

The sprT Gene of Bacillus velezensis FZB42 Is Involved in Biofilm Formation and Bacilysin Production



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Analysis
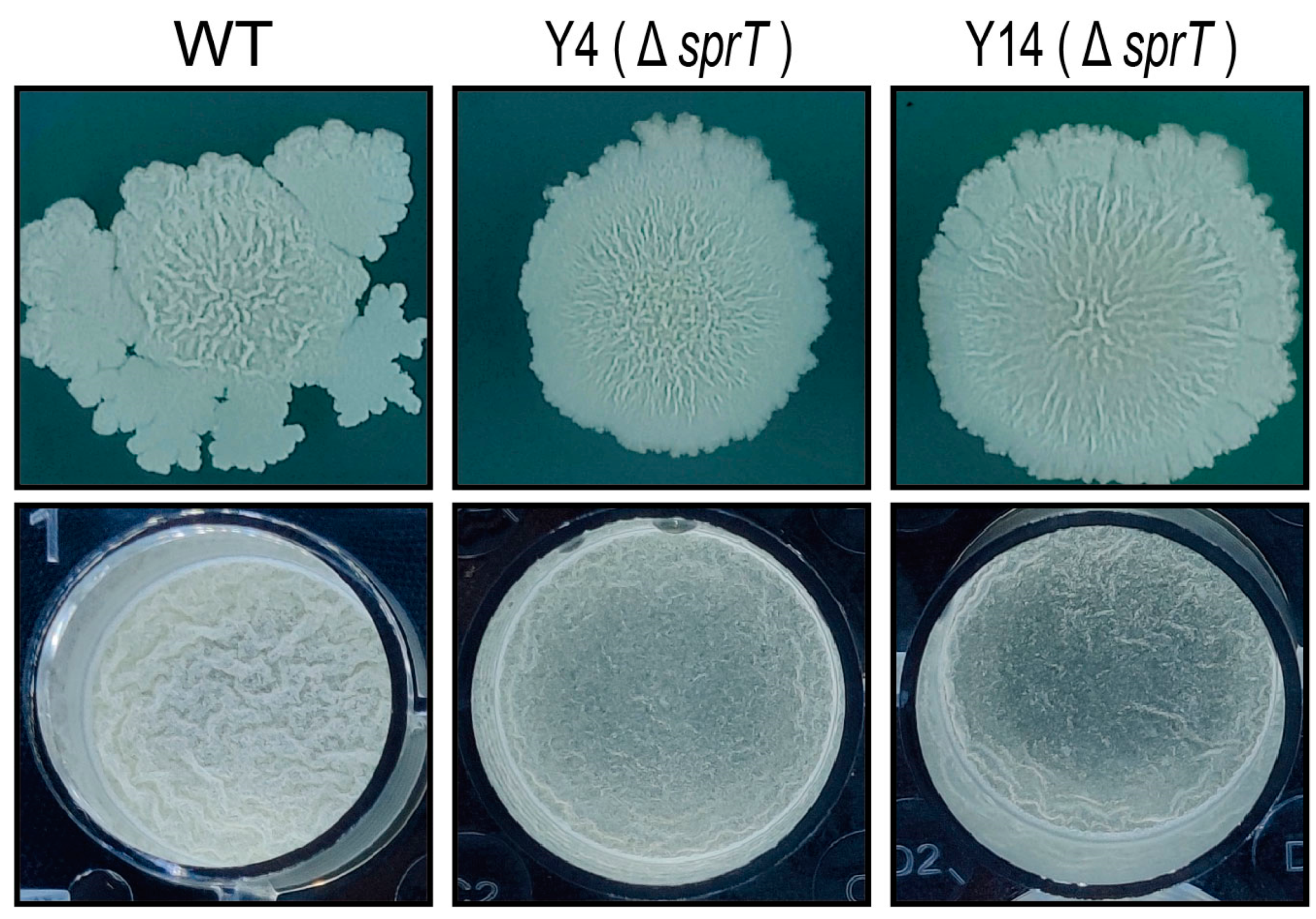
2.1. sprT Is Involved in the Biofilm Formation of B. velezensis FZB42
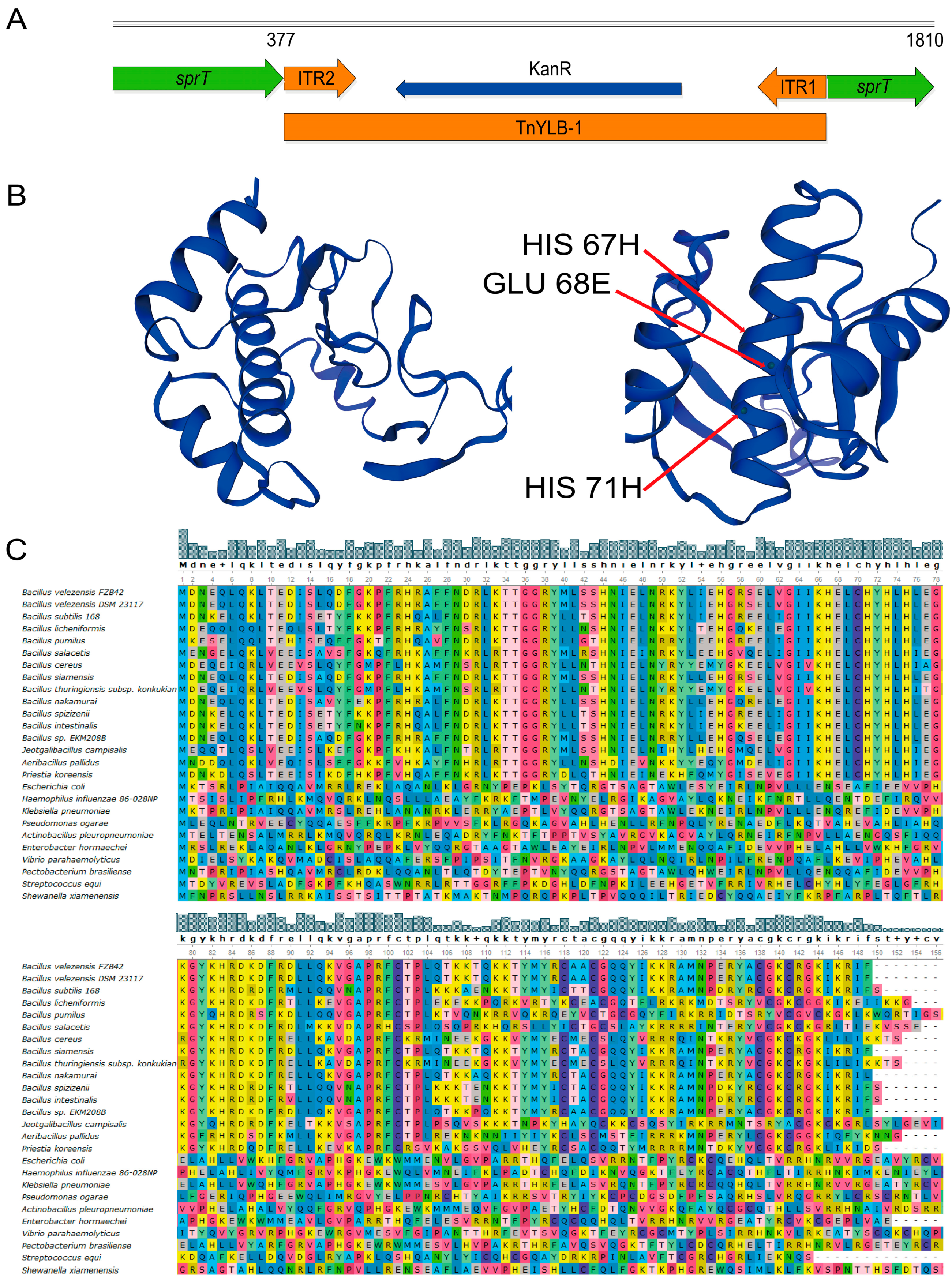
2.2. Bioinformatic Analysis Shows sprT Is Conserved among Bacillus Species
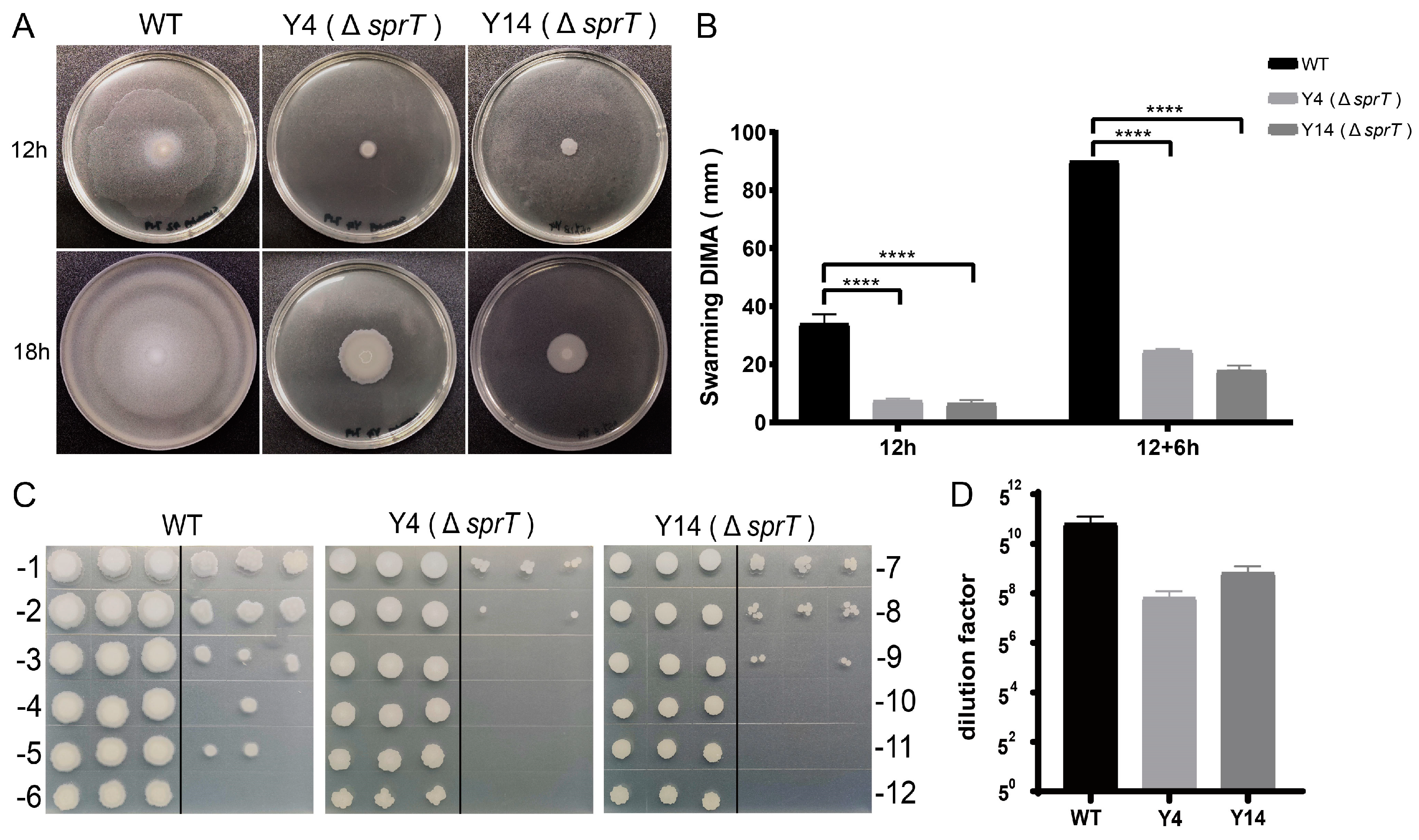
2.3. sprT Deletion Resulted in a Defect in Swarming Ability and Sporulation of FZB42
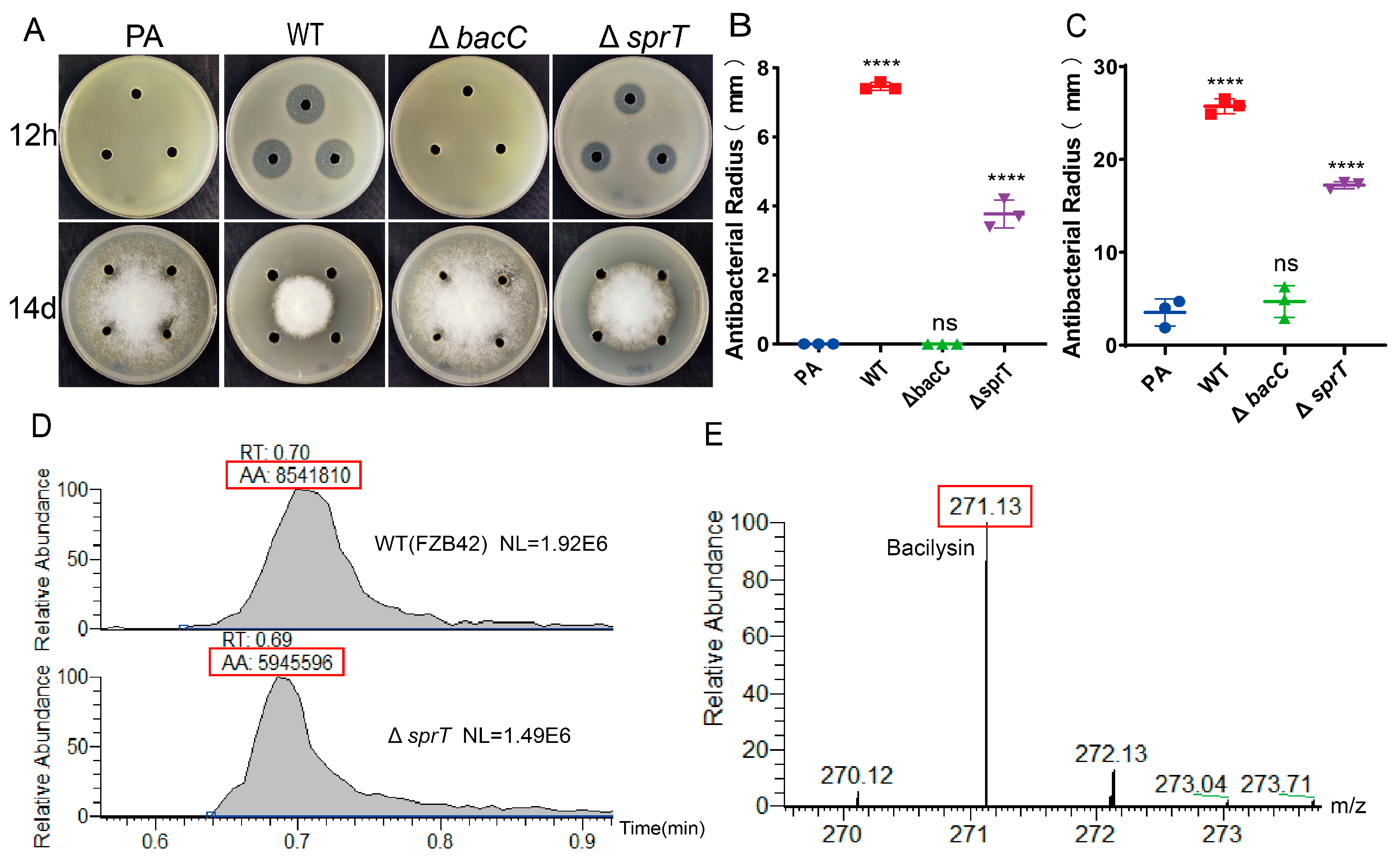
2.4. sprT Deletion Reduces Production of the Antibiotic Bacilysin
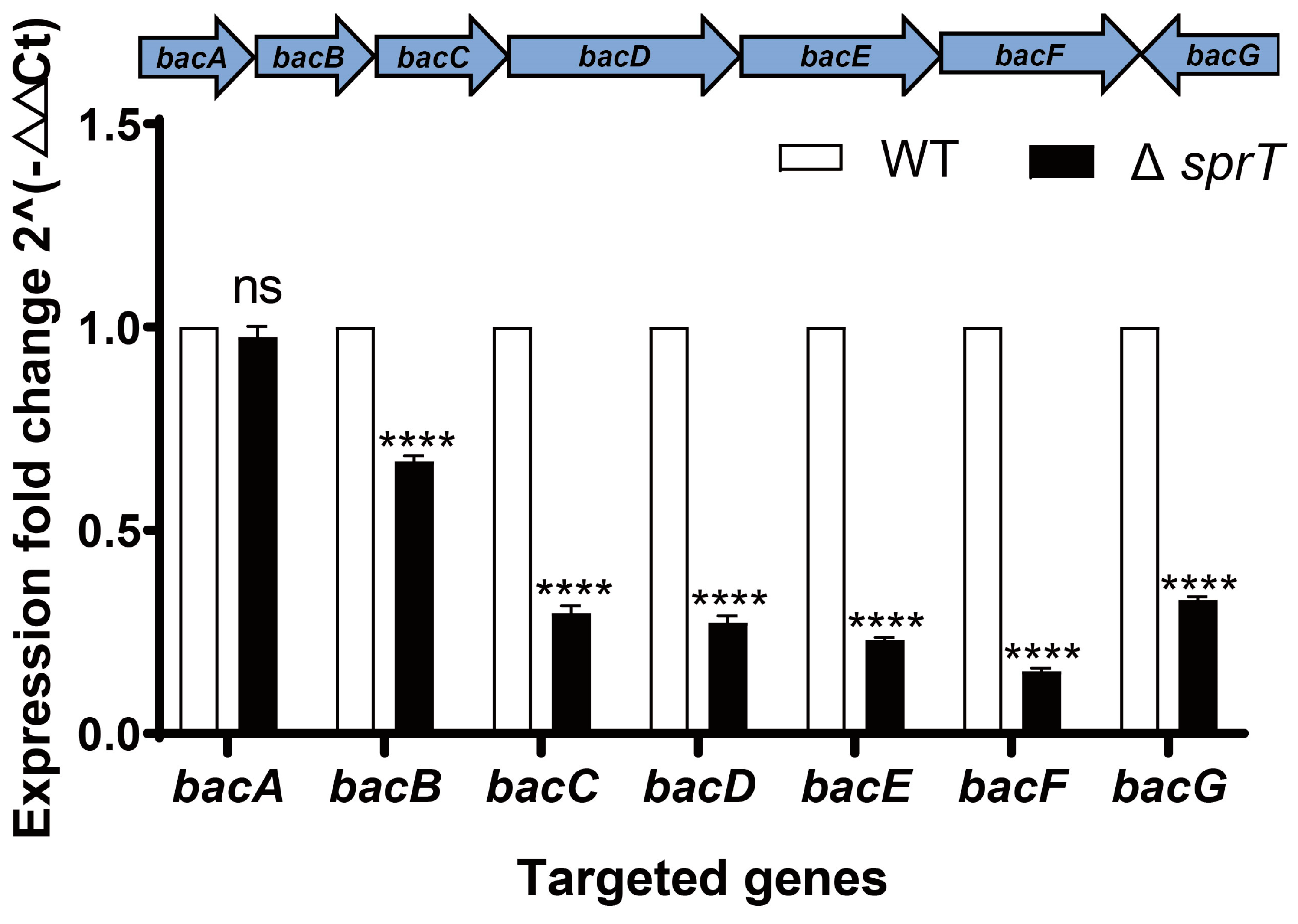
2.5. Transcription of Bac Operon Is Down-Regulated via ΔsprT Deletion
3. Discussion
4. Materials and Methods
4.1. Strains, Growth Conditions, and Primers
4.2. Strain Construction
4.3. Transposon TnYLB-1 Insertion Site Mapping
4.4. Biofilm Formation Test
4.5. Bioinformatics Analysis
4.6. Swarming Test
4.7. Sporulation Test
4.8. Antagonistic Test of Antibiotic Bacilysin
4.9. HPLC-MS
4.10. Real-Time Quantitative PCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singla, J.; Krattinger, S.G. Biotic Stress Resistance Genes in Wheat. In The Production and Genetics of Food Grains; Elsevier: Amsterdam, The Netherlands, 2016; pp. 388–392. [Google Scholar]
- Han, X.; Shen, D.; Xiong, Q.; Bao, B.; Zhang, W.; Dai, T.; Zhao, Y.; Borriss, R.; Fan, B. The Plant-Beneficial Rhizobacterium Bacillus velezensis FZB42 Controls the Soybean Pathogen Phytophthora sojae Due to Bacilysin Production. Appl. Environ. Microbiol. 2021, 87, e0160121. [Google Scholar] [CrossRef]
- Ricci, P.; Bonnet, P.; Huet, J.C.; Sallantin, M.; Beauvais-Cante, F.; Bruneteau, M.; Billard, V.; Michel, G.; Pernollet, J.C. Structure and activity of proteins from pathogenic fungi Phytophthora eliciting necrosis and acquired resistance in tobacco. Eur. J. Biochem. 1989, 183, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.P.; Uhl, J.; Grosch, R.; Alquéres, S.; Pittroff, S.; Dietel, K.; Schmitt-Kopplin, P.; Borriss, R.; Hartmann, A. Cyclic Lipopeptides of Bacillus amyloliquefaciens subsp. plantarum Colonizing the Lettuce Rhizosphere Enhance Plant Defense Responses Toward the Bottom Rot Pathogen Rhizoctonia solani. Mol. Plant-Microbe Interact. 2015, 28, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Borriss, R.; Bleiss, W.; Wu, X. Gram-positive rhizobacterium Bacillus amyloliquefaciens FZB42 colonizes three types of plants in different patterns. J. Microbiol. 2012, 50, 38–44. [Google Scholar] [CrossRef]
- Liu, S.; Tian, Y.; Jia, M.; Lu, X.; Yue, L.; Zhao, X.; Jin, W.; Wang, Y.; Zhang, Y.; Xie, Z.; et al. Induction of Salt Tolerance in Arabidopsis thaliana by Volatiles from Bacillus amyloliquefaciens FZB42 via the Jasmonic Acid Signaling Pathway. Front Microbiol. 2020, 11, 562934. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O.; et al. Comparative analysis of the complete genome sequence of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, J.Y.; Kim, S.H.; Bae, H.J.; Yi, H.; Yoon, S.H.; Koo, B.S.; Kwon, M.; Cho, J.Y.; Lee, C.E.; et al. Surfactin from Bacillus subtilis displays anti-proliferative effect via apoptosis induction, cell cycle arrest and survival signaling suppression. FEBS Lett. 2007, 581, 865–871. [Google Scholar] [CrossRef]
- Doornbos, R.F.; Van Loon, L.C.; Bakker, P.A.H.M. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. A review. Agron. Sustain. Dev. 2011, 32, 227–243. [Google Scholar] [CrossRef]
- Cao, X.; Li, Y.; Fan, J.; Zhao, Y.; Borriss, R.; Fan, B. Two Lysine Sites That Can Be Malonylated Are Important for LuxS Regulatory Roles in Bacillus velezensis. Microorganisms 2021, 9, 1338. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef]
- Schneider, K.; Chen, X.H.; Vater, J.; Franke, P.; Nicholson, G.; Borriss, R.; Süssmuth, R.D. Macrolactin is the polyketide biosynthesis product of the pks2 cluster of Bacillus amyloliquefaciens FZB42. J. Nat. Prod. 2007, 70, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, C. Fengycins, Cyclic Lipopeptides from Marine Bacillus subtilis Strains, Kill the Plant-Pathogenic Fungus Magnaporthe grisea by Inducing Reactive Oxygen Species Production and Chromatin Condensation. Appl. Environ. Microbiol. 2018, 84, e00445-18. [Google Scholar] [CrossRef]
- Chen, X.H.; Vater, J.; Piel, J.; Franke, P.; Scholz, R.; Schneider, K.; Koumoutsi, A.; Hitzeroth, G.; Grammel, N.; Strittmatter, A.W.; et al. Structural and functional characterization of three polyketide synthase gene clusters in Bacillus amyloliquefaciens FZB 42. J. Bacteriol. 2006, 188, 4024–4036. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Schneider, K.; Vater, J.; Süssmuth, R.; Piel, J.; Borriss, R. Genome analysis of Bacillus amyloliquefaciens FZB42 reveals its potential for biocontrol of plant pathogens. J. Biotechnol. 2009, 140, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Borriss, R.; Chen, X.H.; Rueckert, C.; Blom, J.; Becker, A.; Baumgarth, B.; Fan, B.; Pukall, R.; Schumann, P.; Spröer, C.; et al. Relationship of Bacillus amyloliquefaciens clades associated with strains DSM 7T and FZB42T: A proposal for Bacillus amyloliquefaciens subsp. amyloliquefaciens subsp. nov. and Bacillus amyloliquefaciens subsp. plantarum subsp. nov. based on complete genome sequence comparisons. Int. J. Syst. Evol. Microbiol. 2011, 61, 1786–1801. [Google Scholar]
- Wu, L.; Wu, H.; Chen, L.; Yu, X.; Borriss, R.; Gao, X. Difficidin and bacilysin from Bacillus amyloliquefaciens FZB42 have antibacterial activity against Xanthomonas oryzae rice pathogens. Sci. Rep. 2015, 5, 12975. [Google Scholar] [CrossRef]
- Wang, T.; Liu, X.H.; Wu, M.B.; Ge, S. Molecular insights into the antifungal mechanism of bacilysin. J. Mol. Model. 2018, 24, 118. [Google Scholar] [CrossRef]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef]
- Talukdar, P.K.; Olguín-Araneda, V.; Alnoman, M.; Paredes-Sabja, D.; Sarker, M.R. Updates on the sporulation process in species. Res. Microbiol. 2015, 166, 225–235. [Google Scholar] [CrossRef]
- Budiharjo, A.; Chowdhury, S.P.; Dietel, K.; Beator, B.; Dolgova, O.; Fan, B.; Bleiss, W.; Ziegler, J.; Schmid, M.; Hartmann, A.; et al. Transposon Mutagenesis of the Plant-Associated Bacillus amyloliquefaciens ssp. plantarum FZB42 Revealed That the nfrA and RBAM17410 Genes Are Involved in Plant-Microbe-Interactions. PLoS ONE 2014, 9, e98267. [Google Scholar] [CrossRef]
- Garrity, L.F.; Ordal, G.W. Chemotaxis in Bacillus subtilis: How bacteria monitor environmental signals. Pharmacol. Ther. 1995, 68, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus velezensis FZB42 in 2018: The Gram-Positive Model Strain for Plant Growth Promotion and Biocontrol. Front. Microbiol. 2018, 9, 2491. [Google Scholar] [CrossRef] [PubMed]
- Ghoreishi, F.S.; Roghanian, R.; Emtiazi, G. Inhibition of quorum sensing-controlled virulence factors with natural substances and novel protease, obtained from Halobacillus karajensis. Microb. Pathog. 2020, 149, 104555. [Google Scholar] [CrossRef]
- Yokoi, K.J.; Kuzuwa, S.; Iwasaki, S.; Yamakawa, A.; Taketo, A.; Kodaira, K. Aureolysin of Staphylococcus warneri M accelerates its proteolytic cascade, and participates in biofilm formation. Biosci. Biotechnol. Biochem. 2016, 80, 1238–1242. [Google Scholar] [CrossRef] [PubMed]
- Van Wolferen, M.; Orell, A.; Albers, S.V. Archaeal biofilm formation. Nat. Rev. Microbiol. 2018, 16, 699–713. [Google Scholar] [CrossRef]
- Verhamme, D.T.; Kiley, T.B.; Stanley-Wall, N.R. DegU co-ordinates multicellular behaviour exhibited by Bacillus subtilis. Mol. Microbiol. 2007, 65, 554–568. [Google Scholar] [CrossRef]
- Schacherl, M.; Pichlo, C.; Neundorf, I.; Baumann, U. Structural Basis of Proline-Proline Peptide Bond Specificity of the Metalloprotease Zmp1 Implicated in Motility of Clostridium difficile. Structure 2015, 23, 1632–1642. [Google Scholar] [CrossRef]
- Shankweiler, G.W.Z. Primary Subsite Mapping and Carbohydrate Analysis of the Serine Protease, Bacillopeptidase F, from Bacillus subttlis. Ph.D. Dissertation, New Mexico State University, Las Cruces, NM, USA, 1986. [Google Scholar]
- Vishnoi, M.; Narula, J.; Devi, S.N.; Dao, H.A.; Igoshin, O.A.; Fujita, M. Triggering sporulation in Bacillus subtilis with artificial two-component systems reveals the importance of proper Spo0A activation dynamics. Mol. Microbiol. 2013, 90, 181–194. [Google Scholar] [CrossRef]
- Grandvalet, C.; Gominet, M.; Lereclus, D. Identification of genes involved in the activation of the Bacillus thuringiensis inhA metalloprotease gene at the onset of sporulation. Microbiology 2001, 147, 1805–1813. [Google Scholar] [CrossRef]
- Kenig, M.; Abraham, E.P. Antimicrobial activities and antagonists of bacilysin and anticapsin. Microbiology 1976, 94, 37–45. [Google Scholar] [CrossRef]
- Wu, J.W.; Chen, X.L. Extracellular metalloproteases from bacteria. Appl. Microbiol. Biotechnol. 2011, 92, 253–262. [Google Scholar] [CrossRef]
- Diethmaier, C.; Chawla, R.; Canzoneri, A.; Kearns, D.B.; Lele, P.P.; Dubnau, D. Viscous drag on the flagellum activates Bacillus subtilis entry into the K-state. Mol. Microbiol. 2017, 106, 367–380. [Google Scholar] [CrossRef]
- Mariappan, A.; Makarewicz, O.; Chen, X.H.; Borriss, R. Two-component response regulator DegU controls the expression of bacilysin in plant-growth-promoting bacterium Bacillus amyloliquefaciens FZB42. J. Mol. Microbiol. Biotechnol. 2012, 22, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Kanvinde, L.; Sastry, G.R. Agrobacterium tumefaciens Is a Diazotrophic Bacterium. Appl. Environ. Microbiol. 1990, 56, 2087–2092. [Google Scholar] [CrossRef] [PubMed]
- Perry, D.; Abraham, E.P. Transport and metabolism of bacilysin and other peptides by suspensions of Staphylococcus aureus. J. Gen. Microbiol. 1979, 115, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Alves, S.H.; Linares, C.E.; Loreto, E.S.; Rodrigues, M.; Thomazi, D.I.; Souza, F.; Santurio, J.M. Utilization of tomato juice agar (V8 agar) in the presumptive identification of Candida dubliniensis. Rev. Da Soc. Bras. De Med. Trop. 2006, 39, 92–93. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Chen, R.; Rahman, M.U.; Wei, C.; Fan, B. The sprT Gene of Bacillus velezensis FZB42 Is Involved in Biofilm Formation and Bacilysin Production. Int. J. Mol. Sci. 2023, 24, 16815. https://doi.org/10.3390/ijms242316815
Yang Y, Chen R, Rahman MU, Wei C, Fan B. The sprT Gene of Bacillus velezensis FZB42 Is Involved in Biofilm Formation and Bacilysin Production. International Journal of Molecular Sciences. 2023; 24(23):16815. https://doi.org/10.3390/ijms242316815
Chicago/Turabian StyleYang, Yihan, Ruofu Chen, Mati Ur Rahman, Chunyue Wei, and Ben Fan. 2023. "The sprT Gene of Bacillus velezensis FZB42 Is Involved in Biofilm Formation and Bacilysin Production" International Journal of Molecular Sciences 24, no. 23: 16815. https://doi.org/10.3390/ijms242316815
APA StyleYang, Y., Chen, R., Rahman, M. U., Wei, C., & Fan, B. (2023). The sprT Gene of Bacillus velezensis FZB42 Is Involved in Biofilm Formation and Bacilysin Production. International Journal of Molecular Sciences, 24(23), 16815. https://doi.org/10.3390/ijms242316815