
Anti-Obesity Effects of Metformin: A Scoping Review Evaluating the Feasibility of Brown Adipose Tissue as a Therapeutic Target

 , , ,
, , ,  and
and
Abstract
:1. Introduction
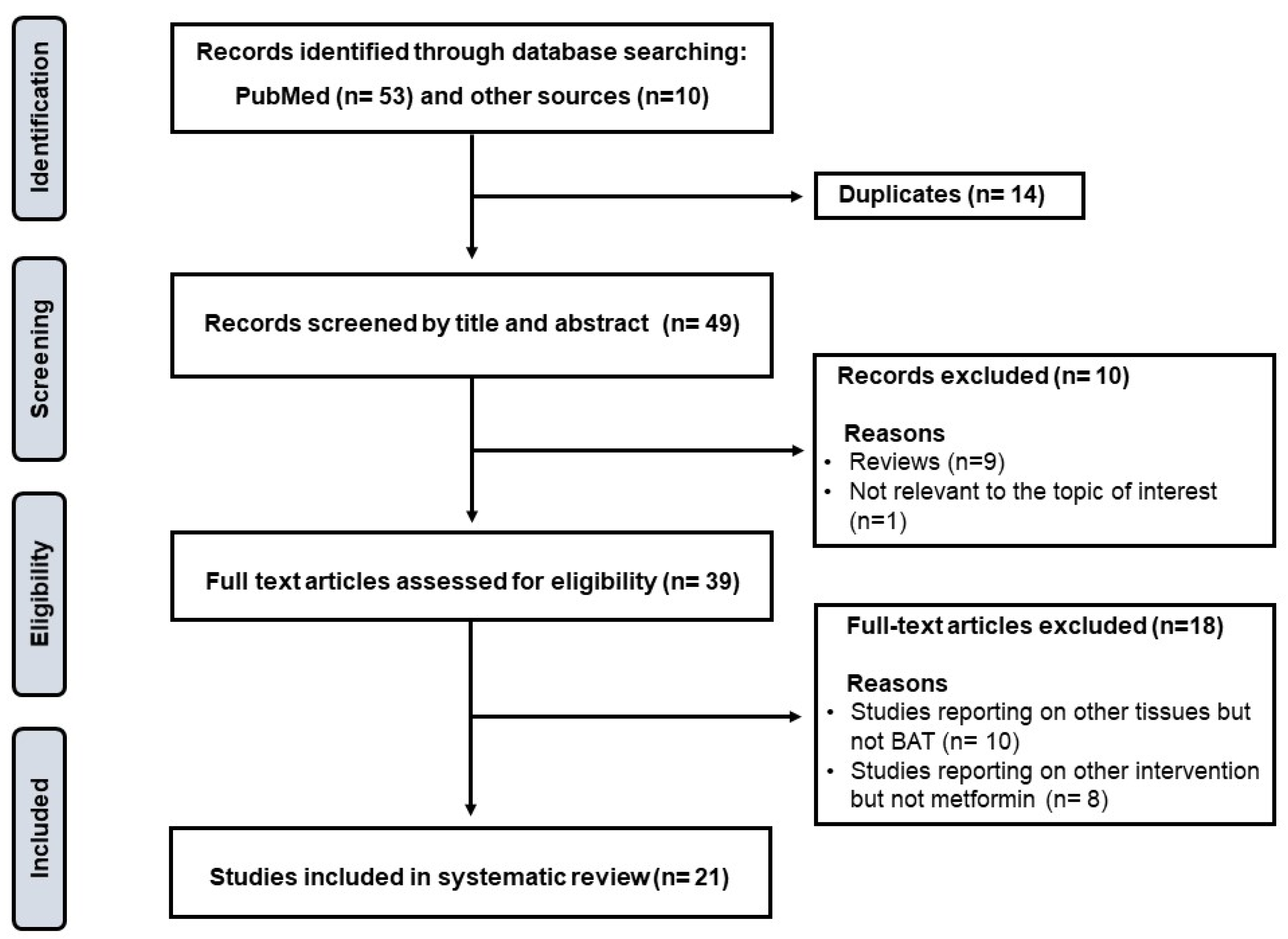
2. Methods: Search Strategy, Study Eligibility Criteria, and Data Items
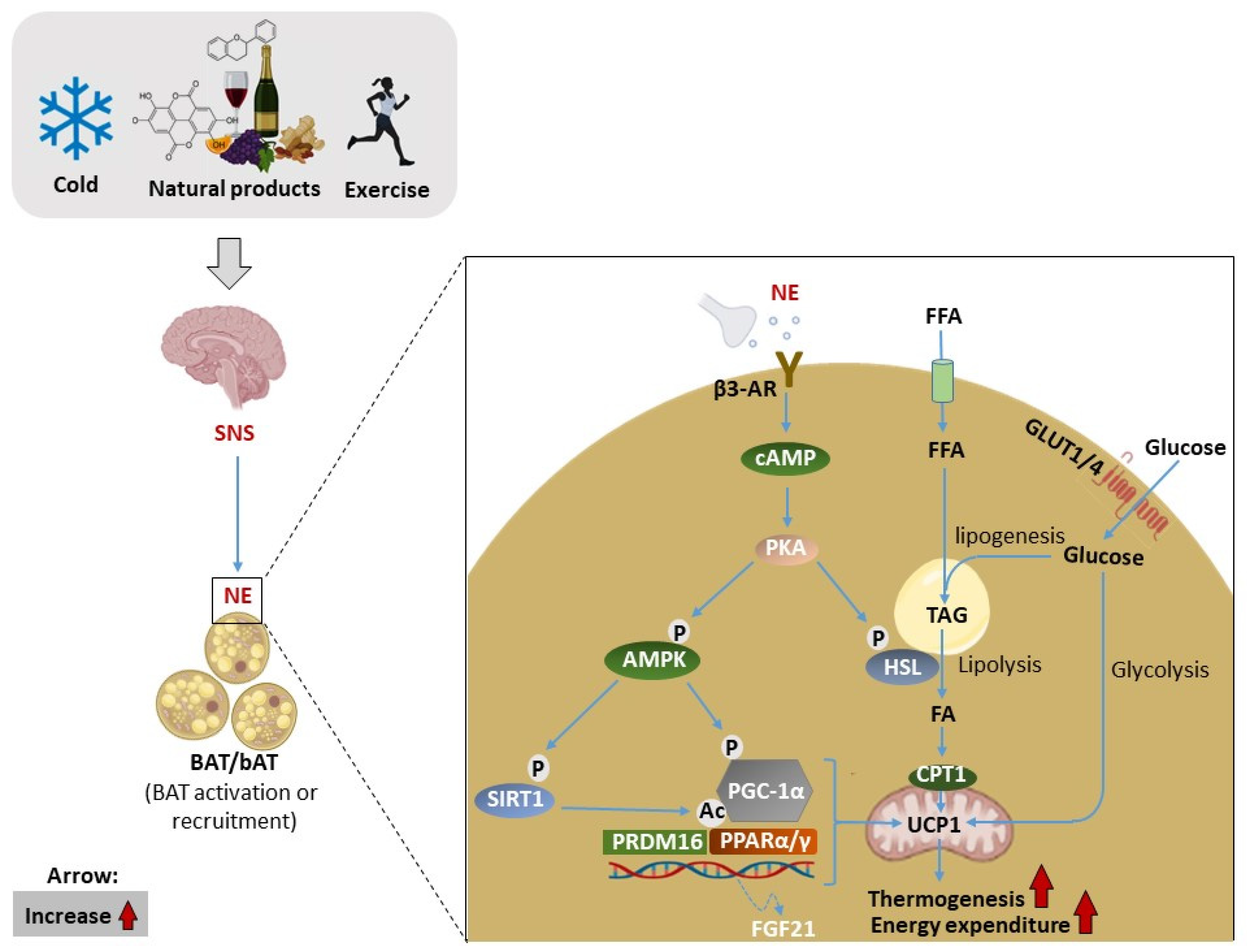
3. The Significance of BAT and Thermogenesis as a Therapeutic Target
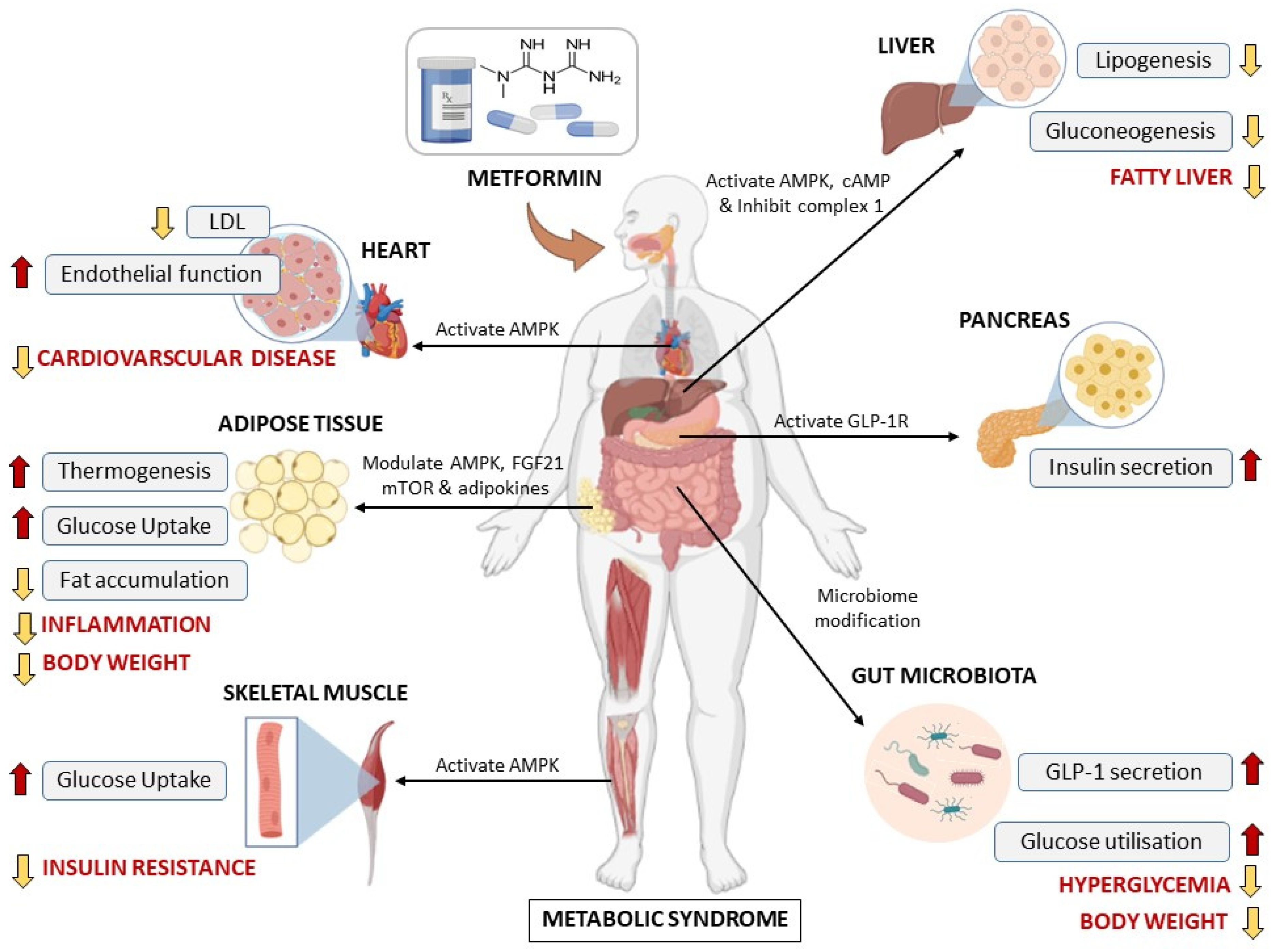
4. An Overview of Metformin and Its Therapeutic Potential against Metabolic Diseases
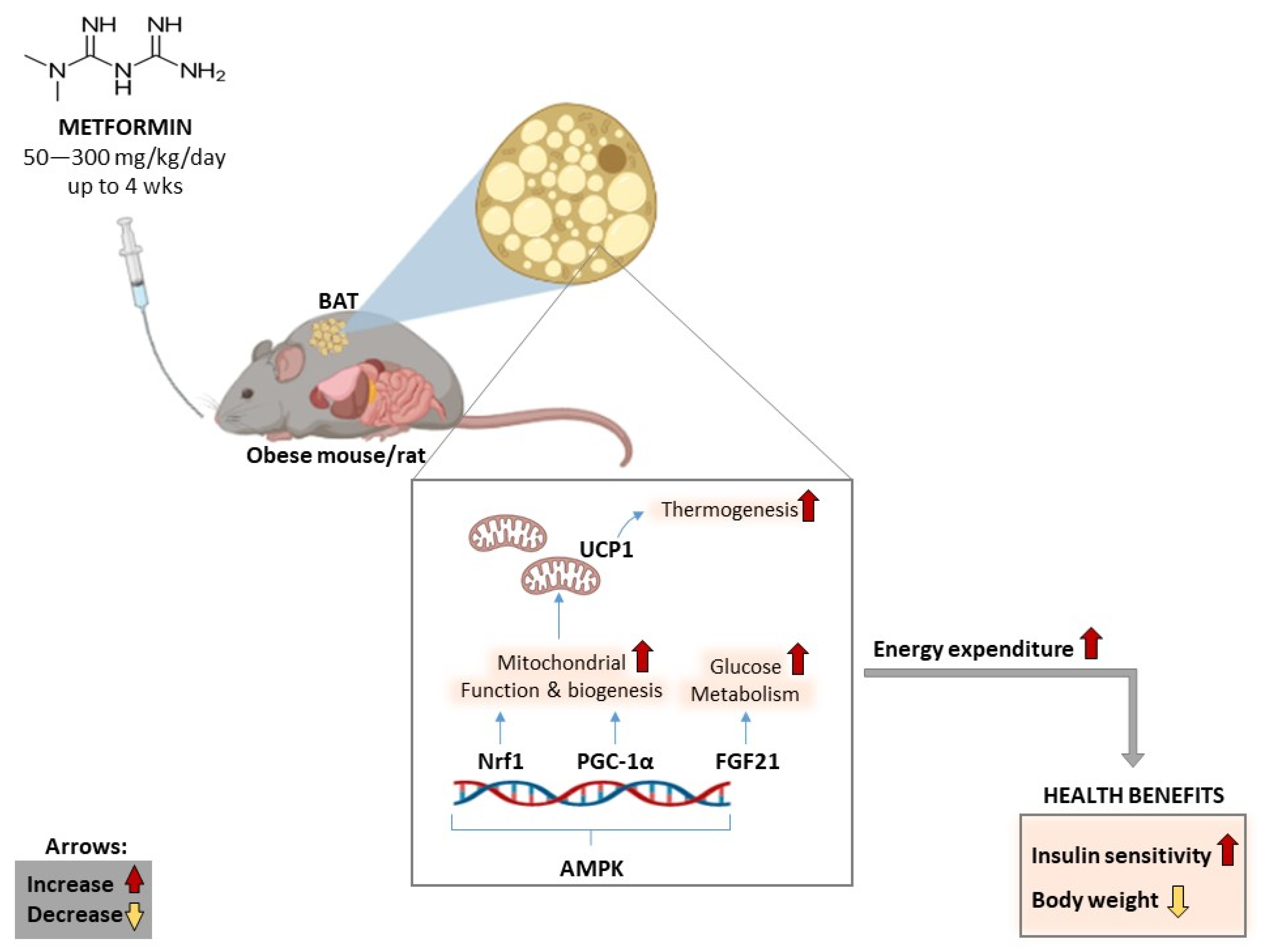
5. Results: Impact of Metformin on Energy Expenditure and BAT Activity
5.1. General Characteristics and Overview of Included Literature
5.2. Evidence on the Short-Term Treatment Effects of Metformin
5.3. Evidence on the Long-Term Treatment Effects of Metformin
6. Clinical Translation of Results
7. Conclusions and Future Perspective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chooi, Y.C.; Ding, C.; Magkos, F. The epidemiology of obesity. Metabolism 2019, 92, 6–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 20 October 2022).
- Anthony, S.R.; Guarnieri, A.R.; Gozdiff, A.; Helsley, R.N.; Owens, A.P.; Tranteret, M. Mechanisms linking adipose tissue inflammation to cardiac hypertrophy and fibrosis. Clin. Sci. 2019, 133, 2329–2344. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M. Metabolic syndrome update. Trends. Cardiovasc. Med. 2016, 26, 364–373. [Google Scholar] [CrossRef]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [Green Version]
- Mazibuko-Mbeje, S.E.; Dludla, P.V.; Roux, C.; Johnson, R.; Ghoor, S.; Joubert, E.; Louw, J.; Opoku, A.R.; Muller, C.J.F. Aspalathin-enriched green rooibos extract reduces hepatic insulin resistance by modulating PI3K/AKT and AMPK pathways. Int. J. Mol. Sci. 2019, 20, 633. [Google Scholar] [CrossRef] [Green Version]
- Sam, S.; Mazzone, T. Adipose tissue changes in obesity and the impact on metabolic function. Transl. Res. 2014, 164, 284–292. [Google Scholar] [CrossRef]
- Saely, C.H.; Geiger, K.; Drexel, H. Brown versus white adipose tissue: A mini-review. Gerontology 2012, 58, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Aldiss, P.; Betts, J.; Sale, C.; Pope, M.; Budge, H.; Symonds, M.E. Exercise-induced ‘browning’ of adipose tissues. Metabolism 2018, 81, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Nyawo, T.A.; Pheiffer, C.; Mazibuko-Mbeje, S.E.; Mthembu, S.X.H.; Nyambuya, T.M.; Nkambule, B.B.; Sadie-Van Gijsen, H.; Strijdom, H.; Tiano, L.; Dludla, P.V. Physical exercise potentially targets epicardial adipose tissue to reduce cardiovascular disease risk in patients with metabolic diseases: Oxidative stress and inflammation emerge as major therapeutic targets. Antioxidants 2021, 10, 1758. [Google Scholar] [CrossRef]
- Mthembu, S.X.H.; Mazibuko-Mbeje, S.E.; Ziqubu, K.; Nyawo, T.A.; Obonye, N.; Nyambuya, T.M.; Nkambule, B.B.; Silvestri, S.; Tiano, L.; Muller, C.J.F.; et al. Impact of physical exercise and caloric restriction in patients with type 2 diabetes: Skeletal muscle insulin resistance and mitochondrial dysfunction as ideal therapeutic targets. Life Sci. 2022, 297, 120467. [Google Scholar] [CrossRef]
- Kuryłowicz, A.; Puzianowska-Kuźnicka, M. Induction of adipose tissue browning as a strategy to combat obesity. Int. J. Mol. Sci. 2020, 21, 6241. [Google Scholar] [CrossRef] [PubMed]
- Fenzl, A.; Kiefer, F.W. Brown adipose tissue and thermogenesis. Horm. Mol. Biol. Clin. Investig. 2014, 19, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, A.; Krause, F.N.; Moran, A.; MacCannell, A.D.V.; Scragg, J.L.; McNally, B.D.; Boatng, E.; Murfitt, S.A.; Virtue, S.; Wright, J.; et al. Brown and beige adipose tissue regulate systemic metabolism through a metabolite interorgan signaling axis. Nat. Commun. 2021, 12, 1905. [Google Scholar] [CrossRef] [PubMed]
- Melguizo Rodríguez, L.; Illescas-Montes, R.; Costela-Ruiz, V.J.; García-Martínez, O. Stimulation of brown adipose tissue by polyphenols in extra virgin olive oil. Crit. Rev. Food Sci. Nutr. 2021, 61, 3481–3488. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Z.; Tan, B.K.; Christiand, M. Dietary polyphenols turn fat “brown”: A narrative review of the possible mechanisms. Trends Food Sci. Technol. 2021, 97, 221–232. [Google Scholar] [CrossRef]
- Engler, C.; Leo, M.; Pfeifer, B.; Juchum, M.; Chen-Koenig, D.; Poelzl, K.; Schoenherr, H.; Vill, D.; Oberdanner, J.; Eisendle, E.; et al. Long-term trends in the prescription of antidiabetic drugs: Real-world evidence from the Diabetes Registry Tyrol 2012–2018. BMJ Open Diabetes Res. Care 2020, 8, e001279. [Google Scholar] [CrossRef]
- Lv, Z.; Guo, Y. Metformin and Its Benefits for Various Diseases. Front. Endocrinol. 2020, 11, 191. [Google Scholar] [CrossRef]
- Prattichizzo, F.; Giuliani, A.; Mensà, E.; Sabbatinelli, J.; De Nigris, V.; Rippo, M.R.; La Sala, L.; Procopio, A.D.; Olivieri, F.; Ceriello, A. Pleiotropic effects of metformin: Shaping the microbiome to manage type 2 diabetes and postpone ageing. Ageing Res. Rev. 2018, 48, 87–98. [Google Scholar] [CrossRef]
- Lee, A.; Morley, J.E. Metformin decreases food consumption and induces weight loss in subjects with obesity with type II non-insulin-dependent diabetes. Obes. Res. 1998, 6, 47–53. [Google Scholar] [CrossRef]
- Seifarth, C.; Schehler, B.; Schneider, H.J. Effectiveness of metformin on weight loss in non-diabetic individuals with obesity. Exp. Clin. Endocrinol. Diabetes 2013, 121, 27–31. [Google Scholar] [CrossRef]
- Ejtahed, H.S.; Tito, R.Y.; Siadat, S.D.; Hasani-Ranjbar, S.; Hoseini-Tavassol, Z.; Rymenans, L.; Verbeke, K.; Soroush, A.R.; Raes, J.; Larijani, B. Metformin induces weight loss associated with gut microbiota alteration in non-diabetic obese women: A randomized double-blind clinical trial. Eur. J. Endocrinol. 2019, 180, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, J.P.; Alemzadeh, R.; Langley, G.; D’Angelo, L.; Smith, P.; Holshouser, S. Beneficial effects of metformin in normoglycemic morbidly obese adolescents. Metabolism 2001, 50, 1457–1461. [Google Scholar] [CrossRef] [PubMed]
- Levri, K.M.; Slaymaker, E.; Last, A.; Yeh, J.; Ference, J.; D’Amico, F.; Wilson, S.A. Metformin as treatment for overweight and obese adults: A systematic review. Ann. Fam. Med. 2005, 3, 457–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masarwa, R.; Brunetti, V.C.; Aloe, S.; Henderson, M.; Platt, R.W.; Filion, K.B. Efficacy and Safety of Metformin for Obesity: A Systematic Review. Pediatrics 2021, 147. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, A.; Mousavi, S.M.; Mokhtari, T.; Parohan, M.; Milajerdi, A. Metformin Therapy Reduces Obesity Indices in Children and Adolescents: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. Child. Obes. 2020, 16, 174–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerevanian, A.; Soukas, A.A. Metformin: Mechanisms in Human Obesity and Weight Loss. Curr. Obes. Rep. 2019, 8, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Nyambuya, T.M.; Mxinwa, V.; Mokgalaboni, K.; Ziqubu, K.; Cirilli, I.; Marcheggiani, F.; Louw, J.; et al. Adipokines as a therapeutic target by metformin to improve metabolic function: A systematic review of randomized controlled trials. Pharmacol. Res. 2021, 163, 105219. [Google Scholar] [CrossRef]
- Yuan, T.; Li, J.; Zhao, W.G.; Sun, W.; Liu, S.N.; Liu, Q.; Fu, Y.; Shen, Z.F. Effects of metformin on metabolism of white and brown adipose tissue in obese C57BL/6J mice. Diabetol. Metab. Syndr. 2019, 11, 96. [Google Scholar] [CrossRef]
- Kim, E.K.; Lee, S.H.; Jhun, J.Y.; Byun, J.K.; Jeong, J.H.; Lee, S.Y.; Kim, J.K.; Choi, J.Y.; Cho, M.L. Metformin Prevents Fatty Liver and Improves Balance of White/Brown Adipose in an Obesity Mouse Model by Inducing FGF21. Mediat. Inflamm. 2016, 2016, 5813030. [Google Scholar] [CrossRef] [Green Version]
- Breining, P.; Jensen, J.B.; Sundelin, E.I.; Gormsen, L.C.; Jakobsen, S.; Busk, M.; Rolighed, L.; Bross, P.; Fernandez-Guerra, P.; Markussen, L.K.; et al. Metformin targets brown adipose tissue in vivo and reduces oxygen consumption in vitro. Diabetes Obes. Metab. 2018, 20, 2264–2273. [Google Scholar] [CrossRef]
- Tokubuchi, I.; Tajiri, Y.; Iwata, S.; Hara, K.; Wada, N.; Hashinaga, T.; Nakayama, H.; Mifune, H.; Yamada, K. Beneficial effects of metformin on energy metabolism and visceral fat volume through a possible mechanism of fatty acid oxidation in human subjects and rats. PLoS ONE 2017, 12, e0171293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Yang, Q.; Zhang, L.; Maricelli, J.W.; Rodgers, B.D.; Zhu, M.J.; Du, M. Maternal high-fat diet during lactation impairs thermogenic function of brown adipose tissue in offspring mice. Sci. Rep. 2016, 6, 34345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karise, I.; Bargut, T.C.; Del Sol, M.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Metformin enhances mitochondrial biogenesis and thermogenesis in brown adipocytes of mice. Biomed. Pharmacother. 2019, 111, 1156–1165. [Google Scholar] [CrossRef]
- Singh, R.; Barrios, A.; Dirakvand, G.; Pervin, S. Human Brown Adipose Tissue and Metabolic Health: Potential for Therapeutic Avenues. Cells 2021, 10, 3030. [Google Scholar] [CrossRef]
- Carey, A.L.; Kingwell, B.A. Brown adipose tissue in humans: Therapeutic potential to combat obesity. Pharmacol. Ther. 2013, 140, 26–33. [Google Scholar] [CrossRef]
- Smith, R.E.; Horwitz, B.A. Brown fat and thermogenesis. Physiol. Rev. 1969, 49, 330–425. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.J.; Enerbäck, S.; et al. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.H.; Doria, A.; et al. Identification and importance of brown adipose tissue in adult humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.H.; Wei, Y.H. Therapeutic Perspectives of Thermogenic Adipocytes in Obesity and Related Complications. Int. J. Mol. Sci. 2021, 22, 7177. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Fang, H.; Guo, F.; Li, F.; Chen, A.; Huang, S. Flavonoids as inducers of white adipose tissue browning and thermogenesis: Signalling pathways and molecular triggers. Nutr. Metab. 2019, 16, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonet, M.L.; Oliver, P.; Palou, A. Pharmacological and nutritional agents promoting browning of white adipose tissue. Biochim. Et Biophys. Acta 2013, 1831, 969–985. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, D.; Xiang, J.; Zhou, J.; Cao, H.; Che, Q.; Bai, Y.; Guo, J.; Su, Z. Non-shivering Thermogenesis Signalling Regulation and Potential Therapeutic Applications of Brown Adipose Tissue. Int. J. Biol. Sci. 2021, 17, 2853–2870. [Google Scholar] [CrossRef] [PubMed]
- Jastroch, M.; Wuertz, S.; Kloas, W.; Klingenspor, M. Uncoupling protein 1 in fish uncovers an ancient evolutionary history of mammalian nonshivering thermogenesis. Physiol. Genom. 2005, 22, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Townsend, K.L.; Tseng, Y.H. Brown fat fuel utilization and thermogenesis. Trends Endocrinol. Metab. TEM 2014, 25, 168–177. [Google Scholar] [CrossRef] [Green Version]
- McNeill, B.T.; Morton, N.M.; Stimson, R.H. Substrate Utilization by Brown Adipose Tissue: What’s Hot and What’s Not? Front. Endocrinol. 2020, 11, 571659. [Google Scholar] [CrossRef]
- Randle, P.J.; Garland, P.B.; Hales, C.N.; Newsholme, E.A. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1963, 1, 785–789. [Google Scholar] [CrossRef]
- Ouellet, V.; Labbé, S.M.; Blondin, D.P.; Phoenix, S.; Guérin, B.; Haman, F.; Turcotte, E.E.; Richard, D.; Carpentier, A.C. Brown adipose tissue oxidative metabolism contributes to energy expenditure during acute cold exposure in humans. J. Clin. Investig. 2012, 122, 545–552. [Google Scholar] [CrossRef]
- Mazibuko-Mbeje, S.E.; Ziqubu, K.; Dludla, P.V.; Tiano, L.; Silvestri, S.; Orlando, P.; Nyawo, T.A.; Louw, J.; Kappo, A.P.; Muller, C.J.F. Isoorientin ameliorates lipid accumulation by regulating fat browning in palmitate-exposed 3T3-L1 adipocytes. Metab. Open 2020, 6, 100037. [Google Scholar] [CrossRef]
- Dallner, O.S.; Chernogubova, E.; Brolinson, K.A.; Bengtsson, T. Beta3-adrenergic receptors stimulate glucose uptake in brown adipocytes by two mechanisms independently of glucose transporter 4 translocation. Endocrinology 2006, 147, 5730–5739. [Google Scholar] [CrossRef]
- Saari, T.J.; Raiko, J.; M, U.D.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.; Haaparanta-Solin, M.; Nuutila, P.; Virtanen, K.A. Basal and cold-induced fatty acid uptake of human brown adipose tissue is impaired in obesity. Sci. Rep. 2020, 10, 14373. [Google Scholar] [CrossRef] [PubMed]
- Alcalá, M.; Calderon-Dominguez, M.; Serra, D.; Herrero, L.; Viana, M. Mechanisms of Impaired Brown Adipose Tissue Recruitment in Obesity. Front. Physiol. 2019, 10, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, I.; Walsh, K. The Whitening of Brown Fat and Its Implications for Weight Management in Obesity. Curr. Obes. Rep. 2015, 4, 224–229. [Google Scholar] [CrossRef] [PubMed]
- van der Vaart, J.I.; Boon, M.R.; Houtkooper, R.H. The Role of AMPK Signaling in Brown Adipose Tissue Activation. Cells 2021, 10, 1122. [Google Scholar] [CrossRef]
- Danysz, W.; Han, Y.; Li, F.; Nicoll, J.; Buch, P.; Hengl, T.; Ruitenberg, M.; Parsons, C. Browning of white adipose tissue induced by the ß3 agonist CL-316,243 after local and systemic treatment—PK-PD relationship. Biochim. Et Biophys. Acta. Mol. Basis Dis. 2018, 1864 (9 Pt B), 2972–2982. [Google Scholar] [CrossRef]
- Cypess, A.M.; Weiner, L.S.; Roberts-Toler, C.; Franquet Elía, E.; Kessler, S.H.; Kahn, P.A.; English, J.; Chatman, K.; Trauger, S.A.; Doria, A.; et al. Activation of human brown adipose tissue by a β3-adrenergic receptor agonist. Cell Metab. 2015, 21, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Baskin, A.S.; Linderman, J.D.; Brychta, R.J.; McGehee, S.; Anflick-Chames, E.; Cero, C.; Johnson, J.W.; O’Mara, A.E.; Fletcher, L.A.; Leitner, B.P.; et al. Regulation of Human Adipose Tissue Activation, Gallbladder Size, and Bile Acid Metabolism by a β3-Adrenergic Receptor Agonist. Diabetes 2018, 67, 2113–2125. [Google Scholar] [CrossRef] [Green Version]
- Ziqubu, K.; Dludla, P.V.; Joubert, E.; Muller, C.J.F.; Louw, J.; Tiano, L.; Nkambule, B.B.; Kappo, A.P.; Mazibuko-Mbeje, S.E. Isoorientin: A dietary flavone with the potential to ameliorate diverse metabolic complications. Pharmacol. Res. 2020, 158, 104867. [Google Scholar] [CrossRef]
- Silvester, A.J.; Aseer, K.R.; Yun, J.W. Dietary polyphenols and their roles in fat browning. J. Nutr. Biochem. 2019, 64, 1–12. [Google Scholar] [CrossRef]
- Wang, S.; Liang, X.; Yang, Q.; Fu, X.; Rogers, C.J.; Zhu, M.; Rodgers, B.D.; Jiang, Q.; Dodson, M.V.; Du, M. Resveratrol induces brown-like adipocyte formation in white fat through activation of AMP-activated protein kinase (AMPK) α1. Int. J. Obes. 2015, 39, 967–976. [Google Scholar] [CrossRef]
- Lakka, T.A.; Laaksonen, D.E. Physical activity in prevention and treatment of the metabolic syndrome. Appl. Physiol. Nutr. Metab.=Physiol. Appl. Nutr. Metab. 2007, 32, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Kim, S.H.; Min, Y.K.; Yang, H.M.; Lee, J.B.; Lee, M.S. Acute exercise induces FGF21 expression in mice and in healthy humans. PLoS ONE 2013, 8, e63517. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Linderman, J.D.; Smith, S.; Brychta, R.J.; Wang, J.; Idelson, C.; Perron, R.M.; Werner, C.D.; Phan, G.Q.; Kammula, U.S.; et al. Irisin and FGF21 are cold-induced endocrine activators of brown fat function in humans. Cell Metab. 2014, 19, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Szczepańska, E.; Gietka-Czernel, M. FGF21: A Novel Regulator of Glucose and Lipid Metabolism and Whole-Body Energy Balance. Horm. Metab. Res. = Horm. Stoffwechs. = Horm. Metab. 2022, 54, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yan, J.; Goh, H.J.; Govindharajulu, P.; Verma, S.; Michael, N.; Sadananthan, S.A.; Henry, C.J.; Velan, S.S.; Leow, M.K. Fibroblast Growth Factor-21, Leptin, and Adiponectin Responses to Acute Cold-Induced Brown Adipose Tissue Activation. J. Clin. Endocrinol. Metab. 2020, 105, e520–e531. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Schönke, M.; Zhou, E.; Li, Z.; Kooijman, S.; Boon, M.R.; Larsson, M.; Wallenius, K.; Dekker, N.; Barlind, L.; et al. Pharmacological treatment with FGF21 strongly improves plasma cholesterol metabolism to reduce atherosclerosis. Cardiovasc. Res. 2022, 118, 489–502. [Google Scholar] [CrossRef]
- Kaufman, A.; Abuqayyas, L.; Denney, W.S.; Tillman, E.J.; Rolph, T. AKR-001, an Fc-FGF21 Analog, Showed Sustained Pharmacodynamic Effects on Insulin Sensitivity and Lipid Metabolism in Type 2 Diabetes Patients. Cell Rep. Med. 2020, 1, 100057. [Google Scholar] [CrossRef]
- Bailey, C.J. Metformin: Historical overview. Diabetologia 2017, 60, 1566–1576. [Google Scholar] [CrossRef] [Green Version]
- Modak, M.; Dixit, P.; Londhe, J.; Ghaskadbi, S.; Devasagayam, T.P. Indian herbs and herbal drugs used for the treatment of diabetes. J. Clin. Biochem. Nutr. 2007, 40, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.D. C, Metformin: Its botanical background. Pract. Diabetes Int. 2004, 21, 15–117. [Google Scholar] [CrossRef]
- White, J.R., Jr. A Brief History of the Development of Diabetes Medications. Diabetes Spectr. Publ. Am. Diabetes Assoc. 2014, 27, 82–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García Rubiño, M.E.; Carrillo, E.; Ruiz Alcalá, G.; Domínguez-Martín, A.; J, A.M.; Boulaiz, H. Phenformin as an Anticancer Agent: Challenges and Prospects. Int. J. Mol. Sci. 2019, 20, 3316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Model Lists of Essential Medicines; World Health Organization: Geneva, Switzerland, 2022.
- Yang, X.; Xu, Z.; Zhang, C.; Cai, Z.; Zhang, J. Metformin, beyond an insulin sensitizer, targeting heart and pancreatic β cells. Biochim. Et Biophys. Acta. Mol. Basis Dis. 2017, 1863, 1984–1990. [Google Scholar] [CrossRef] [PubMed]
- Adeshirlarijaney, A.; Zou, J.; Tran, H.Q.; Chassaing, B.; Gewirtz, A.T. Amelioration of metabolic syndrome by metformin associates with reduced indices of low-grade inflammation independently of the gut microbiota. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E1121–E1130. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Kimura, T.; Obata, A.; Shimoda, M.; Kaku, K. Multifaceted Mechanisms of Action of Metformin Which Have Been Unraveled One after Another in the Long History. Int. J. Mol. Sci. 2021, 22, 2596. [Google Scholar] [CrossRef]
- LaMoia, T.E.; Shulman, G.I. Cellular and Molecular Mechanisms of Metformin Action. Endocr. Rev. 2021, 42, 77–96. [Google Scholar] [CrossRef]
- Nyambuya, T.M.; Dludla, P.V.; Mxinwa, V.; Mokgalaboni, K.; Ngcobo, S.R.; Tiano, L.; Nkambule, B.B. The impact of metformin and aspirin on T-cell mediated inflammation: A systematic review of in vitro and in vivo findings. Life Sci. 2020, 255, 117854. [Google Scholar] [CrossRef]
- Dludla, P.V.; Silvestri, S.; Orlando, P.; Gabuza, K.B.; Mazibuko-Mbeje, S.E.; Nyambuya, T.M.; Mxinwa, V.; Mokgalaboni, K.; Johnson, R.; Muller, C.J.F.; et al. Exploring the Comparative Efficacy of Metformin and Resveratrol in the Management of Diabetes-associated Complications: A Systematic Review of Preclinical Studies. Nutrients 2020, 12, 739. [Google Scholar] [CrossRef] [Green Version]
- Agius, L.; Ford, B.E.; Chachra, S.S. The Metformin Mechanism on Gluconeogenesis and AMPK Activation: The Metabolite Perspective. Int. J. Mol. Sci. 2020, 21, 3240. [Google Scholar] [CrossRef]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef]
- Steinberg, G.R.; Carling, D. AMP-activated protein kinase: The current landscape for drug development. Nat. Rev. Drug Discov. 2019, 18, 527–551. [Google Scholar] [CrossRef] [PubMed]
- Nyambuya, T.M.; Dludla, P.V.; Nkambule, B.B. The aberrant expression of CD69 on peripheral T-helper cells in diet-induced inflammation is ameliorated by low-dose aspirin and metformin treatment. Cell. Immunol. 2021, 363, 104313. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, T.; Galiero, R.; Caturano, A.; Vetrano, E.; Rinaldi, L.; Coviello, F.; Di Martino, A.; Albanese, G.; Marfella, R.; Sardu, C.; et al. Effects of Metformin in Heart Failure: From Pathophysiological Rationale to Clinical Evidence. Biomolecules 2021, 11, 1834. [Google Scholar] [CrossRef] [PubMed]
- Sardu, C.; D’Onofrio, N.; Torella, M.; Portoghese, M.; Mureddu, S.; Loreni, F.; Ferraraccio, F.; Panarese, I.; Trotta, M.C.; Gatta, G.; et al. Metformin Therapy Effects on the Expression of Sodium-Glucose Cotransporter 2, Leptin, and SIRT6 Levels in Pericoronary Fat Excised from Pre-Diabetic Patients with Acute Myocardial Infarction. Biomedicines 2021, 9, 904. [Google Scholar] [CrossRef]
- Jiang, L.L.; Liu, L. Effect of metformin on stem cells: Molecular mechanism and clinical prospect. World J. Stem Cells 2020, 12, 1455–1473. [Google Scholar] [CrossRef]
- Chen, S.C.; Brooks, R.; Houskeeper, J.; Bremner, S.K.; Dunlop, J.; Viollet, B.; Logan, P.J.; Salt, I.P.; Ahmed, S.F.; Yarwood, S.J. Metformin suppresses adipogenesis through both AMP-activated protein kinase (AMPK)-dependent and AMPK-independent mechanisms. Mol. Cell. Endocrinol. 2017, 440, 57–68. [Google Scholar] [CrossRef]
- Pastor-Villaescusa, B.; Cañete, M.D.; Caballero-Villarraso, J.; Hoyos, R.; Latorre, M.; Vázquez-Cobela, R.; Plaza-Díaz, J.; Maldonado, J.; Bueno, G.; Leis, R.; et al. Metformin for Obesity in Prepubertal and Pubertal Children: A Randomized Controlled Trial. Pediatrics 2017, 140, e20164285. [Google Scholar] [CrossRef] [Green Version]
- Lyu, X.; Lyu, T.; Wang, X.; Zhu, H.; Pan, H.; Wang, L.; Yang, H.; Gong, F. The Antiobesity Effect of GLP-1 Receptor Agonists Alone or in Combination with Metformin in Overweight /Obese Women with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. Int. J. Endocrinol. 2021, 2021, 6616693. [Google Scholar] [CrossRef]
- Silamiķele, L.; Silamiķelis, I.; Ustinova, M.; Kalniņa, Z.; Elbere, I.; Petrovska, R.; Kalniņa, I.; Kloviņš, J. Metformin Strongly Affects Gut Microbiome Composition in High-Fat Diet-Induced Type 2 Diabetes Mouse Model of Both Sexes. Front. Endocrinol. 2021, 12, 626359. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, N. Effects of Metformin on the Gut Microbiota in Obesity and Type 2 Diabetes Mellitus. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 5003–5014. [Google Scholar] [CrossRef]
- Oliveira, F.R.; Mamede, M.; Bizzi, M.F.; Rocha, A.L.L.; Ferreira, C.N.; Gomes, K.B.; Cândido, A.L.; Reis, F.M. Effects of Short Term Metformin Treatment on Brown Adipose Tissue Activity and Plasma Irisin Levels in Women with Polycystic Ovary Syndrome: A Randomized Controlled Trial. Horm. Metab. Res. = Horm.-Und Stoffwechs. = Horm. Metab. 2020, 52, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Vangoori, Y.; Suresh, B.S.; Midde, M.L.; Anusha, D.; Uppala, P.K. A review on drug induced obesity and rodent experimental models of obesity in animals. Maedica (Bucur) 2022, 17(3), 706–713. [Google Scholar] [PubMed]
- Kanasaki, K.; Koya, D. Biology of obesity: Lessons from animal models of obesity. J. Biomed. Biotechnol. 2011, 2011, 197636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dludla, P.V.; Mazibuko-Mbeje, S.E.; Nyambuya, T.M.; Mxinwa, V.; Tiano, L.; Marcheggiani, F.; Cirilli, I.; Louw, J.; Nkambule, B.B. The beneficial effects of N-acetyl cysteine (NAC) against obesity associated complications: A systematic review of pre-clinical studies. Pharmacol. Res. 2019, 146, 104332. [Google Scholar] [CrossRef]
- Brozek, J. Body composition: Models and estimation equations. Am. J. Phys. Anthropol. 1966, 24, 239–246. [Google Scholar] [CrossRef]
- de Moura, E.D.M.; Dos Reis, S.A.; da Conceição, L.L.; Sediyama, C.; Pereira, S.S.; de Oliveira, L.L.; Gouveia Peluzio, M.D.C.; Martinez, J.A.; Milagro, F.I. Diet-induced obesity in animal models: Points to consider and influence on metabolic markers. Diabetol. Metab. Syndr. 2021, 13, 32. [Google Scholar] [CrossRef]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schürmann, A.; et al. Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Rouru, J.; Isaksson, K.; Santti, E.; Huupponen, R.; Koulu, M. Metformin and brown adipose tissue thermogenetic activity in genetically obese Zucker rats. Eur. J. Pharmacol. 1993, 246, 67–71. [Google Scholar] [CrossRef]
- Kumar, V.B.; Bernardo, A.E.; Vyas, K.; Franko, M.; Farr, S.; Lakshmanan, L.; Buddhiraju, C.; Morley, J.E. Effect of metformin on nitric oxide synthase in genetically obese (ob/ob) mice. Life Sci. 2001, 69, 2789–2799. [Google Scholar] [CrossRef]
- Klein, J.; Westphal, S.; Kraus, D.; Meier, B.; Perwitz, N.; Ott, V.; Fasshauer, M.; Klein, H.H. Metformin inhibits leptin secretion via a mitogen-activated protein kinase signalling pathway in brown adipocytes. J. Endocrinol. 2004, 183, 299–307. [Google Scholar] [CrossRef]
- Le Gall, M.; Chambard, J.C.; Breittmayer, J.P.; Grall, D.; Pouysségur, J.; Van Obberghen-Schilling, E. The p42/p44 MAP kinase pathway prevents apoptosis induced by anchorage and serum removal. Mol. Biol. Cell 2000, 11, 1103–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Young, A.J.; Ehli, E.A.; Nowotny, D.; Davies, P.S.; Droke, E.A.; Soundy, T.J.; Davies, G.E. Metformin and berberine prevent olanzapine-induced weight gain in rats. PLoS ONE 2014, 9, e93310. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liang, X.; Sun, X.; Zhang, L.; Fu, X.; Rogers, C.J.; Berim, A.; Zhang, S.; Wang, S.; Wang, B.; et al. AMPK/α-Ketoglutarate Axis Dynamically Mediates DNA Demethylation in the Prdm16 Promoter and Brown Adipogenesis. Cell Metab. 2016, 24, 542–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savontaus, E.; Rouru, J.; Boss, O.; Huupponen, R.; Koulu, M. Differential regulation of uncoupling proteins by chronic treatments with beta 3-adrenergic agonist BRL 35135 and metformin in obese fa/fa Zucker rats. Biochem. Biophys. Res. Commun. 1998, 246, 899–904. [Google Scholar] [CrossRef]
- Geerling, J.J.; Boon, M.R.; van der Zon, G.C.; van den Berg, S.A.; van den Hoek, A.M.; Lombès, M.; Princen, H.M.; Havekes, L.M.; Rensen, P.C.; Guigas, B. Metformin lowers plasma triglycerides by promoting VLDL-triglyceride clearance by brown adipose tissue in mice. Diabetes 2014, 63, 880–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, K.; Valdez, J.; Nguyen, J.; Vawter, M.; Galke, B.; Kurtz, T.W.; Chan, J.Y. Increased Energy Expenditure, Ucp1 Expression, and Resistance to Diet-induced Obesity in Mice Lacking Nuclear Factor-Erythroid-2-related Transcription Factor-2 (Nrf2). J. Biol. Chem. 2016, 291, 7754–7766. [Google Scholar] [CrossRef] [Green Version]
- Cantó, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef] [Green Version]
- Dludla, P.V.; Nkambule, B.B.; Tiano, L.; Louw, J.; Jastroch, M.; Mazibuko-Mbeje, S.E. Uncoupling proteins as a therapeutic target to protect the diabetic heart. Pharmacol. Res. 2018, 137, 11–24. [Google Scholar] [CrossRef]
- Day, E.A.; Ford, R.J.; Steinberg, G.R. AMPK as a Therapeutic Target for Treating Metabolic Diseases. Trends Endocrinol. Metab. TEM 2017, 28, 545–560. [Google Scholar] [CrossRef]
- Mehdi, F.; Keihan, G.S.; Asadollah, A.S.; Effat, F. The Effects of Resveratrol, Metformin, Cold and Strength Training on the Level of Perilipin 5 in the Heart, Skeletal Muscle and Brown Adipose Tissues in Mouse. Cell Biochem. Biophys. 2018, 76, 471–476. [Google Scholar] [CrossRef]
- Kim, E.K.; Lee, S.H.; Lee, S.Y.; Kim, J.K.; Jhun, J.Y.; Na, H.S.; Kim, S.Y.; Choi, J.Y.; Yang, C.W.; Park, S.H.; et al. Metformin ameliorates experimental-obesity-associated autoimmune arthritis by inducing FGF21 expression and brown adipocyte differentiation. Exp. Mol. Med. 2018, 50, e432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mthembu, S.X.H.; Dludla, P.V.; Nyambuya, T.M.; Kappo, A.P.; Madoroba, E.; Ziqubu, K.; Nyawo, T.A.; Nkambule, B.B.; Silvestri, S.; Muller, C.J.F.; et al. Experimental models of lipid overload and their relevance in understanding skeletal muscle insulin resistance and pathological changes in mitochondrial oxidative capacity. Biochimie 2021, 196, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, C.; Sul, H.S. Signaling Pathways Regulating Thermogenesis. Front. Endocrinol. 2021, 12, 595020. [Google Scholar] [CrossRef] [PubMed]
- Tezze, C.; Romanello, V.; Sandri, M. FGF21 as Modulator of Metabolism in Health and Disease. Front. Physiol. 2019, 10, 419. [Google Scholar] [CrossRef]
- Gallardo-Montejano, V.I.; Yang, C.; Hahner, L.; McAfee, J.L.; Johnson, J.A.; Holland, W.L.; Fernandez-Valdivia, R.; Bickel, P.E. Perilipin 5 links mitochondrial uncoupled respiration in brown fat to healthy white fat remodeling and systemic glucose tolerance. Nat. Commun. 2021, 12, 3320. [Google Scholar] [CrossRef]
- Abdel-Rehim, W.M.; El-Tahan, R.A.; El-Tarawy, M.A.; Shehata, R.R.; Kamel, M.A. The possible antidiabetic effects of vitamin D receptors agonist in rat model of type 2 diabetes. Mol. Cell. Biochem. 2019, 450, 105–112. [Google Scholar] [CrossRef]
- Stojnić, B.; Serrano, A.; Sušak, L.; Palou, A.; Bonet, M.L.; Ribot, J. Protective Effects of Individual and Combined Low Dose Beta-Carotene and Metformin Treatments against High-Fat Diet-Induced Responses in Mice. Nutrients 2021, 13, 3607. [Google Scholar] [CrossRef] [PubMed]
- Pescador, N.; Francisco, V.; Vázquez, P.; Esquinas, E.M.; González-Páramos, C.; Valdecantos, M.P.; García-Martínez, I.; Urrutia, A.A.; Ruiz, L.; Escalona-Garrido, C.; et al. Metformin reduces macrophage HIF1α-dependent proinflammatory signaling to restore brown adipocyte function in vitro. Redox Biol. 2021, 48, 102171. [Google Scholar] [CrossRef]
- Novitzke, J.M. The significance of clinical trials. J. Vasc. Interv. Neurol. 2008, 1, 31. [Google Scholar]
- U.S. Food and Drug Administration. FDA Drug Safety Communication: FDA Revises Warnings Regarding Use of the Diabetes Medicine Metformin in Certain Patients with Reduced Kidney Function; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2022.
- Davidson, M.B.; Peters, A.L. An overview of metformin in the treatment of type 2 diabetes mellitus. Am. J. Med. 1997, 102, 99–110. [Google Scholar] [CrossRef]
- Rena, G.; Hardie, D.G.; Pearson, E.R. The mechanisms of action of metformin. Diabetologia 2017, 60, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Srinivasa, S.; Wong, K.; Fitch, K.V.; Wei, J.; Petrow, E.; Cypess, A.M.; Torriani, M.; Grinspoon, S.K. Effects of lifestyle modification and metformin on irisin and FGF21 among HIV-infected subjects with the metabolic syndrome. Clin. Endocrinol. 2015, 82, 678–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author, Year | Experimental Model and Metformin Dose | Main Findings |
|---|---|---|
| Short-term effects of metformin (treatment duration equivalent to 2 weeks or less) | ||
| Rouru et al., 1993 [100] | Obese Zucker rats treated with metformin (dissolved in drinking water) at 320 mg/kg/day for 12 days | Reduced body weight and cumulative food intake; however, did not affect thermogenesis, measured using the binding of [3H]GDP to BAT mitochondria and the expression of uncoupling protein mRNA in brown adipose tissue (BAT). |
| Kumar et al., 2001 [101] | Genetically modified obese (ob/ob) mice treated with metformin (subcutaneously) at 200 mg/kg/day for 10 days | Reduced food intake, but did not affect serum glucose levels. Metformin did not affect the expression of nitric oxide synthase in the BAT of obese mice. |
| Klein et al., 2004 [102] | SV40T-immortalized brown adipocytes from the FVB strain of mice were treated with metformin (500 µM and 1 mM) for 8 days | Dose dependently reduced leptin secretion without affecting adipocyte differentiation. Metformin also acutely stimulated p44/p42 MAP kinase and inhibited leptin secretion in a dose-dependent manner in BAT. |
| Hu et al., 2014 [104] | Olanzapine-induced weigh gain in Sprague Dawley rats treated with metformin (oral gavage) at 300 mg/kg for 2 weeks | Prevented weight gain and loss of BAT. Mechanistically, metformin upregulated BAT genes involved in energy expenditure such as AMP-activated protein kinase (AMPK) and uncoupling protein (UCP)3 and those cited in the regulation of lipid metabolism such as resistin, fatty acid synthase, insulin-induced gene 2, CCAAT/enhancer binding protein alpha (C/EBPa), and peroxisome-proliferator-activated receptor gamma (PPARγ) |
| Yang et al., 2016 [105] | Newborn offspring of C57BL/6 mice fed a high-fat diet (HFD) and injected intraperitoneally with metformin at 250 mg/kg for 15 consecutive days | Rescued obesity-induced suppression of brown adipogenesis and thermogenesis. Metformin also activated AMPKα and upregulated the expression of PR domain containing 16 (Prdm16) in BAT. |
| Tokubuchi et al., 2017 [32] | Sprague Dawley rats were treated with metformin (dissolved in drinking water) at 2.5 mg/mL for 2 weeks | Increased plasma levels of lactate and pyruvate. Metformin also significantly reduced visceral fat mass, upregulated fat oxidation-related enzyme in the liver, UCP1 in BAT, and UCP3 in the skeletal muscle. |
| Breining et al., 2018 [31] | Organic cation transporter (Oct)1/2−/− mice on an FVB background received [11C]-metformin (0.2–1.0 GBq) containing 0.1–0.5 µg/mL metformin for 60 min, whereas, brown adipocytes of human origin were treated with metformin (0, 0.1 or 0.5 mM) for 24 h | Metformin was taken up in murine interscapular BAT depots, and this was associated with increased expression of UCP1. Notably, metformin reduced cellular oxygen consumption in human brown adipocyte cells. |
| Long-term effects of metformin (treatment duration equivalent to >2 weeks) | ||
| Savontaus et al., 1998 [106] | Obese Zucker rats treated with metformin (dissolved in drinking water) at 300 mg/kg/day for 3 weeks | Reduced weight gain, as well as food and water intake; however, did not affect mRNA expressions of UCP1, UCP2, or UCP3 in BAT. The observed effect of metformin on the expression of UCPs was when combined with β3-adrenoceptor agonist (BRL 35135) when administered at 0.5 mg/kg/day. |
| Geerling et al., 2014 [107] | E3L. CETP mice fed a Western-type diet supplemented with 200 mg/kg body weight/day (0.2%, w/w) metformin for 4 weeks | Lowered plasma total cholesterol and triglyceride levels, in addition to reducing BAT mass and lipid droplet. This effect was linked to increases in AMP-activated protein kinase a1 (AMPKa1) expression and activity, including hormone-sensitive lipase and mitochondrial respiratory chain complexes in BAT. |
| Liang et al., 2016 [33] | Offspring of C57/BL mice fed an HFD and treated with metformin (dissolved in saline) at 200 mg/kg for 21 days | Decreased serotonin concentration and promoted BAT thermogenic activity by upregulating the expression of UCP1. |
| Kim et al., 2016 [30] | C57BL/6 mice fed an HFD and treated with metformin (oral gavage) at 10 mg/kg or 50 mg/kg for 14 weeks | Improved glucose metabolism and suppressed white adipocyte differentiation via induction of fibroblast growth factor (FGF) 21 in the liver and in white adipocytes. |
| Mehdi et al., 2018 [112] | C57BL/6 J mice treated with metformin (oral gavage) at 250 mg/kg/day for 45 days | Significantly increased the mRNA expression of perilipin 5 in BAT. |
| Kim et al., 2018 [113] | Collagen-induced arthritis DBA/1J mice treated with metformin (oral gavage) at 50 mg/kg for 13 weeks | Displayed a small normalizing effect on the metabolic profile of obese mice. In addition, metformin promoted BAT differentiation while increasing the production of pAMPKα and fibroblast growth factor 21 (FGF21). |
| Karise et al., 2019 [34] | C57Bl/6 mice fed a HFD and treated with metformin (oral gavage) at 250 mg/kg/day for 8 weeks | Increased BAT content and function, as shown by an increase in adipocyte proliferation and differentiation. Metformin further promoted the activation of AMPK and enhanced thermogenic markers (UCP1 and PGC1-α) through adrenergic stimuli and FGF21. Metformin also improved mitochondrial biogenesis in BAT by upregulating nuclear respiratory factor (NRF) 1 and transcription factor A, mitochondrial (TFAM). |
| Yuan et al., 2019 [29] | C57BL/6J mice fed and treated with metformin (oral gavage) at 200 mg/kg/day for 8 weeks | Improved the body weight and insulin sensitivity, while affecting differential expression of 3486 proteins in BAT that were mainly assigned to the pathways of EIF2 signaling and mitochondrial dysfunction. Furthermore, carnitine palmitoyltransferase (CPT)1b and CPT2 in BAT were downregulated by metformin significantly. |
| Abdel-Rehim et al., 2019 [118] | Sprague Dawley rats fed an HFD and treated with metformin (oral gavage) at 200 mg/kg/day for 4 weeks | Improved glucose homeostasis and lipid profile parameters. Metformin also significantly reduced the expression of SREBP-1c, which regulates lipid synthesis in BAT. |
| Stojnic et al., 2021 [119] | C57BL/6J mice fed an HFD and metformin (dissolved in drinking water) at 100 mg/kg/day for 4 weeks | Improved glucose control and insulin sensitivity. Treatment did not affect energy intake, but increased systemic energy expenditure and BAT activation |
| Pescador et al., 2021 [120] | C75BL/6J mice fed HFD and treated with metformin (oral gavage) at 100 mg/kg/day for 6 weeks | Reduced inflammatory features in BAT such as macrophage infiltration, proinflammatory signaling, and gene expression and restored the response to cold exposure. Furthermore, suppressed a HIF1α-dependent pro-inflammatory program that was likely responsible for a secondary beneficial effect on insulin-mediated glucose uptake and β-adrenergic responses in BAT. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziqubu, K.; Mazibuko-Mbeje, S.E.; Mthembu, S.X.H.; Mabhida, S.E.; Jack, B.U.; Nyambuya, T.M.; Nkambule, B.B.; Basson, A.K.; Tiano, L.; Dludla, P.V. Anti-Obesity Effects of Metformin: A Scoping Review Evaluating the Feasibility of Brown Adipose Tissue as a Therapeutic Target. Int. J. Mol. Sci. 2023, 24, 2227. https://doi.org/10.3390/ijms24032227
Ziqubu K, Mazibuko-Mbeje SE, Mthembu SXH, Mabhida SE, Jack BU, Nyambuya TM, Nkambule BB, Basson AK, Tiano L, Dludla PV. Anti-Obesity Effects of Metformin: A Scoping Review Evaluating the Feasibility of Brown Adipose Tissue as a Therapeutic Target. International Journal of Molecular Sciences. 2023; 24(3):2227. https://doi.org/10.3390/ijms24032227
Chicago/Turabian StyleZiqubu, Khanyisani, Sithandiwe E. Mazibuko-Mbeje, Sinenhlanhla X. H. Mthembu, Sihle E. Mabhida, Babalwa U. Jack, Tawanda M. Nyambuya, Bongani B. Nkambule, Albertus K. Basson, Luca Tiano, and Phiwayinkosi V. Dludla. 2023. "Anti-Obesity Effects of Metformin: A Scoping Review Evaluating the Feasibility of Brown Adipose Tissue as a Therapeutic Target" International Journal of Molecular Sciences 24, no. 3: 2227. https://doi.org/10.3390/ijms24032227
APA StyleZiqubu, K., Mazibuko-Mbeje, S. E., Mthembu, S. X. H., Mabhida, S. E., Jack, B. U., Nyambuya, T. M., Nkambule, B. B., Basson, A. K., Tiano, L., & Dludla, P. V. (2023). Anti-Obesity Effects of Metformin: A Scoping Review Evaluating the Feasibility of Brown Adipose Tissue as a Therapeutic Target. International Journal of Molecular Sciences, 24(3), 2227. https://doi.org/10.3390/ijms24032227