

Reactive Oxygen Species Mediate Transcriptional Responses to Dopamine and Cocaine in Human Cerebral Organoids


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Ventral Forebrain Organoids Exhibit Ca2+ and Intracellular cAMP Responses to Dopamine
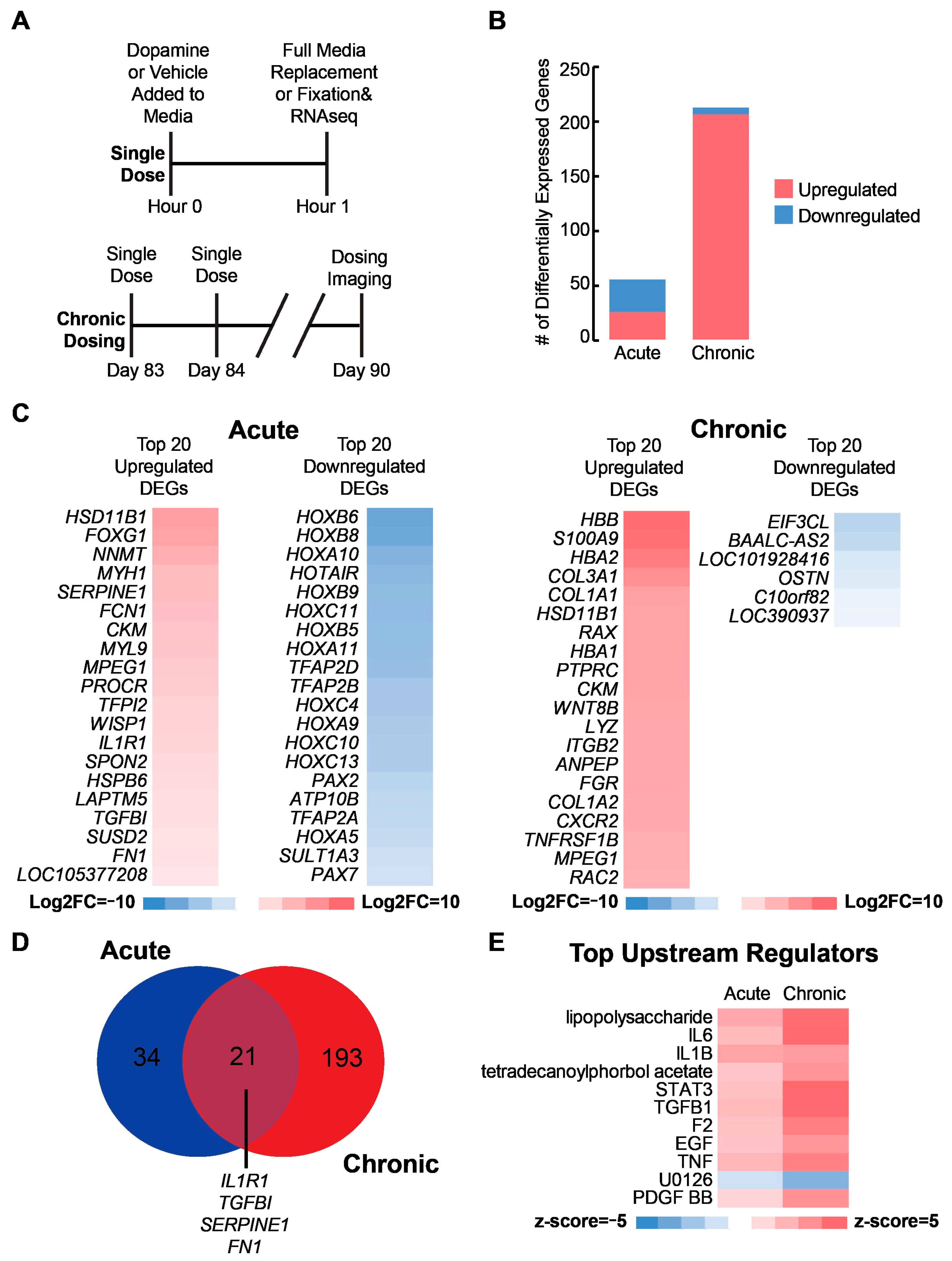
2.2. Transcriptional Analysis Reveals Immune-Related Response to Acute and Chronic Dopamine
2.3. Organoid Response to Dopamine Driven by Reactive Oxygen Species
2.4. ROS Also Mediates the Transcriptomic Response to Cocaine
3. Discussion
4. Materials and Methods
4.1. hESC Cell Lines
4.2. Organoid Culture
4.3. Cryosectioning and Immunohistochemistry
4.4. Real Time Quantitative PCR
4.5. Live Cell Imaging
4.6. Dosing Experiments
4.7. ROS Measurement
4.8. RNA Extraction and Sequencing Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinta, S.J.; Andersen, J.K. Dopaminergic neurons. Int. J. Biochem. Cell Biol. 2005, 37, 942–946. [Google Scholar] [CrossRef]
- Wise, R.A. Brain reward circuitry: Insights from unsensed incentives. Neuron 2002, 36, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Soares-Cunha, C.; Coimbra, B.; Sousa, N.; Rodrigues, A.J. Reappraising striatal D1- and D2-neurons in reward and aversion. Neurosci. Biobehav. Rev. 2016, 68, 370–386. [Google Scholar] [CrossRef] [PubMed]
- Buchsbaum, I.Y.; Cappello, S. Neuronal migration in the CNS during development and disease: Insights from in vivo and in vitro models. Development 2019, 146, dev163766. [Google Scholar] [CrossRef]
- Ohtani, N.; Goto, T.; Waeber, C.; Bhide, P.G. Dopamine modulates cell cycle in the lateral ganglionic eminence. J. Neurosci. 2003, 23, 2840–2850. [Google Scholar] [CrossRef] [PubMed]
- Hunt, C.P.J.; Pouton, C.W.; Haynes, J.M. Characterising the developmental profile of human embryonic stem cell-derived medium spiny neuron progenitors and assessing mature neuron function using a CRISPR-generated human DARPP-32WT/eGFP-AMP reporter line. Neurochem. Int. 2017, 106, 3–13. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Duffy, P. Time course of extracellular dopamine and behavioral sensitization to cocaine. I. Dopamine axon terminals. J. Neurosci. 2011, 102, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Hyman, S.E.; Malenka, R.C.; Nestler, E.J. Neural Mechanisms of Addiction: The Role of Reward-Related Learning and Memory. Annu. Rev. Neurosci. 2006, 29, 565–598. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.A.; Salery, M.; Scarpa, J.R.; Calipari, E.S.; Hamilton, P.J.; Ku, S.M.; Kronman, H.; Purushothaman, I.; Juarez, B.; Heshmati, M.; et al. Transcriptional and physiological adaptations in nucleus accumbens somatostatin interneurons that regulate behavioral responses to cocaine. Nat. Commun. 2018, 9, 3149. [Google Scholar] [CrossRef]
- Russo, S.J.; Dietz, D.M.; Dumitriu, D.; Malenka, R.C.; Nestler, E.J. The Addicted Synapse: Mechanisms of Synaptic and Structural Plasticity in Nucleus Accumbens. Trends Neurosci. 2011, 33, 267–276. [Google Scholar] [CrossRef]
- Esiri, M.M. Ageing and the brain. J. Pathol. 2007, 211, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Meiser, J.; Weindl, D.; Hiller, K. Complexity of dopamine metabolism. Cell Commun. Signal. 2013, 11, 34. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yang, F. The interplay of dopamine metabolism abnormalities and mitochondrial defects in the pathogenesis of schizophrenia. Transl. Psychiatry 2022, 12, 464. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J.; Shen, W.; Day, M.; Gertler, T.; Chan, S.; Tian, X.; Plotkin, J.L. The role of dopamine in modulating the structure and function of striatal circuits. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2010; Volume 183, pp. 148–167. [Google Scholar]
- Kippin, T.E.; Kapur, S.; Van Der Kooy, D. Dopamine specifically inhibits forebrain neural stem cell proliferation, suggesting a novel effect of antipsychotic drugs. J. Neurosci. 2005, 25, 5815–5823. [Google Scholar] [CrossRef]
- Gerfen, C.R.; Bolam, J.P. The Neuroanatomical Organization of the Basal Ganglia. Handb. Behav. Neurosci. 2010, 20, 3–28. [Google Scholar]
- Beauchamp, A.; Yee, Y.; Darwin, B.C.; Raznahan, A.; Mars, R.B.; Lerch, J.P. Whole-brain comparison of rodent and human brains using spatial transcriptomics. eLife 2022, 11, e79418. [Google Scholar] [CrossRef] [PubMed]
- Gokce, O.; Stanley, G.M.; Treutlein, B.; Neff, N.F.; Camp, J.G.; Malenka, R.C.; Rothwell, P.E.; Fuccillo, M.V.; Südhof, T.C.; Quake, S.R. Cellular Taxonomy of the Mouse Striatum as Revealed by Single-Cell RNA-Seq. Cell Rep. 2016, 16, 1126–1137. [Google Scholar] [CrossRef]
- Marshall, J.J.; Mason, J.O. Mouse vs man: Organoid models of brain development & disease. Brain Res. 2019, 1724, 146427. [Google Scholar]
- Cambray, S.; Arber, C.; Little, G.; Dougalis, A.G.; de Paola, V.; Ungless, M.A.; Li, M.; Rodríguez, T.A. Activin induces cortical interneuron identity and differentiation in embryonic stem cell-derived telencephalic neural precursors. Nat. Commun. 2012, 3, 841. [Google Scholar] [CrossRef]
- Arber, C.; Precious, S.V.; Cambray, S.; Risner-Janiczek, J.R.; Kelly, C.; Noakes, Z.; Fjodorova, M.; Heuer, A.; Ungless, M.A.; Rodríguez, T.A.; et al. Activin a directs striatal projection neuron differentiation of human pluripotent stem cells. Development 2015, 142, 1375–1386. [Google Scholar] [CrossRef]
- Nicoleau, C.; Varela, C.; Bonnefond, C.; Maury, Y.; Bugi, A.; Aubry, L.; Viegas, P.; Bourgois-Rocha, F.; Peschanski, M.; Perrier, A.L. Embryonic stem cells neural differentiation qualifies the role of Wnt/β-Catenin signals in human telencephalic specification and regionalization. Stem Cells 2013, 31, 1763–1774. [Google Scholar] [CrossRef] [PubMed]
- Fjodorova, M.; Noakes, Z.; Li, M. How to make striatal projection neurons. Neurogenesis 2015, 2, e1100227. [Google Scholar] [CrossRef] [PubMed]
- Fasano, C.; Bourque, M.-J.; Lapointe, G.; Leo, D.; Thibault, D.; Haber, M.; Kortleven, C.; DesGroseillers, L.; Murai, K.K.; Trudeau, L. Dopamine facilitates dendritic spine formation by cultured striatal medium spiny neurons through both D1 and D2 dopamine receptors. Neuropharmacology 2013, 67, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, D.; Bi, C.; Mi, T.; Zhu, W.; Xia, L.; Teng, Z.; Hu, B.; Wu, Y. A Chemical Recipe for Generation of Clinical-Grade Striatal Neurons from hESCs. Stem Cell Rep. 2018, 11, 635–650. [Google Scholar] [CrossRef] [PubMed]
- Tam, R.W.; Keung, A.J. Human Pluripotent Stem Cell-Derived Medium Spiny Neuron-like Cells Exhibit Gene Desensitization. Cells 2022, 11, 1411. [Google Scholar] [CrossRef]
- Bagley, J.A.; Reumann, D.; Bian, S.; Lévi-Strauss, J.; Knoblich, J.A. Fused cerebral organoids model interactions between brain regions. Nat. Methods 2017, 14, 743–751. [Google Scholar] [CrossRef]
- Xiang, Y.; Tanaka, Y.; Patterson, B.; Kang, Y.J.; Govindaiah, G.; Roselaar, N.; Cakir, B.; Kim, K.Y.; Lombroso, A.P.; Hwang, S.M.; et al. Fusion of Regionally Specified hPSC-Derived Organoids Models Human Brain Development and Interneuron Migration. Cell Stem Cell 2017, 21, 383–398.e7. [Google Scholar] [CrossRef]
- Kindberg, A.A.; Bendriem, R.M.; Spivak, C.E.; Chen, J.; Handreck, A.; Lupica, C.R.; Liu, J.; Freed, W.J.; Lee, C.T. An in vitro model of human neocortical development using pluripotent stem cells: Cocaine-induced cy-toarchitectural alterations. DMM Dis. Model. Mech. 2014, 7, 1397–1405. [Google Scholar]
- Yao, H.; Wu, W.; Cerf, I.; Zhao, H.W.; Wang, J.; Negraes, P.D.; Muotri, A.R.; Haddad, G.G. Methadone interrupts neural growth and function in human cortical organoids. Stem Cell Res. 2020, 49, 102065. [Google Scholar] [CrossRef]
- Dang, J.; Tiwari, S.K.; Agrawal, K.; Hui, H.; Qin, Y.; Rana, T.M. Glial cell diversity and methamphetamine-induced neuroin-flammation in human cerebral organoids. Mol. Psychiatry 2020, 26, 9–11. [Google Scholar]
- Lee, C.-T.; Chen, J.; Kindberg, A.A.; Bendriem, R.M.; Spivak, C.E.; Williams, M.P.; Richie, C.T.; Handreck, A.; Mallon, B.S.; Lupica, C.R.; et al. CYP3A5 Mediates Effects of Cocaine on Human Neocorticogenesis: Studies using an in Vitro 3D Self-Organized hPSC Model with a Single Cortex-Like Unit. Neuropsychopharmacology 2017, 42, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.M.; Gaj, T.; Rao, A.T.; Kulkarni, R.U.; Fuentes, C.M.; Ramadoss, G.N.; Ekman, F.K.; Miller, E.W.; Schaffer, D.V. hPSC-Derived Striatal Cells Generated Using a Scalable 3D Hydrogel Promote Recovery in a Huntington Disease Mouse Model. Stem Cell Rep. 2018, 10, 1481–1491. [Google Scholar] [CrossRef] [PubMed]
- Rudibaugh, T.; Tam, R.; Estridge, R.C.; Keung, A.J. Single cell assessment of human stem cell derived mesolimbic models and their responses to substances of abuse. bioRxiv 2023, preprint. [Google Scholar]
- Scimemi, A.; Beato, M. Determining the neurotransmitter concentration profile at active synapses. Mol. Neurobiol. 2009, 40, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Garris, P.A.; Ciolkowski, E.L.; Pastore, P.; Wightman, R.M. Efflux of dopamine from the synaptic cleft in the nucleus accumbens of the rat brain. J. Neurosci. 1994, 14, 6084–6093. [Google Scholar] [CrossRef]
- Liu, C.; Goel, P.; Kaeser, P.S. Spatial and temporal scales of dopamine transmission. Nat. Rev. Neurosci. 2021, 22, 345–358. [Google Scholar] [CrossRef]
- Martel, J.C.; McArthur, S.G. Dopamine Receptor Subtypes, Physiology and Pharmacology: New Ligands and Concepts in Schizophrenia. Front. Pharmacol. 2020, 11, 1003. [Google Scholar] [CrossRef]
- Al-muhtasib, N.; Forcelli, P.A.; Vicini, S. Differential electrophysiological properties of D1 and D2 spiny projection neurons in the mouse nucleus accumbens core. Physiol. Rep. 2018, 6, e13784. [Google Scholar] [CrossRef]
- Yapo, C.; Nair, A.G.; Clement, L.; Castro, L.R.; Kotaleski, J.H.; Vincent, P. Detection of phasic dopamine by D1 and D2 striatal medium spiny neurons. J. Physiol. 2017, 595, 7451–7475. [Google Scholar] [CrossRef]
- Matamales, M.; Girault, J.-A. Signaling from the Cytoplasm to the Nucleus in Striatal Medium-Sized Spiny Neurons. Front. Neuroanat. 2011, 5, 37. [Google Scholar] [CrossRef]
- Zaccolo, M. cAMP Signaling: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2015. [Google Scholar]
- Lee, H.; Jiang, X.; Perwaiz, I.; Yu, P.; Wang, J.; Wang, Y.; Hüttemann, M.; Felder, R.A.; Sibley, D.R.; Polster, B.M.; et al. Dopamine D5 receptor-mediated decreases in mitochondrial reactive oxygen species production are cAMP and autophagy dependent. Hypertens. Res. 2022, 44, 628–641. [Google Scholar] [CrossRef]
- Robison, A.J.; Nestler, E.J. Transcriptional and epigenetic mechanisms of addiction. Nat. Rev. Neurosci. 2011, 12, 623–637. [Google Scholar] [CrossRef]
- Carlezon, W.A.; Duman, R.S.; Nestler, E.J. The many faces of CREB. Trends Neurosci. 2005, 28, 436–445. [Google Scholar] [CrossRef]
- Asl, M.M.; Vahabie, A.H.; Valizadeh, A. Dopaminergic modulation of synaptic plasticity, its role in neuropsy-chiatric disorders, and its computational modeling. Basic Clin. Neurosci. 2019, 10, 1–12. [Google Scholar]
- Robinson, T.E.; Kolb, B. Persistent structural modifications in nucleus accumbens and prefrontal cortex neurons produced by previous experience with amphetamine. J. Neurosci. 1997, 17, 8491–8497. [Google Scholar] [CrossRef]
- Hyman, S.E. Addiction: A disease of learning and memory. Am. J. Psychiatry 2005, 162, 1414–1422. [Google Scholar] [CrossRef]
- Nestler, E.J. Molecular basis of long-term plasticity underlying addiction. Nat. Rev. Neurosci. 2001, 2, 119–128. [Google Scholar] [CrossRef]
- Tanaka, Y.; Cakir, B.; Xiang, Y.; Sullivan, G.J.; Park, I.H. Synthetic analyses of single-cell transcriptomes from multiple brain organoids and fetal brain. Cell Rep. 2020, 30, 1682–1689. [Google Scholar] [CrossRef]
- Luo, C.; Lancaster, M.A.; Castanon, R.; Nery, J.R.; Knoblich, J.A.; Ecker, J.R. Cerebral organoids recapitulate epigenomic sig-natures of the human fetal brain. Cell Rep. 2016, 17, 3369–3384. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Miura, M. Programmed cell death in neurodevelopment. Dev. Cell 2015, 32, 478–490. [Google Scholar] [CrossRef]
- Sánchez-Herrero, E. Hox Targets and Cellular Functions. Scientifica 2013, 2013, 738257. [Google Scholar] [CrossRef]
- Andrews, R.N.; Caudell, D.L.; Metheny-Barlow, L.J.; Peiffer, A.M.; Tooze, J.A.; Bourland, J.D.; Hampson, R.E.; Deadwyler, S.A.; Cline, J.M. Fibronectin Produced by Cerebral Endothelial and Vascular Smooth Muscle Cells Contributes to Peri-vascular Extracellular Matrix in Late-Delayed Radiation-Induced Brain Injury. Radiat. Res. 2018, 190, 361–373. [Google Scholar] [CrossRef]
- Palomo, J.; Dietrich, D.; Martin, P.; Palmer, G.; Gabay, C. The interleukin (IL)-1 cytokine family—Balance between agonists and antagonists in inflammatory diseases. Cytokine 2015, 76, 25–37. [Google Scholar] [CrossRef]
- Katayama, M.; Ota, K.; Nagi-Miura, N.; Ohno, N.; Yabuta, N.; Nojima, H.; Kumanogoh, A.; Hirano, T. Ficolin-1 is a promising therapeutic target for autoimmune diseases. Int. Immunol. 2018, 31, 23–32. [Google Scholar] [CrossRef]
- Simone, T.M.; Higgins, C.E.; Czekay, R.P.; Law, B.K.; Higgins, S.P.; Archambeault, J.; Kutz, S.M.; Higgins, P.J. SERPINE1: A Molecular Switch in the Proliferation-Migration Dichotomy in Wound-‘Activated’ Keratinocytes. Adv. Wound Care 2014, 3, 281–290. [Google Scholar] [CrossRef]
- Zhou, P.; Xiang, C.X.; Wei, J.F. The clinical significance of spondin 2 eccentric expression in peripheral blood mononuclear cells in bronchial asthma. J. Clin. Lab. Anal. 2021, 35, e23764. [Google Scholar] [CrossRef]
- Gancarz-Kausch, A.M.; Schroeder, G.L.; Panganiban, C.; Adank, D.; Humby, M.S.; Kausch, M.A.; Clark, S.D.; Dietz, D.M. Transforming growth factor beta receptor 1 is increased following abstinence from cocaine self-administration, but not cocaine sensitization. PLoS ONE 2013, 8, e83834. [Google Scholar] [CrossRef]
- Wrana, J.L. Signaling by the TGFβ superfamily. Cold Spring Harb. Perspect. Biol. 2013, 5, a011197. [Google Scholar] [CrossRef]
- Yee, N.; Hamerman, J. β2 integrins inhibit TLR responses by regulating NF-κB pathway and p38 MAPK activation. Eur. J. Immunol. 2014, 43, 779–792. [Google Scholar] [CrossRef]
- Balanis, N.; Carlin, C. Stress-induced EGF Receptor Signaling through STAT3 and Tumor Progression in Triple-Negative Breast Cancer. Mol. Cell Endocrinol. 2017, 176, 24–30. [Google Scholar] [CrossRef]
- Liu, Y.N.; Peng, Y.L.; Liu, L.; Wu, T.Y.; Zhang, Y.; Lian, Y.J.; Yang, Y.Y.; Kelley, K.W.; Jiang, C.L.; Wang, Y.X. TNFα mediates stress-induced depression by upregulating indoleamine 2,3-dioxygenase in a mouse model of unpredictable chronic mild stress. Eur. Cytokine Netw. 2015, 26, 15–25. [Google Scholar] [CrossRef]
- Dainichi, T.; Kabashima, K.; Ivanov, I.I.; Goto, Y. Regulation of Immunity by Non-Immune Cells. Front. Immunol. 2021, 12, 770847. [Google Scholar] [CrossRef]
- Assis-De-Lemos, G.; Monteiro, J.; Oliveira-Valença, V.M.; Melo, G.A.; Reis, R.A.D.M.; Rehen, S.K.; Silveira, M.S.; Galina, A. Dopamine signaling impairs ROS modulation by mitochondrial hexokinase in human neural progenitor cells. Biosci. Rep. 2021, 41, BSR20211191. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Takeshima, Y.; Fujio, K. Basic mechanism of immune system activation by mitochondria. Immunol. Med. 2020, 43, 142–147. [Google Scholar] [CrossRef]
- Martins, S.G.; Zilhão, R.; Thorsteinsdóttir, S.; Carlos, A.R. Linking Oxidative Stress and DNA Damage to Changes in the Ex-pression of Extracellular Matrix Components. Front. Genet. 2021, 12, 673002. [Google Scholar] [CrossRef]
- David, O.; Barrera, I.; Gould, N.; Gal-Ben-Ari, S.; Rosenblum, K. D1 Dopamine Receptor Activation Induces Neuronal eEF2 Pathway-Dependent Protein Synthesis. Front. Mol. Neurosci. 2020, 13, 67. [Google Scholar] [CrossRef]
- Guo, M.-L.; Liao, K.; Periyasamy, P.; Yang, L.; Cai, Y.; Callen, S.E.; Buch, S. Cocaine-mediated microglial activation involves the ER stress-autophagy axis. Autophagy 2015, 11, 995–1009. [Google Scholar] [CrossRef]
- Lee, C.-T.; Chen, J.; Hayashi, T.; Tsai, S.-Y.; Sanchez, J.F.; Errico, S.L.; Amable, R.; Su, T.-P.; Lowe, R.H.; Huestis, M.A.; et al. A mechanism for the inhibition of neural progenitor cell proliferation by cocaine. PLoS Med. 2008, 5, 0987–1004. [Google Scholar] [CrossRef]
- Chong, W.C.; Shastri, M.D.; Eri, R. Endoplasmic Reticulum Stress and Oxidative Stress: A Vicious Nexus Implicated in Bowel Disease Pathophysiology. Int. J. Mol. Sci. 2017, 18, 771. [Google Scholar] [CrossRef]
- Lidow, M.S.; Bozian, D.; Song, Z.M. Cocaine affects cerebral neocortical cytoarchitecture in primates only if administered during neocortical neuronogenesis. Dev. Brain Res. 2001, 128, 45–52. [Google Scholar] [CrossRef]
- Ares-Santos, S.; Granado, N.; Moratalla, R. The role of dopamine receptors in the neurotoxicity of methamphetamine. J. Intern. Med. 2013, 273, 437–453. [Google Scholar] [CrossRef]
- Dimberg, J.; Ström, K.; Löfgren, S.; Zar, N.; Hugander, A.; Matussek, A. Expression of the serine protease inhibitor serpinA3 in human colorectal adenocarcinomas. Oncol. Lett. 2011, 2, 413–418. [Google Scholar] [CrossRef]
- Olguín, H.J.; Guzmán, D.C.; García, E.H.; Mejía, G.B. The role of dopamine and its dysfunction as a consequence of oxidative stress. Oxid. Med. Cell. Longev. 2016, 2016, 9730467. [Google Scholar]
- Money, K.M.; Stanwood, G.D. Developmental origins of brain disorders: Roles for dopamine. Front. Cell. Neurosci. 2013, 7, 260. [Google Scholar] [CrossRef]
- Park, D.S.; Kozaki, T.; Tiwari, S.K.; Moreira, M.; Khalilnezhad, A.; Torta, F.; Olivié, N.; Thiam, C.H.; Liani, O.; Silvin, A.; et al. iPS-cell-derived microglia promote brain organoid maturation via cholesterol transfer. Nature 2023, 623, 397–405. [Google Scholar] [CrossRef]
- McCarthy, D.M.; Zhang, X.; Darnell, S.B.; Sangrey, G.R.; Yanagawa, Y.; Sadri-Vakili, G.; Bhide, P.G. Cocaine alters BDNF expression and neuronal migration in the embryonic mouse forebrain. J. Neurosci. 2011, 31, 13400–13411. [Google Scholar] [CrossRef]
- Glatt, S.J.; Bolaños, C.A.; Trksak, G.H.; Jackson, D. Effects of prenatal cocaine exposure on dopamine system development: A meta-analysis. Neurotoxicol. Teratol. 2000, 22, 617–629. [Google Scholar] [CrossRef]
- D’Sa, C.; Fox, H.C.; Hong, A.K.; Dileone, R.J.; Sinha, R. Increased serum brain-derived neurotrophic factor is predictive of cocaine relapse outcomes: A prospective study. Biol. Psychiatry 2011, 70, 706–711. [Google Scholar] [CrossRef]
- McCarthy, D.M.; Mueller, K.A.; Cannon, E.N.; Huizenga, M.N.; Darnell, S.B.; Bhide, P.G.; Sadri-Vakili, G. Prenatal Cocaine Exposure Alters BDNF-TrkB Signaling in the Embryonic and Adult Brain. Dev. Neurosci. 2017, 38, 365–374. [Google Scholar] [CrossRef]
- Motz, K.; Lina, I.; Murphy, M.K.; Drake, V.; Davis, R.; Tsai, H.; Feeley, M.; Yin, L.X.; Ding, D.; Hillel, A. Alexander Hillel, M2 Macrophages Promote Collagen Expression and Synthesis in Laryngotracheal Stenosis Fibroblasts. Laryngoscope 2021, 131, 678–687. [Google Scholar] [CrossRef]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Channer, B.; Matt, S.M.; Nickoloff-Bybel, E.A.; Pappa, V.; Agarwal, Y.; Wickman, J.; Gaskill, P.J. Dopamine, Immunity, and Disease. Pharmacol. Rev. 2023, 75, 62–158. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Kelava, I.; Lancaster, M.A. Stem Cell Models of Human Brain Development. Cell Stem Cell 2016, 18, 736–748. [Google Scholar] [CrossRef]
- Koo, B.; Choi, B.; Park, H.; Yoon, K.J. Past, Present, and Future of Brain Organoid Technology. Mol. Cells 2019, 42, 617–627. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudibaugh, T.T.; Stuppy, S.R.; Keung, A.J. Reactive Oxygen Species Mediate Transcriptional Responses to Dopamine and Cocaine in Human Cerebral Organoids. Int. J. Mol. Sci. 2023, 24, 16474. https://doi.org/10.3390/ijms242216474
Rudibaugh TT, Stuppy SR, Keung AJ. Reactive Oxygen Species Mediate Transcriptional Responses to Dopamine and Cocaine in Human Cerebral Organoids. International Journal of Molecular Sciences. 2023; 24(22):16474. https://doi.org/10.3390/ijms242216474
Chicago/Turabian StyleRudibaugh, Thomas T., Samantha R. Stuppy, and Albert J. Keung. 2023. "Reactive Oxygen Species Mediate Transcriptional Responses to Dopamine and Cocaine in Human Cerebral Organoids" International Journal of Molecular Sciences 24, no. 22: 16474. https://doi.org/10.3390/ijms242216474
APA StyleRudibaugh, T. T., Stuppy, S. R., & Keung, A. J. (2023). Reactive Oxygen Species Mediate Transcriptional Responses to Dopamine and Cocaine in Human Cerebral Organoids. International Journal of Molecular Sciences, 24(22), 16474. https://doi.org/10.3390/ijms242216474