale, A., Jr.; Farid, Z.; Sandstead, H.H.; Schulert, A.R. Zinc metabolism in patients with the syndrome of iron deficiency anemia, hepatosplenomegaly, dwarfism, and hypognadism. J. Lab. Clin. Med. 1963, 61, 537–549. [Google Scholar] [PubMed]
- Prasad, A.S.; Halsted, J.A.; Nadimi, M. Syndrome of iron deficiency anemia, hepatosplenomegaly, hypogonadism, dwarfism and geophagia. Am. J. Med. 1961, 31, 532–546. [Google Scholar] [CrossRef]
- Sandstead, H.H.; Prasad, A.S.; Schulert, A.R.; Farid, Z.; Miale, A.; Bassilly, S.; Darby, W.J. Human zinc deficiency, endocrine manifestations and response to treatment. Am. J. Clin. Nutr. 1967, 20, 422–442. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; He, J.; Zheng, H.; Chen, C.; Jiang, H.; Lan, S. Zinc supplementation increased bone mineral density, improves bone histomorphology, and prevents bone loss in diabetic rat. Biol. Trace Elem. Res. 2019, 194, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Yan, G.; Guan, M. Zinc Homeostasis in bone: Zinc transporters and bone diseases. Int. J. Mol. Sci. 2020, 21, 1236. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, W.; Wang, M.; Backman, L.J.; Chen, J. Effects of zinc, magnesium, and iron ions on bone tissue engineering. ACS Biomater. Sci. Eng. 2022, 8, 2321–2335. [Google Scholar] [CrossRef] [PubMed]
- Nizet, A.; Cavalier, E.; Stenvinkel, P.; Haarhaus, M.; Magnusson, P. Bone alkaline phosphatase: An important biomarker in chronic kidney disease—Mineral and bone disorder. Clin. Chim. Acta 2019, 501, 198–206. [Google Scholar] [CrossRef]
- Vimalraj, S. Alkaline phosphatase: Structure, expression and its function in bone mineralization. Gene 2020, 754, 144855. [Google Scholar] [CrossRef]
- Seo, H.-J.; Cho, Y.-E.; Kim, T.; Shin, H.-I.; Kwun, I.-S. Zinc may increase bone formation through stimulating cell proliferation, alkaline phosphatase activity and collagen synthesis in osteoblastic MC3T3-E1 cells. Nutr. Res. Pract. 2010, 4, 356–361. [Google Scholar] [CrossRef]
- Bortolin, R.H.; Abreu, B.J.d.G.A.; Ururahy, M.A.G.; de Souza, K.S.C.; Bezerra, J.F.; Loureiro, M.B.; da Silva, F.S.; Marques, D.E.d.S.; Batista, A.A.d.S.; Oliveira, G.; et al. Protection against T1DM-Induced bone loss by zinc supplementation: Biomechanical, histomorphometric, and molecular analyses in STZ-induced diabetic rats. PLoS ONE 2015, 10, e0125349. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, A.; Yamaguchi, M. Increase in bone growth factors with healing rat fractures: The enhancing effect of zinc. Int. J. Mol. Med. 2001, 8, 433–438. [Google Scholar] [CrossRef]
- Igarashi, A.; Yamaguchi, M. Increase in bone protein components with healing rat fractures: Enhancement by zinc treatment. Int. J. Mol. Med. 1999, 4, 615–635. [Google Scholar] [CrossRef]
- Igarashi, A.; Yamaguchi, M. Great increase in bone 66 kDa protein and osteocalcin at later stages with healing rat fractures: Effect of zinc treatment. Int. J. Mol. Med. 2003, 11, 223–228. [Google Scholar] [CrossRef]
- Matsui, T.; Yamaguchi, M. Zinc modulation of insulin-like growth factor’s effect in osteoblastic MC3T3-E1 cells. Peptides 1995, 16, 1063–1068. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Goto, M.; Uchiyama, S.; Nakagawa, T. Effect of zinc on gene expression in osteoblastic MC3T3-E1 cells: Enhancement of Runx2, OPG, and regucalcin mRNA expressions. Mol. Cell. Biochem. 2008, 312, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.-O.; Kwun, I.-S. Zinc upregulates bone-specific transcription factor Runx2 expression via BMP-2 signaling and Smad-1 phosphorylation in osteoblasts. J. Nutr. Health 2018, 51, 23–30. [Google Scholar] [CrossRef]
- Fu, X.; Li, Y.; Huang, T.; Yu, Z.; Ma, K.; Yang, M.; Liu, Q.; Pan, H.; Wang, H.; Wang, J.; et al. Runx2/Osterix and zinc uptake synergize to orchestrate osteogenic differentiation and citrate containing bone apatite formation. Adv. Sci. 2018, 5, 1700755. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Choi, Y.; Yoon, D.S.; Lee, K.-M.; Kim, D.; Lee, J.W. Zinc promotes osteoblast differentiation in human mesenchymal stem cells via activation of the cAMP-PKA-CREB signaling pathway. Stem Cells Dev. 2018, 27, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Fathi, E.; Farahzadi, R. Enhancement of osteogenic differentiation of rat adipose tissue-derived mesenchymal stem cells by zinc sulphate under electromagnetic field via the PKA, ERK1/2 and Wnt/β-catenin signaling pathways. PLoS ONE 2017, 12, e0173877. [Google Scholar] [CrossRef]
- Liang, D.; Yang, M.; Guo, B.; Cao, J.; Yang, L.; Guo, X.; Li, Y.; Gao, Z. Zinc inhibits H2O2-induced MC3T3-E1 cells apoptosis via mapk and pi3k/akt pathways. Biol. Trace Elem. Res. 2012, 148, 420–429. [Google Scholar] [CrossRef]
- Moonga, B.S.; Dempster, D.W. Zinc is a potent inhibitor of osteoclastic bone resorption in vitro. J. Bone Miner. Res. 1995, 10, 453–457. [Google Scholar] [CrossRef]
- Sasaki, S.; Tsukamoto, M.; Saito, M.; Hojyo, S.; Fukada, T.; Takami, M.; Furuichi, T. Disruption of the mouse Slc39a14 gene encoding zinc transporter ZIP14 is associated with decreased bone mass, likely caused by enhanced bone resorption. FEBS Open Bio 2018, 8, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Weitzmann, M.N. Zinc stimulates osteoblastogenesis and suppresses osteoclastogenesis by antagonizing NF-κB activation. Mol. Cell. Biochem. 2011, 355, 179–186. [Google Scholar] [CrossRef]
- Park, K.H.; Park, B.; Yoon, D.S.; Kwon, S.-H.; Shin, D.M.; Lee, J.W.; Lee, H.G.; Shim, J.-H.; Park, J.H.; Lee, J.M. Zinc inhibits osteoclast differentiation by suppression of Ca2+-Calcineurin-NFATc1 signaling pathway. Cell Commun. Signal. 2013, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.Y.; Kang, N.; Yang, Y.-M.; Hong, J.H.; Shin, D.M. The role of Ca2+-NFATc1 signaling and its modulation on osteoclastogenesis. Int. J. Mol. Sci. 2020, 21, 3646. [Google Scholar] [CrossRef]
- Amin, N.; Clark, C.C.; Taghizadeh, M.; Djafarnejad, S. Zinc supplements and bone health: The role of the RANKL-RANK axis as a therapeutic target. J. Trace Elem. Med. Biol. 2019, 57, 126417. [Google Scholar] [CrossRef]
- Ferreira, E.C.; Bortolin, R.H.; Freire-Neto, F.P.; Souza, K.S.; Bezerra, J.F.; Ururahy, M.A.; Ramos, A.M.; Himelfarb, S.T.; Abreu, B.J.; Didone, T.V.; et al. Zinc supplementation reduces RANKL/OPG ratio and prevents bone architecture alterations in ovariectomized and type 1 diabetic rats. Nutr. Res. 2017, 40, 48–56. [Google Scholar] [CrossRef]
- Li, X.; Senda, K.; Ito, A.; Sogo, Y.; Yamazaki, A. Effect of Zn and Mg in tricalcium phosphate and in culture medium on apoptosis and actin ring formation of mature osteoclasts. Biomed. Mater. 2008, 3, 045002. [Google Scholar] [CrossRef]
- Wang, B.; Yang, M.; Liu, L.; Yan, G.; Yan, H.; Feng, J.; Li, Z.; Li, D.; Sun, H.; Yang, B. Osteogenic potential of Zn2+-passivated carbon dots for bone regeneration in vivo. Biomater. Sci. 2019, 7, 5414–5423. [Google Scholar] [CrossRef]
- Preeth, D.R.; Saravanan, S.; Shairam, M.; Selvakumar, N.; Raja, I.S.; Dhanasekaran, A.; Vimalraj, S.; Rajalakshmi, S. Bioactive Zinc(II) complex incorporated PCL/gelatin electrospun nanofiber enhanced bone tissue regeneration. Eur. J. Pharm. Sci. 2021, 160, 105768. [Google Scholar] [CrossRef]
- Jin, G.; Cao, H.; Qiao, Y.; Meng, F.; Zhu, H.; Liu, X. Osteogenic activity and antibacterial effect of zinc ion implanted titanium. Colloids Surf. B Biointerfaces 2014, 117, 158–165. [Google Scholar] [CrossRef]
- Maleki-Ghaleh, H.; Siadati, M.H.; Fallah, A.; Koc, B.; Kavanlouei, M.; Khademi-Azandehi, P.; Moradpur-Tari, E.; Omidi, Y.; Barar, J.; Beygi-Khosrowshahi, Y.; et al. Antibacterial and cellular behaviors of novel zinc-doped hydroxyapatite/graphene nanocomposite for bone tissue engineering. Int. J. Mol. Sci. 2021, 22, 9564. [Google Scholar] [CrossRef]
- Song, Y.; Wu, H.; Gao, Y.; Li, J.; Lin, K.; Liu, B.; Lei, X.; Cheng, P.; Zhang, S.; Wang, Y.; et al. Zinc silicate/nano-hydroxyapatite/collagen scaffolds promote angiogenesis and bone regeneration via the p38 MAPK pathway in activated monocytes. ACS Appl. Mater. Interfaces 2020, 12, 16058–16075. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Leach, R.M.; Fosmire, G.J.; Gay, C.V. Short-term zinc deficiency inhibits chondrocyte proliferation and induces cell apoptosis in the epiphyseal growth plate of young chickens. J. Nutr. 2002, 132, 665–673. [Google Scholar] [CrossRef]
- Rodríguez, J.P.; Rosselot, G. Effects of zinc on cell proliferation and proteoglycan characteristics of epiphyseal chondrocytes. J. Cell. Biochem. 2001, 82, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Hozain, S.; Hernandez, A.; Fuller, J.; Sharp, G.; Cottrell, J. Zinc chloride affects chondrogenesis via VEGF signaling. Exp. Cell Res. 2021, 399, 112436. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-C.; Chang, W.-T.; Hu, Y.-C.; Hsieh, B.-S.; Cheng, H.-L.; Yen, J.-H.; Chiu, P.-R.; Chang, K.-L. Zinc protects articular chondrocytes through changes in Nrf2-mediated antioxidants, cytokines and matrix metalloproteinases. Nutrients 2018, 10, 471. [Google Scholar] [CrossRef]
- Bhardwaj, V.; Deepika, P.; Basavarajaiah, S. Zinc incorporated nano hydroxyapatite: A novel bone graft used for regeneration of intrabony defects. Contemp. Clin. Dent. 2018, 9, 427–433. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Li, S.; Huang, X.; Chen, X.; Shan, H.; Zhang, M. The role of copper homeostasis in brain disease. Int. J. Mol. Sci. 2022, 23, 13850. [Google Scholar] [CrossRef]
- Festa, R.A.; Thiele, D.J. Copper: An essential metal in biology. Curr. Biol. 2011, 21, R877–R883. [Google Scholar] [CrossRef]
- Vallet, S.D.; Ricard-Blum, S. Lysyl oxidases: From enzyme activity to extracellular matrix cross-links. Essays Biochem. 2019, 63, 349–364. [Google Scholar] [CrossRef]
- Trist, B.G.; Hilton, J.B.; Hare, D.J.; Crouch, P.J.; Double, K.L. Superoxide Dismutase 1 in Health and Disease: How a Frontline Antioxidant Becomes Neurotoxic. Angew. Chem. Int. Ed. 2020, 60, 9215–9246. [Google Scholar] [CrossRef]
- Swaminathan, A.B.; Gohil, V.M. The role of COA6 in the mitochondrial copper delivery pathway to cytochrome c oxidase. Biomolecules 2022, 12, 125. [Google Scholar] [CrossRef] [PubMed]
- Alasvand, N.; Behnamghader, A.; Milan, P.B.; Simorgh, S.; Mobasheri, A.; Mozafari, M. Tissue-engineered small-diameter vascular grafts containing novel copper-doped bioactive glass biomaterials to promote angiogenic activity and endothelial regeneration. Mater. Today Bio 2023, 20, 100647. [Google Scholar] [CrossRef] [PubMed]
- Wazir, S.M.; Ghobrial, I. Copper deficiency, a new triad: Anemia, leucopenia, and myeloneuropathy. J. Community Hosp. Intern. Med. Perspect. 2017, 7, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Baxter, J.H.; Van Wyk, J.J.; Follis, R.H., Jr. A bone disorder associated with copper deficiency. II. Histological and chemical studies on the bones. Bull. Johns Hopkins Hosp. 1953, 93, 25–39. [Google Scholar]
- Wu, Z.; Yuan, Y.; Tian, J.; Long, F.; Luo, W. The associations between serum trace elements and bone mineral density in children under 3 years of age. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- Qu, X.; He, Z.; Qiao, H.; Zhai, Z.; Mao, Z.; Yu, Z.; Dai, K. Serum copper levels are associated with bone mineral density and total fracture. J. Orthop. Transl. 2018, 14, 34–44. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, F.; Liu, J.; Smått, J.-H.; Gepperth, D.; Lastusaari, M.; Xu, C.; Hupa, L. Biocomposites of copper-containing mesoporous bioactive glass and nanofibrillated cellulose: Biocompatibility and angiogenic promotion in chronic wound healing application. Acta Biomater. 2016, 46, 286–298. [Google Scholar] [CrossRef]
- Baino, F.; Potestio, I.; Vitale-Brovarone, C. Production and physicochemical characterization of Cu-doped silicate bioceramic scaffolds. Materials 2018, 11, 1524. [Google Scholar] [CrossRef]
- Jin, S.; Ren, L.; Yang, K. Bio-functional Cu containing biomaterials: A new way to enhance bio-adaption of biomaterials. J. Mater. Sci. Technol. 2016, 32, 835–839. [Google Scholar] [CrossRef]
- Bari, A.; Bloise, N.; Fiorilli, S.; Novajra, G.; Vallet-Regí, M.; Bruni, G.; Torres-Pardo, A.; González-Calbet, J.M.; Visai, L.; Vitale-Brovarone, C. Copper-containing mesoporous bioactive glass nanoparticles as multifunctional agent for bone regeneration. Acta Biomater. 2017, 55, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhou, Y.; Xu, M.; Han, P.; Chen, L.; Chang, J.; Xiao, Y. Copper-containing mesoporous bioactive glass scaffolds with multifunctional properties of angiogenesis capacity, osteostimulation and antibacterial activity. Biomaterials 2013, 34, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Popescu, R.A.; Magyari, K.; Vulpoi, A.; Trandafir, D.L.; Licarete, E.; Todea, M.; Ştefan, R.; Voica, C.; Vodnar, D.C.; Simon, S.; et al. Bioactive and biocompatible copper containing glass-ceramics with remarkable antibacterial properties and high cell viability designed for future in vivo trials. Biomater. Sci. 2016, 4, 1252–1265. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, W.; Yao, Q. Copper-based biomaterials for bone and cartilage tissue engineering. J. Orthop. Transl. 2021, 29, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Rivera, L.R.; Cochis, A.; Biser, S.; Canciani, E.; Ferraris, S.; Rimondini, L.; Boccaccini, A.R. Antibacterial, pro-angiogenic and pro-osteointegrative zein-bioactive glass/copper based coatings for implantable stainless steel aimed at bone healing. Bioact. Mater. 2020, 6, 1479–1490. [Google Scholar] [CrossRef]
- Rucker, R.B.; Kosonen, T.; Clegg, M.S.; E Mitchell, A.; Rucker, B.R.; Uriu-Hare, J.Y.; Keen, C.L. Copper, lysyl oxidase, and extracellular matrix protein cross-linking. Am. J. Clin. Nutr. 1998, 67, 996S–1002S. [Google Scholar] [CrossRef]
- Jacobs, A.; Renaudin, G.; Forestier, C.; Nedelec, J.-M.; Descamps, S. Biological properties of copper-doped biomaterials for orthopedic applications: A review of antibacterial, angiogenic and osteogenic aspects. Acta Biomater. 2020, 117, 21–39. [Google Scholar] [CrossRef]
- Burghardt, I.; Lüthen, F.; Prinz, C.; Kreikemeyer, B.; Zietz, C.; Neumann, H.-G.; Rychly, J. A dual function of copper in designing regenerative implants. Biomaterials 2015, 44, 36–44. [Google Scholar] [CrossRef]
- Applerot, G.; Lellouche, J.; Lipovsky, A.; Nitzan, Y.; Lubart, R.; Gedanken, A.; Banin, E. Understanding the antibacterial mechanism of cuo nanoparticles: Revealing the route of induced oxidative stress. Small 2012, 8, 3326–3337. [Google Scholar] [CrossRef]
- Climova, A.; Pivovarova, E.; Szczesio, M.; Gobis, K.; Ziembicka, D.; Korga-Plewko, A.; Kubik, J.; Iwan, M.; Antos-Bielska, M.; Krzyżowska, M.; et al. Anticancer and antimicrobial activity of new copper (II) complexes. J. Inorg. Biochem. 2023, 240, 112108. [Google Scholar] [CrossRef]
- Tan, J.; Ren, L.; Xie, K.; Wang, L.; Jiang, W.; Guo, Y.; Hao, Y. Functionalized TiCu/TiCuN coating promotes osteoporotic fracture healing by upregulating the Wnt/β-catenin pathway. Regen. Biomater. 2022, 10, rbac092. [Google Scholar] [CrossRef]
- Vlashi, R.; Zhang, X.; Wu, M.; Chen, G. Wnt signaling: Essential roles in osteoblast differentiation, bone metabolism and therapeutic implications for bone and skeletal disorders. Genes Dis. 2023, 10, 1291–1317. [Google Scholar] [CrossRef] [PubMed]
- Maupin, K.A.; Droscha, C.J.; Williams, B.O. A Comprehensive overview of skeletal phenotypes associated with alterations in wnt/β-catenin signaling in humans and mice. Bone Res. 2013, 1, 27–71. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Xiao, Z.; Chen, Y.; Deng, Y.; Zeng, D.; Liu, Y.; Huang, F.; Wang, J.; Liu, Y.; Bellanti, J.A.; et al. CD39 produced from human gmscs regulates the balance of osteoclasts and osteoblasts through the wnt/β-catenin pathway in osteoporosis. Mol. Ther. 2020, 28, 1518–1532. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Feng, C.; Wang, W.; Wu, G.; Hu, Y.; Li, S.; Gao, X.; Chen, X.; Ji, Q. Copper-deposited diatom-biosilica enhanced osteogenic potential in periodontal ligament stem cells and rat cranium. J. Biomed. Mater. Res. Part B Appl. Biomater. 2023, 111, 1286–1298. [Google Scholar] [CrossRef]
- Li, S.; Zhang, L.; Liu, C.; Kim, J.; Su, K.; Chen, T.; Zhao, L.; Lu, X.; Zhang, H.; Cui, Y.; et al. Spontaneous immunomodulation and regulation of angiogenesis and osteogenesis by Sr/Cu-borosilicate glass (BSG) bone cement to repair critical bone defects. Bioact. Mater. 2023, 23, 101–117. [Google Scholar] [CrossRef]
- Wang, L.-J.; Ni, X.-H.; Zhang, F.; Peng, Z.; Yu, F.-X.; Zhang, L.-B.; Li, B.; Jiao, Y.; Li, Y.-K.; Yang, B.; et al. Osteoblast response to copper-doped microporous coatings on titanium for improved bone integration. Nanoscale Res. Lett. 2021, 16, 146. [Google Scholar] [CrossRef]
- Wang, L.; Li, G.; Ren, L.; Kong, X.; Wang, Y.; Han, X.; Jiang, W.; Dai, K.; Yang, K.; Hao, Y. Nano-copper-bearing stainless steel promotes fracture healing by accelerating the callus evolution process. Int. J. Nanomed. 2017, 12, 8443–8457. [Google Scholar] [CrossRef]
- Lin, W.; Xu, L.; Li, G. Molecular insights into lysyl oxidases in cartilage regeneration and rejuvenation. Front. Bioeng. Biotechnol. 2020, 8, 359. [Google Scholar] [CrossRef]
- Makris, E.A.; MacBarb, R.F.; Responte, D.J.; Hu, J.C.; Athanasiou, K.A. A copper sulfate and hydroxylysine treatment regimen for enhancing collagen cross-linking and biomechanical properties in engineered neocartilage. FASEB J. 2013, 27, 2421–2430. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, J.; Zhu, X.; Zhang, Z.; Ai, Y.; Sun, G.; Wang, Z.; Liu, G. Effect of copper on levels of collagen and alkaline phosphatase activity from chondrocytes in newborn piglets in vitro. Biol. Trace Elem. Res. 2011, 144, 597–605. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, X.; Li, X.; Wang, W.; Wang, X.; Liu, L.; Deng, Q.; Bai, G.; Wang, J.; Feng, H.; et al. Effects of copper on proliferation and autocrine secretion of insulin-like growth factor-1 (IGF-1) and IGF-binding protein-3 (IGFBP-3) in chondrocytes from newborn pigs in vitro. Biol. Trace Elem. Res. 2011, 144, 588–596. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, X.; Xie, G.; Zhang, Y.; Liu, Z.; Zhang, L.; Zhao, C.; Yang, W.; Shi, X.; Wang, Z.; et al. Effect of copper on the expression of IGF-1 from chondrocytes in newborn piglets in vitro. Biol. Trace Elem. Res. 2012, 148, 178–181. [Google Scholar] [CrossRef]
- Madzovska-Malagurski, I.; Vukasinovic-Sekulic, M.; Kostic, D.; Levic, S. Towards antimicrobial yet bioactive Cu-alginate hydrogels. Biomed. Mater. 2016, 11, 35015. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Chen, J.; Li, L.; Pu, X.; Chu, X.; Wang, X.; Li, M.; Lu, Y.; Zheng, X. Promotion of chondrogenic differentiation of mesenchymal stem cells by copper: Implications for new cartilage repair biomaterials. Mater. Sci. Eng. C 2018, 93, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Deng, C.; Li, X.; Liu, Y.; Zhang, M.; Qin, C.; Yao, Q.; Wang, L.; Wu, C. Copper-incorporated bioactive glass-ceramics inducing anti-inflammatory phenotype and regeneration of cartilage/bone interface. Theranostics 2019, 9, 6300–6313. [Google Scholar] [CrossRef]
- Wang, X.; Cai, Y.; Wu, C.; Liang, J.; Tang, K.; Lin, Z.; Chen, L.; Lu, Y.; Wang, Q. Conversion of senescent cartilage into a pro-chondrogenic microenvironment with antibody-functionalized copper sulfate nanoparticles for efficient osteoarthritis therapy. J. Nanobiotechnology 2023, 21, 258. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Han, S.; Zeng, X.; Zhu, C.; Pu, Y.; Sun, Y. Multifunctional thermo-sensitive hydrogel for modulating the microenvironment in Osteoarthritis by polarizing macrophages and scavenging RONS. J. Nanobiotechnology 2022, 20, 221. [Google Scholar] [CrossRef]
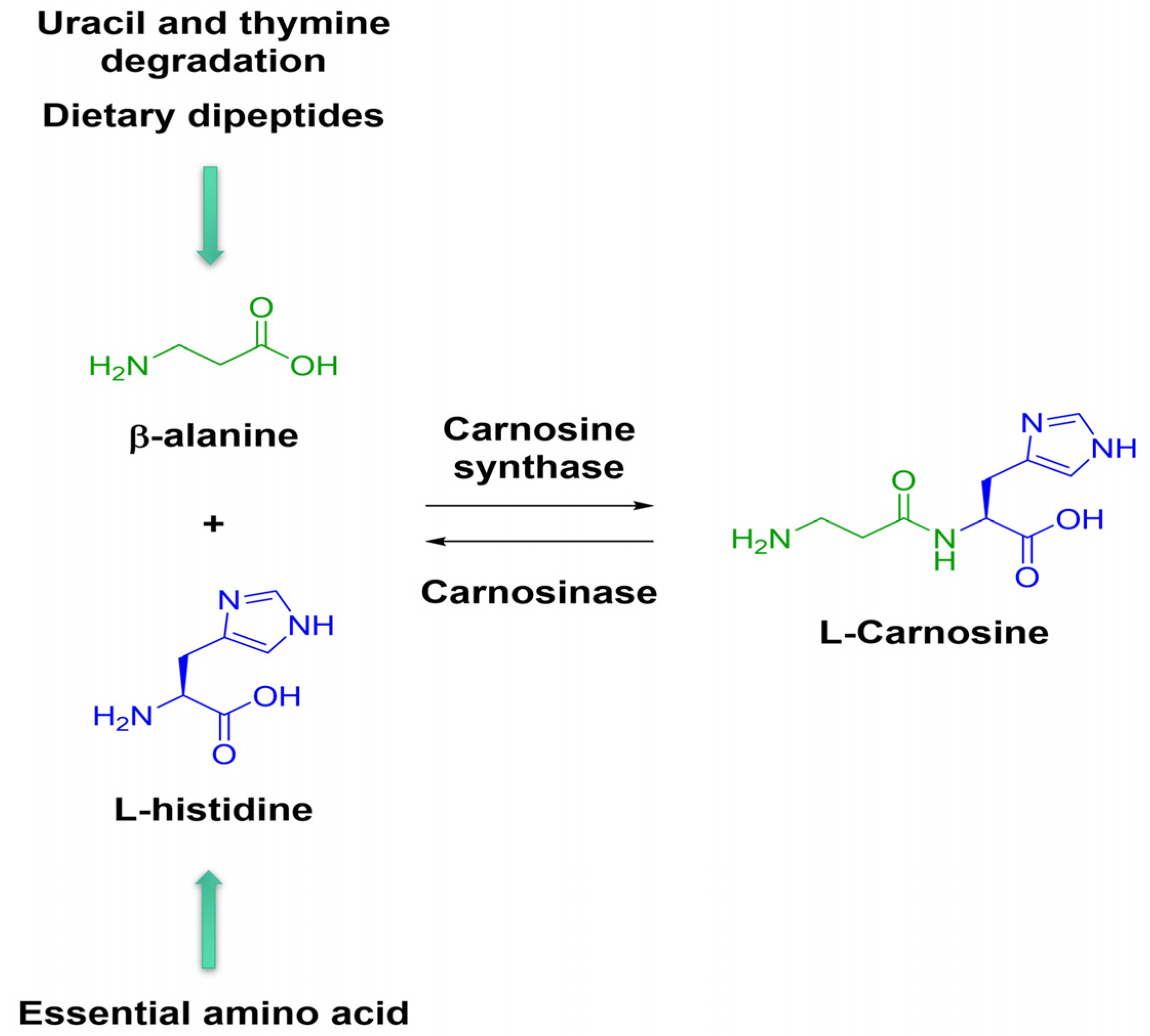
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and Pathophysiology of Carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef]
- Fresta, C.G.; Hogard, M.L.; Caruso, G.; Melo Costa, E.E.; Lazzarino, G.; Lunte, S.M. Monitoring carnosine uptake by RAW 264.7 macrophage cells using microchip electrophoresis with fluorescence detection. Anal. Methods 2017, 9, 402–408. [Google Scholar] [CrossRef]
- Chmielewska, K.; Dzierzbicka, K.; Inkielewicz-Stępniak, I.; Przybyłowska, M. Therapeutic potential of carnosine and its derivatives in the treatment of human diseases. Chem. Res. Toxicol. 2020, 33, 1561–1578. [Google Scholar] [CrossRef] [PubMed]
- Ito-Kato, E.; Suzuki, N.; Maeno, M.; Takada, T.; Tanabe, N.; Takayama, T.; Ito, K.; Otsuka, K. Effect of carnosine on runt-related transcription factor-2/core binding factor alpha-1 and Sox9 expressions of human periodontal ligament cells. J. Periodontal Res. 2004, 39, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; de Courten, B.; Regazzoni, L.; Gilardoni, E.; Ferrario, G.; Baron, G.; Altomare, A.; D’amato, A.; Vistoli, G.; Carini, M. Understanding the antioxidant and carbonyl sequestering activity of carnosine: Direct and indirect mechanisms. Free. Radic. Res. 2020, 55, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Solana-Manrique, C.; Sanz, F.J.; Martínez-Carrión, G.; Paricio, N. Antioxidant and neuroprotective effects of carnosine: Therapeutic implications in neurodegenerative diseases. Antioxidants 2022, 11, 848. [Google Scholar] [CrossRef]
- Zhao, K.; Li, Y.; Wang, Z.; Han, N.; Wang, Y. Carnosine protects mouse podocytes from high glucose induced apoptosis through pi3k/akt and nrf2 pathways. BioMed Res. Int. 2019, 2019, 4348973. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Y.; Kong, Y.; Zhang, X.; Zhang, H.; Gang, Y.; Bai, L. Carnosine prevents type 2 diabetes-induced osteoarthritis through the ROS/NF-κB pathway. Front. Pharmacol. 2018, 9, 598. [Google Scholar] [CrossRef]
- Ponist, S.; Drafi, F.; Kuncirova, V.; Mihalova, D.; Rackova, L.; Danisovic, L.; Ondrejickova, O.; Tumova, I.; Trunova, O.; Fedorova, T.; et al. Effect of carnosine in experimental arthritis and on primary culture chondrocytes. Oxidative Med. Cell. Longev. 2016, 2016, 8470589. [Google Scholar] [CrossRef]
- Fitzpatrick, M.; Young, S.P. Metabolomics—A novel window into inflammatory disease. Swiss Med Wkly. 2013, 143, w13743. [Google Scholar] [CrossRef]
- Qiu, S.; Cai, Y.; Yao, H.; Lin, C.; Xie, Y.; Tang, S.; Zhang, A. Small molecule metabolites: Discovery of biomarkers and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 132. [Google Scholar] [CrossRef]
- Bartikoski, B.J.; De Oliveira, M.S.; Santo, R.C.D.E.; Dos Santos, L.P.; Dos Santos, N.G.; Xavier, R.M. A review of metabolomic profiling in rheumatoid arthritis: Bringing new insights in disease pathogenesis, treatment and comorbidities. Metabolites 2022, 12, 394. [Google Scholar] [CrossRef]
- Sobczak, A.I.S.; Blindauer, C.A.; Stewart, A.J. Changes in plasma free fatty acids associated with type-2 diabetes. Nutrients 2019, 11, 2022. [Google Scholar] [CrossRef]
- Xu, L.; Chang, C.; Jiang, P.; Wei, K.; Zhang, R.; Jin, Y.; Zhao, J.; Xu, L.; Shi, Y.; Guo, S.; et al. Metabolomics in rheumatoid arthritis: Advances and review. Front. Immunol. 2022, 13, 961708. [Google Scholar] [CrossRef]
- Seiki, M.; Ueki, S.; Tanaka, Y.; Soeda, M.; Hori, Y.; Aita, H.; Yoneta, T.; Morita, H.; Tagashira, E.; Okabe, S. Studies on anti-ulcer effects of a new compound, zinc L-carnosine (Z-103). Folia Pharmacol. Jpn. 1990, 95, 257–269. [Google Scholar] [CrossRef]
- Furihata, K.; Tsuchikawa, M.; Miwa, T.; Naito, Y.; Oba, K.; Sakagami, M. Efficacy and safety of polaprezinc (zinc compound) on zinc deficiency: A systematic review and dose–response meta-analysis of randomized clinical trials using individual patient data. Nutrients 2020, 12, 1128. [Google Scholar] [CrossRef]
- Li, M.; Sun, Z.; Zhang, H.; Liu, Z. Recent advances on polaprezinc for medical use (Review). Exp. Ther. Med. 2021, 22, 1445. [Google Scholar] [CrossRef]
- Udechukwu, M.C.; Collins, S.A.; Udenigwe, C.C. Prospects of enhancing dietary zinc bioavailability with food-derived zinc-chelating peptides. Food Funct. 2016, 7, 4137–4144. [Google Scholar] [CrossRef]
- O’connor, J.P.; Kanjilal, D.; Teitelbaum, M.; Lin, S.S.; Cottrell, J.A. Zinc as a therapeutic agent in bone regeneration. Materials 2020, 13, 2211. [Google Scholar] [CrossRef]
- Hashizume, M.; Yamaguchi, M. Effect of ?-alanyl-L-histidinato zinc on differentiation of osteoblastic MC3T3-El cells: Increases in alkaline phosphatase activity and protein concentration. Mol. Cell. Biochem. 1994, 131, 19–24. [Google Scholar] [CrossRef]
- Maeno, M.; Ito-Kato, E.; Suzuki, N.; Takada, T.; Takayama, T.; Ito, K.; Otsuka, K. Effect of β-alanyl-L-histidinato zinc on the differentiation pathway of human periodontal ligament cells. Life Sci. 2004, 74, 2493–2504. [Google Scholar] [CrossRef]
- Takada, T.; Suzuki, N.; Ito-Kato, E.; Noguchi, Y.; Ito, M.; Maeno, M.; Otsuka, K. Effect of β-alanyl-L-histidinato zinc on the differentiation of C2C12 cells. Life Sci. 2004, 76, 509–520. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Kishi, S. Effect of zinc-chelating dipeptide on bone metabolism in weanling rats: Comparison with β-alanyl-l-histidinato zinc-related compounds. Peptides 1994, 15, 671–673. [Google Scholar] [CrossRef]
- Kisi, S.; Yamaguchi, M. Stimulatory effect of BETA.-alanyl-L-histidinato zinc on alkaline phosphatase activity in bone tissues from elderly rats: Comparison with zinc sulfate action. Biol. Pharm. Bull. 1994, 17, 345–347. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Kishi, S. Zinc compounds inhibit osteoclast-like cell formation at the earlier stage of rat marrow culture but not osteoclast function. Mol. Cell. Biochem. 1996, 158, 171–177. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Kishi, S. Inhibitory effect of zinc-chelating dipeptide on parathyroid hormone-stimulated osteoclast-like cell formation in mouse marrow cultures: Involvement of calcium signaling. Peptides 1995, 16, 629–633. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Kishi, S. Differential effects of transforming growth factor-β on osteoclast-like cell formation in mouse marrow culture: Relation to the effect of zinc-chelating dipeptides. Peptides 1995, 16, 1483–1488. [Google Scholar] [CrossRef]
- Kishi, S.; Yamaguchi, M. Inhibitory effect of zinc compounds on osteoclast-like cell formation in mouse marrow cultures. Biochem. Pharmacol. 1994, 48, 1225–1230. [Google Scholar] [CrossRef]
- Gao, X.; A Al-Baadani, M.; Wu, M.; Tong, N.; Shen, X.; Ding, X.; Liu, J. Study on the local anti-osteoporosis effect of polaprezinc-loaded antioxidant electrospun membrane. Int. J. Nanomed. 2022, 17, 17–29. [Google Scholar] [CrossRef]
- Branham, M.L.; Singh, P.; Bisetty, K.; Sabela, M.; Govender, T. Preparation, spectrochemical, and computational analysis of L-carnosine (2-[(3-aminopropanoyl)amino]-3-(1H-imidazol-5-yl)propanoic acid) and its ruthenium (II) coordination complexes in aqueous solution. Molecules 2011, 16, 10269–10291. [Google Scholar] [CrossRef]
- Abate, C.; Aiello, D.; Cordaro, M.; Giuffrè, O.; Napoli, A.; Foti, C. Binding ability of l-carnosine towards Cu2+, Mn2+ and Zn2+ in aqueous solution. J. Mol. Liq. 2022, 368, 120772. [Google Scholar] [CrossRef]
- Keenan, J.; Horgan, K.; Clynes, M.; Sinkunaite, I.; Ward, P.; Murphy, R.; O’sullivan, F. Unexpected fluctuations of trace element levels in cell culture medium in vitro: Caveat emptor. Vitr. Cell. Dev. Biol.-Anim. 2018, 54, 555–558. [Google Scholar] [CrossRef]
- Falcone, E.; Okafor, M.; Vitale, N.; Raibaut, L.; Sour, A.; Faller, P. Extracellular Cu2+ pools and their detection: From current knowledge to next-generation probes. Coord. Chem. Rev. 2021, 433, 213727. [Google Scholar] [CrossRef]
- Irving, H.; Williams, R.J.P. Order of stability of metal complexes. Nature 1948, 162, 746–747. [Google Scholar] [CrossRef]
- Stacy, A.E.; Palanimuthu, D.; Bernhardt, P.V.; Kalinowski, D.S.; Jansson, P.J.; Richardson, D.R. Zinc(II)–thiosemicarbazone complexes are localized to the lysosomal compartment where they transmetallate with copper ions to induce cytotoxicity. J. Med. Chem. 2016, 59, 4965–4984. [Google Scholar] [CrossRef] [PubMed]
- Oliveri, I.P.; Consiglio, G.; Munzi, G.; Failla, S.; Di Bella, S. Deaggregation properties and transmetalation studies of a zinc(ii) salen-type Schiff-base complex. Dalton Trans. 2022, 51, 11859–11867. [Google Scholar] [CrossRef]
- Oliveri, I.P.; Attinà, A.; Di Bella, S. A zinc(II) schiff base complex as fluorescent chemosensor for the selective and sensitive detection of copper(II) in aqueous solution. Sensors 2023, 23, 3925. [Google Scholar] [CrossRef]
- Christlieb, M.; Struthers, H.S.R.; Bonnitcha, P.D.; Cowley, A.R.; Dilworth, J.R. The exocyclic functionalisation of bis(thiosemicarbazonate) complexes of zinc and copper: The synthesis of monomeric and dimeric species. Dalton Trans. 2007, 5043–5054. [Google Scholar] [CrossRef]
- McAllum, E.J.; Roberts, B.R.; Hickey, J.L.; Dang, T.N.; Grubman, A.; Donnelly, P.S.; Liddell, J.R.; White, A.R.; Crouch, P.J. ZnII(atsm) is protective in amyotrophic lateral sclerosis model mice via a copper delivery mechanism. Neurobiol. Dis. 2015, 81, 20–24. [Google Scholar] [CrossRef]




| Cu Source | Cell Type | Animal Model | Effects | Outcomes | Ref |
|---|---|---|---|---|---|
| Cu-titanium implants | MSCs | - | ↑ ALP activity, COL1, OPG, OPN expression, ↓ E. coli and S. aureus growth | ↑ osteoblasts proliferation, differentiation, antimicrobial activity | [169] |
| Cu-titanium implants | BMSCs | rat femur fracture model | ↑ Wnt/β-catenin pathway | ↑ osteoporotic fractures healing | [172] |
| Cu-mineralized diatom | PDLSCs | rat cranial defect/full-thickness skin incision models | ↑ ALP, Runx2, BSP, OCN, OPN expression | ↑ osteogenesis, angiogenesis | [176] |
| Cu-borosilicate glass bone cement | HUVECs, hBMSCs | femoral condylar bone defects in rats and rabbits | ↑ Runx2, BGLAP, SPP1, IL-1Ra, TGF-β1, VEGF, FGF2 ↓ IL-1β, IL-6 expression | antiinflammatory, angiogenic, osteogenic properties | [177] |
| Cu- coatings on titanium | MC3T3-E1 | rabbit femoral condyle | ↑ cell adhesion, proliferation, differentiation | ↑ bone formation ↑ osseointegration | [178] |
| Nano Cu- bearing stainless steel | BMSCs | rat fracture model | ↑ LOX activity, osteogenic differentiation ↑ callus evolution | ↑ bone formation, fracture healing | [179] |
| Copper sulfate | chondrocytes from bovine joints | - | ↑ LOX activity | ↑ biomechanical properties in engineered neocartilage | [181] |
| Copper sulfate | chondrocytes from piglets | - | ↑ ALP activity, collagen content | ↑ chondrocytes proliferation | [182] |
| Copper sulfate | chondrocytes from pigs | - | ↑ IGF-1 and IGFBP-3 secretion | ↑ chondrocytes proliferation | [183] |
| Cu-alginate hydrogels | bovine calf chondrocytes | - | ↑ cell proliferation, ECM synthesis | ↑ chondrogenesis | [185] |
| Cu-alginate porous scaffold | MSCs | mice muscle pouch | ↑ extracellular GAG deposition, ↑ SOX9, ACAN, COL2 expression | ↑ chondrogenic differentiation, ↑ chondrogenesis | [186] |
| Cu-bioactive glass ceramics | rabbit chondrocytes, RAW264.7 cells | rabbit osteochondral defect model | ↑ HIF pathway ↑ anti-inflammatory cytokines | ↑ osteochondral regeneration, ↓ inflammation | [187] |
| Copper sulfide nanoparticles | ATDC5 | OA mouse model | ↑ GAG and COL2 synthesis, ↑ apoptosis in senescent chondrocytes | ↑ cartilage regeneration | [188] |
| Cu-doped hydrogel | chondrocytes, L929, RAW264.7 | rat OA model | ROS scavenging, macrophages repolarization | antioxidant activity, ↓ cartilage degradation, ↓ inflammation | [189] |
| Compound (Tested Conc.) | Cell Type | Medium | Serum | Supplements | Duration | Ref. |
|---|---|---|---|---|---|---|
| Polaprezinc (5 × 10−5 M) | hBMSCs | DMEM-LG | 10% FBS | 1% antibiotic– antimycotic solution, 10 mM β-glycerophosphate, 50 μg/mL ascorbic acid | 5–12 days | [17] |
| Car (5 × 10−5 M) | hBMSCs | DMEM-LG | 10% FBS | 1% antibiotic– antimycotic solution, 10 mM β-glycerophosphate, 50 μg/mL ascorbic acid | 5–12 days | [17] |
| Polaprezinc (5 × 10−5 M) | mBMMs | α-MEM | 10% FBS | 1% antibiotic– antimycotic solution, 10 ng/mL mM-CSF and 10 ng/mL mRANKL | - | [17] |
| Car (5 × 10−5 M) | mBMMs | α-MEM | 10% FBS | 1% antibiotic–antimycotic solution, 10 ng/mL mM-CSF and 10 ng/mL mRANKL | - | [17] |
| Car (10−4–10−5 M) | PDLSCs | α-MEM | 10% FBS | antibiotics | up to 10 days | [193] |
| Car (10−4 M) | FLSs | DMEM | 10% FBS | antibiotics (100 U/mL penicillin and 100 mg/mL streptomycin) | - | [197] |
| Car (10−4 M) | FLSs | DMEM | - | antibiotics penicillin and streptomycin (100 mg/mL) | - | [13] |
| Car (10−5 M) | Rat primary chondrocytes | DMEM/ Ham’s F-12 | 10% FBS | 1 mmol/L glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin | - | [198] |
| Polaprezinc (10−7–10−5 M) | MC3T3-E1 | α-MEM | 10% FBS | - | up to 10 days | [209] |
| Polaprezinc (10−4–10−5 M) | PDLSCs | α-MEM | 10% FBS | 1% (vol/vol) penicillin streptomycin solution (5000 units/mL penicillin and 50 mg/mL streptomycin) | up to 10 days | [210] |
| Polaprezinc (10−7–10−5 M) | Rat marrow culture | α-MEM | 10% HI FBS | - | 7 days | [214] |
| Polaprezinc (10−7–10−5 M) | osteoclasts isolated from rat femoral tissues | α-MEM | 10% FBS | - | 24 h | [214] |
| Polaprezinc (10−6 M) | Bone marrow cells isolated from mice | α-MEM | 10% HI FBS | Penicillin–streptomycin solution (5000 U/mL penicillin; 5000 pg/mL streptomycin) | 7 days | [215] |
| Polaprezinc (10−6 M) | Bone marrow cells isolated from mice | α-MEM | 10% HI FBS | Penicillin–streptomycin solution (5000 U/mL penicillin; 5000 pg/mL streptomycin) | 7 days | [216] |
| Polaprezinc (10−5 M) | MC3T3-E1 | α-MEM | 10% FBS | Penicillin–streptomycin (5000 U/mL penicillin; 5000 lg/mL streptomycin) | 24–72 h | [125] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciaffaglione, V.; Rizzarelli, E. Carnosine, Zinc and Copper: A Menage a Trois in Bone and Cartilage Protection. Int. J. Mol. Sci. 2023, 24, 16209. https://doi.org/10.3390/ijms242216209
Ciaffaglione V, Rizzarelli E. Carnosine, Zinc and Copper: A Menage a Trois in Bone and Cartilage Protection. International Journal of Molecular Sciences. 2023; 24(22):16209. https://doi.org/10.3390/ijms242216209
Chicago/Turabian StyleCiaffaglione, Valeria, and Enrico Rizzarelli. 2023. "Carnosine, Zinc and Copper: A Menage a Trois in Bone and Cartilage Protection" International Journal of Molecular Sciences 24, no. 22: 16209. https://doi.org/10.3390/ijms242216209
APA StyleCiaffaglione, V., & Rizzarelli, E. (2023). Carnosine, Zinc and Copper: A Menage a Trois in Bone and Cartilage Protection. International Journal of Molecular Sciences, 24(22), 16209. https://doi.org/10.3390/ijms242216209