
Fiber-Type Shifting in Sarcopenia of Old Age: Proteomic Profiling of the Contractile Apparatus of Skeletal Muscles


Abstract
:1. Introduction
2. Proteomic Profiling of Skeletal Muscle Tissues
2.1. Proteomic Analysis Platforms and Associated Biochemical and Cell Biological Methodology
2.1.1. Two-Dimensional Gel Electrophoresis
2.1.2. Antibody-Based Methodology
2.1.3. Sample Preparation for Proteomic Analysis
2.1.4. Protein Digestion for Peptide Mass Spectrometry
2.1.5. Mass Spectrometric Analysis
2.1.6. Data Acquisition by Mass Spectrometry
2.1.7. Single-Cell Proteomics
2.1.8. Aptamer-Based Proteomics
2.2. Proteomic Profiling of Fiber-Type Specification in Skeletal Muscles
2.3. Composition of the Acto-Myosin Apparatus and Its Proteomic Profile
3. Proteomics of Age-Related Muscle Wasting
3.1. Pathobiological Hallmarks of Sarcopenia of Old Age
3.2. Proteomics of Aged Skeletal Muscle
4. Age-Related Muscular Atrophy, Biomarker Discovery and Therapeutic Approaches
4.1. Mechanisms of Age-Related Muscular Atrophy
- Progressive neurodegeneration: loss of neuromuscular junction integrity; degeneration of motor neurons and resulting denervation; faulty patterns of reinnervation; loss of entire motor units;
- Excitation–contraction uncoupling at the level of the transverse tubules, triad junction and sarcoplasmic reticulum;
- Impaired calcium homeostasis;
- Abnormal mitochondrial functioning;
- Fast-to-slow transitions due to increased susceptibility of fast fibers to atrophy;
- Tendency of bioenergetic glycolytic-to-oxidative shifting;
- Increased cellular stress due to proteotoxic abnormalities;
- Abnormal protein turnover and synthesis causing dysregulated proteostasis;
- Hormonal imbalance and disturbed cellular signaling;
- Visceral obesity causing abnormal muscle-fat-axis signaling;
- Metabolic syndrome and insulin resistance;
- Increased levels of reactive myofibrosis triggering loss of fiber elasticity;
- Chronic low-level sterile inflammation;
- Reduced regenerative capacity due to satellite cell exhaustion;
- Epigenetic changes.
4.2. Biomarker Discovery for the Improved Evaluation of Sarcopenia of Old Age
4.3. Therapeutic Approaches to Counteract Age-Related Muscular Atrophy
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACTA | Alpha-actin-1, skeletal muscle |
| ACTC | Alpha-actin-1, cardiac muscle |
| ACTN-2 | Alpha-actinin-2 |
| ACTN-3 | Alpha-actinin-3 |
| AI | Artificial intelligence |
| BAC | Benzyldimethyl-n-hexadecylammonium chloride |
| BAC-DROP | BAC-gel dissolution to digest PAGE-resolved objective proteins |
| BN | Blue native |
| CAF | Carboxy-terminal fragment of agrin |
| CBB | Coomassie brilliant blue |
| CyTOF | Mass cytometry |
| DAPs | Dystrophin-associated proteins |
| DDA | Data-dependent acquisition |
| DG | Dystroglycan |
| DIA | Data-independent acquisition |
| DIGE | Difference gel electrophoresis |
| DL | Deep learning |
| ELISA | Enzyme-linked immunosorbent assays |
| ESI | Electrospray ionization |
| FACS | Fluorescence-activated cell sorting |
| FASP | Filter-aided sample preparation |
| FC | Flow cytometry |
| FFPE | Formalin-fixed, paraffin-embedded |
| GE | Gel electrophoresis |
| GeLC-MS/MS | Gel electrophoresis liquid chromatography mass spectrometry |
| IB | Immunoblotting |
| ICAT | Isotope-coded affinity tags |
| IFM | Immunofluorescence microscopy |
| IHC | Immunohistochemistry |
| iST | In-StageTip |
| iTRAQ | Isobaric tags for relative and absolute quantitation |
| LC | Liquid chromatography |
| MALDI | Matrix-assisted laser desorption/ionization |
| MARP | Muscle ankyrin repeat protein (ANKRD-2) |
| mIHC/IF | Multiplex immunohistochemistry/immunofluorescence |
| ML | Machine learning |
| MLC-2f | Myosin light chain, fast, regulatory (MYL-11) |
| MLC-2s | Myosin light chain, slow/cardiac, regulatory (MYL-2) |
| MLC-1/3f | Myosin light chain, fast, essential (MLC-1 and MLC-3) |
| MLC-1s | Myosin light chain, slow, essential (MLC-1sa and MLC-1sb) |
| MS | Mass spectrometry |
| MS/MS | Tandem mass spectrometry |
| MudPIT | Multidimensional protein identification technology |
| MuSCs | Muscle stem cells |
| MYBP-C1 | Myosin-binding protein C1, slow |
| MYBP-C2 | Myosin-binding protein C2, fast |
| MyHC-1 | Myosin heavy chain, slow type-I (Myosin-7) |
| MyHC-2a | Myosin heavy chain, fast type-IIA (Myosin-2) |
| MyHC-2b | Myosin heavy chain, fast type-IIB (Myosin-4) |
| MyHC-2x | Myosin heavy chain, fast type-IIX (Myosin-1) |
| MyHC-6 | Myosin heavy chain MYH-6 (Myosin-6) |
| MyHC-7B | Myosin heavy chain MYH-7B (Myosin-7B) |
| MyHC-13 | Myosin heavy chain MYH-13, extraocular muscle (Myosin-13) |
| MyHC-14 | Myosin heavy chain MYH-14 (Myosin-14) |
| MyHC-15 | Myosin heavy chain MYH-15 (Myoisn-15) |
| MyHC-16 | Myosin heavy chain MYH-16 (Myosin-16) |
| MyHC-emb | Myosin heavy chain, embryonic muscle, MyHC-3 (Myosin-3), |
| MyHC-neo | Myosin heavy chain, perinatal muscle, MyHC-8 (Myosin-8) |
| MYOM-1 | Myomesin-1 |
| MYOM-2 | Myomesin-2 |
| MYOZ-1 | Myozenin-1 |
| MYOZ-2 | Myozenin-2 |
| MYOZ-3 | Myozenin-3 |
| nAChR | Nicotinic acetylcholine receptor |
| NEB | Nebulin |
| OBSCN | Obscurin |
| PAGE | Polyacrylamide gel electrophoresis |
| PCT | Pressure-cycling technology |
| PRM | Parallel Reaction Monitoring |
| PTM | Post-translational modification |
| sCAF | Serum carboxy-terminal fragment of agrin |
| SCoPE-MS | Single Cell ProtEomics by Mass Spectrometry |
| SCoPE2 | Second-generation protocol called Single Cell ProtEomics |
| SDOC | Sarcopenia Definitions and Outcomes Consortium |
| SDS | Sodium dodecyl sulfate |
| SILAC | Stable isotope labelling by amino acids in cell culture |
| SP3 | Single-pot solid-phase-enhanced sample preparation |
| SRM/MRM | Selected/Multiple Reaction Monitoring |
| SWATH-MS | Sequential window acquisition of all theoretical mass spectra |
| TCAP | Telethonin/titin-cap |
| TDA | Targeted data acquisition |
| TMT | Tandem mass tag |
| TNC-1 | Troponin TnC, slow/cardiac |
| TNC-2 | Troponin TnC, skeletal muscle |
| TNI-1 | Troponin TnI, slow muscle |
| TNI-2 | Troponin TnI, fast muscle |
| TNT-1 | Troponin TnT, slow muscle |
| TNT-3 | Troponin TnT, fast muscle |
| TOF | Time-of-flight |
| TPM-1 | Alpha-1-tropomyosin |
| TPM-2 | Beta-tropomyosin, slow muscle |
| TPM-3 | Alpha-3-tropomyosin, muscle |
| TPM-4 | Alpha-4-tropomyosin, cytoskeletal |
| TTN | Titin |
| USP3 | Universal solid-phase protein preparation |
References
- Bonaldo, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Model. Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, R.Y.; Li, J.; Dai, Q.; Li, Q.; Yang, J. Muscle Atrophy: Present and Future. Adv. Exp. Med. Biol. 2018, 1088, 605–624. [Google Scholar] [PubMed]
- Capri, M.; Morsiani, C.; Santoro, A.; Moriggi, M.; Conte, M.; Martucci, M.; Bellavista, E.; Fabbri, C.; Giampieri, E.; Albracht, K.; et al. Recovery from 6-month spaceflight at the International Space Station: Muscle-related stress into a proinflammatory setting. FASEB J. 2019, 33, 5168–5180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Huang, Y.; Chen, Y.; Shen, X.; Pan, H.; Yu, W. Impact of Muscle Mass on Survival in Patients with Sepsis: A Systematic Review and Meta-Analysis. Ann. Nutr. Metab. 2021, 77, 330–336. [Google Scholar] [CrossRef]
- Cohen, S.; Nathan, J.A.; Goldberg, A.L. Muscle wasting in disease: Molecular mechanisms and promising therapies. Nat. Rev. Drug Discov. 2015, 14, 58–74. [Google Scholar] [CrossRef]
- Shum, A.M.Y.; Poljak, A.; Bentley, N.L.; Turner, N.; Tan, T.C.; Polly, P. Proteomic profiling of skeletal and cardiac muscle in cancer cachexia: Alterations in sarcomeric and mitochondrial protein expression. Oncotarget 2018, 9, 22001–22022. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Li, N.; Jia, W.; Wang, N.; Liang, M.; Yang, X.; Du, G. Skeletal muscle atrophy: From mechanisms to treatments. Pharmacol. Res. 2021, 172, 105807. [Google Scholar] [CrossRef]
- Feldman, E.L.; Goutman, S.A.; Petri, S.; Mazzini, L.; Savelieff, M.G.; Shaw, P.J.; Sobue, G. Amyotrophic lateral sclerosis. Lancet 2022, 400, 1363–1380. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet 2019, 393, 2636–2646. [Google Scholar] [CrossRef]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef]
- Nishikawa, H.; Fukunishi, S.; Asai, A.; Yokohama, K.; Nishiguchi, S.; Higuchi, K. Pathophysiology and mechanisms of primary sarcopenia (Review). Int. J. Mol. Med. 2021, 48, 156. [Google Scholar] [CrossRef]
- Lippi, G.; Sanchis-Gomar, F.; Montagnana, M. Biological markers in older people at risk of mobility limitations. Curr. Pharm. Des. 2014, 20, 3222–3244. [Google Scholar] [CrossRef]
- Dhillon, R.J.; Hasni, S. Pathogenesis and Management of Sarcopenia. Clin. Geriatr. Med. 2017, 33, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Zullo, A.; Fleckenstein, J.; Schleip, R.; Hoppe, K.; Wearing, S.; Klingler, W. Structural and Functional Changes in the Coupling of Fascial Tissue, Skeletal Muscle, and Nerves During Aging. Front. Physiol. 2020, 11, 592. [Google Scholar] [CrossRef]
- Cho, M.R.; Lee, S.; Song, S.K. A Review of Sarcopenia Pathophysiology, Diagnosis, Treatment and Future Direction. J. Korean Med. Sci. 2022, 37, e146. [Google Scholar] [CrossRef]
- Lynch, D.H.; Spangler, H.B.; Franz, J.R.; Krupenevich, R.L.; Kim, H.; Nissman, D.; Zhang, J.; Li, Y.Y.; Sumner, S.; Batsis, J.A. Multimodal Diagnostic Approaches to Advance Precision Medicine in Sarcopenia and Frailty. Nutrients 2022, 14, 1384. [Google Scholar] [CrossRef]
- Dos Santos, L.; Cyrino, E.S.; Antunes, M.; Santos, D.A.; Sardinha, L.B. Sarcopenia and physical independence in older adults: The independent and synergic role of muscle mass and muscle function. J. Cachexia Sarcopenia Muscle 2017, 8, 245–250. [Google Scholar] [CrossRef]
- Yeung, S.S.Y.; Reijnierse, E.M.; Pham, V.K.; Trappenburg, M.C.; Lim, W.K.; Meskers, C.G.M.; Maier, A.B. Sarcopenia and its association with falls and fractures in older adults: A systematic review and meta-analysis. J. Cachexia Sarcopenia Muscle 2019, 10, 485–500. [Google Scholar] [CrossRef] [Green Version]
- Dent, E.; Martin, F.C.; Bergman, H.; Woo, J.; Romero-Ortunom, R.; Walston, J.D. Management of frailty: Opportunities, challenges, and future directions. Lancet 2019, 394, 1376–1386. [Google Scholar] [CrossRef]
- Smith, L.; Sanchez, G.F.L.; Veronese, N.; Soysal, P.; Kostev, K.; Jacob, L.; Oh, H.; Tully, M.A.; Butler, L.; Parsa, A.D.; et al. Association between sarcopenia and quality of life among adults aged ≥ 65 years from low- and middle-income countries. Aging Clin. Exp. Res. 2022, 34, 2779–2787. [Google Scholar] [CrossRef]
- Veronese, N.; Koyanagi, A.; Cereda, E.; Maggi, S.; Barbagallo, M.; Dominguez, L.J.; Smith, L. Sarcopenia reduces quality of life in the long-term: Longitudinal analyses from the English longitudinal study of ageing. Eur. Geriatr. Med. 2022, 13, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Chianca, V.; Albano, D.; Messina, C.; Gitto, S.; Ruffo, G.; Guarino, S.; Del Grande, F.; Sconfienza, L.M. Sarcopenia: Imaging assessment and clinical application. Abdom. Radiol. 2022, 47, 3205–3216. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Landi, F.; Schneider, S.M.; Zúñiga, C.; Arai, H.; Boiriem, Y.; Chenm, L.K.; Fielding, R.A.; Martin, F.C.; Michel, J.P.; et al. Prevalence of and interventions for sarcopenia in ageing adults: A systematic review. Report of the International Sarcopenia Initiative (EWGSOP and IWGS). Age Ageing 2014, 43, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Writing Group for the European Working Group on Sarcopenia in Older People 2 (EWGSOP2), and the Extended Group for EWGSOP2. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Morley, J.E.; Abbatecola, A.M.; Argiles, J.M.; Baracos, V.; Bauer, J.; Bhasin, S.; Cederholm, T.; Coats, A.J.; Cummings, S.R.; Evans, W.J.; et al. Society on Sarcopenia, Cachexia and Wasting Disorders Trialist Workshop. Sarcopenia with limited mobility: An international consensus. J. Am. Med. Dir. Assoc. 2011, 12, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Studenski, S.A.; Peters, K.W.; Alley, D.E.; Cawthon, P.M.; McLean, R.R.; Harris, T.B.; Ferrucci, L.; Guralnik, J.M.; Fragala, M.S.; Kenny, A.M.; et al. The FNIH sarcopenia project: Rationale, study description, conference recommendations, and final estimates. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 547–558. [Google Scholar] [CrossRef]
- Ackermans, L.L.G.C.; Rabou, J.; Basrai, M.; Schweinlin, A.; Bischoff, S.C.; Cussenot, O.; Cancel-Tassin, G.; Renken, R.J.; Gómez, E.; Sánchez-González, P.; et al. Screening, diagnosis and monitoring of sarcopenia: When to use which tool? Clin. Nutr. ESPEN 2022, 48, 36–44. [Google Scholar] [CrossRef]
- Kirkeby, S.; Garbarsch, C. Aging affects different human muscles in various ways. An image analysis of the histomorphometric characteristics of fiber types in human masseter and vastus lateralis muscles from young adults and the very old. Histol. Histopathol. 2000, 15, 61–71. [Google Scholar]
- Hoffman, J.M.; Lyu, Y.; Pletcher, S.D.; Promislow, D.E.L. Proteomics and metabolomics in ageing research: From biomarkers to systems biology. Essays Biochem. 2017, 61, 379–388. [Google Scholar]
- McHugh, D.; Gil, J. Senescence and aging: Causes, consequences, and therapeutic avenues. J. Cell Biol. 2018, 217, 65–77. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Aunan, J.R.; Watson, M.M.; Hagland, H.R.; Søreide, K. Molecular and biological hallmarks of ageing. Br. J. Surg. 2016, 103, e29–e46. [Google Scholar] [CrossRef] [Green Version]
- Perazza, L.R.; Brown-Borg, H.M.; Thompson, L.V. Physiological Systems in Promoting Frailty. Compr. Physiol. 2022, 12, 3575–3620. [Google Scholar]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef]
- Omholt, S.W.; Kirkwood, T.B.L. Aging as a consequence of selection to reduce the environmental risk of dying. Proc. Natl. Acad. Sci. USA 2021, 118, e2102088118. [Google Scholar] [CrossRef]
- Kowald, A.; Passos, J.F.; Kirkwood, T.B.L. On the evolution of cellular senescence. Aging Cell 2020, 19, e13270. [Google Scholar] [CrossRef]
- Schmeer, C.; Kretz, A.; Wengerodt, D.; Stojiljkovic, M.; Witte, O.W. Dissecting Aging and Senescence-Current Concepts and Open Lessons. Cells 2019, 8, 1446. [Google Scholar] [CrossRef] [Green Version]
- Campisi, J.; Kapahi, P.; Lithgow, G.J.; Melov, S.; Newman, J.C.; Verdin, E. From discoveries in ageing research to therapeutics for healthy ageing. Nature 2019, 571, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Doran, P.; Donoghue, P.; O’Connell, K.; Gannon, J.; Ohlendieck, K. Proteomics of skeletal muscle aging. Proteomics 2009, 9, 989–1003. [Google Scholar] [CrossRef]
- Ohlendieck, K. Proteomics of skeletal muscle differentiation, neuromuscular disorders and fiber aging. Expert Rev. Proteom. 2010, 7, 283–296. [Google Scholar] [CrossRef]
- Baraibar, M.A.; Gueugneau, M.; Duguez, S.; Butler-Browne, G.; Bechet, D.; Friguet, B. Expression and modification proteomics during skeletal muscle ageing. Biogerontology 2013, 14, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Danese, E.; Montagnana, M.; Lippi, G. Proteomics and frailty: A clinical overview. Expert Rev. Proteom. 2018, 15, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Moaddel, R.; Ubaida-Mohien, C.; Tanaka, T.; Lyashkov, A.; Basisty, N.; Schilling, B.; Semba, R.D.; Franceschi, C.; Gorospe, M.; Ferrucci, L. Proteomics in aging research: A roadmap to clinical, translational research. Aging Cell 2021, 20, e13325. [Google Scholar] [CrossRef] [PubMed]
- Aebersold, R.; Mann, M. Mass-spectrometric exploration of proteome structure and function. Nature 2016, 537, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Lill, J.R.; Mathews, W.R.; Rose, C.M.; Schirle, M. Proteomics in the pharmaceutical and biotechnology industry: A look to the next decade. Expert Rev. Proteom. 2021, 18, 503–526. [Google Scholar] [CrossRef]
- Sobsey, C.A.; Ibrahim, S.; Richard, V.R.; Gaspar, V.; Mitsa, G.; Lacasse, V.; Zahedi, R.P.; Batist, G.; Borchers, C.H. Targeted and Untargeted Proteomics Approaches in Biomarker Development. Proteomics 2020, 20, e1900029. [Google Scholar] [CrossRef]
- Mann, S.P.; Treit, P.V.; Geyer, P.E.; Omenn, G.S.; Mann, M. Ethical Principles, Constraints and Opportunities in Clinical Proteomics. Mol. Cell. Proteom. 2021, 20, 100046. [Google Scholar] [CrossRef]
- Rivero-Segura, N.A.; Bello-Chavolla, O.Y.; Barrera-Vázquez, O.S.; Gutierrez-Robledo, L.M.; Gomez-Verjan, J.C. Promising biomarkers of human aging: In search of a multi-omics panel to understand the aging process from a multidimensional perspective. Ageing Res. Rev. 2020, 64, 101164. [Google Scholar] [CrossRef]
- Pan, Y.; Ji, T.; Li, Y.; Ma, L. Omics biomarkers for frailty in older adults. Clin. Chim. Acta. 2020, 510, 363–372. [Google Scholar] [CrossRef]
- Ohlendieck, K. Proteomic Profiling of Fast-To-Slow Muscle Transitions during Aging. Front. Physiol. 2011, 2, 105. [Google Scholar] [CrossRef] [Green Version]
- Dowling, P.; Murphy, S.; Ohlendieck, K. Proteomic profiling of muscle fibre type shifting in neuromuscular diseases. Expert Rev. Proteom. 2016, 13, 783–799. [Google Scholar] [CrossRef]
- Dowling, P.; Zweyer, M.; Swandulla, D.; Ohlendieck, K. Characterization of Contractile Proteins from Skeletal Muscle Using Gel-Based Top-Down Proteomics. Proteomes 2019, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Wasinger, V.C.; Cordwell, S.J.; Cerpa-Poljak, A.; Yan, J.X.; Gooley, A.A.; Wilkins, M.R.; Duncan, M.W.; Harris, R.; Williams, K.L.; Humphery-Smith, I. Progress with gene-product mapping of the Mollicutes: Mycoplasma genitalium. Electrophoresis 1995, 16, 1090–1094. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Sanchez, J.C.; Gooley, A.A.; Appel, R.D.; Humphery-Smith, I.; Hochstrasser, D.F.; Williams, K.L. Progress with proteome projects: Why all proteins expressed by a genome should be identified and how to do it. Biotechnol. Genet. Eng. Rev. 1996, 13, 19–50. [Google Scholar] [CrossRef] [Green Version]
- Manes, N.P.; Nita-Lazar, A. Application of targeted mass spectrometry in bottom-up proteomics for systems biology research. J. Proteom. 2018, 189, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Dupree, E.J.; Jayathirtha, M.; Yorkey, H.; Mihasan, M.; Petre, B.A.; Darie, C.C. A Critical Review of Bottom-Up Proteomics: The Good, the Bad, and the Future of this Field. Proteomes 2010, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Révész, Á.; Hevér, H.; Steckel, A.; Schlosser, G.; Szabó, D.; Vékey, K.; Drahos, L. Collision energies: Optimization strategies for bottom-up proteomics. Mass Spectrom. Rev. 2021, 2, e21763. [Google Scholar] [CrossRef] [PubMed]
- Padula, M.P.; Berry, I.J.; Rourke, M.B.O.; Raymond, B.B.; Santos, J.; Djordjevic, S.P. A Comprehensive Guide for Performing Sample Preparation and Top-Down Protein Analysis. Proteomes 2017, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Cupp-Sutton, K.A.; Wu, S. High-throughput quantitative top-down proteomics. Mol. Omics 2020, 16, 91–99. [Google Scholar] [CrossRef]
- Brown, K.A.; Melby, J.A.; Roberts, D.S.; Ge, Y. Top-down proteomics: Challenges, innovations, and applications in basic and clinical research. Expert Rev. Proteom. 2020, 17, 719–733. [Google Scholar] [CrossRef]
- Carbonara, K.; Andonovski, M.; Coorssen, J.R. Proteomes Are of Proteoforms: Embracing the Complexity. Proteomes 2021, 9, 38. [Google Scholar] [CrossRef]
- Schaffer, L.V.; Millikin, R.J.; Miller, R.M.; Anderson, L.C.; Fellers, R.T.; Ge, Y.; Kelleher, N.L.; LeDuc, R.D.; Liu, X.; Payne, S.H.; et al. Identification and Quantification of Proteoforms by Mass Spectrometry. Proteomics 2019, 19, e1800361. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Z.; Lu, H. Separation methods for system-wide profiling of protein terminome. Proteomics 2022, e2100374, in press. [Google Scholar] [CrossRef]
- Bludau, I.; Aebersold, R. Proteomic and interactomic insights into the molecular basis of cell functional diversity. Nat. Rev. Mol. Cell Biol. 2020, 21, 327–340. [Google Scholar] [CrossRef]
- Ohlendieck, K. Skeletal muscle proteomics: Current approaches, technical challenges and emerging techniques. Skelet. Muscle 2011, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Murphy, S.; Dowling, P.; Ohlendieck, K. Comparative Skeletal Muscle Proteomics Using Two-Dimensional Gel Electrophoresis. Proteomes 2016, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Capitanio, D.; Moriggi, M.; Gelfi, C. Mapping the human skeletal muscle proteome: Progress and potential. Expert Rev. Proteom. 2017, 14, 825–839. [Google Scholar] [CrossRef]
- Hesketh, S.J.; Stansfield, B.N.; Stead, C.A.; Burniston, J.G. The application of proteomics in muscle exercise physiology. Expert Rev. Proteom. 2020, 17, 813–825. [Google Scholar] [CrossRef]
- Domon, B.; Aebersold, R. Mass spectrometry and protein analysis. Science 2006, 312, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fonslow, B.R.; Shan, B.; Baek, M.C.; Yates, J.R., 3rd. Protein analysis by shotgun/bottom-up proteomics. Chem. Rev. 2013, 113, 2343–2394. [Google Scholar] [CrossRef] [Green Version]
- Yates, J.R.; Ruse, C.I.; Nakorchevsky, A. Proteomics by mass spectrometry: Approaches, advances, and applications. Annu. Rev. Biomed. Eng. 2009, 11, 49–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yates, J.R., 3rd. Recent technical advances in proteomics. F1000Res 2019, 8, F1000 Faculty Rev-351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabidó, E.; Selevsek, N.; Aebersold, R. Mass spectrometry-based proteomics for systems biology. Curr. Opin. Biotechnol. 2012, 23, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Omenn, G.S.; Lane, L.; Overall, C.M.; Corrales, F.J.; Schwenk, J.M.; Paik, Y.K.; Van Eyk, J.E.; Liu, S.; Snyder, M.; Baker, M.S.; et al. Progress on Identifying and Characterizing the Human Proteome: 2018 Metrics from the HUPO Human Proteome Project. J. Proteome Res. 2018, 17, 4031–4041. [Google Scholar] [CrossRef]
- Omenn, G.S. Reflections on the HUPO Human Proteome Project, the Flagship Project of the Human Proteome Organization, at 10 Years. Mol. Cell. Proteom. 2021, 20, 100062. [Google Scholar] [CrossRef]
- Omenn, G.S.; Lane, L.; Overall, C.M.; Pineau, C.; Packer, N.H.; Cristea, I.M.; Lindskog, C.; Weintraub, S.T.; Orchard, S.; Roehrl, M.H.A.; et al. The 2022 Report on the Human Proteome from the HUPO Human Proteome Project. J. Proteome Res. 2022, in press. [Google Scholar] [CrossRef]
- Wilhelm, M.; Schlegl, J.; Hahne, H.; Gholami, A.M.; Lieberenz, M.; Savitski, M.M.; Ziegler, E.; Butzmann, L.; Gessulat, S.; Marx, H.; et al. Mass-spectrometry-based draft of the human proteome. Nature 2014, 509, 582–587. [Google Scholar] [CrossRef]
- Kim, M.S.; Pinto, S.M.; Getnet, D.; Nirujogi, R.S.; Manda, S.S.; Chaerkady, R.; Madugundu, A.K.; Kelkar, D.S.; Isserlin, R.; Jain, S.; et al. A draft map of the human proteome. Nature 2014, 509, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, S.; Nice, E.C.; Deutsch, E.W.; Lane, L.; Omenn, G.S.; Pennington, S.R.; Paik, Y.K.; Overall, C.M.; Corrales, F.J.; Cristea, I.M.; et al. A high-stringency blueprint of the human proteome. Nat. Commun. 2020, 11, 5301. [Google Scholar] [CrossRef]
- Westermeier, R. 2D gel-based Proteomics: There’s life in the old dog yet. Arch. Physiol. Biochem. 2016, 122, 236–237. [Google Scholar] [CrossRef]
- Zhan, X.; Li, B.; Zhan, X.; Schlüter, H.; Jungblut, P.R.; Coorssen, J.R. Innovating the Concept and Practice of Two-Dimensional Gel Electrophoresis in the Analysis of Proteomes at the Proteoform Level. Proteomes 2019, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Marcus, K.; Lelong, C.; Rabilloud, T. What Room for Two-Dimensional Gel-Based Proteomics in a Shotgun Proteomics World? Proteomes 2020, 8, 17. [Google Scholar] [CrossRef]
- Friedman, D.B.; Hoving, S.; Westermeier, R. Isoelectric focusing and two-dimensional gel electrophoresis. Methods Enzymol. 2009, 463, 515–540. [Google Scholar]
- Rabilloud, T.; Chevallet, M.; Luche, S.; Lelong, C. Two-dimensional gel electrophoresis in proteomics: Past, present and future. J. Proteom. 2010, 73, 2064–2077. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, B.M.; Coorssen, J.R.; Martins-de-Souza, D. 2DE: The phoenix of proteomics. J. Proteom. 2014, 104, 140–150. [Google Scholar] [CrossRef]
- Westermeier, R. Looking at proteins from two dimensions: A review on five decades of 2D electrophoresis. Arch. Physiol. Biochem. 2014, 120, 168–172. [Google Scholar] [CrossRef]
- Rabilloud, T.; Lelong, C. Two-dimensional gel electrophoresis in proteomics: A tutorial. J. Proteom. 2011, 74, 1829–1841. [Google Scholar] [CrossRef]
- Lee, P.Y.; Saraygord-Afshari, N.; Low, T.Y. The evolution of two-dimensional gel electrophoresis—From proteomics to emerging alternative applications. J. Chromatogr. A 2020, 1615, 460763. [Google Scholar] [CrossRef]
- Carbonara, K.; Coorssen, J.R. Sometimes faster can be better: Microneedling IPG strips enables higher throughput for integrative top-down proteomics. Proteomics 2023, 23, e2200307. [Google Scholar] [CrossRef]
- Görg, A.; Weiss, W.; Dunn, M.J. Current two-dimensional electrophoresis technology for proteomics. Proteomics 2004, 4, 3665–3685. [Google Scholar] [CrossRef]
- Carrette, O.; Burkhard, P.R.; Sanchez, J.C.; Hochstrasser, D.F. State-of-the-art two-dimensional gel electrophoresis: A key tool of proteomics research. Nat. Protoc. 2006, 1, 812–823. [Google Scholar] [CrossRef] [PubMed]
- Yoneten, K.K.; Kasap, M.; Akpinar, G.; Kanli, A.; Karaoz, E. Comparative Proteomics Analysis of Four Commonly Used Methods for Identification of Novel Plasma Membrane Proteins. J. Membr. Biol. 2019, 252, 587–608. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, R.P.; Moebius, J.; Sickmann, A. Two-dimensional BAC/SDS-PAGE for membrane proteomics. Subcell. Biochem. 2002, 43, 13–20. [Google Scholar]
- Wittig, I.; Braun, H.P.; Schägger, H. Blue native PAGE. Nat. Protoc. 2006, 1, 418–428. [Google Scholar] [CrossRef]
- Fernandez-Vizarra, E.; Zeviani, M. Blue-Native Electrophoresis to Study the OXPHOS Complexes. Methods Mol. Biol. 2021, 2192, 287–311. [Google Scholar]
- Sunderhaus, S.; Eubel, H.; Braun, H.P. Two-dimensional blue native/blue native polyacrylamide gel electrophoresis for the characterization of mitochondrial protein complexes and supercomplexes. Methods Mol. Biol. 2007, 372, 315–324. [Google Scholar]
- Vincis Pereira Sanglard, L.; des Francs-Small, C.C. High-Throughput BN-PAGE for Mitochondrial Respiratory Complexes. Methods Mol. Biol. 2022, 2363, 111–119. [Google Scholar]
- Singh, K.; Duchen, M.R. Analysis of Organization and Activity of Mitochondrial Respiratory Chain Complexes in Primary Fibroblasts Using Blue Native PAGE. Methods Mol. Biol. 2022, 2497, 339–348. [Google Scholar]
- Maguire, P.B.; Briggs, F.N.; Lennon, N.J.; Ohlendieck, K. Oligomerization is an intrinsic property of calsequestrin in normal and transformed skeletal muscle. Biochem. Biophys. Res. Commun. 1997, 240, 721–727. [Google Scholar] [CrossRef]
- Froemming, G.R.; Murray, B.E.; Ohlendieck, K. Self-aggregation of triadin in the sarcoplasmic reticulum of rabbit skeletal muscle. Biochim. Biophys. Acta. 1999, 1418, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Culligan, K.; Banville, N.; Dowling, P.; Ohlendieck, K. Drastic reduction of calsequestrin-like proteins and impaired calcium binding in dystrophic mdx muscle. J. Appl. Physiol. 2002, 92, 435–445. [Google Scholar] [CrossRef]
- Noaman, N.; Coorssen, J.R. Coomassie does it (better): A Robin Hood approach to total protein quantification. Anal. Biochem. 2018, 556, 53–56. [Google Scholar] [CrossRef]
- Noaman, N.; Abbineni, P.S.; Withers, M.; Coorssen, J.R. Coomassie staining provides routine (sub)femtomole in-gel detection of intact proteoforms: Expanding opportunities for genuine Top-down Proteomics. Electrophoresis 2017, 38, 3086–3099. [Google Scholar] [CrossRef]
- Chevalier, F. Standard Dyes for Total Protein Staining in Gel-Based Proteomic Analysis. Materials 2010, 3, 4784–4792. [Google Scholar] [CrossRef] [Green Version]
- Panfoli, I.; Calzia, D.; Santucci, L.; Ravera, S.; Bruschi, M.; Candiano, G. A blue dive: From ‘blue fingers’ to ‘blue silver’. A comparative overview of staining methods for in-gel proteomics. Expert Rev. Proteom. 2012, 9, 627–634. [Google Scholar] [CrossRef]
- Sundaram, P. Protein Stains and Applications. Methods Mol. Biol. 2018, 1853, 1–14. [Google Scholar]
- Meleady, P. Two-Dimensional Gel Electrophoresis and 2D-DIGE. Methods Mol. Biol. 2023, 2596, 3–15. [Google Scholar]
- Murphy, S.; Dowling, P.; Zweyer, M.; Swandulla, D.; Ohlendieck, K. Proteomic profiling of giant skeletal muscle proteins. Expert Rev. Proteom. 2019, 16, 241–256. [Google Scholar] [CrossRef]
- Murphy, S.; Henry, M.; Meleady, P.; Ohlendieck, K. Utilization of dried and long-term stored polyacrylamide gels for the advanced proteomic profiling of mitochondrial contact sites from rat liver. Biol. Methods Protoc. 2018, 3, bpy008. [Google Scholar] [CrossRef]
- Murphy, S.; Ohlendieck, K. Proteomic profiling of large myofibrillar proteins from dried and long-term stored polyacrylamide gels. Anal. Biochem. 2018, 543, 8–11. [Google Scholar] [CrossRef]
- Banks, C.A.; Kong, S.E.; Washburn, M.P. Affinity purification of protein complexes for analysis by multidimensional protein identification technology. Protein Expr. Purif. 2012, 86, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Elschenbroich, S.; Ignatchenko, V.; Sharma, P.; Schmitt-Ulms, G.; Gramolini, A.O.; Kislinger, T. Peptide separations by on-line MudPIT compared to isoelectric focusing in an off-gel format: Application to a membrane-enriched fraction from C2C12 mouse skeletal muscle cells. J. Proteome Res. 2009, 8, 4860–4869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unlü, M.; Morgan, M.E.; Minden, J.S. Difference gel electrophoresis: A single gel method for detecting changes in protein extracts. Electrophoresis 1997, 18, 2071–2077. [Google Scholar] [CrossRef] [PubMed]
- Minden, J.S.; Dowd, S.R.; Meyer, H.E.; Stühler, K. Difference gel electrophoresis. Electrophoresis 2009, 30, S156–S161. [Google Scholar] [CrossRef]
- Arentz, G.; Weiland, F.; Oehler, M.K.; Hoffmann, P. State of the art of 2D DIGE. Proteom. Clin. Appl. 2015, 9, 277–288. [Google Scholar] [CrossRef]
- Timms, J.F.; Cramer, R. Difference gel electrophoresis. Proteomics 2008, 8, 4886–4897. [Google Scholar] [CrossRef]
- Blundon, M.; Ganesan, V.; Redler, B.; Van, P.T.; Minden, J.S. Two-Dimensional Difference Gel Electrophoresis. Methods Mol. Biol. 2019, 1855, 229–247. [Google Scholar]
- Gelfi, C.; Capitanio, D. DIGE Analysis of Clinical Specimens. Methods Mol. Biol. 2023, 2596, 177–199. [Google Scholar]
- Di Luca, A.; Hamill, R.; Mullen, A.M.; Elia, G. DIGE Analysis of Animal Tissues. Methods Mol. Biol. 2023, 2596, 201–216. [Google Scholar]
- Holland, A. Two-Dye Versus Three-Dye DIGE for Comparative Testis Tissue Proteomic Analysis. Methods Mol. Biol. 2023, 2596, 245–263. [Google Scholar]
- Ohlendieck, K. Top-Down Proteomics and Comparative 2D-DIGE Analysis. Methods Mol. Biol. 2023, 2596, 19–38. [Google Scholar]
- Ohlendieck, K. Comparative 3-Sample 2D-DIGE Analysis of Skeletal Muscles. Methods Mol. Biol. 2023, 2596, 127–146. [Google Scholar]
- Tonge, R.; Shaw, J.; Middleton, B.; Rowlinson, R.; Rayner, S.; Young, J.; Pognan, F.; Hawkins, E.; Currie, I.; Davison, M. Validation and development of fluorescence two-dimensional differential gel electrophoresis proteomics technology. Proteomics 2001, 1, 377–396. [Google Scholar] [CrossRef]
- Marouga, R.; David, S.; Hawkins, E. The development of the DIGE system: 2D fluorescence difference gel analysis technology. Anal. Bioanal. Chem. 2005, 382, 669–678. [Google Scholar] [CrossRef]
- Dowling, P. DIGE Analysis Software and Protein Identification Approaches. Methods Mol. Biol. 2023, 2596, 39–50. [Google Scholar]
- Dani, D.; Dencher, N.A. Native DIGE for Quantitative and Functional Analysis of Protein Interactomes. Methods Mol. Biol. 2023, 2596, 53–69. [Google Scholar]
- Ackermann, D.; König, S. Comparative Two-Dimensional Fluorescence Gel Electrophoresis. Methods Mol. Biol. 2023, 2596, 71–81. [Google Scholar]
- Stasyk, T.; Huber, L.A. DIGE-Based Phosphoproteomic Analysis. Methods Mol. Biol. 2023, 2596, 97–104. [Google Scholar]
- Carberry, S.; Zweyer, M.; Swandulla, D.; Ohlendieck, K. Application of fluorescence two-dimensional difference in-gel electrophoresis as a proteomic biomarker discovery tool in muscular dystrophy research. Biology 2013, 2, 1438–1464. [Google Scholar] [CrossRef]
- Forgrave, L.M.; Wang, M.; Yang, D.; DeMarco, M.L. Proteoforms and their expanding role in laboratory medicine. Pract. Lab Med. 2021, 28, e00260. [Google Scholar] [CrossRef]
- Landsberger, M.; Brinkmeier, H. Immunoblot Analysis of DIGE-Based Proteomics. Methods Mol. Biol. 2023, 2596, 429–443. [Google Scholar] [PubMed]
- Zweyer, M.; Ohlendieck, K.; Swandulla, D. Histological and Histochemical Microscopy Used to Verify 2D-DIGE Pathoproteomics. Methods Mol. Biol. 2023, 2596, 465–480. [Google Scholar] [PubMed]
- Zweyer, M.; Ohlendieck, K.; Swandulla, D. Verification of Protein Changes Determined by 2D-DIGE Based Proteomics Using Immunofluorescence Microscopy. Methods Mol. Biol. 2023, 2596, 445–464. [Google Scholar] [PubMed]
- Tabatabaei, M.S.; Ahmed, M. Enzyme-Linked Immunosorbent Assay (ELISA). Methods Mol. Biol. 2022, 2508, 115–134. [Google Scholar]
- Siddiqui, S.; Livák, F. Principles of Advanced Flow Cytometry: A Practical Guide. Methods Mol. Biol. 2023, 2580, 89–114. [Google Scholar]
- Pakula, A.; Spinazzola, J.M.; Gussoni, E. Purification of Myogenic Progenitors from Human Muscle Using Fluorescence-Activated Cell Sorting (FACS). Methods Mol. Biol. 2019, 1889, 1–15. [Google Scholar]
- Behbehani, G.K. Immunophenotyping by Mass Cytometry. Methods Mol. Biol. 2019, 2032, 31–51. [Google Scholar]
- Helali, Y.; Sharma, S.; Vandeput, M.; Welba, D.; Van Antwerpen, P.; Marchant, A.; Delporte, C. Fc Glycosylation Characterization of Human Immunoglobulins G Using Immunocapture and LC-MS. Methods Mol. Biol. 2021, 2271, 57–71. [Google Scholar]
- Ghosh, R.; Gilda, J.E.; Gomes, A.V. The necessity of and strategies for improving confidence in the accuracy of western blots. Expert Rev. Proteom. 2014, 11, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Zweyer, M.; Sabir, H.; Dowling, P.; Gargan, S.; Murphy, S.; Swandulla, D.; Ohlendieck, K. Histopathology of Duchenne muscular dystrophy in correlation with changes in proteomic biomarkers. Histol. Histopathol. 2022, 37, 101–116. [Google Scholar]
- Dowd, A. Elucidating Cellular Metabolism and Protein Difference Data from DIGE Proteomics Experiments Using Enzyme Assays. Methods Mol. Biol. 2023, 2596, 399–419. [Google Scholar]
- Dowd, A. Enzyme Assay Methods to Validate DIGE Proteomics Data. Methods Mol. Biol. 2023, 2596, 421–428. [Google Scholar]
- Mishra, M.; Tiwari, S.; Gomes, A.V. Protein purification and analysis: Next generation Western blotting techniques. Expert Rev. Proteom. 2017, 14, 1037–1053. [Google Scholar] [CrossRef]
- Meola, G. Advanced microscopic and histochemical techniques: Diagnostic tools in the molecular era of myology. Eur. J. Histochem. 2005, 49, 93–96. [Google Scholar]
- Nix, J.S.; Moore, S.A. What Every Neuropathologist Needs to Know: The Muscle Biopsy. J. Neuropathol. Exp. Neurol. 2020, 79, 719–733. [Google Scholar] [CrossRef]
- Su, T.; Wang, S.; Huang, S.; Cai, H.; McKinley, E.T.; Beeghly-Fadiel, A.; Zheng, W.; Shu, X.O.; Cai, Q. Multiplex immunohistochemistry and high-throughput image analysis for evaluation of spatial tumor immune cell markers in human breast cancer. Cancer Biomark. 2022, 35, 193–206. [Google Scholar] [CrossRef]
- Tan, W.C.C.; Nerurkar, S.N.; Cai, H.Y.; Ng, H.H.M.; Wu, D.; Wee, Y.T.F.; Lim, C.T.; Yeong, J.; Lim, T.K.H. Overview of multiplex immunohistochemistry/immunofluorescence techniques in the era of cancer immunotherapy. Cancer Commun. 2020, 40, 135–153. [Google Scholar] [CrossRef] [Green Version]
- Alcazar, J.; Frandsen, U.; Prokhorova, T.; Kamper, R.S.; Haddock, B.; Aagaard, P.; Suetta, C. Changes in systemic GDF15 across the adult lifespan and their impact on maximal muscle power: The Copenhagen Sarcopenia Study. J. Cachexia Sarcopenia Muscle 2021, 12, 1418–1427. [Google Scholar] [CrossRef]
- Mengeste, A.M.; Nikolić, N.; Fernandez, A.D.; Feng, Y.Z.; Nyman, T.A.; Kersten, S.; Haugen, F.; Kase, E.T.; Aas, V.; Rustan, A.C.; et al. Insight Into the Metabolic Adaptations of Electrically Pulse-Stimulated Human Myotubes Using Global Analysis of the Transcriptome and Proteome. Front. Physiol. 2022, 13, 928195. [Google Scholar] [CrossRef]
- Krüger, K.; Seimetz, M.; Ringseis, R.; Wilhelm, J.; Pichl, A.; Couturier, A.; Eder, K.; Weissmann, N.; Mooren, F.C. Exercise training reverses inflammation and muscle wasting after tobacco smoke exposure. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R366–R376. [Google Scholar] [CrossRef]
- San Segundo-Acosta, P.; Montero-Calle, A.; Jernbom-Falk, A.; Alonso-Navarro, M.; Pin, E.; Andersson, E.; Hellström, C.; Sánchez-Martínez, M.; Rábano, A.; Solís-Fernández, G.; et al. Multiomics Profiling of Alzheimer’s Disease Serum for the Identification of Autoantibody Biomarkers. J. Proteome Res. 2021, 20, 5115–5130. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.L.; Shoemaker, M.E.; Gawel, S.; Davis, G.J.; Luo, M.; Mustad, V.A.; Cramer, J.T. Biomarker Changes in Response to a 12-Week Supplementation of an Oral Nutritional Supplement Enriched with Protein, Vitamin D and HMB in Malnourished Community Dwelling Older Adults with Sarcopenia. Nutrients 2022, 14, 1196. [Google Scholar] [CrossRef] [PubMed]
- Assarsson, E.; Lundberg, M.; Holmquist, G.; Björkesten, J.; Thorsen, S.B.; Ekman, D.; Eriksson, A.; Rennel Dickens, E.; Ohlsson, S.; Edfeldt, G.; et al. Homogenous 96-plex PEA immunoassay exhibiting high sensitivity, specificity, and excellent scalability. PLoS One 2014, 9, e95192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Saedi, A.; Phu, S.; Vogrin, S.; Gunawardene, P.; Duque, G. Association between Circulating Osteoprogenitor Cells and Sarcopenia. Gerontology 2022, 68, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Kirk, B.; Zanker, J.; Bani Hassan, E.; Bird, S.; Brennan-Olsen, S.; Duque, G. Sarcopenia Definitions and Outcomes Consortium (SDOC) Criteria are Strongly Associated With Malnutrition, Depression, Falls, and Fractures in High-Risk Older Persons. J. Am. Med. Dir. Assoc. 2021, 22, 741–745. [Google Scholar] [CrossRef]
- Bandura, D.R.; Baranov, V.I.; Ornatsky, O.I.; Antonov, A.; Kinach, R.; Lou, X.; Pavlov, S.; Vorobiev, S.; Dick, J.E.; Tanner, S.D. Mass cytometry: Technique for real time single cell multitarget immunoassay based on inductively coupled plasma time-of-flight mass spectrometry. Anal. Chem. 2009, 81, 6813–6822. [Google Scholar] [CrossRef]
- Tan, T.; Gray, D.H.D.; The, C.E. Single-Cell Profiling of the Intrinsic Apoptotic Pathway by Mass Cytometry (CyTOF). Methods Mol. Biol. 2022, 2543, 83–97. [Google Scholar]
- Porpiglia, E.; Samusik, N.; Ho, A.T.V.; Cosgrove, B.D.; Mai, T.; Davis, K.L.; Jager, A.; Nolan, G.P.; Bendall, S.C.; Fantl, W.J.; et al. High-resolution myogenic lineage mapping by single-cell mass cytometry. Nat. Cell Biol. 2017, 19, 558–567. [Google Scholar] [CrossRef] [Green Version]
- Petrilli, L.L.; Riccio, F.; Giuliani, G.; Palma, A.; Gargioli, C.; Vumbaca, S.; Faron, M.; Palmieri, G.; Pasquini, L.; Sacco, F.; et al. Skeletal Muscle Subpopulation Rearrangements upon Rhabdomyosarcoma Development through Single-Cell Mass Cytometry. J. Clin. Med. 2021, 10, 823. [Google Scholar] [CrossRef]
- Borok, M.; Didier, N.; Gattazzo, F.; Ozturk, T.; Corneau, A.; Rouard, H.; Relaix, F. Progressive and Coordinated Mobilization of the Skeletal Muscle Niche throughout Tissue Repair Revealed by Single-Cell Proteomic Analysis. Cells 2021, 10, 744. [Google Scholar] [CrossRef]
- Porpiglia, E.; Mai, T.; Kraft, P.; Holbrook, C.A.; de Morree, A.; Gonzalez, V.D.; Hilgendorf, K.I.; Frésard, L.; Trejo, A.; Bhimaraju, S.; et al. Elevated CD47 is a hallmark of dysfunctional aged muscle stem cells that can be targeted to augment regeneration. Cell Stem Cell 2022, 29, 1653–1668.e8. [Google Scholar] [CrossRef]
- Swaney, D.L.; Villén, J. Enrichment of Modified Peptides via Immunoaffinity Precipitation with Modification-Specific Antibodies. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot088013. [Google Scholar] [CrossRef]
- Kumar, A.; Baycin-Hizal, D.; Shiloach, J.; Bowen, M.A.; Betenbaugh, M.J. Coupling enrichment methods with proteomics for understanding and treating disease. Proteom. Clin. Appl. 2015, 9, 33–47. [Google Scholar] [CrossRef]
- Beltran, L.; Cutillas, P.R. Advances in phosphopeptide enrichment techniques for phosphoproteomics. Amino Acids 2012, 43, 1009–1024. [Google Scholar] [CrossRef]
- Gargan, S.; Ohlendieck, K. Sample Preparation and Protein Determination for 2D-DIGE Proteomics. Methods Mol. Biol. 2023, 2596, 325–337. [Google Scholar]
- Murphy, S. Subcellular Fractionation for DIGE-Based Proteomics. Methods Mol. Biol. 2023, 2596, 351–362. [Google Scholar]
- Murphy, S.; Zweyer, M.; Henry, M.; Meleady, P.; Mundegar, R.R.; Swandulla, D.; Ohlendieck, K. Proteomic analysis of the sarcolemma-enriched fraction from dystrophic mdx-4cv skeletal muscle. J. Proteom. 2019, 191, 212–227. [Google Scholar] [CrossRef]
- Murphy, S.; Zweyer, M.; Mundegar, R.R.; Swandulla, D.; Ohlendieck, K. Comparative gel-based proteomic analysis of chemically crosslinked complexes in dystrophic skeletal muscle. Electrophoresis 2018, 39, 1735–1744. [Google Scholar] [CrossRef] [Green Version]
- Sielaff, M.; Kuharev, J.; Bohn, T.; Hahlbrock, J.; Bopp, T.; Tenzer, S.; Distler, U. Evaluation of FASP, SP3, and iST Protocols for Proteomic Sample Preparation in the Low Microgram Range. J. Proteome Res. 2017, 16, 4060–4072. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Mann, M. Combination of FASP and StageTip-based fractionation allows in-depth analysis of the hippocampal membrane proteome. J. Proteome Res. 2009, 8, 5674–5678. [Google Scholar] [CrossRef]
- Xie, Z.; Feng, Q.; Zhang, S.; Yan, Y.; Deng, C.; Ding, C.F. Advances in proteomics sample preparation and enrichment for phosphorylation and glycosylation analysis. Proteomics 2022, 22, e2200070. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R. Filter Aided Sample Preparation—A tutorial. Anal. Chim. Acta 2019, 1090, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Kulak, N.A.; Pichler, G.; Paron, I.; Nagaraj, N.; Mann, M. Minimal, encapsulated proteomic-sample processing applied to copy-number estimation in eukaryotic cells. Nat. Methods 2014, 11, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.S.; Moggridge, S.; Müller, T.; Sorensen, P.H.; Morin, G.B.; Krijgsveld, J. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 2019, 14, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Dagley, L.F.; Infusini, G.; Larsen, R.H.; Sandow, J.J.; Webb, A.I. Universal Solid-Phase Protein Preparation (USP3) for Bottom-up and Top-down Proteomics. J. Proteome Res. 2019, 18, 2915–2924. [Google Scholar] [CrossRef] [PubMed]
- Dapic, I.; Baljeu-Neuman, L.; Uwugiaren, N.; Kers, J.; Goodlett, D.R.; Corthals, G.L. Proteome analysis of tissues by mass spectrometry. Mass Spectrom. Rev. 2019, 38, 403–441. [Google Scholar] [CrossRef]
- Cai, X.; Xue, Z.; Wu, C.; Sun, R.; Qian, L.; Yue, L.; Ge, W.; Yi, X.; Liu, W.; Chen, C.; et al. High-throughput proteomic sample preparation using pressure cycling technology. Nat. Protoc. 2022, 17, 2307–2325. [Google Scholar] [CrossRef]
- Vit, O.; Petrak, J. Integral membrane proteins in proteomics. How to break open the black box? J. Proteom. 2017, 153, 8–20. [Google Scholar] [CrossRef]
- Kar, U.K.; Simonian, M.; Whitelegge, J.P. Integral membrane proteins: Bottom-up, top-down and structural proteomics. Expert Rev. Proteom. 2017, 14, 715–723. [Google Scholar] [CrossRef]
- Dafun, A.S.; Marcoux, J. Structural mass spectrometry of membrane proteins. Biochim. Biophys. Acta Proteins Proteom. 2022, 1870, 140813. [Google Scholar] [CrossRef]
- Boeri Erba, E.; Signor, L.; Petosa, C. Exploring the structure and dynamics of macromolecular complexes by native mass spectrometry. J. Proteom. 2020, 222, 103799. [Google Scholar] [CrossRef]
- Tamara, S.; den Boer, M.A.; Heck, A.J.R. High-Resolution Native Mass Spectrometry. Chem. Rev. 2022, 122, 7269–7326. [Google Scholar] [CrossRef]
- Santambrogio, C.; Ponzini, E.; Grandori, R. Native mass spectrometry for the investigation of protein structural (dis)order. Biochim. Biophys. Acta Proteins Proteom. 2022, 1870, 140828. [Google Scholar] [CrossRef]
- Choksawangkarn, W.; Edwards, N.; Wang, Y.; Gutierrez, P.; Fenselau, C. Comparative study of workflows optimized for in-gel, in-solution, and on-filter proteolysis in the analysis of plasma membrane proteins. J. Proteome Res. 2012, 11, 3030–3034. [Google Scholar] [CrossRef] [Green Version]
- Glatter, T.; Ludwig, C.; Ahrné, E.; Aebersold, R.; Heck, A.J.; Schmidt, A. Large-scale quantitative assessment of different in-solution protein digestion protocols reveals superior cleavage efficiency of tandem Lys-C/trypsin proteolysis over trypsin digestion. J. Proteome Res. 2012, 11, 5145–5156. [Google Scholar] [CrossRef]
- Goodman, J.K.; Zampronio, C.G.; Jones, A.M.E.; Hernandez-Fernaud, J.R. Updates of the In-Gel Digestion Method for Protein Analysis by Mass Spectrometry. Proteomics 2018, 18, e1800236. [Google Scholar] [CrossRef] [Green Version]
- Gauci, V.J.; Wright, E.P.; Coorssen, J.R. Quantitative proteomics: Assessing the spectrum of in-gel protein detection methods. J. Chem. Biol. 2011, 4, 3–29. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.; Ohlendieck, K. Mass spectrometric identification of dystrophin isoform Dp427 by on-membrane digestion of sarcolemma from skeletal muscle. Anal. Biochem. 2010, 404, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Staunton, L.; Ohlendieck, K. Mass spectrometric characterization of the sarcoplasmic reticulum from rabbit skeletal muscle by on-membrane digestion. Protein Pept. Lett. 2012, 19, 252–263. [Google Scholar] [CrossRef]
- Murphy, S.; Brinkmeier, H.; Krautwald, M.; Henry, M.; Meleady, P.; Ohlendieck, K. Proteomic profiling of the dystrophin complex and membrane fraction from dystrophic mdx muscle reveals decreases in the cytolinker desmoglein and increases in the extracellular matrix stabilizers biglycan and fibronectin. J. Muscle Res. Cell. Motil. 2017, 38, 251–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, S.; Ohlendieck, K. Protein Digestion for 2D-DIGE Analysis. Methods Mol. Biol. 2023, 2596, 339–349. [Google Scholar] [PubMed]
- Tsiatsiani, L.; Heck, A.J. Proteomics beyond trypsin. FEBS J. 2015, 282, 2612–2626. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, P.; Tsiatsiani, L.; Low, T.Y.; Heck, A.J. Six alternative proteases for mass spectrometry-based proteomics beyond trypsin. Nat. Protoc. 2016, 11, 993–1006. [Google Scholar] [CrossRef]
- Zhang, X. Less is More: Membrane Protein Digestion Beyond Urea-Trypsin Solution for Next-level Proteomics. Mol. Cell. Proteom. 2015, 14, 2441–2453. [Google Scholar] [CrossRef] [Green Version]
- Goldman, A.R.; Beer, L.A.; Tang, H.Y.; Hembach, P.; Zayas-Bazan, D.; Speicher, D.W. Proteome Analysis Using Gel-LC-MS/MS. Curr. Protoc. Protein Sci. 2019, 96, e93. [Google Scholar] [CrossRef]
- Takemori, A.; Ishizaki, J.; Nakashima, K.; Shibata, T.; Kato, H.; Kodera, Y.; Suzuki, T.; Hasegawa, H.; Takemori, N. BAC-DROP: Rapid Digestion of Proteome Fractionated via Dissolvable Polyacrylamide Gel Electrophoresis and Its Application to Bottom-Up Proteomics Workflow. J. Proteome Res. 2021, 20, 1535–1543. [Google Scholar] [CrossRef]
- Rešetar Maslov, D.; Svirkova, A.; Allmaier, G.; Marchetti-Deschamann, M.; Kraljević Pavelić, S. Optimization of MALDI-TOF mass spectrometry imaging for the visualization and comparison of peptide distributions in dry-cured ham muscle fibers. Food Chem. 2019, 283, 275–286. [Google Scholar] [CrossRef]
- Evangelista, A.J.; Ferreira, T.L. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry in the diagnosis of microorganisms. Future Microbiol. 2022, 17, 1409–1419. [Google Scholar] [CrossRef]
- Nesvizhskii, A.I.; Vitek, O.; Aebersold, R. Analysis and validation of proteomic data generated by tandem mass spectrometry. Nat. Methods 2007, 4, 787–797. [Google Scholar] [CrossRef]
- Dowling, P.; Gargan, S.; Zweyer, M.; Henry, M.; Meleady, P.; Swandulla, D.; Ohlendieck, K. Protocol for the Bottom-Up Proteomic Analysis of Mouse Spleen. STAR Protoc. 2020, 1, 100196. [Google Scholar] [CrossRef]
- Théron, L.; Gueugneau, M.; Coudy, C.; Viala, D.; Bijlsma, A.; Butler-Browne, G.; Maier, A.; Béchet, D.; Chambon, C. Label-free quantitative protein profiling of vastus lateralis muscle during human aging. Mol. Cell. Proteom. 2014, 13, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Rauniyar, N.; Yates, J.R., 3rd. Isobaric labeling-based relative quantification in shotgun proteomics. J. Proteome Res. 2014, 13, 5293–5309. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Sun, Y.; Zhang, T.; Shu, L.; Roepstorff, P.; Yang, F. Quantitative Proteomics Using Isobaric Labeling: A Practical Guide. Genom. Proteom. Bioinform. 2021, 19, 689–706. [Google Scholar] [CrossRef]
- Beller, N.C.; Hummon, A.B. Advances in stable isotope labeling: Dynamic labeling for spatial and temporal proteomic analysis. Mol. Omics 2022, 18, 579–590. [Google Scholar] [CrossRef]
- Xing, T.; Wang, C.; Zhao, X.; Dai, C.; Zhou, G.; Xu, X. Proteome Analysis Using Isobaric Tags for Relative and Absolute Analysis Quantitation (iTRAQ) Reveals Alterations in Stress-Induced Dysfunctional Chicken Muscle. J. Agric. Food Chem. 2017, 65, 2913–2922. [Google Scholar] [CrossRef]
- Chahrour, O.; Cobice, D.; Malone, J. Stable isotope labelling methods in mass spectrometry-based quantitative proteomics. J. Pharm. Biomed. Anal. 2015, 113, 2–20. [Google Scholar] [CrossRef]
- Westbrook, J.A.; Noirel, J.; Brown, J.E.; Wright, P.C.; Evans, C.A. Quantitation with chemical tagging reagents in biomarker studies. Proteom. Clin. Appl. 2015, 9, 295–300. [Google Scholar] [CrossRef]
- Chaves, D.F.; Carvalho, P.C.; Lima, D.B.; Nicastro, H.; Lorenzeti, F.M.; Siqueira-Filho, M.; Hirabara, S.M.; Alves, P.H.; Moresco, J.J.; Yates, J.R., 3rd; et al. Comparative proteomic analysis of the aging soleus and extensor digitorum longus rat muscles using TMT labeling and mass spectrometry. J. Proteome Res. 2013, 12, 4532–4546. [Google Scholar] [CrossRef] [Green Version]
- Hoedt, E.; Zhang, G.; Neubert, T.A. Stable Isotope Labeling by Amino Acids in Cell Culture (SILAC) for Quantitative Proteomics. Adv. Exp. Med. Biol. 2019, 1140, 531–539. [Google Scholar]
- Shenoy, A.; Geiger, T. Super-SILAC: Current trends and future perspectives. Expert Rev. Proteom. 2015, 12, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Geiger, T.; Wisniewski, J.R.; Cox, J.; Zanivan, S.; Kruger, M.; Ishihama, Y.; Mann, M. Use of stable isotope labeling by amino acids in cell culture as a spike-in standard in quantitative proteomics. Nat. Protoc. 2011, 6, 147–157. [Google Scholar] [CrossRef]
- Rayavarapu, S.; Coley, W.; Cakir, E.; Jahnke, V.; Takeda, S.; Aoki, Y.; Grodish-Dressman, H.; Jaiswal, J.K.; Hoffman, E.P.; Brown, K.J.; et al. Identification of disease specific pathways using in vivo SILAC proteomics in dystrophin deficient mdx mouse. Mol. Cell. Proteom. 2013, 12, 1061–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, M.V.; Tawalbeh, S.M.; Canessa, E.H.; Hathout, Y. Temporal Proteomic Profiling During Differentiation of Normal and Dystrophin-Deficient Human Muscle Cells. J. Neuromuscul. Dis. 2021, 8, S205–S222. [Google Scholar] [CrossRef] [PubMed]
- Kallabis, S.; Abraham, L.; Müller, S.; Dzialas, V.; Türk, C.; Wiederstein, J.L.; Bock, T.; Nolte, H.; Nogara, L.; Blaauw, B.; et al. High-throughput proteomics fiber typing (ProFiT) for comprehensive characterization of single skeletal muscle fibers. Skelet. Muscle. 2020, 10, 7. [Google Scholar] [CrossRef]
- Lang, F.; Aravamudhan, S.; Nolte, H.; Türk, C.; Hölper, S.; Müller, S.; Günther, S.; Blaauw, B.; Braun, T.; Krüger, M. Dynamic changes in the mouse skeletal muscle proteome during denervation-induced atrophy. Dis. Model Mech. 2017, 10, 881–896. [Google Scholar]
- Sadygov, R.G. Using Heavy Mass Isotopomers for Protein Turnover in Heavy Water Metabolic Labeling. J. Proteome Res. 2021, 20, 2035–2041. [Google Scholar] [CrossRef]
- Srisawat, K.; Hesketh, K.; Cocks, M.; Strauss, J.; Edwards, B.J.; Lisboa, P.J.; Shepherd, S.; Burniston, J.G. Reliability of Protein Abundance and Synthesis Measurements in Human Skeletal Muscle. Proteomics 2020, 20, e1900194. [Google Scholar] [CrossRef]
- Stansfield, B.N.; Brown, A.D.; Stewart, C.E.; Burniston, J.G. Dynamic Profiling of Protein Mole Synthesis Rates during C2C12 Myoblast Differentiation. Proteomics 2021, 21, e2000071. [Google Scholar] [CrossRef]
- Brown, A.D.; Stewart, C.E.; Burniston, J.G. Degradation of ribosomal and chaperone proteins is attenuated during the differentiation of replicatively aged C2C12 myoblasts. J. Cachexia Sarcopenia Muscle 2022, 13, 2562–2575. [Google Scholar] [CrossRef]
- Murphy, C.H.; Shankaran, M.; Churchward-Venne, T.A.; Mitchell, C.J.; Kolar, N.M.; Burke, L.M.; Hawley, J.A.; Kassis, A.; Karagounis, L.G.; Li, K.; et al. Effect of resistance training and protein intake pattern on myofibrillar protein synthesis and proteome kinetics in older men in energy restriction. J. Physiol. 2018, 596, 2091–2120. [Google Scholar] [CrossRef] [Green Version]
- Mann, M.; Kumar, C.; Zeng, W.F.; Strauss, M.T. Artificial intelligence for proteomics and biomarker discovery. Cell Syst. 2021, 12, 759–770. [Google Scholar] [CrossRef]
- Fenaille, F.; Barbier Saint-Hilaire, P.; Rousseau, K.; Junot, C. Data acquisition workflows in liquid chromatography coupled to high resolution mass spectrometry-based metabolomics: Where do we stand? J. Chromatogr. A 2017, 1526, 1–12. [Google Scholar] [CrossRef]
- Kitata, R.B.; Yang, J.C.; Chen, Y.J. Advances in data-independent acquisition mass spectrometry towards comprehensive digital proteome landscape. Mass Spectrom. Rev. 2022, e21781, in press. [Google Scholar] [CrossRef]
- Abdollahi, M.; Segura, P.A.; Beaudry, F. Is nontargeted data acquisition for target analysis (nDATA) in mass spectrometry a forward-thinking analytical approach? Biomed. Chromatogr. 2022, e5531, in press. [Google Scholar] [CrossRef]
- Krasny, L.; Huang, P.H. Data-independent acquisition mass spectrometry (DIA-MS) for proteomic applications in oncology. Mol. Omics 2021, 17, 29–42. [Google Scholar] [CrossRef]
- Kawashima, Y.; Watanabe, E.; Umeyama, T.; Nakajima, D.; Hattori, M.; Honda, K.; Ohara, O. Optimization of Data-Independent Acquisition Mass Spectrometry for Deep and Highly Sensitive Proteomic Analysis. Int. J. Mol. Sci. 2019, 20, 5932. [Google Scholar] [CrossRef] [Green Version]
- Gillet, L.C.; Navarro, P.; Tate, S.; Röst, H.; Selevsek, N.; Reiter, L.; Bonner, R.; Aebersold, R. Targeted data extraction of the MS/MS spectra generated by data-independent acquisition: A new concept for consistent and accurate proteome analysis. Mol. Cell. Proteom. 2012, 11, O111.016717. [Google Scholar] [CrossRef] [Green Version]
- Kontostathi, G.; Makridakis, M.; Bitsika, V.; Tsolakos, N.; Vlahou, A.; Zoidakis, J. Development and Validation of Multiple Reaction Monitoring (MRM) Assays for Clinical Applications. Methods Mol. Biol. 2019, 1959, 205–223. [Google Scholar]
- Cho, B.G.; Gutierrez Reyes, C.D.; Goli, M.; Gautam, S.; Banazadeh, A.; Mechref, Y. Targeted N-Glycan Analysis with Parallel Reaction Monitoring Using a Quadrupole-Orbitrap Hybrid Mass Spectrometer. Anal. Chem. 2022, 94, 15215–15222. [Google Scholar] [CrossRef]
- Ives, A.N.; Dunn, H.A.; Afsari, H.S.; Seckler, H.D.S.; Foroutan, M.J.; Chavez, E.; Melani, R.D.; Fellers, R.T.; LeDuc, R.D.; Thomas, P.M.; et al. Middle-Down Mass Spectrometry Reveals Activity-Modifying Phosphorylation Barcode in a Class C G Protein-Coupled Receptor. J. Am. Chem. Soc. 2022, 144, 23104–23114. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, E.; Biezuner, T.; Linnarsson, S. Single-cell sequencing-based technologies will revolutionize whole-organism science. Nat. Rev. Genet. 2013, 14, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, E.; Slavov, N. Single cell protein analysis for systems biology. Essays Biochem. 2018, 62, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Marx, V. A dream of single-cell proteomics. Nat. Methods 2019, 16, 809–812. [Google Scholar] [CrossRef] [Green Version]
- Budnik, B.; Levy, E.; Harmange, G.; Slavov, N. SCoPE-MS: Mass spectrometry of single mammalian cells quantifies proteome heterogeneity during cell differentiation. Genome Biol. 2018, 19, 161. [Google Scholar] [CrossRef] [Green Version]
- Specht, H.; Emmott, E.; Petelski, A.A.; Huffman, R.G.; Perlman, D.H.; Serra, M.; Kharchenko, P.; Koller, A.; Slavov, N. Single-cell proteomic and transcriptomic analysis of macrophage heterogeneity using SCoPE2. Genome Biol. 2021, 22, 50. [Google Scholar] [CrossRef]
- Brunner, A.D.; Thielert, M.; Vasilopoulou, C.; Ammar, C.; Coscia, F.; Mund, A.; Hoerning, O.B.; Bache, N.; Apalategui, A.; Lubeck, M.; et al. Ultra-high sensitivity mass spectrometry quantifies single-cell proteome changes upon perturbation. Mol. Syst. Biol. 2022, 18, e10798. [Google Scholar] [CrossRef]
- Petelski, A.A.; Emmott, E.; Leduc, A.; Huffman, R.G.; Specht, H.; Perlman, D.H.; Slavov, N. Multiplexed single-cell proteomics using SCoPE2. Nat. Protoc. 2021, 16, 5398–5425. [Google Scholar] [CrossRef]
- Cheung, T.K.; Lee, C.Y.; Bayer, F.P.; McCoy, A.; Kuster, B.; Rose, C.M. Defining the carrier proteome limit for single-cell proteomics. Nat. Methods 2021, 18, 76–83. [Google Scholar] [CrossRef]
- Huffman, R.G.; Chen, A.; Specht, H.; Slavov, N. DO-MS: Data-Driven Optimization of Mass Spectrometry Methods. J. Proteome Res. 2019, 18, 2493–2500. [Google Scholar] [CrossRef]
- Cong, Y.; Motamedchaboki, K.; Misal, S.A.; Liang, Y.; Guise, A.J.; Truong, T.; Huguet, R.; Plowey, E.D.; Zhu, Y.; Lopez-Ferrer, D.; et al. Ultrasensitive single-cell proteomics workflow identifies >1000 protein groups per mammalian cell. Chem. Sci. 2020, 12, 1001–1006. [Google Scholar] [CrossRef]
- Murgia, M.; Nogara, L.; Baraldo, M.; Reggiani, C.; Mann, M.; Schiaffino, S. Protein profile of fiber types in human skeletal muscle: A single-fiber proteomics study. Skelet. Muscle 2021, 11, 24. [Google Scholar] [CrossRef]
- Rai, M.F.; Wu, C.L.; Capellini, T.D.; Guilak, F.; Dicks, A.R.; Muthuirulan, P.; Grandi, F.; Bhutani, N.; Westendorf, J.J. Single Cell Omics for Musculoskeletal Research. Curr. Osteoporos. Rep. 2021, 19, 131–140. [Google Scholar] [CrossRef]
- Arias-Hidalgo, C.; Juanes-Velasco, P.; Landeira-Viñuela, A.; García-Vaquero, M.L.; Montalvillo, E.; Góngora, R.; Hernández, Á.P.; Fuentes, M. Single-Cell Proteomics: The Critical Role of Nanotechnology. Int. J. Mol. Sci. 2022, 23, 6707. [Google Scholar] [CrossRef]
- Huang, J.; Chen, X.; Fu, X.; Li, Z.; Huang, Y.; Liang, C. Advances in Aptamer-Based Biomarker Discovery. Front. Cell Dev. Biol. 2021, 9, 659760. [Google Scholar] [CrossRef]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Hathout, Y.; Liang, C.; Ogundele, M.; Xu, G.; Tawalbeh, S.M.; Dang, U.J.; Hoffman, E.P.; Gordish-Dressman, H.; Conklin, L.S.; van den Anker, J.N.; et al. Disease-specific and glucocorticoid-responsive serum biomarkers for Duchenne Muscular Dystrophy. Sci. Rep. 2019, 9, 12167. [Google Scholar] [CrossRef] [Green Version]
- Ogundele, M.; Zhang, J.S.; Goswami, M.V.; Barbieri, M.L.; Dang, U.J.; Novak, J.S.; Hoffman, E.P.; Nagaraju, K.; Cinrg-Dnhs Investigators; Hathout, Y. Validation of Chemokine Biomarkers in Duchenne Muscular Dystrophy. Life 2021, 11, 827. [Google Scholar] [CrossRef]
- Bottinelli, R.; Reggiani, C. Human skeletal muscle fibres: Molecular and functional diversity. Prog. Biophys. Mol. Biol. 2000, 73, 195–262. [Google Scholar] [CrossRef]
- Schiaffino, S. Fibre types in skeletal muscle: A personal account. Acta Physiol. 2010, 199, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Reggiani, C. Fiber types in mammalian skeletal muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciciliot, S.; Rossi, A.C.; Dyar, K.A.; Blaauw, B.; Schiaffino, S. Muscle type and fiber type specificity in muscle wasting. Int. J. Biochem. Cell Biol. 2013, 45, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Sawano, S.; Mizunoya, W. History and development of staining methods for skeletal muscle fiber types. Histol. Histopathol. 2022, 37, 493–503. [Google Scholar]
- Murach, K.A.; Dungan, C.M.; Kosmac, K.; Voigt, T.B.; Tourville, T.W.; Miller, M.S.; Bamman, M.M.; Peterson, C.A.; Toth, M.J. Fiber typing human skeletal muscle with fluorescent immunohistochemistry. J. Appl. Physiol. 2019, 127, 1632–1639. [Google Scholar] [CrossRef]
- Gonzalez-Freire, M.; Semba, R.D.; Ubaida-Mohien, C.; Fabbri, E.; Scalzo, P.; Højlund, K.; Dufresne, C.; Lyashkov, A.; Ferrucci, L. The human skeletal muscle proteome project: A reappraisal of the current literature. J. Cachexia Sarcopenia Muscle 2017, 8, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Dowling, P.; Gargan, S.; Swandulla, D.; Ohlendieck, K. Identification of Subproteomic Markers for Skeletal Muscle Profiling. Methods Mol. Biol. 2023, 2596, 291–302. [Google Scholar]
- Deshmukh, A.S.; Murgia, M.; Nagaraj, N.; Treebak, J.T.; Cox, J.; Mann, M. Deep proteomics of mouse skeletal muscle enables quantitation of protein isoforms, metabolic pathways, and transcription factors. Mol. Cell. Proteom. 2015, 14, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Højlund, K.; Yi, Z.; Hwang, H.; Bowen, B.; Lefort, N.; Flynn, C.R.; Langlais, P.; Weintraub, S.T.; Mandarino, L.J. (Characterization of the human skeletal muscle proteome by one-dimensional gel electrophoresis and HPLC-ESI-MS/MS. Mol. Cell. Proteom. 2008, 7, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Parker, K.C.; Walsh, R.J.; Salajegheh, M.; Amato, A.A.; Krastins, B.; Sarracino, D.A.; Greenberg, S.A. Characterization of human skeletal muscle biopsy samples using shotgun proteomics. J. Proteome Res. 2009, 8, 3265–3277. [Google Scholar] [CrossRef]
- Malik, Z.A.; Cobley, J.N.; Morton, J.P.; Close, G.L.; Edwards, B.J.; Koch, L.G.; Britton, S.L.; Burniston, J.G. Label-Free LC-MS Profiling of Skeletal Muscle Reveals Heart-Type Fatty Acid Binding Protein as a Candidate Biomarker of Aerobic Capacity. Proteomes 2013, 1, 290–308. [Google Scholar] [CrossRef] [Green Version]
- Burniston, J.G.; Connolly, J.; Kainulainen, H.; Britton, S.L.; Koch, L.G. Label-free profiling of skeletal muscle using high-definition mass spectrometry. Proteomics 2014, 14, 2339–2344. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wang, M.; Lin, S.; Jian, R.; Li, X.; Chan, J.; Robinson, A.E.; GTEx Consortium; Snyder, M.P. A quantitative proteome map of the human body. Cell 2020, 183, 269–283.e19. [Google Scholar] [CrossRef]
- Capitanio, D.; Viganò, A.; Ricci, E.; Cerretelli, P.; Wait, R.; Gelfi, C. Comparison of protein expression in human deltoideus and vastus lateralis muscles using two-dimensional gel electrophoresis. Proteomics 2005, 5, 2577–2586. [Google Scholar] [CrossRef]
- Hadrévi, J.; Hellström, F.; Kieselbach, T.; Malm, C.; Pedrosa-Domellöf, F. Protein differences between human trapezius and vastus lateralis muscles determined with a proteomic approach. BMC Musculoskelet. Disord. 2011, 12, 181. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, A.S.; Steenberg, D.E.; Hostrup, M.; Birk, J.B.; Larsen, J.K.; Santos, A.; Kjøbsted, R.; Hingst, J.R.; Schéele, C.C.; Murgia, M.; et al. Deep muscle-proteomic analysis of freeze-dried human muscle biopsies reveals fiber type-specific adaptations to exercise training. Nat. Commun. 2021, 12, 304. [Google Scholar] [CrossRef]
- Geiger, T.; Velic, A.; Macek, B.; Lundberg, E.; Kampf, C.; Nagaraj, N.; Uhlen, M.; Cox, J.; Mann, M. Initial quantitative proteomic map of 28 mouse tissues using the SILAC mouse. Mol. Cell. Proteom. 2013, 12, 1709–1722. [Google Scholar] [CrossRef] [Green Version]
- Raddatz, K.; Albrecht, D.; Hochgräfe, F.; Hecker, M.; Gotthardt, M. A proteome map of murine heart and skeletal muscle. Proteomics 2008, 8, 1885–1897. [Google Scholar] [CrossRef]
- Murphy, S.; Zweyer, M.; Raucamp, M.; Henry, M.; Meleady, P.; Swandulla, D.; Ohlendieck, K. Proteomic profiling of the mouse diaphragm and refined mass spectrometric analysis of the dystrophic phenotype. J. Muscle Res. Cell. Motil. 2019, 40, 9–28. [Google Scholar] [CrossRef]
- Murgia, M.; Nagaraj, N.; Deshmukh, A.S.; Zeiler, M.; Cancellara, P.; Moretti, I.; Reggiani, C.; Schiaffino, S.; Mann, M. Single muscle fiber proteomics reveals un-expected mitochondrial specialization. EMBO Rep. 2015, 16, 387–395. [Google Scholar] [CrossRef]
- Fomchenko, K.M.; Walsh, E.M.; Yang, X.; Verma, R.X.; Lin, B.L.; Nieuwenhuis, T.O.; Patil, A.H.; Fox-Talbot, K.; McCall, M.N.; Kass, D.A.; et al. Spatial proteomic approach to characterize skeletal muscle myofibers. J. Proteome Res. 2021, 20, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Okumura, N.; Hashida-Okumura, A.; Kita, K.; Matsubae, M.; Matsubara, T.; Takao, T.; Nagai, K. Proteomic analysis of slow- and fast-twitch skeletal muscles. Proteomics 2005, 5, 2896–2906. [Google Scholar] [CrossRef] [PubMed]
- Gelfi, C.; Viganò, A.; De Palma, S.; Ripamonti, M.; Begum, S.; Cerretelli, P.; Wait, R. 2-D protein maps of rat gastrocnemius and soleus muscles: A tool for muscle plasticity assessment. Proteomics 2006, 6, 321–340. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, R.; Ferreira, R.; Neuparth, M.; Guedes, S.; Williams, J.; Tomer, K.B.; Domingues, P.M.; Appell, H.J.; Duarte, J.A.; Amado, F.M. Subcellular proteomics of mice gastrocnemius and soleus muscles. Anal. Biochem. 2007, 366, 156–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drexler, H.C.; Ruhs, A.; Konzer, A.; Mendler, L.; Bruckskotten, M.; Looso, M.; Günther, S.; Boettger, T.; Krüger, M.; Braun, T. On marathons and Sprints: An integrated quantitative proteomics and transcriptomics analysis of differences between slow and fast muscle fibers. Mol. Cell. Proteom. 2012, 11, M111.010801. [Google Scholar] [CrossRef] [Green Version]
- Eggers, B.; Schork, K.; Turewicz, M.; Barkovits, K.; Eisenacher, M.; Schröder, R.; Clemen, C.S.; Marcus, K. Advanced fiber type- specific protein profiles derived from adult murine skeletal muscle. Proteomes 2021, 9, 28. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C.; Murgia, M. Fiber type diversity in skeletal muscle explored by mass spectrometry-based single fiber proteomics. Histol. Histopathol. 2020, 35, 239–246. [Google Scholar]
- Sweeney, H.L.; Hammers, D.W. Muscle Contraction. Cold Spring Harb. Perspect. Biol. 2018, 10, a023200. [Google Scholar] [CrossRef] [Green Version]
- Squire, J. Special Issue: The Actin-Myosin Interaction in Muscle: Background and Overview. Int. J. Mol. Sci. 2019, 20, 5715. [Google Scholar] [CrossRef] [Green Version]
- Powers, J.D.; Malingen, S.A.; Regnier, M.; Daniel, T.L. The Sliding Filament Theory Since Andrew Huxley: Multiscale and Multidisciplinary Muscle Research. Annu. Rev. Biophys. 2021, 50, 373–400. [Google Scholar] [CrossRef]
- Lin, B.L.; Song, T.; Sadayappan, S. Myofilaments: Movers and Rulers of the Sarcomere. Compr. Physiol. 2017, 7, 675–692. [Google Scholar] [PubMed]
- Ojima, K. Myosin: Formation and maintenance of thick filaments. Anim. Sci. J. 2019, 90, 801–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobacman, L.S. Troponin Revealed: Uncovering the Structure of the Thin Filament On-Off Switch in Striated Muscle. Biophys. J. 2021, 120, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Pinotsis, N.; Agarkova, I.; Ehler, E. The M-band: The underestimated part of the sarcomere. Biochim. Biophys. Acta Mol. Cell. Res. 2020, 1867, 118440. [Google Scholar] [CrossRef] [PubMed]
- Wadmore, K.; Azad, A.J.; Gehmlich, K. The Role of Z-disc Proteins in Myopathy and Cardiomyopathy. Int. J. Mol. Sci. 2021, 22, 3058. [Google Scholar] [CrossRef]
- Gordon, A.M.; Homsher, E.; Regnier, M. Regulation of contraction in striated muscle. Physiol. Rev. 2000, 80, 853–924. [Google Scholar] [CrossRef]
- Wang, L.; Geist, J.; Grogan, A.; Hu, L.R.; Kontrogianni-Konstantopoulos, A. Thick Filament Protein Network, Functions, and Disease Association. Compr. Physiol. 2018, 8, 631–709. [Google Scholar]
- Henderson, C.A.; Gomez, C.G.; Novak, S.M.; Mi-Mi, L.; Gregorio, C.C. Overview of the Muscle Cytoskeleton. Compr. Physiol. 2017, 7, 891–944. [Google Scholar]
- Reiser, P.J. Current understanding of conventional and novel co-expression patterns of mammalian sarcomeric myosin heavy chains and light chains. Arch. Biochem. Biophys. 2019, 662, 129–133. [Google Scholar] [CrossRef]
- Holland, A.; Ohlendieck, K. Proteomic profiling of the contractile apparatus from skeletal muscle. Expert Rev. Proteom. 2013, 10, 239–257. [Google Scholar] [CrossRef]
- Sitbon, Y.H.; Yadav, S.; Kazmierczak, K.; Szczesna-Cordary, D. Insights into myosin regulatory and essential light chains: A focus on their roles in cardiac and skeletal muscle function, development and disease. J. Muscle Res. Cell. Motil. 2020, 41, 313–327. [Google Scholar] [CrossRef]
- Pette, D.; Staron, R.S. Myosin isoforms, muscle fiber types, and transitions. Microsc. Res. Tech. 2000, 50, 500–509. [Google Scholar] [CrossRef]
- Schiaffino, S.; Rossi, A.C.; Smerdu, V.; Leinwand, L.A.; Reggiani, C. Developmental myosins: Expression patterns and functional significance. Skelet. Muscle 2015, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Hoh, J.F. Laryngeal muscle fibre types. Acta. Physiol. Scand. 2005, 183, 133–149. [Google Scholar] [CrossRef]
- Lee, L.A.; Karabina, A.; Broadwell, L.J.; Leinwand, L.A. The ancient sarcomeric myosins found in specialized muscles. Skelet. Muscle 2019, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Hoh, J.F.Y. Myosin heavy chains in extraocular muscle fibres: Distribution, regulation and function. Acta Physiol. 2021, 231, e13535. [Google Scholar] [CrossRef] [PubMed]
- Gargan, S.; Dowling, P.; Zweyer, M.; Reimann, J.; Henry, M.; Meleady, P.; Swandulla, D.; Ohlendieck, K. Mass Spectrometric Profiling of Extraocular Muscle and Proteomic Adaptations in the mdx-4cv Model of Duchenne Muscular Dystrophy. Life 2021, 11, 595. [Google Scholar] [CrossRef] [PubMed]
- Bozzo, C.; Spolaore, B.; Toniolo, L.; Stevens, L.; Bastide, B.; Cieniewski-Bernard, C.; Fontana, A.; Mounier, Y.; Reggiani, C. Nerve influence on myosin light chain phosphorylation in slow and fast skeletal muscles. FEBS J. 2005, 272, 5771–5785. [Google Scholar] [CrossRef] [PubMed]
- Robinett, J.C.; Hanft, L.M.; Geist, J.; Kontrogianni-Konstantopoulos, A.; McDonald, K.S. Regulation of myofilament force and loaded shortening by skeletal myosin binding protein C. J. Gen. Physiol. 2019, 151, 645–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luther, P.K.; Vydyanath, A. Myosin binding protein-C: An essential protein in skeletal and cardiac muscle. J. Muscle Res. Cell. Motil. 2011, 31, 303–305. [Google Scholar] [CrossRef]
- Lin, B.L.; Li, A.; Mun, J.Y.; Previs, M.J.; Previs, S.B.; Campbell, S.G.; Dos Remedios, C.G.; Tombe, P.P.; Craig, R.; Warshaw, D.M.; et al. Skeletal myosin binding protein-C isoforms regulate thin filament activity in a Ca2+-dependent manner. Sci. Rep. 2018, 8, 2604. [Google Scholar] [CrossRef] [Green Version]
- McNamara, J.W.; Sadayappan, S. Skeletal myosin binding protein-C: An increasingly important regulator of striated muscle physiology. Arch. Biochem. Biophys. 2018, 660, 121–128. [Google Scholar] [CrossRef]
- Prill, K.; Dawson, J.F. Assembly and Maintenance of Sarcomere Thin Filaments and Associated Diseases. Int. J. Mol. Sci. 2020, 21, 542. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, R.; Holmes, K.C. Actin structure and function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef] [Green Version]
- Nowak, K.J.; Ravenscroft, G.; Laing, N.G. Skeletal muscle α-actin diseases (actinopathies): Pathology and mechanisms. Acta Neuropathol. 2013, 125, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.R.; Campbell, S.G.; Lehman, W. Structural determinants of muscle thin filament cooperativity. Arch. Biochem. Biophys. 2016, 594, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Guhathakurta, P.; Prochniewicz, E.; Thomas, D.D. Actin-Myosin Interaction: Structure, Function and Drug Discovery. Int. J. Mol. Sci. 2018, 19, 2628. [Google Scholar] [CrossRef] [Green Version]
- El-Mezgueldi, M. Tropomyosin dynamics. J. Muscle Res. Cell. Motil. 2014, 35, 203–210. [Google Scholar] [CrossRef]
- Hitchcock-DeGregori, S.E.; Barua, B. Tropomyosin Structure, Function, and Interactions: A Dynamic Regulator. Subcell. Biochem. 2017, 82, 253–284. [Google Scholar]
- Lehman, W.; Rynkiewicz, M.J.; Moore, J.R. A new twist on tropomyosin binding to actin filaments: Perspectives on thin filament function, assembly and biomechanics. J. Muscle Res. Cell. Motil. 2020, 41, 23–38. [Google Scholar] [CrossRef]
- Gomes, A.V.; Potter, J.D.; Szczesna-Cordary, D. The role of troponins in muscle contraction. IUBMB Life 2002, 54, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Swartz, D.R.; Yang, Z.; Sen, A.; Tikunova, S.B.; Davis, J.P. Myofibrillar troponin exists in three states and there is signal transduction along skeletal myofibrillar thin filaments. J. Mol. Biol. 2006, 361, 420–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, M.; Jin, J.P. Troponin Variants as Markers of Skeletal Muscle Health and Diseases. Front. Physiol. 2021, 12, 747214. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Feng, X.; Dong, J.; Wang, Z.M.; Lee, J.; Furdui, C.; Files, D.C.; Beavers, K.M.; Kritchevsky, S.; Milligan, C.; et al. Cardiac troponin T and fast skeletal muscle denervation in ageing. J. Cachexia Sarcopenia Muscle 2017, 8, 808–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, D.; Kuhn, C.; Katus, H.A.; Frey, N. The sarcomeric Z-disc: A nodal point in signalling and disease. J. Mol. Med. 2006, 84, 446–468. [Google Scholar] [CrossRef]
- Luther, P.K. The vertebrate muscle Z-disc: Sarcomere anchor for structure and signalling. J. Muscle Res. Cell. Motil. 2009, 30, 171–185. [Google Scholar] [CrossRef] [Green Version]
- Ranta-Aho, J.; Olive, M.; Vandroux, M.; Roticiani, G.; Dominguez, C.; Johari, M.; Torella, A.; Böhm, J.; Turon, J.; Nigro, V.; et al. Mutation update for the ACTN2 gene. Hum. Mutat. 2022, 43, 1745–1756. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Makizako, H.; Nakai, Y.; Kiuchi, Y.; Akaida, S.; Tateishi, M.; Takenaka, T.; Kubozono, T.; Ohishi, M. Associations of the Alpha-Actinin Three Genotype with Bone and Muscle Mass Loss among Middle-Aged and Older Adults. J. Clin. Med. 2022, 11, 6172. [Google Scholar] [CrossRef]
- Gontier, Y.; Taivainen, A.; Fontao, L.; Sonnenberg, A.; van der Flier, A.; Carpen, O.; Faulkner, G.; Borradori, L. The Z-disc proteins myotilin and FATZ-1 interact with each other and are connected to the sarcolemma via muscle-specific filamins. J. Cell Sci. 2005, 118, 3739–3749. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.D.; Romero, M.A.; Mobley, C.B.; Mumford, P.W.; Roberson, P.A.; Haun, C.T.; Vann, C.G.; Osburn, S.C.; Holmes, H.H.; Greer, R.A.; et al. Skeletal muscle mitochondrial volume and myozenin-1 protein differences exist between high versus low anabolic responders to resistance training. PeerJ 2018, 6, e5338. [Google Scholar] [CrossRef] [Green Version]
- Agarkova, I.; Perriard, J.C. The M-band: An elastic web that crosslinks thick filaments in the center of the sarcomere. Trends Cell Biol. 2005, 15, 477–485. [Google Scholar] [CrossRef]
- Van Der Ven, P.F.; Obermann, W.M.; Weber, K.; Fürst, D.O. Myomesin, M-protein and the structure of the sarcomeric M-band. Adv. Biophys. 1996, 33, 91–99. [Google Scholar] [CrossRef]
- Prill, K.; Carlisle, C.; Stannard, M.; Windsor Reid, P.J.; Pilgrim, D.B. Myomesin is part of an integrity pathway that responds to sarcomere damage and disease. PLoS One 2019, 14, e0224206. [Google Scholar] [CrossRef] [Green Version]
- Manring, H.R.; Carter, O.A.; Ackermann, M.A. Obscure functions: The location-function relationship of obscurins. Biophys. Rev. 2017, 9, 245–258. [Google Scholar] [CrossRef] [Green Version]
- Randazzo, D.; Pierantozzi, E.; Rossi, D.; Sorrentino, V. The potential of obscurin as a therapeutic target in muscle disorders. Expert Opin. Ther. Targets 2017, 21, 897–910. [Google Scholar] [CrossRef]
- Chu, M.; Gregorio, C.C.; Pappas, C.T. Nebulin, a multi-functional giant. J. Exp. Biol. 2016, 219, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Yuen, M.; Ottenheijm, C.A.C. Nebulin: Big protein with big responsibilities. J. Muscle Res. Cell. Motil. 2020, 41, 103–124. [Google Scholar] [CrossRef] [Green Version]
- Freundt, J.K.; Linke, W.A. Titin as a force-generating muscle protein under regulatory control. J. Appl. Physiol. 2019, 126, 1474–1482. [Google Scholar] [CrossRef]
- Adewale, A.O.; Ahn, Y.H. Titin N2A Domain and Its Interactions at the Sarcomere. Int. J. Mol. Sci. 2021, 22, 7563. [Google Scholar] [CrossRef]
- Nishikawa, K.; Lindstedt, S.L.; Hessel, A.; Mishra, D. N2A Titin: Signaling Hub and Mechanical Switch in Skeletal Muscle. Int. J. Mol. Sci. 2020, 21, 3974. [Google Scholar] [CrossRef]
- Hessel, A.L.; Ma, W.; Mazara, N.; Rice, P.E.; Nissen, D.; Gong, H.; Kuehn, M.; Irving, T.; Linke, W.A. Titin force in muscle cells alters lattice order, thick and thin filament protein formation. Proc. Natl. Acad. Sci. USA 2022, 119, e2209441119. [Google Scholar] [CrossRef] [PubMed]
- Wette, S.G.; Smith, H.K.; Lamb, G.D.; Murphy, R.M. Characterization of muscle ankyrin repeat proteins in human skeletal muscle. Am. J. Physiol. Cell Physiol. 2017, 313, C327–C339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargan, S.; Dowling, P.; Zweyer, M.; Henry, M.; Meleady, P.; Swandulla, D.; Ohlendieck, K. Proteomic Identification of Markers of Membrane Repair, Regeneration and Fibrosis in the Aged and Dystrophic Diaphragm. Life 2022, 12, 1679. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic, N.; Lim, J.Y.; Miljkovic, I.; Frontera, W.R. Aging of skeletal muscle fibers. Ann. Rehabil. Med. 2015, 39, 155–162. [Google Scholar] [CrossRef]
- Grosicki, G.J.; Zepeda, C.S.; Sundberg, C.W. Single muscle fibre contractile function with ageing. J. Physiol. 2022, 600, 5005–5026. [Google Scholar] [CrossRef]
- Wilkinson, D.J.; Piasecki, M.; Atherton, P.J. The age-related loss of skeletal muscle mass and function: Measurement and physiology of muscle fibre atrophy and muscle fibre loss in humans. Ageing Res. Rev. 2018, 47, 123–132. [Google Scholar] [CrossRef]
- Loh, R.; Tan, R.S.; Lim, W.S.; Koh, A.S. Cardio-sarcopenia: A syndrome of concern in aging. Front. Med. 2022, 9, 1027466. [Google Scholar] [CrossRef]
- Aare, S.; Spendiff, S.; Vuda, M.; Elkrief, D.; Perez, A.; Wu, Q.; Mayaki, D.; Hussain, S.N.; Hettwer, S.; Hepple, R.T. Failed reinnervation in aging skeletal muscle. Skelet. Muscle 2016, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.L. Muscle fibre type adaptation in the elderly human muscle. Scan. J. Med. Sci. Sports 2003, 13, 40–47. [Google Scholar] [CrossRef]
- Wang, Y.; Pessin, J.E. Mechanisms for fiber-type specificity of skeletal muscle atrophy. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 243–250. [Google Scholar] [CrossRef]
- Frontera, W.R.; Hughes, V.A.; Fielding, R.A.; Fiatarone, M.A.; Evans, W.J.; Roubenoff, R. Aging of skeletal muscle: A 12-yr longitudinal study. J. Appl. Physiol. 2000, 88, 1321–1326. [Google Scholar] [CrossRef]
- Frontera, W.R.; Reid, K.F.; Phillips, E.M.; Krivickas, L.S.; Hughes, V.A.; Roubenoff, R.; Fielding, R.A. Muscle fiber size and function in elderly humans: A longitudinal study. J. Appl. Physiol. 2008, 105, 637–642. [Google Scholar] [CrossRef] [Green Version]
- Carvalho do Nascimento, P.R.; Bilodeau, M.; Poitras, S. How do we define and measure sarcopenia? A meta-analysis of observational studies. Age Ageing 2021, 50, 1906–1913. [Google Scholar] [CrossRef]
- Spexoto, M.C.B.; Ramírez, P.C.; de Oliveira Máximo, R.; Steptoe, A.; de Oliveira, C.; Alexandre, T.D.S. European Working Group on Sarcopenia in Older People 2010 (EWGSOP1) and 2019 (EWGSOP2) criteria or slowness: Which is the best predictor of mortality risk in older adults? Age Ageing 2022, 51, afac164. [Google Scholar] [CrossRef]
- Petermann-Rocha, F.; Balntzi, V.; Gray, S.R.; Lara, J.; Ho, F.K.; Pell, J.P.; Celis-Morales, C. Global prevalence of sarcopenia and severe sarcopenia: A systematic review and meta-analysis. J. Cachexia Sarcopenia Muscle. 2022, 13, 86–99. [Google Scholar] [CrossRef]
- Davies, B.; Walter, S.; Rodríguez-Laso, A.; Carnicero Carreño, J.A.; García-García, F.J.; Álvarez-Bustos, A.; Rodríguez-Mañas, L. Differential Association of Frailty and Sarcopenia With Mortality and Disability: Insight Supporting Clinical Subtypes of Frailty. J. Am. Med. Dir. Assoc. 2022, 23, 1712–1716.e3. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, T.; Zhang, Y.; She, Y.; Wang, L.; Gao, Y.; Deng, Y.; Chen, M.; He, Y.; Chen, X.; et al. Natural population cohort study on long-lived adults: West China longevity and ageing procedure (WCLAP). BMJ Open 2022, 12, e055407. [Google Scholar] [CrossRef]
- Mitchell, W.K.; Williams, J.; Atherton, P.; Larvin, M.; Lund, J.; Narici, M. Sarcopenia, Dynapenia, and the impact of advancing age on human skeletal muscle size and strength: A quantitative review. Front. Physiol. 2012, 3, 260. [Google Scholar] [CrossRef] [Green Version]
- Chiba, I.; Lee, S.; Bae, S.; Makino, K.; Shinkai, Y.; Katayama, O.; Harada, K.; Takayanagi, N.; Shimada, H. Difference in sarcopenia characteristics associated with physical activity and disability incidences in older adults. J. Cachexia Sarcopenia Muscle 2021, 12, 1983–1994. [Google Scholar] [CrossRef]
- Frontera, W.R. Rehabilitation of Older Adults with Sarcopenia: From Cell to Functioning. Prog. Rehabil. Med. 2022, 7, 20220044. [Google Scholar] [CrossRef]
- Mcleod, J.C.; Stokes, T.; Phillips, S.M. Resistance Exercise Training as a Primary Countermeasure to Age-Related Chronic Disease. Front. Physiol. 2019, 10, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho-Júnior, H.J.; Picca, A.; Calvani, R.; Marzetti, E. Prescription of resistance training for sarcopenic older adults: Does it require specific attention? Ageing Res. Rev. 2022, 81, 101720. [Google Scholar] [CrossRef] [PubMed]
- de Sá Souza, H.; de Melo, C.M.; Piovezan, R.D.; Miranda, R.E.E.P.C.; Carneiro-Junior, M.A.; Silva, B.M.; Thomatieli-Santos, R.V.; Tufik, S.; Poyares, D.; D’Almeida, V. Resistance Training Improves Sleep and Anti-Inflammatory Parameters in Sarcopenic Older Adults: A Randomized Controlled Trial. Int. J. Environ. Res. Public Health 2022, 19, 16322. [Google Scholar] [CrossRef] [PubMed]
- Cobley, J.N.; Moult, P.R.; Burniston, J.G.; Morton, J.P.; Close, G.L. Exercise improves mitochondrial and redox-regulated stress responses in the elderly: Better late than never! Biogerontology 2015, 16, 249–264. [Google Scholar] [CrossRef] [PubMed]
- El Assar, M.; Álvarez-Bustos, A.; Sosa, P.; Angulo, J.; Rodríguez-Mañas, L. Effect of Physical Activity/Exercise on Oxidative Stress and Inflammation in Muscle and Vascular Aging. Int. J. Mol. Sci. 2022, 23, 8713. [Google Scholar] [CrossRef]
- Hirsch, K.R.; Church, D.D.; Kim, I.Y.; Park, S.; Wolfe, R.R.; Ferrando, A.A. Comparison of basal whole-body protein kinetics and muscle protein synthesis between young and older adults. Physiol. Rep. 2020, 8, e14633. [Google Scholar] [CrossRef]
- Sieber, C.C. Malnutrition and sarcopenia. Aging Clin. Exp. Res. 2019, 31, 793–798. [Google Scholar] [CrossRef]
- Jang, E.H.; Han, Y.J.; Jang, S.E.; Lee, S. Association between Diet Quality and Sarcopenia in Older Adults: Systematic Review of Prospective Cohort Studies. Life 2021, 11, 811. [Google Scholar] [CrossRef]
- Coelho-Junior, H.J.; Calvani, R.; Azzolino, D.; Picca, A.; Tosato, M.; Landi, F.; Cesari, M.; Marzetti, E. Protein Intake and Sarcopenia in Older Adults: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2022, 19, 8718. [Google Scholar] [CrossRef]
- Bradlee, M.L.; Mustafa, J.; Singer, M.R.; Moore, L.L. High-Protein Foods and Physical Activity Protect Against Age-Related Muscle Loss and Functional Decline. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 73, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Coelho-Junior, H.J.; Marzetti, E.; Picca, A.; Cesari, M.; Uchida, M.C.; Calvani, R. Protein Intake and Frailty: A Matter of Quantity, Quality, and Timing. Nutrients 2020, 12, 2915. [Google Scholar] [CrossRef]
- Kinoshita, K.; Otsuka, R.; Nishita, Y.; Tange, C.; Tomida, M.; Zhang, S.; Ando, F.; Shimokata, H.; Arai, H. Breakfast Protein Quality and Muscle Strength in Japanese Older Adults: A Community-Based Longitudinal Study. J. Am. Med. Dir. Assoc. 2022, 23, 729–735.e2. [Google Scholar] [CrossRef]
- Coelho-Júnior, H.J.; Calvani, R.; Tosato, M.; Landi, F.; Picca, A.; Marzetti, E. Protein intake and physical function in older adults: A systematic review and meta-analysis. Ageing Res. Rev. 2022, 81, 101731. [Google Scholar] [CrossRef]
- Rezuş, E.; Burlui, A.; Cardoneanu, A.; Rezuş, C.; Codreanu, C.; Pârvu, M.; Rusu Zota, G.; Tamba, B.I. Inactivity and Skeletal Muscle Metabolism: A Vicious Cycle in Old Age. Int. J. Mol. Sci. 2020, 21, 592. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.M.; Johannsen, D.L.; Ravussin, E. Skeletal muscle mitochondria and aging: A review. J. Aging Res. 2012, 2012, 194821. [Google Scholar] [CrossRef] [Green Version]
- Picca, A.; Lezza, A.M.S.; Leeuwenburgh, C.; Pesce, V.; Calvani, R.; Landi, F.; Bernabei, R.; Marzetti, E. Fueling Inflamm-Aging through Mitochondrial Dysfunction: Mechanisms and Molecular Targets. Int. J. Mol. Sci. 2017, 18, 933. [Google Scholar] [CrossRef] [Green Version]
- Casuso, R.A.; Huertas, J.R. The emerging role of skeletal muscle mitochondrial dynamics in exercise and ageing. Ageing Res. Rev. 2020, 58, 101025. [Google Scholar] [CrossRef]
- Ferri, E.; Marzetti, E.; Calvani, R.; Picca, A.; Cesari, M.; Arosio, B. Role of Age-Related Mitochondrial Dysfunction in Sarcopenia. Int. J. Mol. Sci. 2020, 21, 5236. [Google Scholar] [CrossRef]
- Romanello, V. The Interplay between Mitochondrial Morphology and Myomitokines in Aging Sarcopenia. Int. J. Mol. Sci. 2020, 22, 91. [Google Scholar] [CrossRef]
- Leduc-Gaudet, J.P.; Hussain, S.N.A.; Barreiro, E.; Gouspillou, G. Mitochondrial Dynamics and Mitophagy in Skeletal Muscle Health and Aging. Int. J. Mol. Sci. 2021, 22, 8179. [Google Scholar] [CrossRef]
- Kimoloi, S.; Sen, A.; Guenther, S.; Braun, T.; Brügmann, T.; Sasse, P.; Wiesner, R.J.; Pla-Martín, D.; Baris, O.R. Combined fibre atrophy and decreased muscle regeneration capacity driven by mitochondrial DNA alterations underlie the development of sarcopenia. J. Cachexia Sarcopenia Muscle 2022, 13, 2132–2145. [Google Scholar] [CrossRef] [PubMed]
- Sakellariou, G.K.; McDonagh, B. Redox Homeostasis in Age-Related Muscle Atrophy. Adv. Exp. Med. Biol. 2018, 1088, 281–306. [Google Scholar] [PubMed]
- Cobley, J.N.; Sakellariou, G.K.; Husi, H.; McDonagh, B. Proteomic strategies to unravel age-related redox signalling defects in skeletal muscle. Free Radic. Biol. Med. 2019, 132, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Shally, A.; McDonagh, B. The redox environment and mitochondrial dysfunction in age-related skeletal muscle atrophy. Biogerontology 2020, 21, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Foreman, N.A.; Hesse, A.S.; Ji, L.L. Redox Signaling and Sarcopenia: Searching for the Primary Suspect. Int. J. Mol. Sci. 2021, 22, 9045. [Google Scholar] [CrossRef]
- Protasi, F.; Pietrangelo, L.; Boncompagni, S. Improper Remodeling of Organelles Deputed to Ca2+ Handling and Aerobic ATP Production Underlies Muscle Dysfunction in Ageing. Int. J. Mol. Sci. 2021, 22, 6195. [Google Scholar] [CrossRef]
- Iyer, S.R.; Shah, S.B.; Lovering, R.M. The Neuromuscular Junction: Roles in Aging and Neuromuscular Disease. Int. J. Mol. Sci. 2021, 22, 8058. [Google Scholar] [CrossRef]
- Mancinelli, R.; Checcaglini, F.; Coscia, F.; Gigliotti, P.; Fulle, S.; Fanò-Illic, G. Biological Aspects of Selected Myokines in Skeletal Muscle: Focus on Aging. Int. J. Mol. Sci. 2021, 22, 8520. [Google Scholar] [CrossRef]
- Cannataro, R.; Carbone, L.; Petro, J.L.; Cione, E.; Vargas, S.; Angulo, H.; Forero, D.A.; Odriozola-Martínez, A.; Kreider, R.B.; Bonilla, D.A. Sarcopenia: Etiology, Nutritional Approaches, and miRNAs. Int. J. Mol. Sci. 2021, 22, 9724. [Google Scholar] [CrossRef]
- Francisco, S.; Martinho, V.; Ferreira, M.; Reis, A.; Moura, G.; Soares, A.R.; Santos, M.A.S. The Role of MicroRNAs in Proteostasis Decline and Protein Aggregation during Brain and Skeletal Muscle Aging. Int. J. Mol. Sci. 2022, 23, 3232. [Google Scholar] [CrossRef]
- Prokopidis, K.; Chambers, E.; Ni Lochlainn, M.; Witard, O.C. Mechanisms Linking the Gut-Muscle Axis With Muscle Protein Metabolism and Anabolic Resistance: Implications for Older Adults at Risk of Sarcopenia. Front. Physiol. 2021, 12, 770455. [Google Scholar] [CrossRef]
- Tan, K.T.; Ang, S.J.; Tsai, S.Y. Sarcopenia: Tilting the Balance of Protein Homeostasis. Proteomics 2020, 20, e1800411. [Google Scholar] [CrossRef]
- Fernando, R.; Drescher, C.; Nowotny, K.; Grune, T.; Castro, J.P. Impaired proteostasis during skeletal muscle aging. Free Radic. Biol. Med. 2019, 132, 58–66. [Google Scholar] [CrossRef]
- Wilhelmsen, A.; Tsintzas, K.; Jones, S.W. Recent advances and future avenues in understanding the role of adipose tissue cross talk in mediating skeletal muscle mass and function with ageing. Geroscience 2021, 43, 85–110. [Google Scholar] [CrossRef]
- Campos, G.C.; Lourenço, R.A.; Molina, M.D.C.B. Mortality, sarcopenic obesity, and sarcopenia: Frailty in Brazilian Older People Study-FIBRA-RJ. Rev. Saude Publica 2021, 55, 75. [Google Scholar] [CrossRef]
- Zhang, X.; Li, H.; He, M.; Wang, J.; Wu, Y.; Li, Y. Immune system and sarcopenia: Presented relationship and future perspective. Exp. Gerontol. 2022, 164, 111823. [Google Scholar] [CrossRef]
- Nelke, C.; Dziewas, R.; Minnerup, J.; Meuth, S.G.; Ruck, T. Skeletal muscle as potential central link between sarcopenia and immune senescence. EBioMedicine. 2019, 49, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Gutierrez, G.E.; Martínez-Gómez, L.E.; Martínez-Armenta, C.; Pineda, C.; Martínez-Nava, G.A.; Lopez-Reyes, A. Molecular Mechanisms of Inflammation in Sarcopenia: Diagnosis and Therapeutic Update. Cells 2022, 11, 2359. [Google Scholar] [CrossRef]
- Kadi, F.; Ponsot, E. The biology of satellite cells and telomeres in human skeletal muscle: Effects of aging and physical activity. Scand. J. Med. Sci. Sports 2010, 20, 39–48. [Google Scholar] [CrossRef]
- Sui, S.X.; Williams, L.J.; Holloway-Kew, K.L.; Hyde, N.K.; Pasco, J.A. Skeletal Muscle Health and Cognitive Function: A Narrative Review. Int. J. Mol. Sci. 2020, 22, 255. [Google Scholar] [CrossRef]
- Papadopoulou, S.K.; Voulgaridou, G.; Kondyli, F.S.; Drakaki, M.; Sianidou, K.; Andrianopoulou, R.; Rodopaios, N.; Pritsa, A. Nutritional and Nutrition-Related Biomarkers as Prognostic Factors of Sarcopenia, and Their Role in Disease Progression. Diseases 2022, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Verdijk, L.B.; Koopman, R.; Schaart, G.; Meijer, K.; Savelberg, H.H.; van Loon, L.J. Satellite cell content is specifically reduced in type II skeletal muscle fibers in the elderly. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E151–E157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, V.; Thornell, L.E.; Eriksson, P.O.; Butler-Browne, G.; Mouly, V. Regenerative potential of human skeletal muscle during aging. Aging Cell 2002, 1, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Joanisse, S.; Nederveen, J.P.; Snijders, T.; McKay, B.R.; Parise, G. Skeletal Muscle Regeneration, Repair and Remodelling in Aging: The Importance of Muscle Stem Cells and Vascularization. Gerontology 2017, 63, 91–100. [Google Scholar] [CrossRef]
- Huo, F.; Liu, Q.; Liu, H. Contribution of muscle satellite cells to sarcopenia. Front. Physiol. 2022, 13, 892749. [Google Scholar] [CrossRef]
- Fernández-Lázaro, D.; Garrosa, E.; Seco-Calvo, J.; Garrosa, M. Potential Satellite Cell-Linked Biomarkers in Aging Skeletal Muscle Tissue: Proteomics and Proteogenomics to Monitor Sarcopenia. Proteomes 2022, 10, 29. [Google Scholar] [CrossRef]
- Liu, J.C.; Dong, S.S.; Shen, H.; Yang, D.Y.; Chen, B.B.; Ma, X.Y.; Peng, Y.R.; Xiao, H.M.; Deng, H.W. Multi-omics research in sarcopenia: Current progress and future prospects. Ageing Res. Rev. 2022, 76, 101576. [Google Scholar] [CrossRef]
- Curcio, F.; Ferro, G.; Basile, C.; Liguori, I.; Parrella, P.; Pirozzi, F.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Tocchetti, C.G.; et al. Biomarkers in sarcopenia: A multifactorial approach. Exp. Gerontol. 2016, 85, 1–8. [Google Scholar] [CrossRef]
- Picca, A.; Calvani, R.; Sirago, G.; Coelho-Junior, H.J.; Marzetti, E. Molecular routes to sarcopenia and biomarker development: Per aspera ad astra. Curr. Opin. Pharmacol. 2021, 57, 140–147. [Google Scholar] [CrossRef]
- Picca, A.; Calvani, R.; Marzetti, E. Multisystem derangements in frailty and sarcopenia: A source for biomarker discovery. Curr. Opin. Clin. Nutr. Metab. Care 2022, 25, 173–177. [Google Scholar] [CrossRef]
- Gelfi, C.; Vigano, A.; Ripamonti, M.; Pontoglio, A.; Begum, S.; Pellegrino, M.A.; Grassi, B.; Bottinelli, R.; Wait, R.; Cerretelli, P. The human muscle proteome in aging. J. Proteome Res. 2006, 5, 1344–1353. [Google Scholar] [CrossRef]
- Staunton, L.; Zweyer, M.; Swandulla, D.; Ohlendieck, K. Mass spectrometry-based proteomic analysis of middle-aged vs. aged vastus lateralis reveals increased levels of carbonic anhydrase isoform 3 in senescent human skeletal muscle. Int. J. Mol. Med. 2012, 30, 723–733. [Google Scholar] [CrossRef] [Green Version]
- Ohlendieck, K. Two-CyDye-Based 2D-DIGE Analysis of Aged Human Muscle Biopsy Specimens. Methods Mol. Biol. 2023, 2596, 265–289. [Google Scholar]
- Gueugneau, M.; Coudy-Gandilhon, C.; Gourbeyre, O.; Chambon, C.; Combaret, L.; Polge, C.; Taillandier, D.; Attaix, D.; Friguet, B.; Maier, A.B.; et al. Proteomics of muscle chronological ageing in post-menopausal women. BMC Genom. 2014, 15, 1165. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, S.L.; Baraibar, M.A.; Lundberg, S.; Eeg-Olofsson, O.; Larsson, L.; Friguet, B. Oxidative proteome alterations during skeletal muscle ageing. Redox Biol. 2015, 5, 267–274. [Google Scholar] [CrossRef]
- Cobley, J.N.; Ab Malik, Z.; Morton, J.P.; Close, G.L.; Edwards, B.J.; Burniston, J.G. Age- and Activity-Related Differences in the Abundance of Myosin Essential and Regulatory Light Chains in Human Muscle. Proteomes 2016, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Murgia, M.; Toniolo, L.; Nagaraj, N.; Ciciliot, S.; Vindigni, V.; Schiaffino, S.; Reggiani, C.; Mann, M. Single Muscle Fiber Proteomics Reveals Fiber-Type-Specific Features of Human Muscle Aging. Cell Rep. 2017, 19, 2396–2409. [Google Scholar] [CrossRef] [Green Version]
- Ebhardt, H.A.; Degen, S.; Tadini, V.; Schilb, A.; Johns, N.; Greig, C.A.; Fearon, K.C.H.; Aebersold, R.; Jacobi, C. Comprehensive proteome analysis of human skeletal muscle in cachexia and sarcopenia: A pilot study. J. Cachexia Sarcopenia Muscle 2017, 8, 567–582. [Google Scholar] [CrossRef] [Green Version]
- Brocca, L.; McPhee, J.S.; Longa, E.; Canepari, M.; Seynnes, O.; De Vito, G.; Pellegrino, M.A.; Narici, M.; Bottinelli, R. Structure and function of human muscle fibres and muscle proteome in physically active older men. J. Physiol. 2017, 595, 4823–4844. [Google Scholar] [CrossRef] [Green Version]
- Ubaida-Mohien, C.; Lyashkov, A.; Gonzalez-Freire, M.; Tharakan, R.; Shardell, M.; Moaddel, R.; Semba, R.D.; Chia, C.W.; Gorospe, M.; Sen, R.; et al. Discovery proteomics in aging human skeletal muscle finds change in spliceosome, immunity, proteostasis and mitochondria. Elife 2019, 8, e49874. [Google Scholar] [CrossRef]
- Ubaida-Mohien, C.; Gonzalez-Freire, M.; Lyashkov, A.; Moaddel, R.; Chia, C.W.; Simonsick, E.M.; Sen, R.; Ferrucci, L. Physical Activity Associated Proteomics of Skeletal Muscle: Being Physically Active in Daily Life May Protect Skeletal Muscle From Aging. Front. Physiol. 2019, 10, 312. [Google Scholar] [CrossRef] [PubMed]
- Vann, C.G.; Roberson, P.A.; Osburn, S.C.; Mumford, P.W.; Romero, M.A.; Fox, C.D.; Moore, J.H.; Haun, C.T.; Beck, D.T.; Moon, J.R.; et al. Skeletal Muscle Myofibrillar Protein Abundance Is Higher in Resistance-Trained Men, and Aging in the Absence of Training May Have an Opposite Effect. Sports 2020, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gueugneau, M.; Coudy-Gandilhon, C.; Chambon, C.; Verney, J.; Taillandier, D.; Combaret, L.; Polge, C.; Walrand, S.; Roche, F.; Barthélémy, J.C.; et al. Muscle Proteomic and Transcriptomic Profiling of Healthy Aging and Metabolic Syndrome in Men. Int. J. Mol. Sci. 2021, 22, 4205. [Google Scholar] [CrossRef] [PubMed]
- Deane, C.S.; Phillips, B.E.; Willis, C.R.G.; Wilkinson, D.J.; Smith, K.; Higashitani, N.; Williams, J.P.; Szewczyk, N.J.; Atherton, P.J.; Higashitani, A.; et al. Proteomic features of skeletal muscle adaptation to resistance exercise training as a function of age. Geroscience. 2022, in press. [Google Scholar] [CrossRef]
- Balagopalm, P.; Rooyackers, O.E.; Adey, D.B.; Ades, P.A.; Nair, K.S. Effects of aging on in vivo synthesis of skeletal muscle myosin heavy-chain and sarcoplasmic protein in humans. Am. J. Physiol. 1997, 273, E790–E800. [Google Scholar]
- Lexell, J.; Taylor, C.C.; Sjöström, M. What is the cause of the ageing atrophy? Total number, size and proportion of different fiber types studied in whole vastus lateralis muscle from 15- to 83-year-old men. J. Neurol. Sci. 1988, 84, 275–294. [Google Scholar] [CrossRef]
- Staunton, L.; O'Connell, K.; Ohlendieck, K. Proteomic Profiling of Mitochondrial Enzymes during Skeletal Muscle Aging. J. Aging Res. 2011, 2011, 908035. [Google Scholar] [CrossRef] [Green Version]
- Egan, B.; Dowling, P.; O’Connor, P.L.; Henry, M.; Meleady, P.; Zierath, J.R.; O’Gorman, D.J. 2-D DIGE analysis of the mitochondrial proteome from human skeletal muscle reveals time course-dependent remodelling in response to 14 consecutive days of endurance exercise training. Proteomics 2011, 11, 1413–1428. [Google Scholar] [CrossRef]
- Burniston, J.G.; Hoffman, E.P. Proteomic responses of skeletal and cardiac muscle to exercise. Expert Rev. Proteom. 2011, 8, 361–377. [Google Scholar] [CrossRef] [Green Version]
- Schild, M.; Ruhs, A.; Beiter, T.; Zügel, M.; Hudemann, J.; Reimer, A.; Krumholz-Wagner, I.; Wagner, C.; Keller, J.; Eder, K.; et al. Basal and exercise induced label-free quantitative protein profiling of m. vastus lateralis in trained and untrained individuals. J. Proteom. 2015, 122, 119–132. [Google Scholar] [CrossRef]
- Mckendry, J.; Breen, L.; Shad, B.J.; Greig, C.A. Muscle morphology and performance in master athletes: A systematic review and meta-analyses. Ageing Res. Rev. 2018, 45, 62–82. [Google Scholar] [CrossRef]
- Coudy-Gandilhon, C.; Gueugneau, M.; Chambon, C.; Taillandier, D.; Combaret, L.; Polge, C.; Millet, G.Y.; Féasson, L.; Béchet, D. A Single Bout of Ultra-Endurance Exercise Reveals Early Signs of Muscle Aging in Master Athletes. Int. J. Mol. Sci. 2022, 23, 3713. [Google Scholar] [CrossRef]
- Ubaida-Mohien, C.; Spendiff, S.; Lyashkov, A.; Moaddel, R.; MacMillan, N.J.; Filion, M.E.; Morais, J.A.; Taivassalo, T.; Ferrucci, L.; Hepple, R.T. Unbiased proteomics, histochemistry, and mitochondrial DNA copy number reveal better mitochondrial health in muscle of high-functioning octogenarians. Elife 2022, 11, e74335. [Google Scholar] [CrossRef]
- Poussard, S.; Pires-Alves, A.; Diallo, R.; Dupuy, J.W.; Dargelos, E. A natural antioxidant pine bark extract, Oligopin®, regulates the stress chaperone HSPB1 in human skeletal muscle cells: A proteomics approach. Phytother. Res. 2013, 27, 1529–1535. [Google Scholar] [CrossRef]
- Baraibar, M.A.; Hyzewicz, J.; Rogowska-Wrzesinska, A.; Bulteau, A.L.; Prip-Buus, C.; Butler-Browne, G.; Friguet, B. Impaired energy metabolism of senescent muscle satellite cells is associated with oxidative modifications of glycolytic enzymes. Aging 2016, 8, 3375–3389. [Google Scholar] [CrossRef] [Green Version]
- Medler, S. Mixing it up: The biological significance of hybrid skeletal muscle fibers. J. Exp. Biol. 2019, 222, jeb200832. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, T.; Xu, Z.; Choi, S.J.; Qian, J.; Furdui, C.M.; Register, T.C.; Delbono, O. Myosin heavy chain isoform expression in the Vastus Lateralis muscle of aging African green vervet monkeys. Exp. Gerontol. 2012, 47, 601–607. [Google Scholar] [CrossRef] [Green Version]
- Christian, C.J.; Benian, G.M. Animal models of sarcopenia. Aging Cell 2020, 19, e13223. [Google Scholar] [CrossRef]
- Xie, W.Q.; He, M.; Yu, D.J.; Wu, Y.X.; Wang, X.H.; Lv, S.; Xiao, W.F.; Li, Y.S. Mouse models of sarcopenia: Classification and evaluation. J. Cachexia Sarcopenia Muscle 2021, 12, 538–554. [Google Scholar] [CrossRef]
- Demontis, F.; Piccirillo, R.; Goldberg, A.L.; Perrimon, N. Mechanisms of skeletal muscle aging: Insights from Drosophila and mammalian models. Dis. Model Mech. 2013, 6, 1339–1352. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Cao, Y.; Zhao, J.; Fang, Y.; Liu, N.; Zhang, Y. Multidimensional Proteomics Identifies Declines in Protein Homeostasis and Mitochondria as Early Signals for Normal Aging and Age-associated Disease in Drosophila. Mol. Cell. Proteom. 2019, 18, 2078–2088. [Google Scholar] [CrossRef] [PubMed]
- Yatsenko, A.S.; Kucherenko, M.M.; Xie, Y.; Aweida, D.; Urlaub, H.; Scheibe, R.J.; Cohen, S.; Shcherbata, H.R. Profiling of the muscle-specific dystroglycan interactome reveals the role of Hippo signaling in muscular dystrophy and age-dependent muscle atrophy. BMC Med. 2020, 18, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, A.L. Of worms and women: Sarcopenia and its role in disability and mortality. J. Am. Geriatr. Soc. 2004, 52, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Daya, A.; Donaka, R.; Karasik, D. Zebrafish models of sarcopenia. Dis. Model. Mech. 2020, 13, dmm042689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobon, G.S.; Verrills, N.; Papakostopoulos, P.; Eastwood, H.; Linnane, A.W. The proteomics of ageing. Biogerontology 2002, 3, 133–136. [Google Scholar] [CrossRef]
- Chang, J.; Van Remmen, H.; Cornell, J.; Richardson, A.; Ward, W.F. Comparative proteomics: Characterization of a two-dimensional gel electrophoresis system to study the effect of aging on mitochondrial proteins. Mech. Ageing Dev. 2003, 124, 33–41. [Google Scholar] [CrossRef]
- O'Connell, K.; Ohlendieck, K. Proteomic DIGE analysis of the mitochondria-enriched fraction from aged rat skeletal muscle. Proteomics 2009, 9, 5509–5524. [Google Scholar] [CrossRef] [Green Version]
- Ibebunjo, C.; Chick, J.M.; Kendall, T.; Eash, J.K.; Li, C.; Zhang, Y.; Vickers, C.; Wu, Z.; Clarke, B.A.; Shi, J.; et al. Genomic and proteomic profiling reveals reduced mitochondrial function and disruption of the neuromuscular junction driving rat sarcopenia. Mol. Cell Biol. 2013, 33, 194–212. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.M.; Vitorinom, R.; Figueiredom, P.; Duartem, J.A.; Ferreiram, R.; Amadom, F. Lifelong physical activity modulation of the skeletal muscle mitochondrial proteome in mice. J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65, 832–842. [Google Scholar] [CrossRef] [Green Version]
- Pollard, A.; Shephard, F.; Freed, J.; Liddell, S.; Chakrabarti, L. Mitochondrial proteomic profiling reveals increased carbonic anhydrase II in aging and neurodegeneration. Aging 2016, 8, 2425–2436. [Google Scholar] [CrossRef] [Green Version]
- Stolle, S.; Ciapaite, J.; Reijne, A.C.; Talarovicova, A.; Wolters, J.C.; Aguirre-Gamboa, R.; van der Vlies, P.; de Lange, K.; Neerincx, P.B.; van der Vries, G.; et al. Running-wheel activity delays mitochondrial respiratory flux decline in aging mouse muscle via a post-transcriptional mechanism. Aging Cell 2018, 17, e12700. [Google Scholar] [CrossRef]
- Lu, X.; Gong, Y.; Hu, W.; Mao, Y.; Wang, T.; Sun, Z.; Su, X.; Fu, G.; Wang, Y.; Lai, D. Ultrastructural and proteomic profiling of mitochondria-associated endoplasmic reticulum membranes reveal aging signatures in striated muscle. Cell Death Dis. 2022, 13, 296. [Google Scholar] [CrossRef]
- Lofaro, F.D.; Cisterna, B.; Lacavalla, M.A.; Boschi, F.; Malatesta, M.; Quaglino, D.; Zancanaro, C.; Boraldi, F. Age-Related Changes in the Matrisome of the Mouse Skeletal Muscle. Int. J. Mol. Sci. 2021, 22, 10564. [Google Scholar] [CrossRef]
- Schüler, S.C.; Kirkpatrick, J.M.; Schmidt, M.; Santinha, D.; Koch, P.; Di Sanzo, S.; Cirri, E.; Hemberg, M.; Ori, A.; von Maltzahn, J. Extensive remodeling of the extracellular matrix during aging contributes to age-dependent impairments of muscle stem cell functionality. Cell Rep. 2021, 35, 109223. [Google Scholar] [CrossRef]
- Doran, P.; Gannon, J.; O’Connell, K.; Ohlendieck, K. Aging skeletal muscle shows a drastic increase in the small heat shock proteins alphaB-crystallin/HspB5 and cvHsp/HspB7. Eur. J. Cell Biol. 2007, 86, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Brulé, C.; Dargelos, E.; Diallo, R.; Listrat, A.; Béchet, D.; Cottin, P.; Poussard, S. Proteomic study of calpain interacting proteins during skeletal muscle aging. Biochimie 2010, 92, 1923–1933. [Google Scholar] [CrossRef]
- Wei, L.; Gregorich, Z.R.; Lin, Z.; Cai, W.; Jin, Y.; McKiernan, S.H.; McIlwain, S.; Aiken, J.M.; Moss, R.L.; Diffee, G.M.; et al. Novel Sarcopenia-related Alterations in Sarcomeric Protein Post-translational Modifications (PTMs) in Skeletal Muscles Identified by Top-down Proteomics. Mol. Cell. Proteom. 2018, 17, 134–145. [Google Scholar] [CrossRef] [Green Version]
- O'Connell, K.; Doran, P.; Gannon, J.; Ohlendieck, K. Lectin-based proteomic profiling of aged skeletal muscle: Decreased pyruvate kinase isozyme M1 exhibits drastically increased levels of N-glycosylation. Eur. J. Cell Biol. 2008, 87, 793–805. [Google Scholar] [CrossRef] [Green Version]
- Gannon, J.; Staunton, L.; O’Connell, K.; Doran, P.; Ohlendieck, K. Phosphoproteomic analysis of aged skeletal muscle. Int. J. Mol. Med. 2008, 22, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Xie, H.; Meany, D.L.; Thompson, L.V.; Arriaga, E.A.; Griffin, T.J. Quantitative proteomic profiling of muscle type-dependent and age-dependent protein carbonylation in rat skeletal muscle mitochondria. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 1137–1152. [Google Scholar] [CrossRef] [Green Version]
- Kanski, J.; Alterman, M.A.; Schöneich, C. Proteomic identification of age-dependent protein nitration in rat skeletal muscle. Free Radic. Biol. Med. 2003, 35, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Sharov, V.S.; Galeva, N.A.; Kanski, J.; Williams, T.D.; Schöneich, C. Age-associated tyrosine nitration of rat skeletal muscle glycogen phosphorylase b: Characterization by HPLC-nanoelectrospray-tandem mass spectrometry. Exp. Gerontol. 2006, 41, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Kanski, J.; Schöneich, C. Protein nitration in biological aging: Proteomic and tandem mass spectrometric characterization of nitrated sites. Methods Enzymol. 2005, 396, 160–171. [Google Scholar] [PubMed]
- Kanski, J.; Hong, S.J.; Schöneich, C. Proteomic analysis of protein nitration in aging skeletal muscle and identification of nitrotyrosine-containing sequences in vivo by nanoelectrospray ionization tandem mass spectrometry. J. Biol. Chem. 2005, 280, 24261–24266. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Qiao, X.; Sun, C.; Chu, B.; Meng, J.; Chen, C. GAPDH S-nitrosation contributes to age-related sarcopenia through mediating apoptosis. Nitric Oxide 2022, 120, 1–8. [Google Scholar] [CrossRef]
- Piec, I.; Listrat, A.; Alliot, J.; Chambon, C.; Taylor, R.G.; Béchet, D. Differential proteome analysis of aging in rat skeletal muscle. FASEB J. 2005, 19, 1143–1145. [Google Scholar] [CrossRef]
- O'Connell, K.; Gannon, J.; Doran, P.; Ohlendieck, K. Proteomic profiling reveals a severely perturbed protein expression pattern in aged skeletal muscle. Int. J. Mol. Med. 2007, 20, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Doran, P.; O'Connell, K.; Gannon, J.; Kavanagh, M.; Ohlendieck, K. Opposite pathobiochemical fate of pyruvate kinase and adenylate kinase in aged rat skeletal muscle as revealed by proteomic DIGE analysis. Proteomics 2008, 8, 364–377. [Google Scholar] [CrossRef] [Green Version]
- Capitanio, D.; Vasso, M.; Fania, C.; Moriggi, M.; Viganò, A.; Procacci, P.; Magnaghi, V.; Gelfi, C. Comparative proteomic profile of rat sciatic nerve and gastrocnemius muscle tissues in ageing by 2-D DIGE. Proteomics 2009, 9, 2004–2020. [Google Scholar] [CrossRef]
- Lombardim, A.; Silvestri, E.; Cioffi, F.; Senese, R.; Lanni, A.; Goglia, F.; de Lange, P.; Moreno, M. Defining the transcriptomic and proteomic profiles of rat ageing skeletal muscle by the use of a cDNA array, 2D- and Blue native-PAGE approach. J Proteom. 2009, 72, 708–721. [Google Scholar] [CrossRef]
- Gannon, J.; Doran, P.; Kirwan, A.; Ohlendieck, K. Drastic increase of myosin light chain MLC-2 in senescent skeletal muscle indicates fast-to-slow fibre transition in sarcopenia of old age. Eur. J. Cell Biol. 2009, 88, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Donoghue, P.; Staunton, L.; Mullen, E.; Manning, G.; Ohlendieck, K. DIGE analysis of rat skeletal muscle proteins using nonionic detergent phase extraction of young adult versus aged gastrocnemius tissue. J. Proteom. 2010, 73, 1441–1453. [Google Scholar] [CrossRef] [Green Version]
- Gannon, J.; Ohlendieck, K. Subproteomic analysis of basic proteins in aged skeletal muscle following offgel pre-fractionation. Mol. Med. Rep. 2012, 5, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Carberry, S.; Ohlendieck, K. Gel electrophoresis-based proteomics of senescent tissues. Methods Mol. Biol. 2013, 1048, 229–246. [Google Scholar]
- Faure, C.; Morio, B.; Chafey, P.; Le Plénier, S.; Noirez, P.; Randrianarison-Huetz, V.; Cynober, L.; Aussel, C.; Moinard, C. Citrulline enhances myofibrillar constituents expression of skeletal muscle and induces a switch in muscle energy metabolism in malnourished aged rats. Proteomics. 2013, 13, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Gregorich, Z.R.; Peng, Y.; Cai, W.; Jin, Y.; Wei, L.; Chen, A.J.; McKiernan, S.H.; Aiken, J.M.; Moss, R.L.; Diffee, G.M.; et al. Top-Down Targeted Proteomics Reveals Decrease in Myosin Regulatory Light-Chain Phosphorylation That Contributes to Sarcopenic Muscle Dysfunction. J. Proteome Res. 2016, 15, 2706–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capitanio, D.; Vasso, M.; De Palma, S.; Fania, C.; Torretta, E.; Cammarata, F.P.; Magnaghi, V.; Procacci, P.; Gelfi, C. Specific protein changes contribute to the differential muscle mass loss during ageing. Proteomics 2016, 16, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Li, F.H.; Sun, L.; Wu, D.S.; Gao, H.E.; Min, Z. Proteomics-based identification of different training adaptations of aged skeletal muscle following long-term high-intensity interval and moderate-intensity continuous training in aged rats. Aging 2019, 11, 4159–4182. [Google Scholar] [CrossRef]
- de Sousa Neto, I.V.; Carvalho, M.M.; Marqueti, R.C.; Almeida, J.A.; Oliveira, K.S.; Barin, F.R.; Petriz, B.; de Araújo, H.S.S.; Franco, O.L.; Durigan, J.L.Q. Proteomic changes in skeletal muscle of aged rats in response to resistance training. Cell. Biochem. Funct. 2020, 38, 500–509. [Google Scholar] [CrossRef]
- Gao, H.E.; Wu, D.S.; Sun, L.; Yang, L.D.; Qiao, Y.B.; Ma, S.; Wu, Z.J.; Ruan, L.; Li, F.H. Effects of lifelong exercise on age-related body composition, oxidative stress, inflammatory cytokines, and skeletal muscle proteome in rats. Mech. Ageing Dev. 2020, 189, 111262. [Google Scholar] [CrossRef]
- Kim, J.A.; Vetrivel, P.; Kim, S.M.; Ha, S.E.; Kim, H.H.; Bhosale, P.B.; Heo, J.D.; Lee, W.S.; Senthil, K.; Kim, G.S. Quantitative Proteomics Analysis for the Identification of Differential Protein Expression in Calf Muscles between Young and Old SD Rats Using Mass Spectrometry. ACS Omega 2021, 6, 7422–7433. [Google Scholar] [CrossRef]
- Shembel, A.C.; Kanshin, E.; Ueberheide, B.; Johnson, A.M. Proteomic Characterization of Senescent Laryngeal Adductor and Plantaris Hindlimb Muscles. Laryngoscope 2022, 132, 148–155. [Google Scholar] [CrossRef]
- Barbé, C.; Salles, J.; Chambon, C.; Giraudet, C.; Sanchez, P.; Patrac, V.; Denis, P.; Boirie, Y.; Walrand, S.; Gueugneau, M. Characterization of the Skeletal Muscle Proteome in Undernourished Old Rats. Int. J. Mol. Sci. 2022, 23, 4762. [Google Scholar] [CrossRef]
- Nuss, J.E.; Amaning, J.K.; Bailey, C.E.; DeFord, J.H.; Dimayuga, V.L.; Rabek, J.P.; Papaconstantinou, J. Oxidative modification and aggregation of creatine kinase from aged mouse skeletal muscle. Aging 2009, 1, 557–572. [Google Scholar] [CrossRef] [Green Version]
- Hwang, C.Y.; Kim, K.; Choi, J.Y.; Bahn, Y.J.; Lee, S.M.; Kim, Y.K.; Lee, C.; Kwon, K.S. Quantitative proteome analysis of age-related changes in mouse gastrocnemius muscle using mTRAQ. Proteomics 2014, 14, 121–132. [Google Scholar] [CrossRef]
- McDonagh, B.; Sakellariou, G.K.; Smith, N.T.; Brownridge, P.; Jackson, M.J. Differential cysteine labeling and global label-free proteomics reveals an altered metabolic state in skeletal muscle aging. J. Proteome Res. 2014, 13, 5008–5021. [Google Scholar] [CrossRef]
- Huang, Y.L.; Shen, Z.Q.; Wu, C.Y.; Teng, Y.C.; Liao, C.C.; Kao, C.H.; Chen, L.K.; Lin, C.H.; Tsai, T.F. Comparative proteomic profiling reveals a role for Cisd2 in skeletal muscle aging. Aging Cell 2018, 17, e12705. [Google Scholar] [CrossRef]
- McDonagh, B.; Sakellariou, G.K.; Smith, N.T.; Brownridge, P.; Jackson, M.J. Redox proteomic analysis of the gastrocnemius muscle from adult and old mice. Data Brief 2015, 4, 344–348. [Google Scholar] [CrossRef] [Green Version]
- Smith, N.T.; Soriano-Arroquia, A.; Goljanek-Whysall, K.; Jackson, M.J.; McDonagh, B. Redox responses are preserved across muscle fibres with differential susceptibility to aging. J. Proteom. 2018, 177, 112–123. [Google Scholar] [CrossRef]
- Kelley, R.C.; McDonagh, B.; Ferreira, L.F. Advanced aging causes diaphragm functional abnormalities, global proteome remodeling, and loss of mitochondrial cysteine redox flexibility in mice. Exp. Gerontol. 2018, 103, 69–79. [Google Scholar] [CrossRef]
- Sataranatarajan, K.; Pharaoh, G.; Brown, J.L.; Ranjit, R.; Piekarz, K.M.; Street, K.; Wren, J.D.; Georgescu, C.; Kinter, C.; Kinter, M.; et al. Molecular changes in transcription and metabolic pathways underlying muscle atrophy in the CuZnSOD null mouse model of sarcopenia. Geroscience 2020, 42, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Bareja, A.; Draper, J.A.; Katz, L.H.; Lee, D.E.; Grimsrud, P.A.; White, J.P. Chronic caloric restriction maintains a youthful phosphoproteome in aged skeletal muscle. Mech. Ageing Dev. 2021, 195, 111443. [Google Scholar] [CrossRef]
- Hunt, L.C.; Graca, F.A.; Pagala, V.; Wang, Y.D.; Li, Y.; Yuan, Z.F.; Fan, Y.; Labelle, M.; Peng, J.; Demontis, F. Integrated genomic and proteomic analyses identify stimulus-dependent molecular changes associated with distinct modes of skeletal muscle atrophy. Cell Rep. 2021, 37, 109971. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.M.; Deemer, S.E.; Smith, D.L., Jr.; Mobley, J.A.; Musi, N.; Plaisance, E.P. Effects of an exogenous ketone ester using multi-omics in skeletal muscle of aging C57BL/6J male mice. Front. Nutr. 2022, 9, 1041026. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.D.; Martín-Pérez, M.; Egertson, J.D.; Gaffrey, M.J.; Wang, L.; Bammler, T.; Rabinovitch, P.S.; MacCoss, M.; Qian, W.J.; Villen, J.; et al. Elamipretide effects on the skeletal muscle phosphoproteome in aged female mice. Geroscience 2022, 44, 2913–2924. [Google Scholar] [CrossRef] [PubMed]
- Jessica Lo, H.T.; Yiu, T.L.; Wang, Y.; Feng, L.; Li, G.; Lui, M.P.; Lee, W.Y. Fetal muscle extract improves muscle function and performance in aged mice. Front. Physiol. 2022, 13, 816774. [Google Scholar] [CrossRef]
- Ryan, M.; Carlson, B.M.; Ohlendieck, K. Oligomeric status of the dihydropyridine receptor in aged skeletal muscle. Mol. Cell. Biol. Res. Commun. 2000, 4, 224–229. [Google Scholar] [CrossRef]
- Ryan, M.; Butler-Browne, G.; Erzen, I.; Mouly, V.; Thornell, L.E.; Wernig, A.; Ohlendieck, K. Persistent expression of the alpha1S-dihydropyridine receptor in aged human skeletal muscle: Implications for the excitation-contraction uncoupling hypothesis of sarcopenia. Int. J. Mol. Med. 2003, 11, 425–434. [Google Scholar]
- O'Connell, K.; Gannon, J.; Doran, P.; Ohlendieck, K. Reduced expression of sarcalumenin and related Ca2+-regulatory proteins in aged rat skeletal muscle. Exp. Gerontol. 2008, 43, 958–961. [Google Scholar] [CrossRef] [Green Version]
- Weisleder, N.; Brotto, M.; Komazaki, S.; Pan, Z.; Zhao, X.; Nosek, T.; Parness, J.; Takeshima, H.; Ma, J. Muscle aging is associated with compromised Ca2+ spark signaling and segregated intracellular Ca2+ release. J. Cell Biol. 2006, 174, 639–645. [Google Scholar] [CrossRef]
- Delbono, O. Expression and regulation of excitation-contraction coupling proteins in aging skeletal muscle. Curr. Aging Sci. 2011, 4, 248–259. [Google Scholar] [CrossRef]
- Qaisar, R.; Bhaskaran, S.; Premkumar, P.; Ranjit, R.; Natarajan, K.S.; Ahn, B.; Riddle, K.; Claflin, D.R.; Richardson, A.; Brooks, S.V.; et al. Oxidative stress-induced dysregulation of excitation-contraction coupling contributes to muscle weakness. J. Cachexia Sarcopenia Muscle 2018, 9, 1003–1017. [Google Scholar] [CrossRef]
- Pietrangelo, L.; D'Incecco, A.; Ainbinder, A.; Michelucci, A.; Kern, H.; Dirksen, R.T.; Boncompagni, S.; Protasi, F. Age-dependent uncoupling of mitochondria from Ca2⁺ release units in skeletal muscle. Oncotarget 2015, 6, 35358–35371. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.B.; Wang, J.Q.; Meng, X.H.; Luo, Z.; Liu, X.W.; Shen, H.; Xiao, H.M.; Deng, H.W. Putative Candidate Drug Targets for Sarcopenia-Related Traits Identified Through Mendelian Randomization Analysis of the Blood Proteome. Front. Genet. 2022, 13, 923429. [Google Scholar] [CrossRef]
- Narici, M.; McPhee, J.; Conte, M.; Franchi, M.V.; Mitchell, K.; Tagliaferri, S.; Monti, E.; Marcolin, G.; Atherton, P.J.; Smith, K.; et al. Age-related alterations in muscle architecture are a signature of sarcopenia: The ultrasound sarcopenia index. J. Cachexia Sarcopenia Muscle 2021, 12, 973–982. [Google Scholar] [CrossRef]
- Calvani, R.; Marini, F.; Cesari, M.; Tosato, M.; Picca, A.; Anker, S.D.; von Haehling, S.; Miller, R.R.; Bernabei, R.; Landi, F.; et al. SPRINTT Consortium. Biomarkers for physical frailty and sarcopenia. Aging Clin. Exp. Res. 2017, 29, 29–34. [Google Scholar] [CrossRef]
- Picca, A.; Coelho-Junior, H.J.; Calvani, R.; Marzetti, E.; Vetrano, D.L. Biomarkers shared by frailty and sarcopenia in older adults: A systematic review and meta-analysis. Ageing Res. Rev. 2022, 73, 101530. [Google Scholar] [CrossRef]
- Ohlendieck, K. Proteomic identification of biomarkers of skeletal muscle disorders. Biomark. Med. 2013, 7, 169–186. [Google Scholar] [CrossRef]
- Murphy, S.; Zweyer, M.; Mundegar, R.R.; Swandulla, D.; Ohlendieck, K. Proteomic serum biomarkers for neuromuscular diseases. Expert Rev. Proteom. 2018, 15, 277–291. [Google Scholar] [CrossRef]
- Stalmach, A.; Boehm, I.; Fernandes, M.; Rutter, A.; Skipworth, R.J.E.; Husi, H. Gene Ontology (GO)-Driven Inference of Candidate Proteomic Markers Associated with Muscle Atrophy Conditions. Molecules 2022, 27, 5514. [Google Scholar] [CrossRef]
- Lin, C.H.; Liao, C.C.; Huang, C.H.; Tung, Y.T.; Chang, H.C.; Hsu, M.C.; Huang, C.C. Proteomics Analysis to Identify and Characterize the Biomarkers and Physical Activities of Non-Frail and Frail Older Adults. Int. J. Med. Sci. 2017, 14, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dlamini, S.N.; Norris, S.A.; Mendham, A.E.; Mtintsilana, A.; Ward, K.A.; Olsson, T.; Goedecke, J.H.; Micklesfield, L.K. Targeted proteomics of appendicular skeletal muscle mass and handgrip strength in black South Africans: A cross-sectional study. Sci. Rep. 2022, 12, 9512. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Pan, S.; Xanthakis, V.; Vasan, R.S.; Psaty, B.M.; Austin, T.R.; Newman, A.B.; Sanders, J.L.; Wu, C.; Tracy, R.P.; et al. Plasma proteomic signature of decline in gait speed and grip strength. Aging Cell. 2022, 21, e13736. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Calvani, R.; Coelho-Júnior, H.J.; Marini, F.; Landi, F.; Marzetti, E. Circulating Inflammatory, Mitochondrial Dysfunction, and Senescence-Related Markers in Older Adults with Physical Frailty and Sarcopenia: A BIOSPHERE Exploratory Study. Int. J. Mol. Sci. 2022, 23, 14006. [Google Scholar] [CrossRef]
- Fielding, R.A.; Atkinson, E.J.; Aversa, Z.; White, T.A.; Heeren, A.A.; Achenbach, S.J.; Mielke, M.M.; Cummings, S.R.; Pahor, M.; Leeuwenburgh, C.; et al. Associations between biomarkers of cellular senescence and physical function in humans: Observations from the lifestyle interventions for elders (LIFE) study. Geroscience. 2022, 44, 2757–2770. [Google Scholar] [CrossRef]
- Kwak, J.Y.; Hwang, H.; Kim, S.K.; Choi, J.Y.; Lee, S.M.; Bang, H.; Kwon, E.S.; Lee, K.P.; Chung, S.G.; Kwon, K.S. Prediction of sarcopenia using a combination of multiple serum biomarkers. Sci. Rep. 2018, 8, 8574. [Google Scholar] [CrossRef]
- Wu, J.; Cao, L.; Wang, J.; Wang, Y.; Hao, H.; Huang, L. Characterization of serum protein expression profiles in the early sarcopenia older adults with low grip strength: A cross-sectional study. BMC Musculoskelet. Disord. 2022, 23, 894. [Google Scholar] [CrossRef]
- Van Long, N.; Chienm, P.N.; Tung, T.X.; Van Anh, L.T.; Giang, N.N.; Nga, P.T.; Linh, L.T.T.; Nam, S.Y.; Heo, C.Y. Complementary combination of biomarkers for diagnosis of sarcopenia in C57BL/6J mice. Life Sci. 2022, 312, 121213. [Google Scholar] [CrossRef]
- Chew, J.; Tay, L.; Lim, J.P.; Leung, B.P.; Yeo, A.; Yew, S.; Ding, Y.Y.; Lim, W.S. Serum Myostatin and IGF-1 as Gender-Specific Biomarkers of Frailty and Low Muscle Mass in Community-Dwelling Older Adults. J. Nutr. Health Aging 2019, 23, 979–986. [Google Scholar] [CrossRef]
- Perez, K.; Ciotlos, S.; McGirr, J.; Limbad, C.; Doi, R.; Nederveen, J.P.; Nilsson, M.I.; Winer, D.A.; Evans, W.; Tarnopolsky, M.; et al. Single nuclei profiling identifies cell specific markers of skeletal muscle aging, frailty, and senescence. Aging 2022, 14, 9393–9422. [Google Scholar] [CrossRef]
- Zhao, Q.; Shen, H.; Liu, J.; Chiu, C.Y.; Su, K.J.; Tian, Q.; Kakhniashvili, D.; Qiu, C.; Zhao, L.J.; Luo, Z.; et al. Pathway-based metabolomics study of sarcopenia-related traits in two US cohorts. Aging 2022, 14, 2101–2112. [Google Scholar] [CrossRef]
- Tsai, J.S.; Wang, S.Y.; Chang, C.H.; Chen, C.Y.; Wen, C.J.; Chen, G.Y.; Kuo, C.H.; Tseng, Y.J.; Chen, C.Y. Identification of traumatic acid as a potential plasma biomarker for sarcopenia using a metabolomics-based approach. J. Cachexia Sarcopenia Muscle 2022, 13, 276–286. [Google Scholar] [CrossRef]
- Drey, M.; Sieber, C.C.; Bauer, J.M.; Uter, W.; Dahinden, P.; Fariello, R.G.; Vrijbloed, J.W.; FiAT Intervention Group. C-terminal Agrin Fragment as a potential marker for sarcopenia caused by degeneration of the neuromuscular junction. Exp. Gerontol. 2013, 48, 76–80. [Google Scholar] [CrossRef]
- Hettwer, S.; Dahinden, P.; Kucsera, S.; Farina, C.; Ahmed, S.; Fariello, R.; Drey, M.; Sieber, C.C.; Vrijbloed, J.W. Elevated levels of a C-terminal agrin fragment identifies a new subset of sarcopenia patients. Exp. Gerontol. 2013, 48, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Bütikofer, L.; Zurlinden, A.; Bolliger, M.F.; Kunz, B.; Sonderegger, P. Destabilization of the neuromuscular junction by proteolytic cleavage of agrin results in precocious sarcopenia. FASEB J. 2011, 25, 4378–4393. [Google Scholar] [CrossRef] [Green Version]
- Hettwer, S.; Lin, S.; Kucsera, S.; Haubitz, M.; Oliveri, F.; Fariello, R.G.; Ruegg, M.A.; Vrijbloed, J.W. Injection of a soluble fragment of neural agrin (NT-1654) considerably improves the muscle pathology caused by the disassembly of the neuromuscular junction. PLoS One 2014, 9, e88739. [Google Scholar] [CrossRef]
- Shimada, H.; Makizako, H.; Doi, T.; Yoshida, D.; Tsutsumimoto, K.; Anan, Y.; Uemura, K.; Lee, S.; Park, H.; Suzuki, T. A large, cross-sectional observational study of serum BDNF, cognitive function, and mild cognitive impairment in the elderly. Front. Aging Neurosci. 2014, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, S.; Iino, N.; Koda, R.; Narita, I.; Kaneko, Y. Brain-derived neurotrophic factor is associated with sarcopenia and frailty in Japanese hemodialysis patients. Geriatr. Gerontol. Int. 2021, 21, 27–33. [Google Scholar] [CrossRef]
- Kaneko, Y. Brain-derived neurotrophic factor as a potential biomarker for sarcopenia and frailty in hemodialysis patients. Geriatr. Gerontol. Int. 2021, 21, 874–875. [Google Scholar] [CrossRef]
- Belhasan, D.C.; Akaaboune, M. The role of the dystrophin glycoprotein complex on the neuromuscular system. Neurosci. Lett. 2020, 722, 134833. [Google Scholar] [CrossRef]
- Sirago, G.; Pellegrino, M.A.; Bottinelli, R.; Franchi, M.V.; Narici, M.V. Loss of neuromuscular junction integrity and muscle atrophy in skeletal muscle disuse. Ageing Res. Rev. 2022, 83, 101810. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Pais, A.; Ferreira, R.; Oliveira, P.A.; Duarte, J.A. A neuromuscular perspective of sarcopenia pathogenesis: Deciphering the signaling pathways involved. Geroscience 2022, 44, 1199–1213. [Google Scholar] [CrossRef] [PubMed]
- Guarino, S.R.; Canciani, A.; Forneris, F. Dissecting the Extracellular Complexity of Neuromuscular Junction Organizers. Front. Mol. Biosci. 2020, 6, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolf, R.; Khan, M.M.; Labeit, S.; Deschenes, M.R. Degeneration of neuromuscular junction in age and dystrophy. Front. Aging Neurosci. 2014, 6, 00099. [Google Scholar] [CrossRef] [Green Version]
- da Orssatto, L.B.R.; Wiest, M.J.; Diefenthaeler, F. Neural and musculotendinous mechanisms underpinning age-related force reductions. Mech. Ageing Dev. 2018, 175, 17–23. [Google Scholar] [CrossRef]
- Kwon, Y.N.; Yoon, S.S. Sarcopenia: Neurological point of view. J. Bone Metab. 2017, 24, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Larsson, L. Force-generating capacity of human myosin isoforms extracted from single muscle fibre segments. J. Physiol. 2010, 588, 5105–5114. [Google Scholar] [CrossRef]
- Verdijk, L.B.; Snijders, T.; Beelen, M.; Savelberg, H.H.; Meijer, K.; Kuipers, H.; Van Loon, L.J. Characteristics of muscle fiber type are predictive of skeletal muscle mass and strength in elderly men. J. Am. Geriatr. Soc. 2010, 58, 2069–2075. [Google Scholar] [CrossRef]
- Friedman, S.M. Lifestyle (Medicine) and Healthy Aging. Clin. Geriatr. Med. 2020, 36, 645–653. [Google Scholar] [CrossRef]
- Eckstrom, E.; Neukam, S.; Kalin, L.; Wright, J. Physical Activity and Healthy Aging. Clin. Geriatr. Med. 2020, 36, 671–683. [Google Scholar] [CrossRef]
- Roberts, S.B.; Silver, R.E.; Das, S.K.; Fielding, R.A.; Gilhooly, C.H.; Jacques, P.F.; Kelly, J.M.; Mason, J.B.; McKeown, N.M.; Reardon, M.A.; et al. Healthy Aging-Nutrition Matters: Start Early and Screen Often. Adv. Nutr. 2021, 12, 1438–1448. [Google Scholar] [CrossRef]
- Yeung, S.S.Y.; Kwan, M.; Woo, J. Healthy Diet for Healthy Aging. Nutrients 2021, 13, 4310. [Google Scholar] [CrossRef]
- Kirwan, R.; McCullough, D.; Butler, T.; Perez de Heredia, F.; Davies, I.G.; Stewart, C. Sarcopenia during COVID-19 lockdown restrictions: Long-term health effects of short-term muscle loss. Geroscience 2020, 42, 1547–1578. [Google Scholar] [CrossRef]
- Shur, N.F.; Creedon, L.; Skirrow, S.; Atherton, P.J.; MacDonald, I.A.; Lund, J.; Greenhaff, P.L. Age-related changes in muscle architecture and metabolism in humans: The likely contribution of physical inactivity to age-related functional decline. Ageing Res. Rev. 2021, 68, 101344. [Google Scholar] [CrossRef]
- Demonceau, C.; Beaudart, C.; Reginster, J.Y.; Veronese, N.; Bruyère, O. The interconnection between Covid-19, sarcopenia and lifestyle. Maturitas 2022, S0378-5122(22)00204-3. [Google Scholar] [CrossRef]
- Welch, C.; Greig, C.; Masud, T.; Wilson, D.; Jackson, T.A. COVID-19 and Acute Sarcopenia. Aging Dis. 2020, 11, 1345–1351. [Google Scholar] [CrossRef]
- Soares, M.N.; Eggelbusch, M.; Naddaf, E.; Gerrits, K.H.L.; van der Schaaf, M.; van den Borst, B.; Wiersinga, W.J.; van Vugt, M.; Weijs, P.J.M.; Murray, A.J.; et al. Skeletal muscle alterations in patients with acute Covid-19 and post-acute sequelae of Covid-19. J. Cachexia Sarcopenia Muscle 2022, 13, 11–22. [Google Scholar] [CrossRef]
- Piotrowicz, K.; Gąsowski, J.; Michel, J.P.; Veronese, N. Post-COVID-19 acute sarcopenia: Physiopathology and management. Aging Clin. Exp. Res. 2021, 33, 2887–2898. [Google Scholar] [CrossRef]
- Wierdsma, N.J.; Kruizenga, H.M.; Konings, L.A.; Krebbers, D.; Jorissen, J.R.; Joosten, M.I.; van Aken, L.H.; Tan, F.M.; van Bodegraven, A.A.; Soeters, M.R.; et al. Poor nutritional status, risk of sarcopenia and nutrition related complaints are prevalent in COVID-19 patients during and after hospital admission. Clin. Nutr. ESPEN 2021, 43, 369–376. [Google Scholar] [CrossRef]
- Damanti, S.; Cristel, G.; Ramirez, G.A.; Bozzolo, E.P.; Da Prat, V.; Gobbi, A.; Centurioni, C.; Di Gaeta, E.; Del Prete, A.; Calabrò, M.G.; et al. Influence of reduced muscle mass and quality on ventilator weaning and complications during intensive care unit stay in COVID-19 patients. Clin. Nutr. 2022, 41, 2965–2972. [Google Scholar] [CrossRef]
- Lu, L.; Mao, L.; Feng, Y.; Ainsworth, B.E.; Liu, Y.; Chen, N. Effects of different exercise training modes on muscle strength and physical performance in older people with sarcopenia: A systematic review and meta-analysis. BMC Geriatr. 2021, 21, 708. [Google Scholar] [CrossRef] [PubMed]
- Tsekoura, M.; Billis, E.; Kastrinis, A.; Katsoulaki, M.; Fousekis, K.; Tsepis, E.; Konstantoudaki, X.; Gliatis, J. The Effects of Exercise in Patients with Sarcopenia. Adv. Exp. Med. Biol. 2021, 1337, 281–290. [Google Scholar] [PubMed]
- Gryson, C.; Ratel, S.; Rance, M.; Penando, S.; Bonhomme, C.; Le Ruyet, P.; Duclos, M.; Boirie, Y.; Walrand, S. Four-month course of soluble milk proteins interacts with exercise to improve muscle strength and delay fatigue in elderly participants. J. Am. Med. Dir. Assoc. 2014, 15, 958.e1-9. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Arnau, F.M.; Fonfría-Vivas, R.; Buigues, C.; Castillo, Y.; Molina, P.; Hoogland, A.J.; van Doesburg, F.; Pruimboom, L.; Fernández-Garrido, J.; Cauli, O. Effects of Leucine Administration in Sarcopenia: A Randomized and Placebo-controlled Clinical Trial. Nutrients 2020, 12, 932. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.M.; Verlaan, S.; Bautmans, I.; Brandt, K.; Donini, L.M.; Maggio, M.; McMurdo, M.E.; Mets, T.; Seal, C.; Wijers, S.L.; et al. Effects of a vitamin D and leucine-enriched whey protein nutritional supplement on measures of sarcopenia in older adults, the PROVIDE study: A randomized, double-blind, placebo-controlled trial. J. Am. Med. Dir. Assoc. 2015, 16, 740–747. [Google Scholar] [CrossRef]
- Yoo, J.I.; Chung, H.J.; Kim, B.G.; Jung, Y.K.; Baek, K.W.; Song, M.G.; Cho, M.C. Comparative analysis of the association between various serum vitamin D biomarkers and sarcopenia. J. Clin. Lab. Anal. 2021, 35, e23946. [Google Scholar] [CrossRef]
- Negm, A.M.; Lee, J.; Hamidian, R.; Jones, C.A.; Khadaroo, R.G. Management of Sarcopenia: A Network Meta-Analysis of Randomized Controlled Trials. J. Am. Med. Dir. Assoc. 2022, 23, 707–714. [Google Scholar] [CrossRef]
- Bernabei, R.; Landi, F.; Calvani, R.; Cesari, M.; Del Signore, S.; Anker, S.D.; Bejuit, R.; Bordes, P.; Cherubini, A.; Cruz-Jentoft, A.J.; et al. Multicomponent intervention to prevent mobility disability in frail older adults: Randomised controlled trial (SPRINTT project). BMJ. 2022, 377, e068788. [Google Scholar] [CrossRef]
- Yan, Z.; Okutsu, M.; Akhtar, Y.N.; Lira, V.A. Regulation of exercise-induced fiber type transformation, mitochondrial biogenesis, and angiogenesis in skeletal muscle. J. Appl. Physiol. 2011, 110, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.M.; Loenneke, J.P.; Jo, E.; Wilson, G.J.; Zourdos, M.C.; Kim, J.S. The effects of endurance, strength, and power training on muscle fiber type shifting. J. Strength Cond. Res. 2012, 26, 1724–1729. [Google Scholar] [CrossRef]
- Qaisar, R.; Bhaskaran, S.; Van Remmen, H. Muscle fiber type diversification during exercise and regeneration. Free Radic. Biol. Med. 2016, 98, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Coletti, C.; Acosta, G.F.; Keslacy, S.; Coletti, D. Exercise-mediated reinnervation of skeletal muscle in elderly people: An update. Eur. J. Transl. Myol. 2022, 32, 10416. [Google Scholar] [CrossRef] [PubMed]
- Blocquiaux, S.; Gorski, T.; Van Roie, E.; Ramaekers, M.; Van Thienen, R.; Nielens, H.; Delecluse, C.; De Bock, K.; Thomis, M. The effect of resistance training, detraining and retraining on muscle strength and power, myofibre size, satellite cells and myonuclei in older men. Exp. Gerontol. 2020, 133, 110860. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.S.; Callahan, D.M.; Tourville, T.W.; Slauterbeck, J.R.; Kaplan, A.; Fiske, B.R.; Savage, P.D.; Ades, P.A.; Beynnon, B.D.; Toth, M.J. Moderate-intensity resistance exercise alters skeletal muscle molecular and cellular structure and function in inactive older adults with knee osteoarthritis. J. Appl. Physiol. 2017, 122, 775–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesquita, P.H.C.; Lamb, D.A.; Parry, H.A.; Moore, J.H.; Smith, M.A.; Vann, C.G.; Osburn, S.C.; Fox, C.D.; Ruple, B.A.; Huggins, K.W.; et al. Acute and chronic effects of resistance training on skeletal muscle markers of mitochondrial remodeling in older adults. Physiol. Rep. 2020, 8, e14526. [Google Scholar] [CrossRef]
- Moro, T.; Brightwell, C.R.; Volpi, E.; Rasmussen, B.B.; Fry, C.S. Resistance exercise training promotes fiber type-specific myonuclear adaptations in older adults. J. Appl. Physiol. 2020, 128, 795–804. [Google Scholar] [CrossRef]
- Fry, C.S.; Noehren, B.; Mula, J.; Ubele, M.F.; Westgate, P.M.; Kern, P.A.; Peterson, C.A. Fibre type-specific satellite cell response to aerobic training in sedentary adults. J. Physiol. 2014, 592, 2625–2635. [Google Scholar] [CrossRef]
- Nilwik, R.; Snijders, T.; Leenders, M.; Groen, B.B.; van Kranenburg, J.; Verdijk, L.B.; van Loon, L.J. The decline in skeletal muscle mass with aging is mainly attributed to a reduction in type II muscle fiber size. Exp. Gerontol. 2013, 48, 492–498. [Google Scholar] [CrossRef]
- Kraková, D.; Holwerda, A.M.; Betz, M.W.; Lavin, K.M.; Bamman, M.M.; van Loon, L.J.C.; Verdijk, L.B.; Snijders, T. Muscle fiber type grouping does not change in response to prolonged resistance exercise training in healthy older men. Exp. Gerontol. 2023, 173, 112083. [Google Scholar] [CrossRef]
- Li, Y.; Chen, M.; Zhao, Y.; Li, M.; Qin, Y.; Cheng, S.; Yang, Y.; Yin, P.; Zhang, L.; Tang, P. Advance in Drug Delivery for Ageing Skeletal Muscle. Front. Pharmacol. 2020, 11, 1016. [Google Scholar] [CrossRef]
- Kim, J.W.; Kim, R.; Choi, H.; Lee, S.J.; Bae, G.U. Understanding of sarcopenia: From definition to therapeutic strategies. Arch. Pharm. Res. 2021, 44, 876–889. [Google Scholar] [CrossRef]
- Canfora, I.; Tarantino, N.; Pierno, S. Metabolic Pathways and Ion Channels Involved in Skeletal Muscle Atrophy: A Starting Point for Potential Therapeutic Strategies. Cells 2022, 11, 2566. [Google Scholar] [CrossRef]
- Huang, L.; Li, M.; Deng, C.; Qiu, J.; Wang, K.; Chang, M.; Zhou, S.; Gu, Y.; Shen, Y.; Wang, W.; et al. Potential Therapeutic Strategies for Skeletal Muscle Atrophy. Antioxidants 2023, 12, 44. [Google Scholar] [CrossRef]
- Sartori, R.; Romanello, V.; Sandri, M. Mechanisms of muscle atrophy and hypertrophy: Implications in health and disease. Nat. Commun. 2021, 12, 330. [Google Scholar] [CrossRef]
- Guo, M.; Yao, J.; Li, J.; Zhang, J.; Wang, D.; Zuo, H.; Zhang, Y.; Xu, B.; Zhong, Y.; Shen, F.; et al. Irisin ameliorates age-associated sarcopenia and metabolic dysfunction. J. Cachexia Sarcopenia Muscle 2022, in press. [Google Scholar] [CrossRef]
- Vainshtein, A.; Sandri, M. Signaling Pathways That Control Muscle Mass. Int. J. Mol. Sci. 2020, 21, 4759. [Google Scholar] [CrossRef]
- Penniman, C.M.; Bhardwaj, G.; Nowers, C.J.; Brown, C.U.; Junck, T.L.; Boyer, C.K.; Jena, J.; Fuqua, J.D.; Lira, V.A.; O’Neill, B.T. Loss of FoxOs in muscle increases strength and mitochondrial function during aging. J. Cachexia Sarcopenia Muscle 2022, in press. [Google Scholar] [CrossRef]
- Sirago, G.; Picca, A.; Calvani, R.; Coelho-Júnior, H.J.; Marzetti, E. Mammalian Target of Rapamycin (mTOR) Signaling at the Crossroad of Muscle Fiber Fate in Sarcopenia. Int. J. Mol. Sci. 2022, 23, 13823. [Google Scholar] [CrossRef]
- White, T.A.; LeBrasseur, N.K. Myostatin and sarcopenia: Opportunities and challenges—A mini-review. Gerontology 2014, 60, 289–293. [Google Scholar] [CrossRef]
- Curiel-Cervantes, V.; Solis-Sainz, J.C.; Camacho-Barrón, M.; Aguilar-Galarza, A.; Valencia, M.E.; Anaya-Loyola, M.A. Systematic training in master swimmer athletes increases serum insulin growth factor-1 and decreases myostatin and irisin levels. Growth Factors 2022, 40, 1–12. [Google Scholar] [CrossRef]
- Baig, M.H.; Ahmad, K.; Moon, J.S.; Park, S.Y.; Ho Lim, J.; Chun, H.J.; Qadri, A.F.; Hwang, Y.C.; Jan, A.T.; Ahmad, S.S.; et al. Myostatin and its Regulation: A Comprehensive Review of Myostatin Inhibiting Strategies. Front. Physiol. 2022, 13, 876078. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.S.; Ahmad, K.; Lee, E.J.; Shaikh, S.; Choi, I. Computational Identification of Dithymoquinone as a Potential Inhibitor of Myostatin and Regulator of Muscle Mass. Molecules 2021, 26, 5407. [Google Scholar] [CrossRef] [PubMed]
- Camporez, J.P.; Petersen, M.C.; Abudukadier, A.; Moreira, G.V.; Jurczak, M.J.; Friedman, G.; Haqq, C.M.; Petersen, K.F.; Shulman, G.I. Anti-myostatin antibody increases muscle mass and strength and improves insulin sensitivity in old mice. Proc. Natl. Acad. Sci. USA. 2016, 113, 2212–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, H.O.; Cerqueira, H.S.; Tinsley, G.M. The Effects of Dietary Supplements, Nutraceutical Agents, and Physical Exercise on Myostatin Levels: Hope or Hype? Metabolites 2022, 12, 1146. [Google Scholar] [CrossRef] [PubMed]
- Winnard, A.; Scott, J.; Waters, N.; Vance, M.; Caplan, N. Effect of Time on Human Muscle Outcomes During Simulated Microgravity Exposure Without Countermeasures-Systematic Review. Front. Physiol. 2019, 10, 1046. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.H.U.; Chung, M.; Ren, Z.; Mair, D.B.; Kim, D.H. Factors mediating spaceflight-induced skeletal muscle atrophy. Am. J. Physiol. Cell Physiol. 2022, 322, C567–C580. [Google Scholar] [CrossRef]
- Schulz, H.; Strauch, S.M.; Richter, P.; Wehland, M.; Krüger, M.; Sahana, J.; Corydon, T.J.; Wise, P.; Baran, R.; Lebert, M.; et al. Latest knowledge about changes in the proteome in microgravity. Expert Rev. Proteom. 2022, 19, 43–59. [Google Scholar] [CrossRef]
- Cannavo, A.; Carandina, A.; Corbi, G.; Tobaldini, E.; Montano, N.; Arosio, B. Are Skeletal Muscle Changes during Prolonged Space Flights Similar to Those Experienced by Frail and Sarcopenic Older Adults? Life 2022, 12, 2139. [Google Scholar] [CrossRef]
- Takahashi, H.; Nakamura, A.; Shimizu, T. Simulated microgravity accelerates aging of human skeletal muscle myoblasts at the single cell level. Biochem. Biophys. Res. Commun. 2021, 578, 115–121. [Google Scholar] [CrossRef]







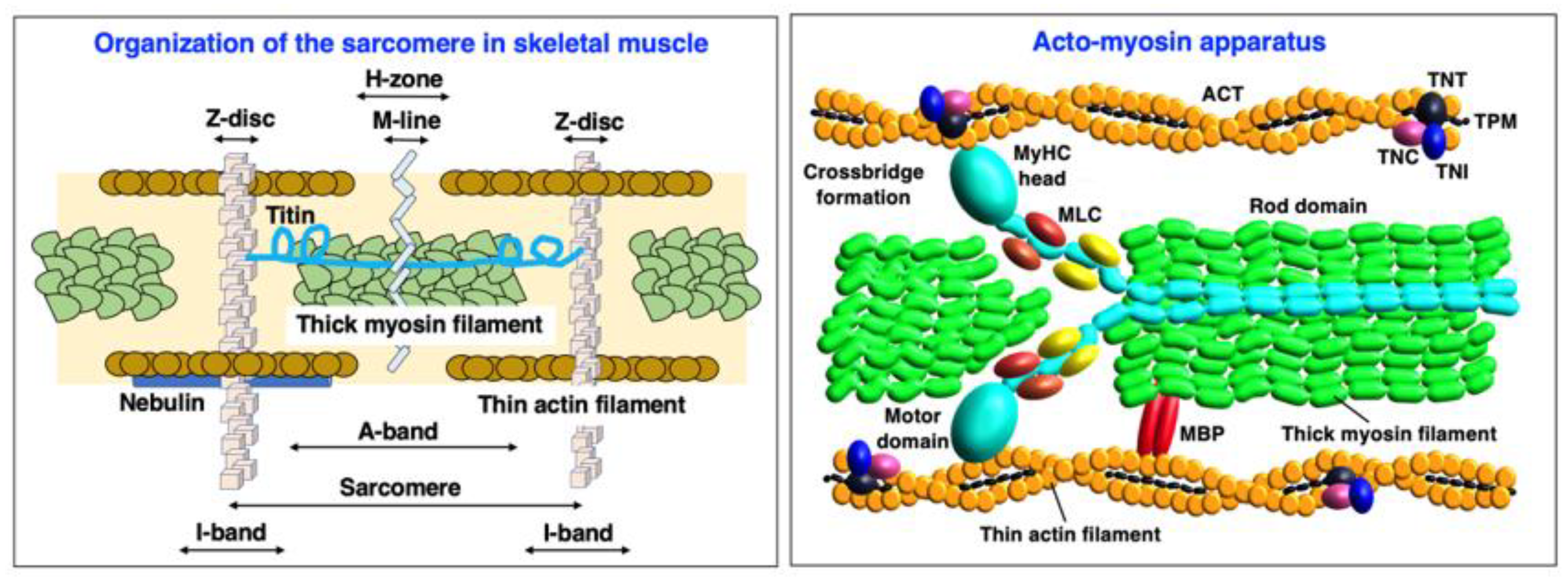{kind=link}
{kind=link}
{kind=link}
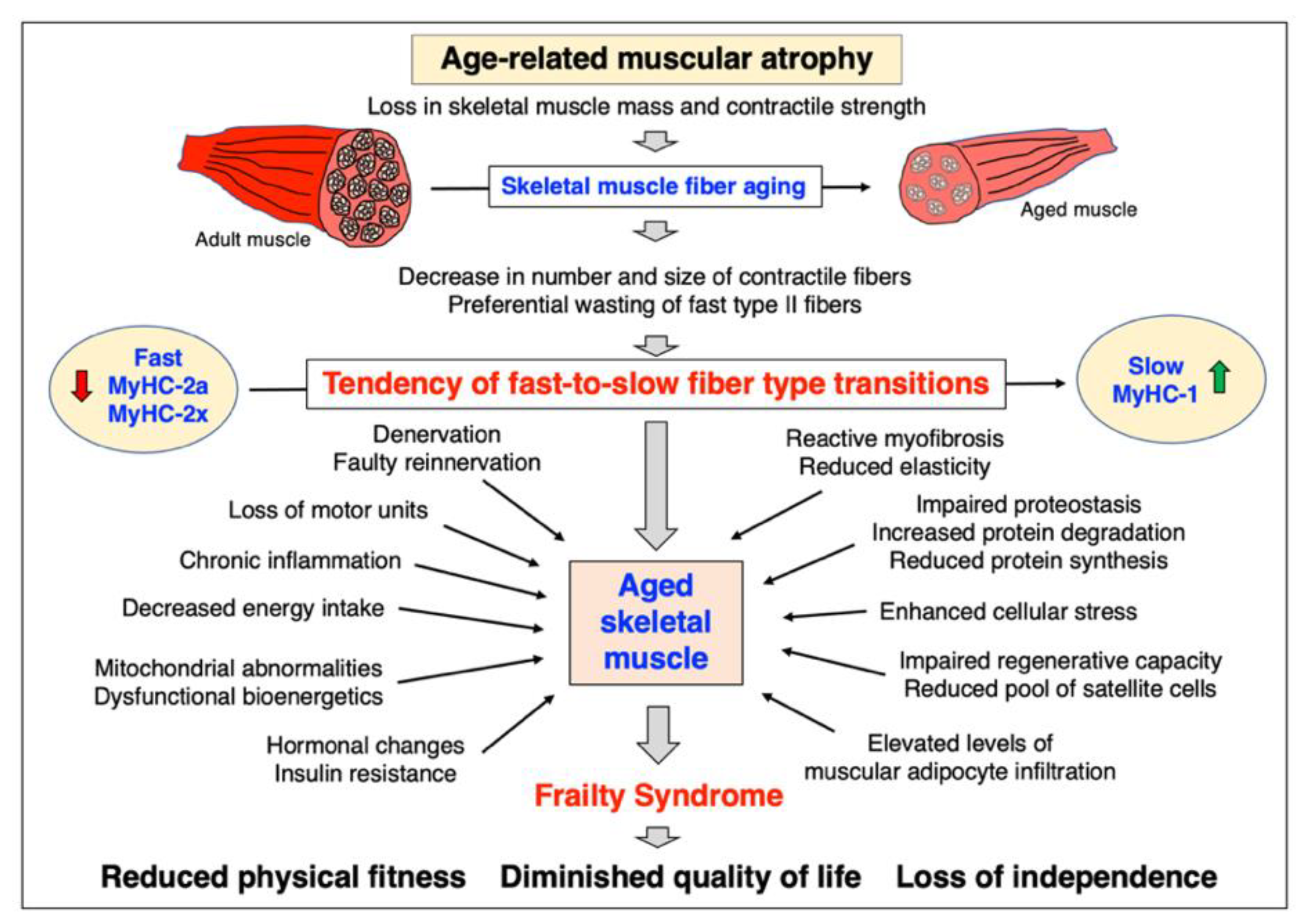{kind=link}
{kind=link}
{kind=link}
| Contractile Protein | Accession Number/Gene | Coverage/Peptides | Molecular Mass |
|---|---|---|---|
| Myosin heavy chains (MyHC) | |||
| MyHC-1, slow muscle (Myosin-7) | Q91Z83/Myh7 | 57.7/139 | 222.7 |
| MyHC-2x, fast muscle (Myosin-1) | Q5SX40/MyH1 | 69.8/192 | 223.2 |
| MyHC-2b, fast muscle (Myosin-4) | Q5SX39/MyH4 | 66.0/174 | 222.7 |
| MyHC-8, perinatal muscle (Myosin-8) | P13542/MyH8 | 46.7/124 | 222.6 |
| Myosin light chains (MLC) | |||
| MLC-1/3, skeletal muscle | P05977/Myl1 | 84.0/19 | 20.6 |
| MLC-2, skeletal muscle | P97457/Mylpf | 88.8 /19 | 18.9 |
| MLC-2, cardiac muscle | P51667/Myl2 | 75.3/12 | 18.9 |
| MLC-3, skeletal muscle | P09542/Myl3 | 78.4 /16 | 22.4 |
| Myosin-binding proteins (MYBP) | |||
| MYBP-C2, fast-type | Q5XKE0/Mybpc2 | 59.5/51 | 127.3 |
| MYBP-H | P70402/Mybph | 25.3/7 | 52.6 |
| Actin (ACT) filament | |||
| Alpha-Actin ACTA, skeletal muscle | P68134/Acta1 | 68.2/25 | 42.0 |
| F-ACT capping protein, subunit a-2 | P47754/Capza2 | 54.9/10 | 32.9 |
| F-ACT capping protein, subunit b | P47757-2/Capzab | 33.8/8 | 30.6 |
| Tropomyosin (TPM) complex | |||
| TPM, alpha-1 chain | P58771/Tpm1 | 77.8/37 | 32.7 |
| TPM, beta chain | P58774/Tpm2 | 76.1/38 | 32.8 |
| TPM, alpha-3 chain | P21107/Tpm 3 | 68.1/25 | 33.0 |
| TPM, alpha-4 chain | Q6IRU2/Tpm 4 | 37.9/9 | 28.5 |
| Troponin (TN) complex | |||
| TNI-1, slow skeletal muscle | Q9WUZ5/Tnni1 | 28.9/7 | 21.7 |
| TNI-2, fast skeletal muscle | P13412/Tnni2 | 44.0/9 | 21.3 |
| TNT-1, slow skeletal muscle | O88346-3/Tnnt1 | 28.4/8 | 30.0 |
| TNT-3, fast skeletal muscle | Q9QZ47-12/Tnnt3 | 40.2/13 | 28.3 |
| TNC-1, slow/cardiac muscle | P19123/Tnnc1 | 47.8/6 | 18.4 |
| TNC-2, skeletal muscle | P20801/Tnnc2 | 79.4/11 | 18.1 |
| Z-disc complex | |||
| Filamin FLNC | Q8VHX6-2/Flnc | 42.0/71 | 287.2 |
| Alpha-Actinin ACTN-2 | Q9JI91/Actn2 | 68.3/50 | 103.8 |
| Alpha-Actinin ACTN-3 | O88990/Actn3 | 65.2/48 | 103.0 |
| Telethonin TCAP | O70548/Tcap | 36.5/5 | 19.1 |
| Myozenin MYOZ-1 | Q9JK37/Myoz1 | 53.7/7 | 31.4 |
| Myozenin MYOZ-2 | Q9JJW5/Myoz2 | 58.3/12 | 29.7 |
| Myozenin MYOZ-3 | Q8R4E4/Myoz3 | 29.4/5 | 27.0 |
| M-line complex | |||
| Myomesin MYOM-1 | Q62234-2/Myom1 | 57.4/70 | 175.3 |
| Myomesin MYOM-3 | A2ABU4/Myom3 | 52.2/47 | 161.7 |
| Obscurin OBSCN | A2AAJ9/Obscn | 31.5/135 | 965.8 |
| Half-sarcomere-spanning titin filament | |||
| Titin TNN | A2ASS6/Ttn | 51.4/1284 | 3904.1 |
| Muscle ankyrin repeat protein MARP | Q9WV06/Ankrd2 | 26.2/7 | 36.7 |
| Specimens | Bioanalytical Approach | Proteomic Changes | References |
|---|---|---|---|
| Vastus lateralis (20–25 years versus 70–76 years) | 2D-DIGE, ESI-MS/MS, Pro-Q Diamond, PAGE analysis of MyHC isoforms | Increase in MLC-2s, ACTC and MyHC-I; decrease in MLC2f, TNT-3, TPM-3 and MyHC-2x; shift in phosphorylated MLC-2f to MLC-2s isoforms | Gelfi et al. [401] |
| Vastus lateralis (47–62 years versus 76–82 years) | 2D-DIGE, MALDI-TOF, IB | Increase in ACTC; decrease in ACTA, MLC-2, TNT-1 and TNC-1 | Staunton et al. [402], Ohlendieck [403] |
| Vastus lateralis (53 years mean age versus 78 years mean age) | Soluble proteins, LC-MS/MS, IB | Increase in MARP/ANKRD2; decreases in MLC-1/3, MyHC-2x and TTN | Théron et al. [202] |
| Vastus lateralis (48–61 years versus 76–82 years post-menopausal women) | 2D-GE (CBB), LC-MS/MS, IB | Increase in MARP/ANKRD2, MLC-1/3f, ACTA, TNT-3 and MYOZ-1; decreases in MLC2s and TNN | Gueugneau et al. [404] |
| Rectus abdominis (0–12 years versus 52–76 years) | Oxi-proteome analysis, 2D-GE, protein carbonyl immuno detection | Detection of age-related carbonylation of MyHC-1, MYBP-C1 and TNT-1 | Dos Santos et al. [405] |
| Vastus lateralis (18–30 years versus >55 years; trained and untrained) | LC-MS/MS, SRM, PAGE analysis of MyHC isoforms | Increase in MyHC-1; decrease in MyHC-2a; establishment of quantitative differences in myosin light chain composition | Cobley et al. [406] |
| Vastus lateralis (22–27 years versus 65–75 years) | Single-muscle-fiber proteomics, LC-MS/MS | Differential effects on fast versus slow fibers based on MyHC-1, MyHC-2a and MyHC-2x distribution analysis; increase in chaperones of MyHC and ACTA | Murgia et al. [407] |
| Quadriceps muscle (66–80 years) of healthy versus cancer patients | LC-ESI-MS/MS, SWATH MS, IFM, IB | Differential expression of MyHC-1, MyHC-2a and MyHC-2x in healthy elderly versus cancer patients with or without weight loss | Ebhardt et al. [408] |
| Vastus lateralis (23 years mean age versus 71 years mean age) | 2D-GE (CBB), Pro-Q Diamond, MALDI-TOF MS, PAGE analysis of MyHC isoforms, IB | Increase in MyHC-1; decrease in MyHC-2a and MyHC-2x; myosin/actin ratio not affected; differential effects on expression of TNT-3, ACTA and ACTC proteoforms | Brocca et al. [409] |
| Vastus lateralis (Obese and healthy older men of average age 66 undergoing resistance training and energy restriction) | LC-MS/MS, deuterated water labeling of newly synthesized skeletal muscle proteins | Determination of synthesis rate of myofibrillar proteins (MyHC, MLC, ACTA, TPM, TNC, TNT, TNI) | Murphy et al. [221] |
| Vastus lateralis (range of individuals from 20 to 87 years of age) | TMT, LC-MS/MS | Decrease in MYBP-H; switch from MyHC-2x/MyHC-2a to MyHC-1; differential effects on TNT-3, TPM-1 and MYOZ-2 expression | Ubaida-Mohien et al. [410,411] |
| Vastus lateralis (25 years mean age versus 62 years mean age) | LC-MS/MS, PAGE analysis of MyHC isoforms | Reduced acto-myosin abundance; decrease in ACTA and MYBP-H; increase in ACTC and TNT-1 | Vann et al. [412] |
| Vastus lateralis (21 years mean age versus 73 years mean age) | 2D-GE (CBB), LC-MS/MS, IB | Increase in TNT-1 and MARP; decrease in ACTA, TNT-3 and MYOZ-1 | Gueugneau et al. [413] |
| Vastus lateralis (25 years mean age versus 67 years mean age) | iTRAQ, LC-MS/MS | Decrease in ACTA and FLNC | Deane et al. [414] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dowling, P.; Gargan, S.; Swandulla, D.; Ohlendieck, K. Fiber-Type Shifting in Sarcopenia of Old Age: Proteomic Profiling of the Contractile Apparatus of Skeletal Muscles. Int. J. Mol. Sci. 2023, 24, 2415. https://doi.org/10.3390/ijms24032415
Dowling P, Gargan S, Swandulla D, Ohlendieck K. Fiber-Type Shifting in Sarcopenia of Old Age: Proteomic Profiling of the Contractile Apparatus of Skeletal Muscles. International Journal of Molecular Sciences. 2023; 24(3):2415. https://doi.org/10.3390/ijms24032415
Chicago/Turabian StyleDowling, Paul, Stephen Gargan, Dieter Swandulla, and Kay Ohlendieck. 2023. "Fiber-Type Shifting in Sarcopenia of Old Age: Proteomic Profiling of the Contractile Apparatus of Skeletal Muscles" International Journal of Molecular Sciences 24, no. 3: 2415. https://doi.org/10.3390/ijms24032415
APA StyleDowling, P., Gargan, S., Swandulla, D., & Ohlendieck, K. (2023). Fiber-Type Shifting in Sarcopenia of Old Age: Proteomic Profiling of the Contractile Apparatus of Skeletal Muscles. International Journal of Molecular Sciences, 24(3), 2415. https://doi.org/10.3390/ijms24032415