

Comparative RNA-Seq Analysis Revealed Tissue-Specific Splicing Variations during the Generation of the PDX Model

 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
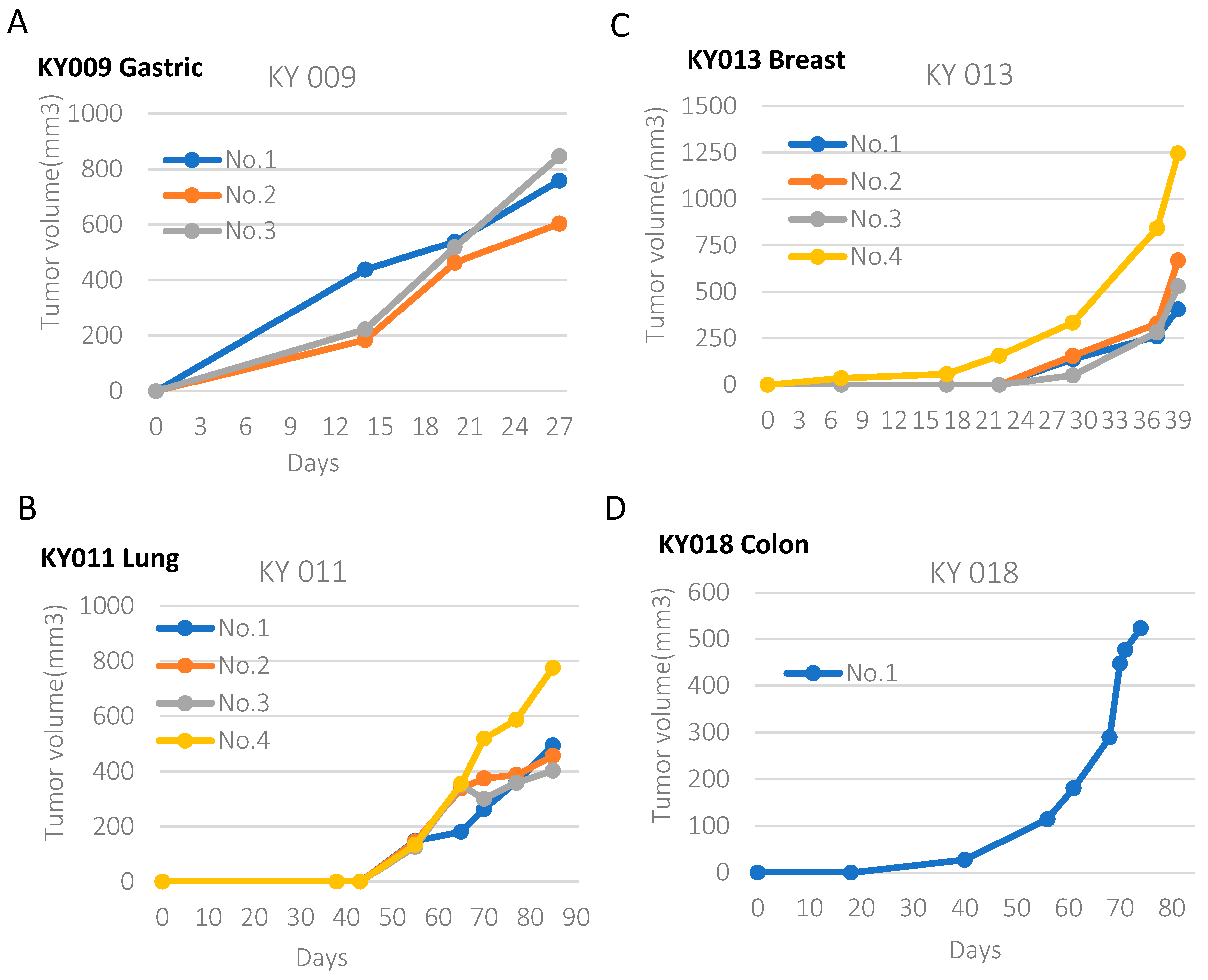
2.1. Generation of PDX Model from Various Types of Primary Tumors
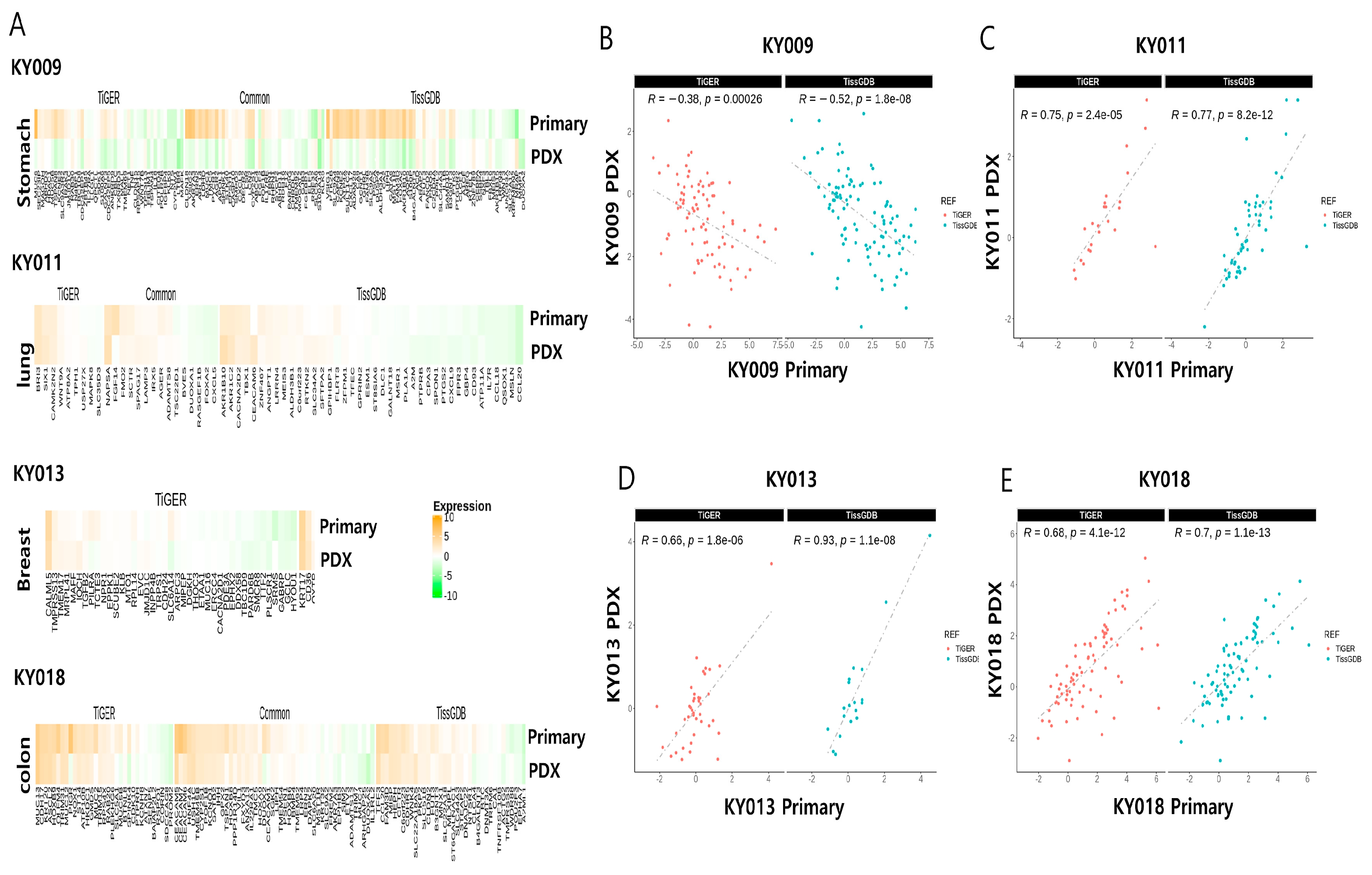
2.2. RNA-Seq Analysis of Four Primary−PDX Pairs Shows Conserved RNA Biotype Distribution with Significant Correlation for the Expression of Tissue-Specific Genes
2.3. RNA-Seq Analysis of Four Primary−PDX Pairs Revealed a Large Number of Splicing Variations in a Tumor-Type-Specific Manner
2.4. Validation of the Skipped Exon Pattern for Each Primary−PDX Pair by RT-PCR
3. Materials and Methods
3.1. Generation of Patient-Derived Xenograft (PDX) Models
3.2. RNA-Seq Analysis
3.3. PCR-Gel Electrophoresis for Validation of the Splicing Events
3.4. Statistical Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jung, J.; Seol, H.S.; Chang, S. The Generation and Application of Patient-Derived Xenograft Model for Cancer Research. Cancer Res. Treat. 2018, 50, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Jang, K.; Ju, J.M.; Lee, E.; Lee, J.W.; Kim, H.J.; Kim, J.; Lee, S.B.; Ko, B.S.; Son, B.H.; et al. Novel cancer gene variants and gene fusions of triple-negative breast cancers (TNBCs) reveal their molecular diversity conserved in the patient-derived xenograft (PDX) model. Cancer Lett. 2018, 428, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Seol, H.S.; Kang, H.J.; Lee, S.I.; Kim, N.E.; Kim, T.I.; Chun, S.M.; Kim, T.W.; Yu, C.S.; Suh, Y.A.; Singh, S.R.; et al. Development and characterization of a colon PDX model that reproduces drug responsiveness and the mutation profiles of its original tumor. Cancer Lett. 2014, 345, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Chen, S.; Guo, Y.; Li, W.; Qi, X.; Yang, H.; Xiao, S.; Fang, G.; Hu, J.; Wen, C.; et al. Establishment and evaluation of four different types of patient-derived xenograft models. Cancer Cell Int. 2017, 17, 122. [Google Scholar] [CrossRef]
- Kawashima, N.; Ishikawa, Y.; Kim, J.H.; Ushijima, Y.; Akashi, A.; Yamaguchi, Y.; Hattori, H.; Nakashima, M.; Ikeno, S.; Kihara, R.; et al. Comparison of clonal architecture between primary and immunodeficient mouse-engrafted acute myeloid leukemia cells. Nat. Commun. 2022, 13, 1624. [Google Scholar] [CrossRef] [PubMed]
- Patient, R.K. Control of gene expression: Tissue-specific expression. Curr. Opin. Biotechnol. 1990, 1, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Biamonti, G.; Infantino, L.; Gaglio, D.; Amato, A. An Intricate Connection between Alternative Splicing and Phenotypic Plasticity in Development and Cancer. Cells 2019, 9, 34. [Google Scholar] [CrossRef]
- Lyu, J.; Cheng, C. Regulation of Alternative Splicing during Epithelial-Mesenchymal Transition. Cells Tissues Organs 2022, 211, 238–251. [Google Scholar] [CrossRef]
- Siegfried, Z.; Karni, R. The role of alternative splicing in cancer drug resistance. Curr. Opin. Genet. Dev. 2018, 48, 16–21. [Google Scholar] [CrossRef]
- Park, J.; Park, J.; Chung, Y.J. Alternative splicing: A new breakthrough for understanding tumorigenesis and potential clinical applications. Genes Genom. 2023, 45, 393–400. [Google Scholar] [CrossRef]
- Bradley, R.K.; Anczukow, O. RNA splicing dysregulation and the hallmarks of cancer. Nat. Rev. Cancer 2023, 23, 135–155. [Google Scholar] [CrossRef] [PubMed]
- Ruggles, K.V.; Tang, Z.; Wang, X.; Grover, H.; Askenazi, M.; Teubl, J.; Cao, S.; McLellan, M.D.; Clauser, K.R.; Tabb, D.L.; et al. An Analysis of the Sensitivity of Proteogenomic Mapping of Somatic Mutations and Novel Splicing Events in Cancer. Mol. Cell Proteom. 2016, 15, 1060–1071. [Google Scholar] [CrossRef] [PubMed]
- Kahles, A.; Lehmann, K.V.; Toussaint, N.C.; Huser, M.; Stark, S.G.; Sachsenberg, T.; Stegle, O.; Kohlbacher, O.; Sander, C.; Cancer Genome Atlas Research Network; et al. Comprehensive Analysis of Alternative Splicing Across Tumors from 8,705 Patients. Cancer Cell 2018, 34, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yao, X.; Zhou, H.; Wu, X.; Tian, J.; Zeng, J.; Yan, L.; Duan, C.; Liu, H.; Li, H.; et al. OncoSplicing: An updated database for clinically relevant alternative splicing in 33 human cancers. Nucleic Acids Res. 2022, 50, D1340–D1347. [Google Scholar] [CrossRef]
- Park, J.; Lee, J.O.; Lee, M.; Chung, Y.J. AS-CMC: A pan-cancer database of alternative splicing for molecular classification of cancer. Sci. Rep. 2022, 12, 21074. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Xiao, L.; Zhou, W.; Jin, X.; Xu, Z.; Xu, C.; Wang, P.; Luo, M.; Wang, M.; Ma, K.; et al. A pan-cancer analysis of alternative splicing of splicing factors in 6904 patients. Oncogene 2021, 40, 5441–5450. [Google Scholar] [CrossRef]
- Tarazona, S.; Garcia-Alcalde, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential expression in RNA-seq: A matter of depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Liu, X.; Yu, X.; Zack, D.J.; Zhu, H.; Qian, J. TiGER: A database for tissue-specific gene expression and regulation. BMC Bioinform. 2008, 9, 271. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.; Park, A.; Han, G.; Sun, H.; Jia, P.; Zhao, Z. TissGDB: Tissue-specific gene database in cancer. Nucleic Acids Res. 2018, 46, D1031–D1038. [Google Scholar] [CrossRef]
- Shen, S.; Park, J.W.; Lu, Z.X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. rMATS: Robust and flexible detection of differential alternative splicing from replicate RNA-Seq data. Proc. Natl. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef] [PubMed]
- Epifano, C.; Megias, D.; Perez-Moreno, M. p120-catenin differentially regulates cell migration by Rho-dependent intracellular and secreted signals. EMBO Rep. 2014, 15, 592–600. [Google Scholar] [CrossRef]
- Warzecha, C.C.; Shen, S.; Xing, Y.; Carstens, R.P. The epithelial splicing factors ESRP1 and ESRP2 positively and negatively regulate diverse types of alternative splicing events. RNA Biol. 2009, 6, 546–562. [Google Scholar] [CrossRef]
- Lee, J.; Pang, K.; Kim, J.; Hong, E.; Lee, J.; Cho, H.J.; Park, J.; Son, M.; Park, S.; Lee, M.; et al. ESRP1-regulated isoform switching of LRRFIP2 determines metastasis of gastric cancer. Nat. Commun. 2022, 13, 6274. [Google Scholar] [CrossRef]
- Banky, B.; Raso-Barnett, L.; Barbai, T.; Timar, J.; Becsagh, P.; Raso, E. Characteristics of CD44 alternative splice pattern in the course of human colorectal adenocarcinoma progression. Mol. Cancer 2012, 11, 83. [Google Scholar] [CrossRef]
- Yae, T.; Tsuchihashi, K.; Ishimoto, T.; Motohara, T.; Yoshikawa, M.; Yoshida, G.J.; Wada, T.; Masuko, T.; Mogushi, K.; Tanaka, H.; et al. Alternative splicing of CD44 mRNA by ESRP1 enhances lung colonization of metastatic cancer cell. Nat. Commun. 2012, 3, 883. [Google Scholar] [CrossRef] [PubMed]
- Dembowski, J.A.; An, P.; Scoulos-Hanson, M.; Yeo, G.; Han, J.; Fu, X.D.; Grabowski, P.J. Alternative Splicing of a Novel Inducible Exon Diversifies the CASK Guanylate Kinase Domain. J. Nucleic Acids 2012, 2012, 816237. [Google Scholar] [CrossRef] [PubMed]
- Matamala, N.; Martinez, M.T.; Lara, B.; Perez, L.; Vazquez, I.; Jimenez, A.; Barquin, M.; Ferrarotti, I.; Blanco, I.; Janciauskiene, S.; et al. Alternative transcripts of the SERPINA1 gene in alpha-1 antitrypsin deficiency. J. Transl. Med. 2015, 13, 211. [Google Scholar] [CrossRef]
- Kwon, C.H.; Park, H.J.; Choi, J.H.; Lee, J.R.; Kim, H.K.; Jo, H.J.; Kim, H.S.; Oh, N.; Song, G.A.; Park, D.Y. Snail and serpinA1 promote tumor progression and predict prognosis in colorectal cancer. Oncotarget 2015, 6, 20312–20326. [Google Scholar] [CrossRef]
- Chan, H.J.; Li, H.; Liu, Z.; Yuan, Y.C.; Mortimer, J.; Chen, S. SERPINA1 is a direct estrogen receptor target gene and a predictor of survival in breast cancer patients. Oncotarget 2015, 6, 25815–25827. [Google Scholar] [CrossRef]
- Koralewski, T.E.; Krutovsky, K.V. Evolution of exon-intron structure and alternative splicing. PLoS ONE 2011, 6, e18055. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Gu, L.; Li, Y.; Zhao, R.; Jian, H.; Xie, W.; Liu, L.; Wu, H.; Ren, F.; Han, Y.; et al. Integrative genomic analysis of drug resistance in MET exon 14 skipping lung cancer using patient-derived xenograft models. Front. Oncol. 2022, 12, 1024818. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Cao, S.; Mashl, R.J.; Mo, C.K.; Zaccaria, S.; Wendl, M.C.; Davies, S.R.; Bailey, M.H.; Primeau, T.M.; Hoog, J.; et al. Comprehensive characterization of 536 patient-derived xenograft models prioritizes candidates for targeted treatment. Nat. Commun. 2021, 12, 5086. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.J.; Noh, S.-J.; Choi, H.; Kim, M.W.; Kim, S.J.; Seo, Y.A.; Jeong, J.E.; Shin, I.; Kim, J.-S.; Choi, J.-K.; et al. Comparative RNA-Seq Analysis Revealed Tissue-Specific Splicing Variations during the Generation of the PDX Model. Int. J. Mol. Sci. 2023, 24, 17001. https://doi.org/10.3390/ijms242317001
Lee EJ, Noh S-J, Choi H, Kim MW, Kim SJ, Seo YA, Jeong JE, Shin I, Kim J-S, Choi J-K, et al. Comparative RNA-Seq Analysis Revealed Tissue-Specific Splicing Variations during the Generation of the PDX Model. International Journal of Molecular Sciences. 2023; 24(23):17001. https://doi.org/10.3390/ijms242317001
Chicago/Turabian StyleLee, Eun Ji, Seung-Jae Noh, Huiseon Choi, Min Woo Kim, Su Jin Kim, Yeon Ah Seo, Ji Eun Jeong, Inkyung Shin, Jong-Seok Kim, Jong-Kwon Choi, and et al. 2023. "Comparative RNA-Seq Analysis Revealed Tissue-Specific Splicing Variations during the Generation of the PDX Model" International Journal of Molecular Sciences 24, no. 23: 17001. https://doi.org/10.3390/ijms242317001
APA StyleLee, E. J., Noh, S. -J., Choi, H., Kim, M. W., Kim, S. J., Seo, Y. A., Jeong, J. E., Shin, I., Kim, J. -S., Choi, J. -K., Cho, D. -Y., & Chang, S. (2023). Comparative RNA-Seq Analysis Revealed Tissue-Specific Splicing Variations during the Generation of the PDX Model. International Journal of Molecular Sciences, 24(23), 17001. https://doi.org/10.3390/ijms242317001