
Impact of Mephedrone on Fear Memory in Adolescent Rats: Involvement of Matrix Metalloproteinase-9 (MMP-9) and N-Methyl-D-aspartate (NMDA) Receptor
 ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
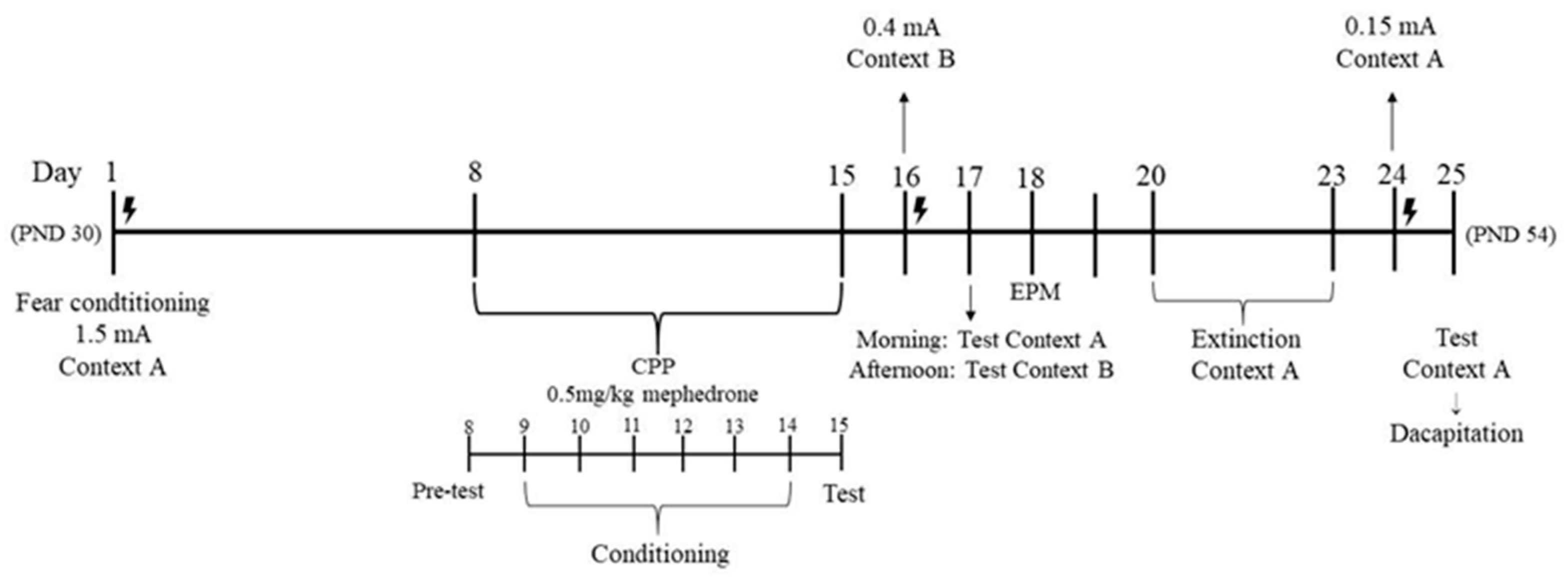
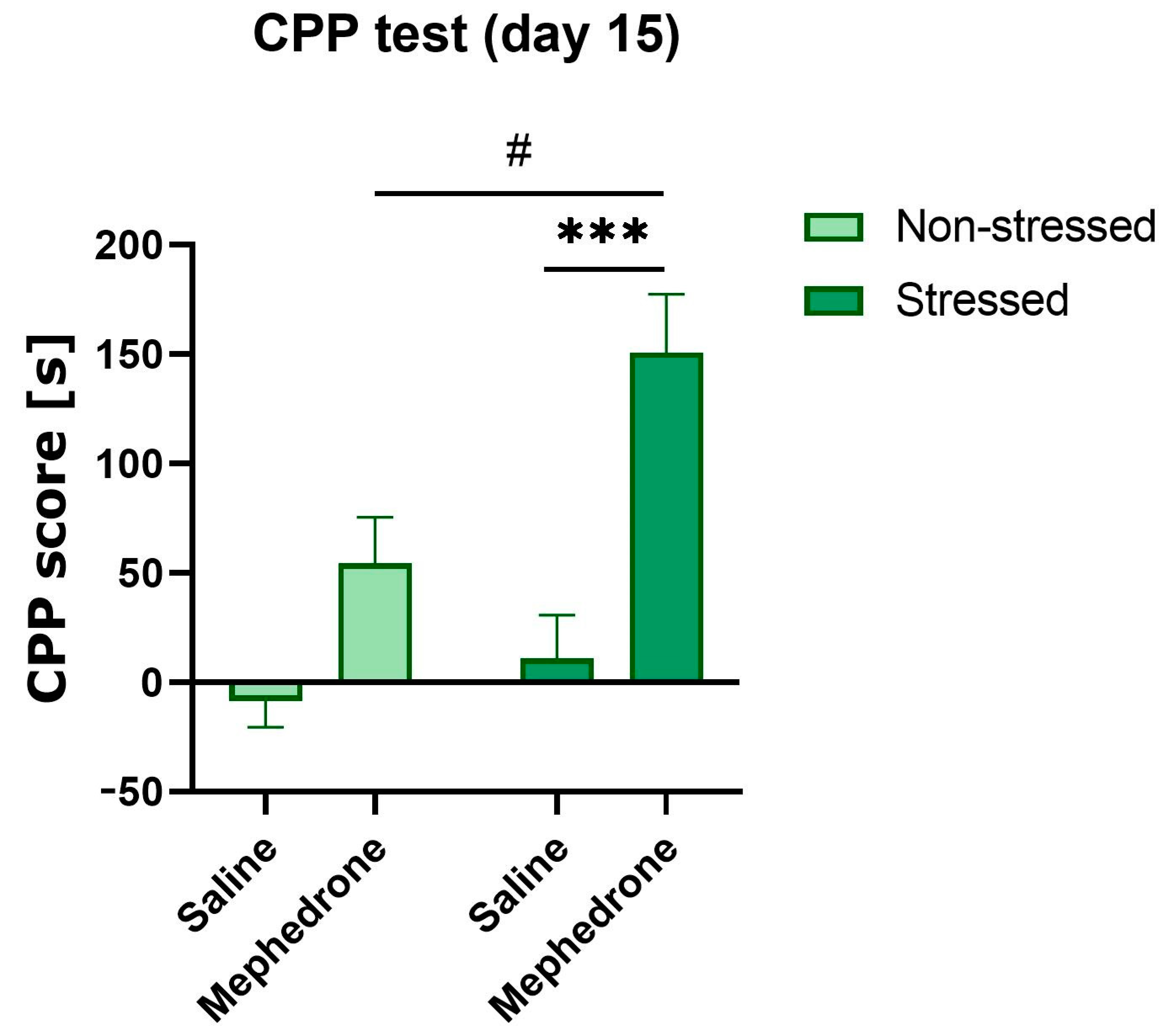
2.1. Stage 1: Impact of Footshock Stress on the Acquisition of Mephedrone-Induced CPP in Adolescent Rats
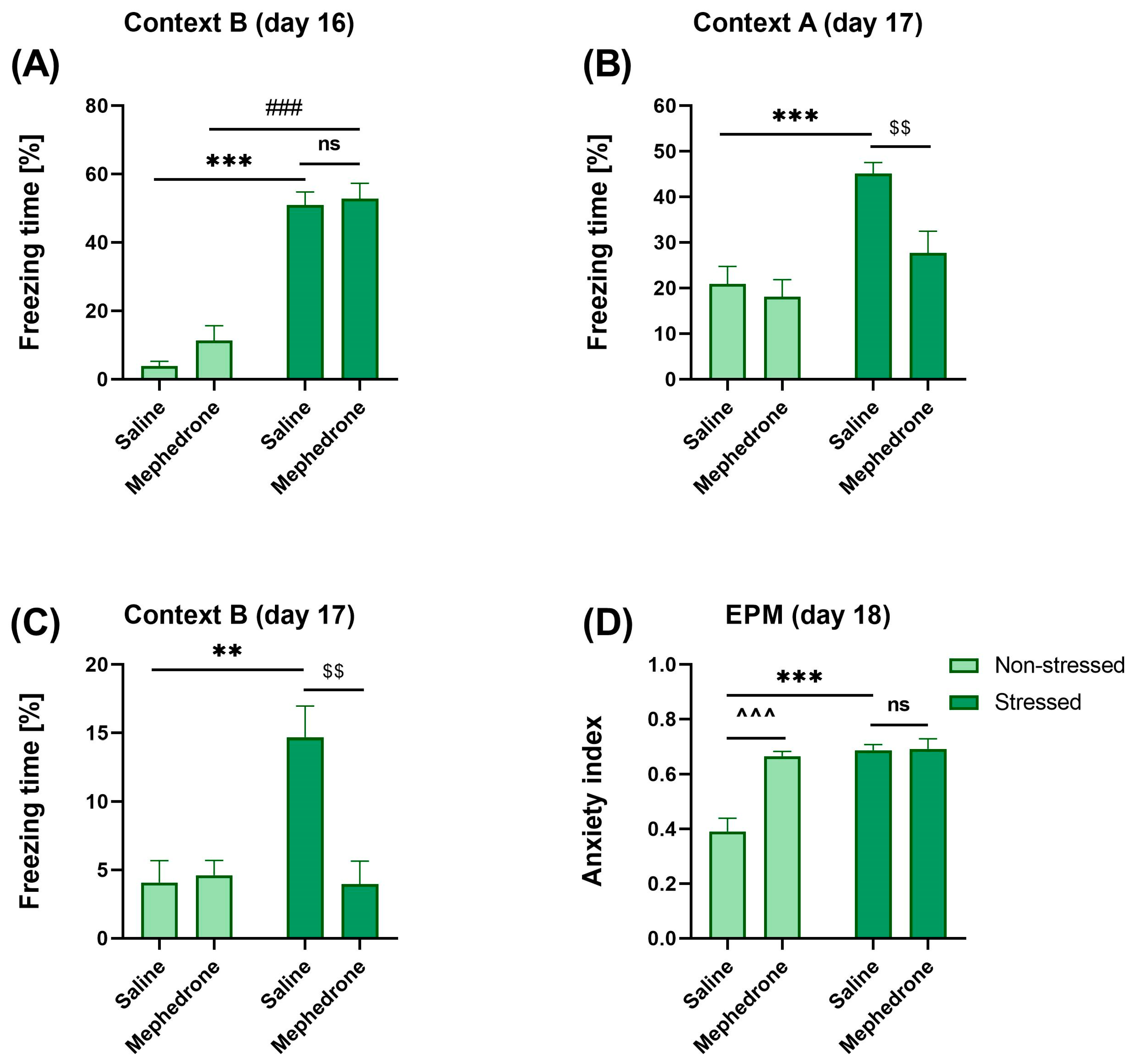
2.2. Stage 2: Impact of Repeated Mephedrone Injections on Fear Sensitization in Adolescent Rats. Anxiety-like Behavior Based on an Elevated Plus-Maze Test
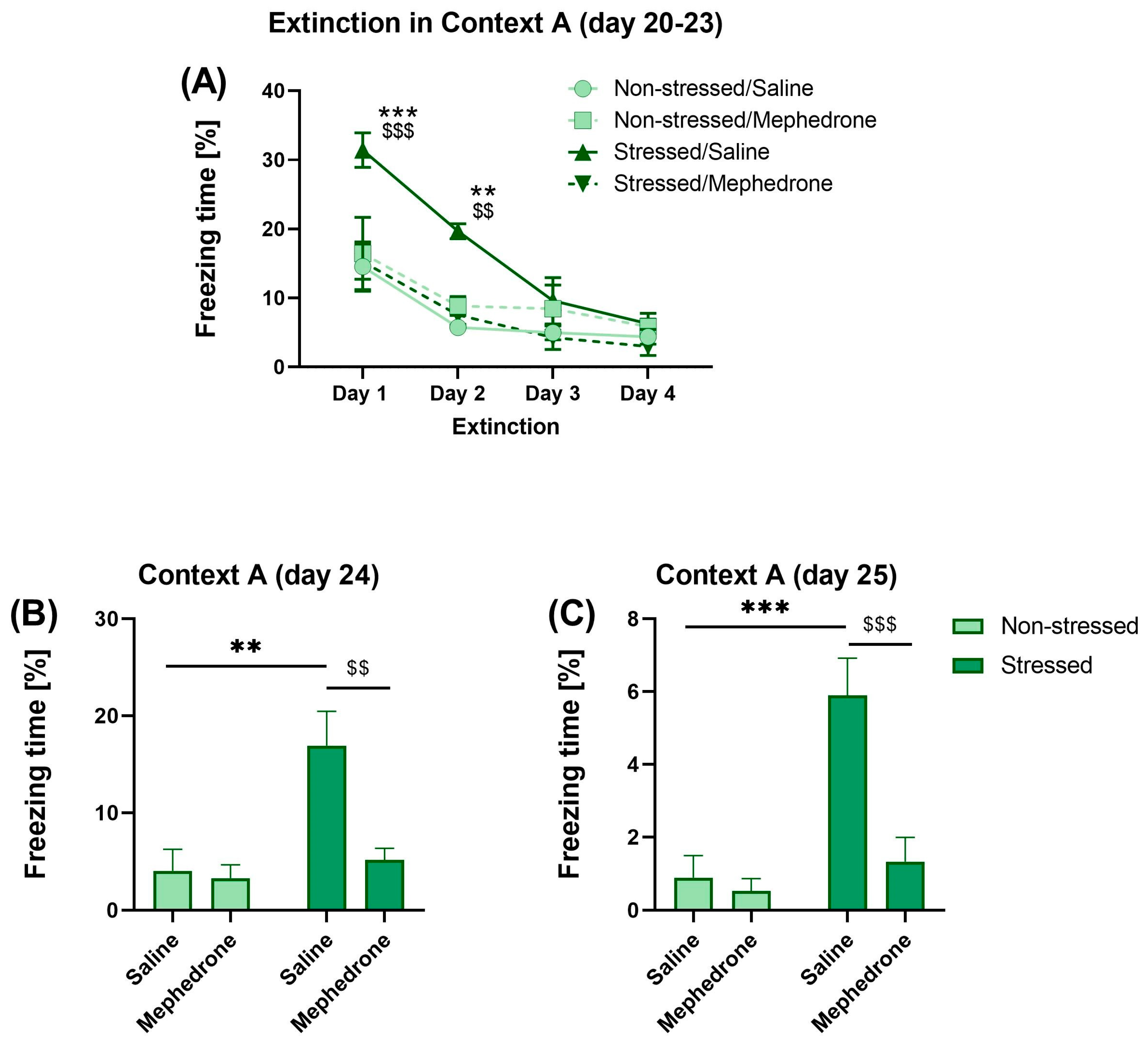
2.3. Stage 3: Impact of Repeated Mephedrone Injections on Extinction, Recall, and Retention of Fear Memory
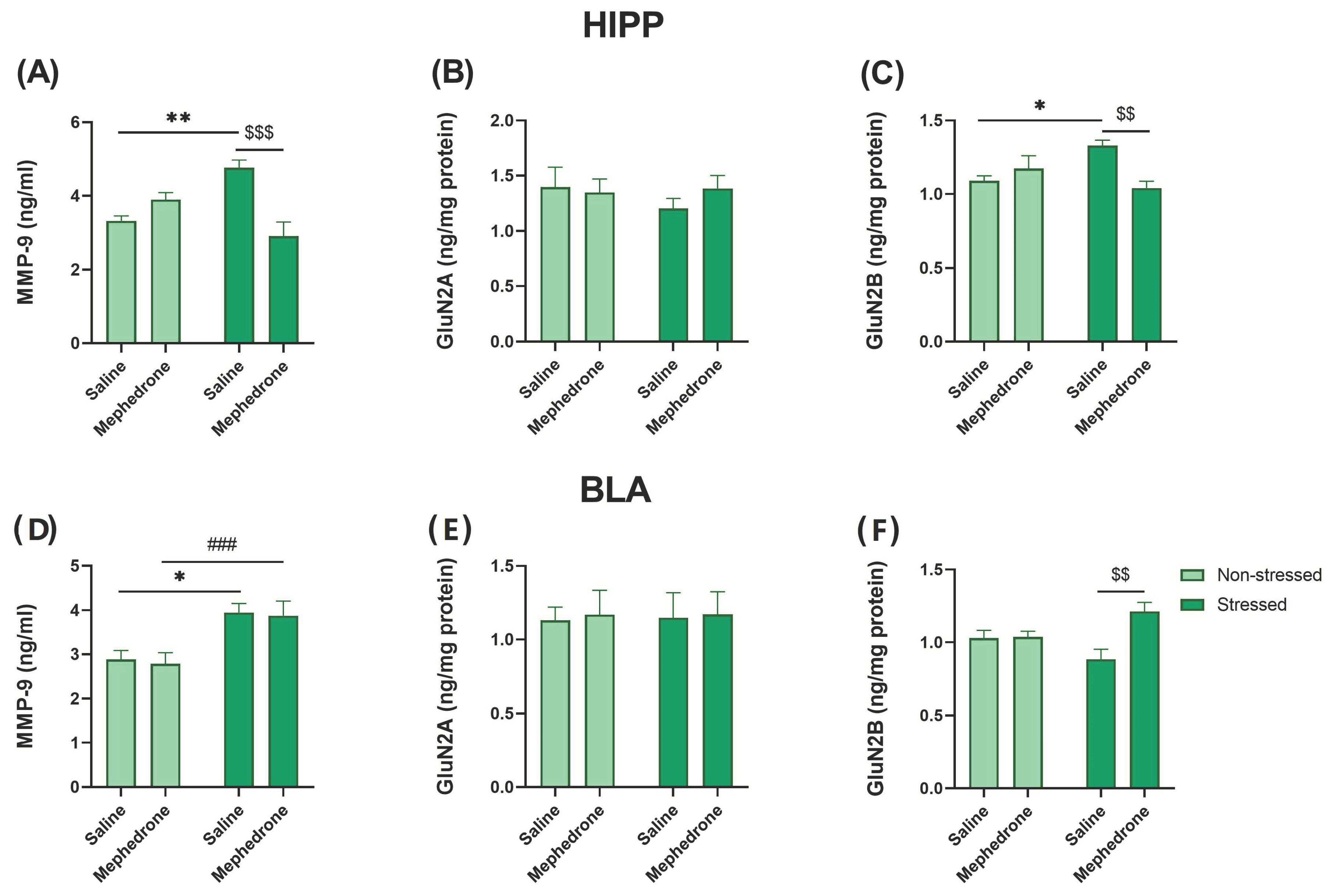
2.4. ELISA Assays: Impact of Footshock Stress and Repeated Mephedrone Injections on the Expression of NMDA Receptor Subunits (GluN2A and GluN2B) and MMP-9 in the HIPP and BLA
3. Discussion
3.1. Acquisition of Mephedrone-Induced CPP in Rats with Footshock Stress
3.2. Impact of Repeated Mephedrone Exposure on Fear Sensitization and Anxiety-like Behavior
3.3. Fear Extinction
3.4. Effects of Post-Stress Repeated Mephedrone Exposure on MPP-9, as Well as GluN2A and GluN2B Subunits of NMDA Receptor Expression in Trauma-Susceptible Rats
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Behavioral Apparatus and Procedures
4.3.1. Fear Conditioning
4.3.2. CPP Paradigm
4.3.3. Elevated Plus Maze (EPM)
4.3.4. ELISA Assays
4.4. Procedures
4.4.1. Stage 1: Impact of Footshock Stress on the Acquisition of Mephedrone-Induced CPP in Adolescent Rats
4.4.2. Stage 2: Impact of Repeated Mephedrone Injections on Fear Sensitization in Adolescent Rats. Anxiety-like Behavior Based on an EPM Test
4.4.3. Stage 3: Impact of Repeated Mephedrone Injections on Recall and Retention of Fear Memory in Adolescent Rats
4.5. Behavioral Scoring and Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bisson, J.I.; Cosgrove, S.; Lewis, C.; Robert, N.P. Post-traumatic stress disorder. BMJ 2015, 351, h6161. [Google Scholar] [CrossRef]
- Shalev, A.Y. Posttraumatic stress disorder and stress-related disorders. Psychiatr. Clin. N. Am. 2009, 32, 687–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, L.K.; Rattel, J.A.; Miedl, S.F.; Danböck, S.K.; Bürkner, P.C.; Wilhelm, F.H. Intrusive memories as conditioned responses to trauma cues: An empirically supported concept? Behav. Res. Ther. 2021, 143, 103848. [Google Scholar] [CrossRef]
- Wessa, M.; Flor, H. Failure of extinction of fear responses in posttraumatic stress disorder: Evidence from second-order conditioning. Am. J. Psychiatry 2007, 164, 1684–1692. [Google Scholar] [CrossRef] [Green Version]
- Seidemann, R.; Duek, O.; Jia, R.; Levy, I.; Harpaz-Rotem, I. The Reward System and Post-Traumatic Stress Disorder: Does Trauma Affect the Way We Interact with Positive Stimuli? Chronic Stress 2021, 5, 2470547021996006. [Google Scholar] [CrossRef]
- Guina, J.; Rossetter, S.R.; De Rhodes, B.J.; Nahhas, R.W.; Welton, R.S. Benzodiazepines for PTSD: A systematic review and meta-analysis. J. Psychiatr. Pract. 2015, 21, 281–303. [Google Scholar] [CrossRef]
- Tipps, M.E.; Raybuck, J.D.; Buck, K.J.; Lattal, K.M. Delay and trace fear conditioning in C57BL/6 and DBA/2 mice: Issues of measurement and performance. Learn. Mem. 2014, 21, 380–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basedow, L.A.; Kuitunen-Paul, S.; Wiedmann, M.F.; Roessner, V.; Golub, Y. Self-reported PTSD is associated with increased use of MDMA in adolescents with substance use disorders. Eur. J. Psychotraumatol. 2021, 12, 1968140. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.M.; Bogenschutz, M.; Lilienstein, A.; Harrison, C.; Kleiman, S.; Parker-Guilbert, K.; Ot’alora, G.M.; Garas, W.; Paleos, C.; Gorman, I.; et al. MDMA-assisted therapy for severe PTSD: A randomized, double-blind, placebo-controlled phase 3 study. Nat. Med. 2021, 27, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Brady, K.T.; Sinha, R. Co-occurring mental and substance use disorders: The neurobiological effects of chronic stress. Am. J. Psychiatry 2005, 162, 1483–1493. [Google Scholar] [CrossRef]
- Cleck, J.N.; Blendy, J.A. Making a bad thing worse: Adverse effects of stress on drug addiction. J. Clin. Investig. 2008, 118, 454–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koob, G.F.; Le Moal, M. Addiction and the brain antireward system. Annu. Rev. Psychol. 2008, 59, 29–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, S.D.; Sumnall, H.R.; Measham, F.; Cole, J. Analyses of second-generation ‘legal highs’ in the UK: Initial findings. Drug Test. Anal. 2010, 2, 377–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hockenhull, J.; Murphy, K.G.; Paterson, S. Mephedrone use is increasing in London. Lancet 2016, 387, 1719–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angoa-Pérez, M.; Kane, M.J.; Francescutti, D.M.; Sykes, K.E.; Shah, M.M.; Mohammed, A.M.; Thomas, D.M.; Kuhn, D.M. Mephedrone, an abused psychoactive component of ‘bath salts’ and methamphetamine congener, does not cause neurotoxicity to dopamine nerve endings of the striatum. J. Neurochem. 2012, 120, 1097–1107. [Google Scholar] [CrossRef] [Green Version]
- Papaseit, E.; Moltó, J.; Muga, R.; Torrens, M.; de la Torre, R.; Farré, M. Clinical pharmacology of the synthetic cathinone mephedrone. Curr. Top. Behav. Neurosci. 2017, 32, 313–331. [Google Scholar] [CrossRef]
- Grochecki, P.; Smaga, I.; Lopatynska-Mazurek, M.; Gibula-Tarlowska, E.; Kedzierska, E.; Listos, J.; Talarek, S.; Marszalek-Grabska, M.; Hubalewska-Mazgaj, M.; Korga-Plewko, A.; et al. Effects of mephedrone and amphetamine exposure during adolescence on spatial memory in adulthood: Behavioral and neurochemical analysis. Int. J. Mol. Sci. 2021, 22, 589. [Google Scholar] [CrossRef]
- Grochecki, P.; Smaga, I.; Marszalek-Grabska, M.; Lopatynska-Mazurek, M.; Slowik, T.; Gibula-Tarlowska, E.; Kedzierska, E.; Listos, J.; Filip, M.; Kotlinska, J.H. Alteration of ethanol reward by prior mephedrone exposure: The role of age and matrix metalloproteinase-9 (MMP-9). Int. J. Mol. Sci. 2022, 23, 2122. [Google Scholar] [CrossRef]
- Papaseit, E.; Pérez-Mañá, C.; de Sousa Fernandes Perna, E.B.; Olesti, E.; Mateus, J.; Kuypers, K.P.C.; Theunissen, E.L.; Fonseca, F.; Torrens, M.; Ramaekers, J.G.; et al. Mephedrone and alcohol interactions in humans. Front. Pharmacol. 2020, 10, 1588. [Google Scholar] [CrossRef]
- Martinelli, S.; Anderzhanova, E.A.; Bajaj, T.; Wiechmann, S.; Dethloff, F.; Weckmann, K.; Heinz, D.E.; Ebert, T.; Hartmann, J.; Geiger, T.M.; et al. Stress-primed secretory autophagy promotes extracellular BDNF maturation by enhancing MMP9 secretion. Nat. Commun. 2021, 12, 4643. [Google Scholar] [CrossRef] [PubMed]
- Michaluk, P.; Mikasova, L.; Groc, L.; Frischknecht, R.; Choquet, D.; Kaczmarek, L. Matrix metalloproteinase-9 controls NMDA receptor surface diffusion through integrin beta1 signaling. J. Neurosci. 2009, 29, 6007–6012. [Google Scholar] [CrossRef] [Green Version]
- Chambers, R.A.; Bremner, J.D.; Moghaddam, B.; Southwick, S.M.; Charney, D.S.; Krystal, J.H. Glutamate and post-traumatic stress disorder: Toward a psychobiology of dissociation. Semin. Clin. Neuropsychiatry 1999, 4, 274–281. [Google Scholar] [CrossRef]
- Dalton, G.L.; Wu, D.C.; Wang, Y.T.; Floresco, S.B.; Phillips, A.G. NMDA GluN2A and GluN2B receptors play separate roles in the induction of LTP and LTD in the amygdala and in the acquisition and extinction of conditioned fear. Neuropharmacology 2012, 62, 797–806. [Google Scholar] [CrossRef]
- Bienvenu, T.C.M.; Dejean, C.; Jercog, D.; Aouizerate, B.; Lemoine, M.; Herry, C. The advent of fear conditioning as an animal model of post-traumatic stress disorder: Learning from the past to shape the future of PTSD research. Neuron 2021, 109, 2380–2397. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.R.; McGuire, J.; Lazarus, R.; Palmer, A.A. Pavlovian fear memory circuits and phenotype models of PTSD. Neuropharmacology 2012, 62, 638–646. [Google Scholar] [CrossRef]
- Ross, D.A.; Arbuckle, M.R.; Travis, M.J.; Dwyer, J.B.; van Schalkwyk, G.I.; Ressler, K.J. An integrated neuroscience perspective on formulation and treatment planning for posttraumatic stress disorder: An educational review. JAMA Psychiatry 2017, 74, 407–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.J.; Fanselow, M.S. Modality-specific retrograde amnesia of fear. Science 1992, 256, 675–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, R.G.; LeDoux, J.E. Differential contribution of amygdala and hippocampus to cued and contextual fear conditioning. Behav. Neurosci. 1992, 106, 274–285. [Google Scholar] [CrossRef]
- Rudy, J.W. Contextual conditioning and auditory cue conditioning dissociate during development. Behav. Neurosci. 1993, 107, 887–891. [Google Scholar] [CrossRef] [PubMed]
- Logue, S.F.; Paylor, R.; Wehner, J.M. Hippocampal lesions cause learning deficits in inbred mice in the Morris water maze and conditioned-fear task. Behav. Neurosci. 1997, 111, 104–113. [Google Scholar] [CrossRef]
- Caprioli, D.; Celentano, M.; Paolone, G.; Badiani, A. Modeling the role of environment in addiction. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2007, 31, 1639–1653. [Google Scholar] [CrossRef] [PubMed]
- Le Moal, M. Drug abuse: Vulnerability and transition to addiction. Pharmacopsychiatry 2009, 42 (Suppl. 1), S42–S55. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Yap, J.J.; Covington, H.E., 3rd. Social stress, therapeutics and drug abuse: Preclinical models of escalated and depressed intake. Pharmacol. Ther. 2008, 120, 102–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, R. New findings on biological factors predicting addiction relapse vulnerability. Curr. Psychiatry Rep. 2011, 13, 398–405. [Google Scholar] [CrossRef] [Green Version]
- Burke, A.R.; Miczek, K.A. Stress in adolescence and drugs of abuse in rodent models: Role of dopamine, CRF, and HPA axis. Psychopharmacology 2014, 231, 1557–1580. [Google Scholar] [CrossRef] [Green Version]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Arias, M.; Valverde, O.; Daza-Losada, M.; Blanco-Gandía, M.C.; Aguilar, M.A.; Miñarro, J. Assessment of the abuse potential of MDMA in the conditioned place preference paradigm: Role of CB1 receptors. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 47, 77–84. [Google Scholar] [CrossRef]
- Sinha, R. Chronic stress, drug use, and vulnerability to addiction. Ann. N. Y. Acad. Sci. 2008, 1141, 105–130. [Google Scholar] [CrossRef] [Green Version]
- Bahi, A.; Dreyer, J.L. Chronic psychosocial stress causes delayed extinction and exacerbates reinstatement of ethanol-induced conditioned place preference in mice. Psychopharmacology 2014, 231, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Filarowska-Jurko, J.; Komsta, L.; Smaga, I.; Surowka, P.; Marszalek-Grabska, M.; Grochecki, P.; Nizio, D.; Filip, M.; Kotlinska, J.H. Maternal separation alters ethanol drinking and reversal learning processes in adolescent rats: The impact of sex and glycine transporter type 1 (GlyT1) inhibitor. Int. J. Mol. Sci. 2022, 23, 5350. [Google Scholar] [CrossRef]
- Cannady, R.; Nguyen, T.; Padula, A.E.; Rinker, J.A.; Lopez, M.F.; Becker, H.C.; Woodward, J.J.; Mulholland, P.J. Interaction of chronic intermittent ethanol and repeated stress on structural and functional plasticity in the mouse medial prefrontal cortex. Neuropharmacology 2021, 182, 108396. [Google Scholar] [CrossRef]
- Barchiesi, R.; Chanthongdee, K.; Domi, E.; Gobbo, F.; Coppola, A.; Asratian, A.; Toivainen, S.; Holm, L.; Augier, G.; Xu, L.; et al. Stress-induced escalation of alcohol self-administration, anxiety-like behavior, and elevated amygdala Avp expression in a susceptible subpopulation of rats. Addict. Biol. 2021, 26, e13009. [Google Scholar] [CrossRef]
- Enman, N.M.; Arthur, K.; Ward, S.J.; Perrine, S.A.; Unterwald, E.M. Anhedonia, reduced cocaine reward, and dopamine dysfunction in a rat model of posttraumatic stress disorder. Biol. Psychiatry 2015, 78, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Lguensat, A.; Montanari, C.; Vielle, C.; Bennis, M.; Ba-M’hamed, S.; Baunez, C.; Garcia, R. Repeated cocaine exposure prior to fear conditioning induces persistency of PTSD-like symptoms and enhancement of hippocampal and amygdala cell density in male rats. Brain Struct. Funct. 2021, 226, 2219–2241. [Google Scholar] [CrossRef]
- Eagle, A.L.; Perrine, S.A. Methamphetamine-induced behavioral sensitization in a rodent model of posttraumatic stress disorder. Drug Alcohol Depend. 2013, 131, 36–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanagel, R.; Noori, H.R.; Heilig, M. Stress and alcohol interactions: Animal studies and clinical significance. Trends Neurosci. 2014, 37, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Weera, M.M.; Gilpin, N.W. Biobehavioral interactions between stress and alcohol. Alcohol. Res. 2019, 40, arcr.v40.1.04. [Google Scholar] [CrossRef] [PubMed]
- Burke, A.R.; Watt, M.J.; Forster, G.L. Adolescent social defeat increases adult amphetamine conditioned place preference and alters D2 dopamine receptor expression. Neuroscience 2011, 197, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Hymel, K.A.; Eans, S.O.; LSitchenko, K.; Gomes, S.M.; Lukowsky, A.L.; Medina, J.M.; Sypek, E.I.; Carey, A.N.; McLaughlin, J.P. Stress-induced increases in depression-like and cocaine place-conditioned behaviors are reversed by disruption of memories during reconsolidation. Behav. Pharmacol. 2014, 25, 599–608. [Google Scholar] [CrossRef]
- McLaughlin, J.P.; Li, S.; Valdez, J.; Chavkin, T.A.; Chavkin, C. Social defeat stress-induced behavioral responses are mediated by the endogenous kappa opioid system. Neuropsychopharmacology 2006, 31, 1241–1248. [Google Scholar] [CrossRef]
- Marquardt, A.R.; Ortiz-Lemos, L.; Lucion, A.B.; Barros, H.M. Influence of handling or aversive stimulation during rats’ neonatal or adolescence periods on oral cocaine self-administration and cocaine withdrawal. Behav. Pharmacol. 2004, 15, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Piazza, P.V.; Deminiere, J.M.; le Moal, M.; Simon, H. Stress- and pharmacologically-induced behavioral sensitization increases vulnerability to acquisition of amphetamine self-administration. Brain Res. 1990, 514, 22–26. [Google Scholar] [CrossRef]
- García-Pardo, M.P.; Blanco-Gandía, M.C.; Valiente-Lluch, M.; Rodríguez-Arias, M.; Miñarro, J.; Aguilar, M.A. Long-term effects of repeated social stress on the conditioned place preference induced by MDMA in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2015, 63, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Siegmund, A.; Wotjak, C.T. A mouse model of posttraumatic stress disorder that distinguishes between conditioned and sensitised fear. J. Psychiatr. Res. 2007, 41, 848–860. [Google Scholar] [CrossRef]
- Cohen, H.; Zohar, J.; Matar, M.A.; Zeev, K.; Loewenthal, U.; Richter-Levin, G. Setting apart the affected: The use of behavioral criteria in animal models of post traumatic stress disorder. Neuropsychopharmacology 2004, 29, 1962–1970. [Google Scholar] [CrossRef]
- Bentefour, Y.; Bennis, M.; Garcia, R.; M’hamed, S.B. Effects of paroxetine on PTSD-like symptoms in mice. Psychopharmacology 2015, 232, 2303–2312. [Google Scholar] [CrossRef]
- Bentefour, Y.; Bennis, M.; Garcia, R.; Ba-M’hamed, S. High-frequency stimulation of the infralimbic cortex, following behavioral suppression of PTSD-like symptoms, prevents symptom relapse in mice. Brain Stimul. 2018, 11, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.C.; Gould, T.J. Nicotine and ethanol enhancements of acoustic startle reflex are mediated in part by dopamine in C57BL/6J mice. Pharmacol. Biochem. Behav. 2003, 76, 179–186. [Google Scholar] [CrossRef]
- Perusini, J.N.; Meyer, E.M.; Long, V.A.; Rau, V.; Nocera, N.; Avershal, J.; Maksymetz, J.; Spigelman, I.; Fanselow, M.S. Induction and expression of fear sensitization caused by acute traumatic stress. Neuropsychopharmacology 2016, 41, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Olatunji, B.O.; Fan, Q.; Wolitzky-Taylor, K. Anxiety sensitivity and post-traumatic stress reactions: Effects of time-varying intrusive thoughts and associated distress. J. Behav. Ther. Exp. Psychiatry 2018, 61, 113–120. [Google Scholar] [CrossRef]
- Marshall, G.N.; Miles, J.N.; Stewart, S.H. Anxiety sensitivity and PTSD symptom severity are reciprocally related: Evidence from a longitudinal study of physical trauma survivors. J. Abnorm. Psychol. 2010, 119, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, E.A.; Reisfield, G.M.; Watson, M.C.; Chronister, C.W.; Goldberger, B.A. Psychoactive “bath salts” intoxication with methylenedioxypyrovalerone. Am. J. Med. 2012, 125, 854–858. [Google Scholar] [CrossRef] [PubMed]
- Valente, M.J.; Guedes de Pinho, P.; de Lourdes Bastos, M.; Carvalho, F.; Carvalho, M. Khat and synthetic cathinones: A review. Arch. Toxicol. 2014, 88, 15–45. [Google Scholar] [CrossRef] [PubMed]
- Perrine, S.A.; Sheikh, I.S.; Nwaneshiudu, C.A.; Schroeder, J.A.; Unterwald, E.M. Withdrawal from chronic administration of cocaine decreases delta opioid receptor signaling and increases anxiety- and depression-like behaviors in the rat. Neuropharmacology 2008, 54, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Kitanaka, N.; Kitanaka, J.; Watabe, K.; Takemura, M. Low-dose pretreatment with clorgyline decreases the levels of 3-methoxy-4-hydroxyphenylglycol in the striatum and nucleus accumbens and attenuates methamphetamine-induced conditioned place preference in rats. Neuroscience 2010, 165, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- El Hage, C.; Rappeneau, V.; Etievant, A.; Morel, A.L.; Scarna, H.; Zimmer, L.; Bérod, A. Enhanced anxiety observed in cocaine withdrawn rats is associated with altered reactivity of the dorsomedial prefrontal cortex. PLoS ONE 2012, 7, e43535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Qiu, Y.; Zhang, Y.; Βai, Y.; Xu, P.; Liu, Y.; Kim, J.H.; Shen, H.W. The effects of 4-methylethcathinone on conditioned place preference, locomotor sensitization, and anxiety-like behavior: A comparison with methamphetamine. Int. J. Neuropsychopharmacol. 2016, 19, pyv120. [Google Scholar] [CrossRef] [Green Version]
- Myers, K.M.; Davis, M. Mechanisms of fear extinction. Mol. Psychiatry 2007, 12, 120–150. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.C.; Fay, J.; Sage, J.R.; Anagnostaras, S.G. Cocaine and Pavlovian fear conditioning: Dose-effect analysis. Behav. Brain Res. 2007, 176, 244–250. [Google Scholar] [CrossRef] [Green Version]
- Shuman, T.; Wood, S.C.; Anagnostaras, S.G. Modafinil and memory: Effects of modafinil on Morris water maze learning and Pavlovian fear conditioning. Behav. Neurosci. 2009, 123, 257–266. [Google Scholar] [CrossRef]
- Carmack, S.A.; Howell, K.K.; Rasaei, K.; Reas, E.T.; Anagnostaras, S.G. Animal model of methylphenidate’s long-term memory-enhancing effects. Learn Mem. 2014, 21, 82–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maren, S.; Quirk, G.J. Neuronal signalling of fear memory. Nat. Rev. Neurosci. 2004, 5, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Pape, H.C.; Pare, D. Plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear. Physiol. Rev. 2010, 90, 419–463. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, K.; Rejmak, E.; Mikosz, M.; Nikolaev, E.; Knapska, E.; Kaczmarek, L. Matrix metalloproteinase (MMP) 9 transcription in mouse brain induced by fear learning. J. Biol. Chem. 2013, 288, 20978–20991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanElzakker, M.B.; Dahlgren, M.K.; Davis, F.C.; Dubois, S.; Shin, L.M. From Pavlov to PTSD: The extinction of conditioned fear in rodents, humans, and anxiety disorders. Neurobiol. Learn. Mem. 2014, 113, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Bach, D.R.; Tzovara, A.; Vunder, J. Blocking human fear memory with the matrix metalloproteinase inhibitor doxycycline. Mol. Psychiatry 2018, 23, 1584–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brahmajothi, M.V.; Abou-Donia, M.B. PTSD susceptibility and challenges: Pathophysiological consequences of behavioral symptoms. Mil. Med. 2020, 185, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Lima, B.B.; Hammadah, M.; Wilmot, K.; Pearce, B.D.; Shah, A.; Levantsevych, O.; Kaseer, B.; Obideen, M.; Gafeer, M.M.; Kim, J.H.; et al. Posttraumatic stress disorder is associated with enhanced interleukin-6 response to mental stress in subjects with a recent myocardial infarction. Brain Behav. Immun. 2019, 75, 26–33. [Google Scholar] [CrossRef]
- Szymanowski, A.; Nijm, J.; Kristenson, M.; Jonasson, L. Elevated levels of circulating matrix metalloproteinase-9 are associated with a dysregulated cortisol rhythm--A case-control study of coronary artery disease. Psychoneuroendocrinology 2011, 36, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, A.K.; Jönsson, S.; Stenmark, J.; Kristenson, M.; Jonasson, L. Stress-induced release of matrix metalloproteinase-9 in patients with coronary artery disease: The possible influence of cortisol. Psychoneuroendocrinology 2016, 73, 117–124. [Google Scholar] [CrossRef]
- Aguayo, F.I.; Pacheco, A.A.; García-Rojo, G.J.; Pizarro-Bauerle, J.A.; Doberti, A.V.; Tejos, M.; García-Pérez, M.A.; Rojas, P.S.; Fiedler, J.L. Matrix metalloproteinase 9 displays a particular time response to acute stress: Variation in its levels and activity distribution in rat hippocampus. ACS Chem. Neurosci. 2018, 9, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.E.; Wilson, A.R.; Cocking, D.L.; Sorg, B.A. Inhibition of matrix metalloproteinase activity disrupts reconsolidation but not consolidation of a fear memory. Neurobiol. Learn. Mem. 2009, 91, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wronikowska, O.; Zykubek, M.; Kurach, Ł.; Michalak, A.; Boguszewska-Czubara, A.; Budzyńska, B. Vulnerability factors for mephedrone-induced conditioned place preference in rats-the impact of sex differences, social-conditioning and stress. Psychopharmacology 2021, 238, 2947–2961. [Google Scholar] [CrossRef] [PubMed]
- Lguensat, A.; Boudjafad, Z.; Giorla, E.; Bennis, M.; Baunez, C.; Garcia, R.; Ba-M’hamed, S. Repeated ethanol exposure following avoidance conditioning impairs avoidance extinction and modifies conditioning-associated prefrontal dendritic changes in a mouse model of post-traumatic stress disorder. Eur. J. Neurosci. 2021, 54, 7710–7732. [Google Scholar] [CrossRef] [PubMed]
- Koltunowska, D.; Gibula-Bruzda, E.; Kotlinska, J.H. The influence of ionotropic and metabotropic glutamate receptor ligands on anxiety-like effect of amphetamine withdrawal in rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 45, 242–249. [Google Scholar] [CrossRef]
- Gibula-Bruzda, E.; Marszalek-Grabska, M.; Witkowska, E.; Izdebski, J.; Kotlinska, J.H. Enkephalin analog, cyclo[N(ε),N(β)-carbonyl-D-Lys(2),Dap(5)] enkephalinamide (cUENK6), inhibits the ethanol withdrawal-induced anxiety-like behavior in rats. Alcohol 2015, 49, 229–236. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grochecki, P.; Smaga, I.; Wydra, K.; Marszalek-Grabska, M.; Slowik, T.; Kedzierska, E.; Listos, J.; Gibula-Tarlowska, E.; Filip, M.; Kotlinska, J.H. Impact of Mephedrone on Fear Memory in Adolescent Rats: Involvement of Matrix Metalloproteinase-9 (MMP-9) and N-Methyl-D-aspartate (NMDA) Receptor. Int. J. Mol. Sci. 2023, 24, 1941. https://doi.org/10.3390/ijms24031941
Grochecki P, Smaga I, Wydra K, Marszalek-Grabska M, Slowik T, Kedzierska E, Listos J, Gibula-Tarlowska E, Filip M, Kotlinska JH. Impact of Mephedrone on Fear Memory in Adolescent Rats: Involvement of Matrix Metalloproteinase-9 (MMP-9) and N-Methyl-D-aspartate (NMDA) Receptor. International Journal of Molecular Sciences. 2023; 24(3):1941. https://doi.org/10.3390/ijms24031941
Chicago/Turabian StyleGrochecki, Pawel, Irena Smaga, Karolina Wydra, Marta Marszalek-Grabska, Tymoteusz Slowik, Ewa Kedzierska, Joanna Listos, Ewa Gibula-Tarlowska, Malgorzata Filip, and Jolanta H. Kotlinska. 2023. "Impact of Mephedrone on Fear Memory in Adolescent Rats: Involvement of Matrix Metalloproteinase-9 (MMP-9) and N-Methyl-D-aspartate (NMDA) Receptor" International Journal of Molecular Sciences 24, no. 3: 1941. https://doi.org/10.3390/ijms24031941
APA StyleGrochecki, P., Smaga, I., Wydra, K., Marszalek-Grabska, M., Slowik, T., Kedzierska, E., Listos, J., Gibula-Tarlowska, E., Filip, M., & Kotlinska, J. H. (2023). Impact of Mephedrone on Fear Memory in Adolescent Rats: Involvement of Matrix Metalloproteinase-9 (MMP-9) and N-Methyl-D-aspartate (NMDA) Receptor. International Journal of Molecular Sciences, 24(3), 1941. https://doi.org/10.3390/ijms24031941