
Moderating Gut Microbiome/Mitochondrial Axis in Oxazolone Induced Ulcerative Colitis: The Evolving Role of β-Glucan and/or, Aldose Reductase Inhibitor, Fidarestat
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of Oxazolone, β-Glucan and Fidarestat on Mitochondrial Parameters
2.2. β-Glucan and Fidarestat Ameliorated Oxazolone-Induced Intestinal Oxidative Stress and Inflammation
2.3. Effect of Oxazolone, β-Glucan and Fidarestat on Intestinal Nrf2 DNA Binding Activity and Peroxisome Proliferator-activated Receptor γ Co-activator 1α (PGC1-α) Levels
2.4. Effect of Oxazolone, β-Glucan and Fidarestat on Microbiota-Derived Metabolites
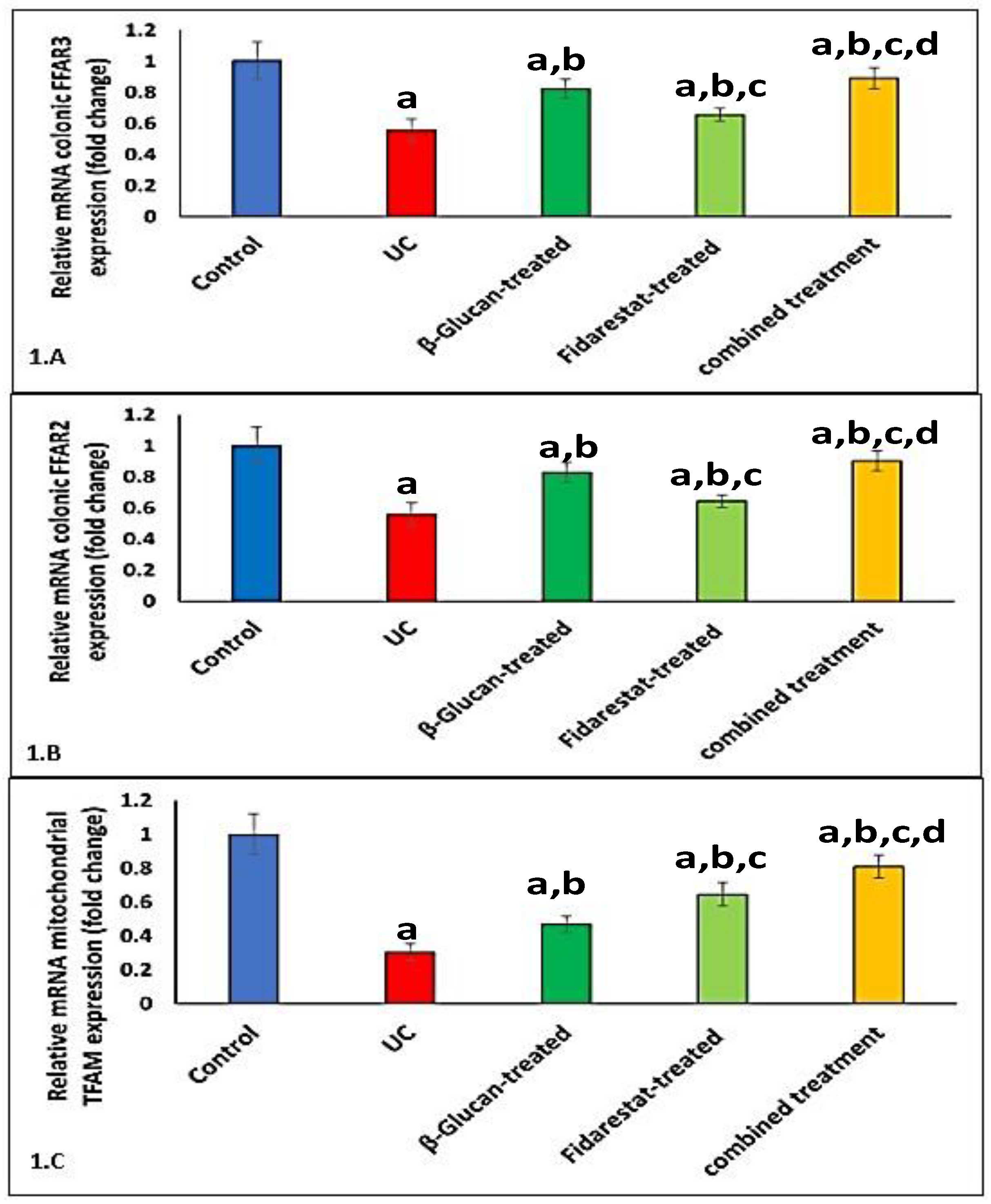
2.5. Effect of Oxazolone, β-Glucan and Fidarestat on mRNA Expressions of Colonic FFAR2, Colonic FFAR3, and Mitochondrial Transcription Factor A (TFAM)
2.6. Correlation between TMAO, SCFAs, FFAR2, FFAR3, TFAM, and Measured Variables in UC Group
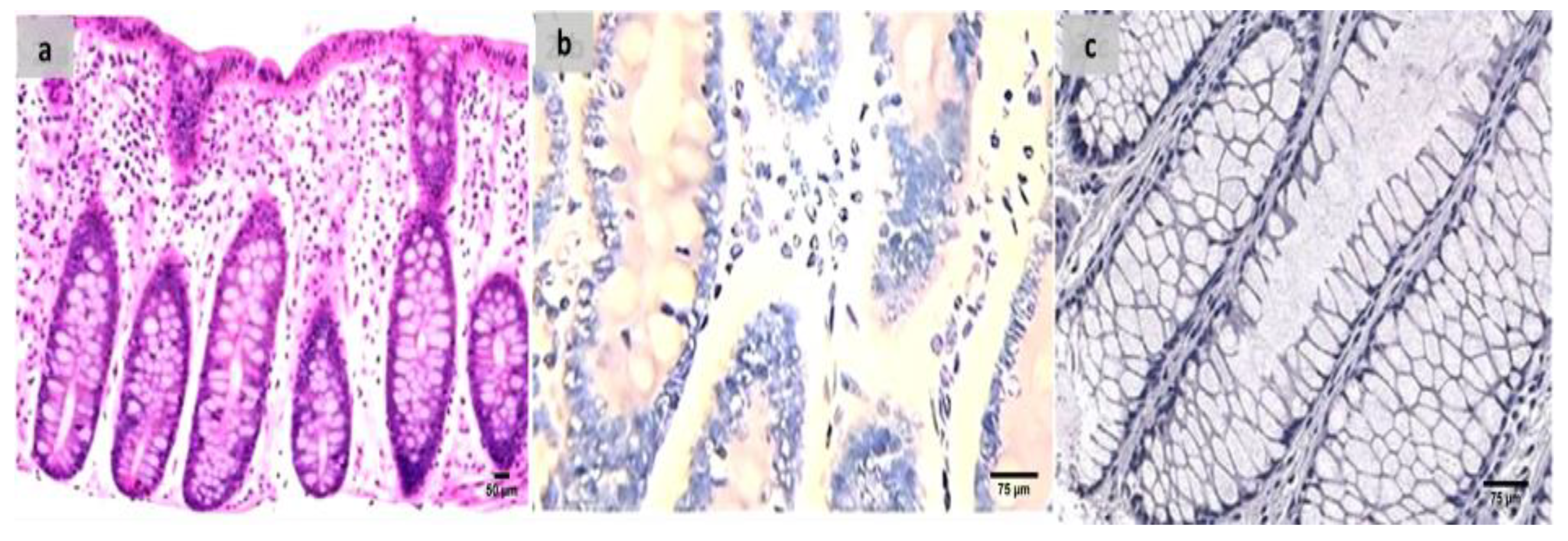
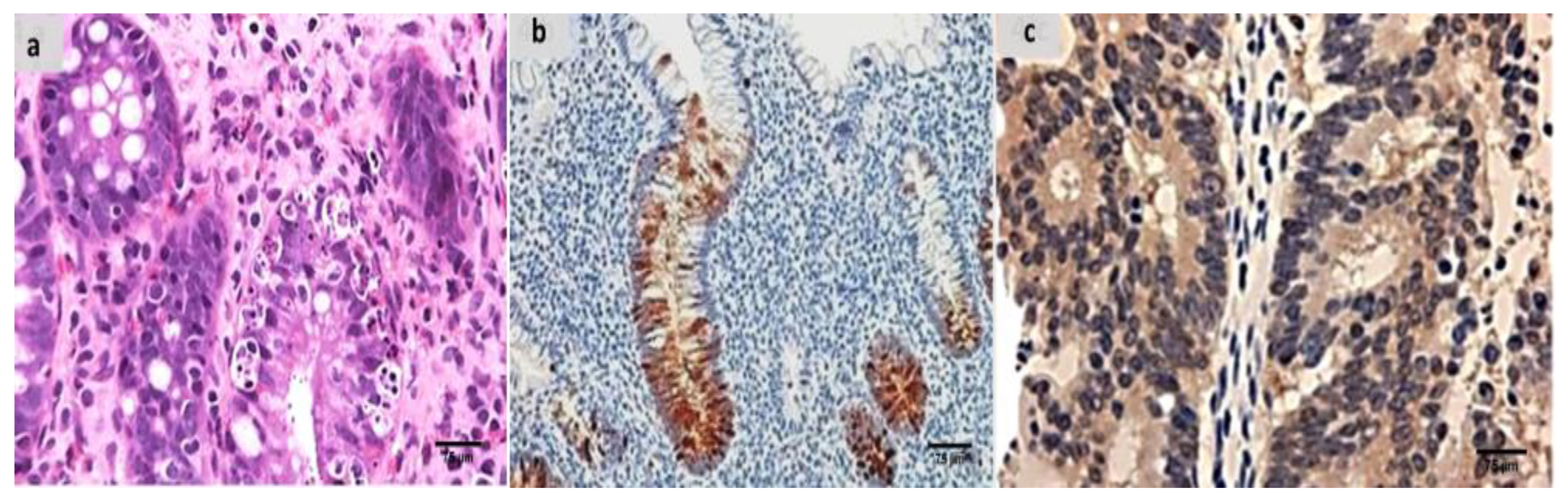
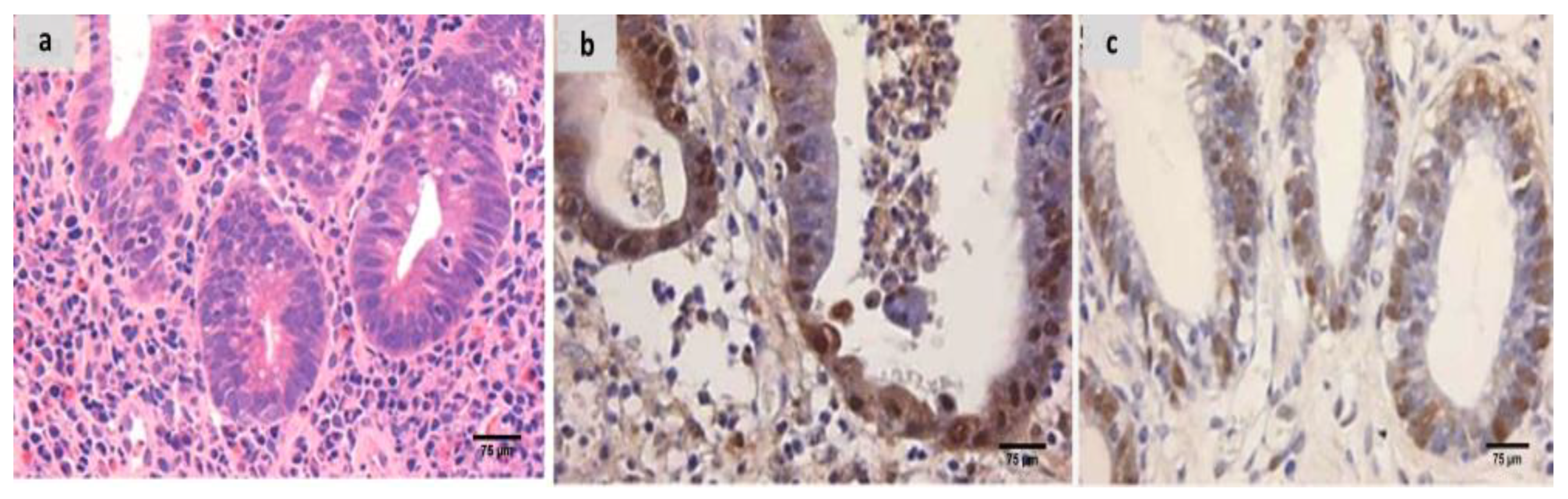
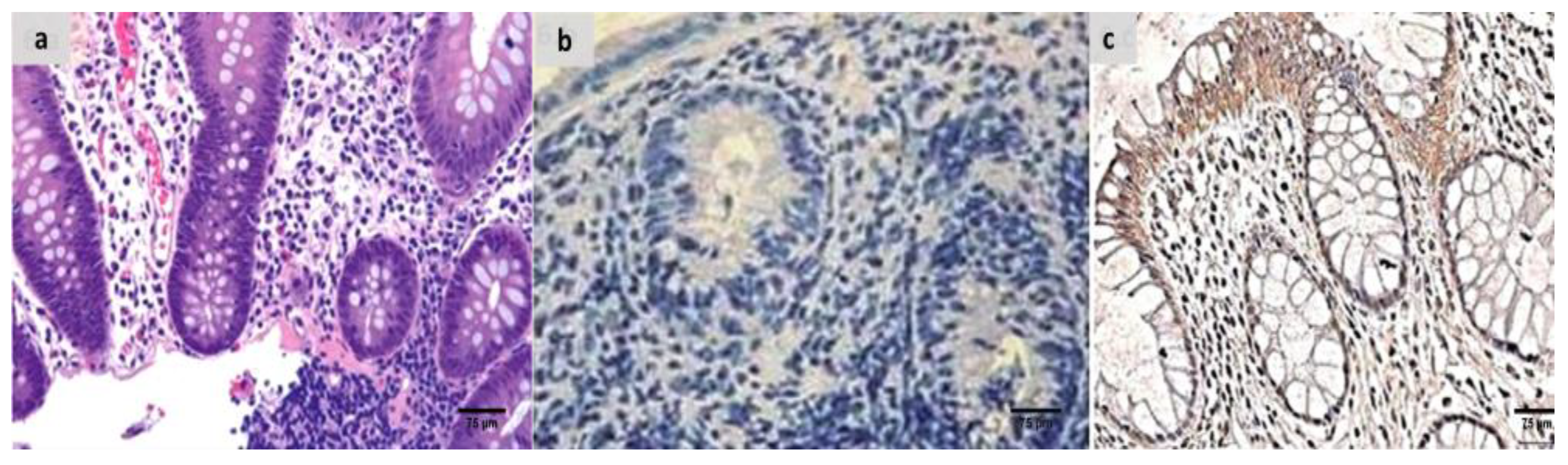
2.7. The Groups’ Histopathology and Immunohistochemistry Findings
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Experimental Design
The Rats Were Divided into Five Equal Groups, Each with Ten Rats
- Control group: received 1.1 mL/rat of 40% ethanol solvent once via a rubber catheter while receiving mild ether anesthesia.
- UC group: was shown the non-treated group with UC caused by oxazolone.
- β-Glucan group: for 28 days, oxazolone-induced UC was combined with intragastric gavage administration of β-glucan at a dosage of 350 mg/kg/day (at a value of 140 mg/mL in filtered water) [58].
- Fidarestat group: received fidarestat by intragastric gavage for 28 days at a dosage of 4 mg/kg at a 1 mg/mL concentration in DMSO after oxazolone-induced UC [59].
- Combined treatment group: UC caused by oxazolone with concomitant administration of fidarestat and β-glucan at similar doses and time frames previously described.
4.4. Tissue Collection and Homogenate Creation
4.5. Mitochondrial Parameters
4.5.1. Mitochondrial ATP Concentration
4.5.2. Mitochondrial Transmembrane Potential (ΔΨm) Assay
4.5.3. Mitochondria ROS Assay
4.5.4. Nrf2 DNA Binding Activity
4.5.5. PGC1α and NF-kꞵ
4.6. Redox-Status Parameters
4.6.1. GSH
4.6.2. SOD
4.7. Microbiota-Derived Metabolite Detection
4.7.1. The Level of Trimethylamine N-Oxide (TMAO)
4.7.2. Determination of SCFA in Fecal Samples
4.8. Numerical Estimation of Colonic (FFAR-2 and FFAR-3 in the Colon and Mitochondrial TFAM mRNA Expression by Numerical Real-Time Reverse Transcription PCR (RT–PCR)
4.9. Histopathological Processing and Group Assessment
4.10. Immunohistochemical Analysis and Assessment of the Groups Examined
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mottawea, W.; Chiang, C.K.; Mühlbauer, M.; Starr, A.E.; Butcher, J.; Abujamel, T.; Deeke, S.A.; Brandel, A.; Zhou, H.; Shokralla, S.; et al. Altered intestinal microbiota–host mitochondria crosstalk in new onset Crohn’s disease. Nat. Commun. 2016, 7, 13419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Georges-Chaumet, Y.; Edeas, M. Microbiota–mitochondria inter-talk: Consequence for microbiota–host interaction. Pathog. Dis. 2016, 74, ftv096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.; Mach, N. The crosstalk between the gut microbiota and mitochondria during exercise. Front. Physiol. 2017, 8, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Circu, M.L.; Aw, T.Y. Intestinal redox biology and oxidative stress. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2012; Volume 23, pp. 729–737. [Google Scholar]
- Bär, F.; Bochmann, W.; Widok, A.; Von Medem, K.; Pagel, R.; Hirose, M.; Yu, X.; Kalies, K.; König, P.; Böhm, R.; et al. Mitochondrial gene polymorphisms that protect mice from colitis. Gastroenterology 2013, 145, 1055–1063. [Google Scholar] [CrossRef]
- Green, D.R.; Galluzzi, L.; Kroemer, G. Mitochondria and the autophagy–inflammation–cell death axis in organismal aging. Science 2011, 333, 1109–1112. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, K.E.; Vincent, G.; Sodhi, C.P.; Novak, E.A.; Ranganathan, S.; Egan, C.E.; Stolz, D.B.; Rogers, M.B.; Firek, B.; Morowitz, M.J.; et al. Peroxisome proliferator-activated receptor-γ coactivator 1-α (PGC1α) protects against experimental murine colitis. J. Biol. Chem. 2016, 291, 10184–10200. [Google Scholar] [CrossRef] [Green Version]
- Veltkamp, C.; Tonkonogy, S.L.; De Jong, Y.P.; Albright, C.; Grenther, W.B.; Balish, E.; Terhorst, C.; Sartor, R.B. Continuous stimulation by normal luminal bacteria is essential for the development and perpetuation of colitis in Tgϵ26 mice. Gastroenterology 2001, 120, 900–913. [Google Scholar] [CrossRef]
- Ohkusa, T.; Okayasu, I.; Ogihara, T.; Morita, K.; Ogawa, M.; Sato, N. Induction of experimental ulcerative colitis by Fusobacterium varium isolated from colonic mucosa of patients with ulcerative colitis. Gut 2003, 52, 79–83. [Google Scholar] [CrossRef]
- Schaubeck, M.; Clavel, T.; Calasan, J.; Lagkouvardos, I.; Haange, S.B.; Jehmlich, N.; Basic, M.; Dupont, A.; Hornef, M.; Von Bergen, M.; et al. Dysbiotic gut microbiota causes transmissible Crohn’s disease-like ileitis independent of failure in antimicrobial defence. Gut 2016, 65, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhang, Y.; Yang, S.; Zhao, D.; Wang, M. A polysaccharide from cultured mycelium of Hericium erinaceus relieves ulcerative colitis by counteracting oxidative stress and improving mitochondrial function. Int. J. Biol. Macromol. 2019, 125, 572–579. [Google Scholar] [CrossRef]
- Hirose, M.; Künstner, A.; Schilf, P.; Sünderhauf, A.; Rupp, J.; Jöhren, O.; Schwaninger, M.; Sina, C.; Baines, J.F.; Ibrahim, S.M. Mitochondrial gene polymorphism is associated with gut microbial communities in mice. Sci. Rep. 2017, 7, 15293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashir, K.M.I.; Choi, J.S. Clinical and Physiological Perspectives of β-Glucans: The Past, Present, and Future. Int. J. Mol. Sci. 2017, 18, 1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchecka, D.; Harasym, J.P.; Wilczak, J.; Gajewska, M.; Oczkowski, M.; Gudej, S.; Błaszczyk, K.; Kamola, D.; Filip, R.; Gromadzka-Ostrowska, J. Antioxidative and anti-inflammatory effects of high beta-glucan concentration purified aqueous extract from oat in experimental model of LPS-induced chronic enteritis. J. Funct. Foods 2015, 14, 244–254. [Google Scholar] [CrossRef]
- Wilczak, J.; Błaszczyk, K.; Kamola, D.; Gajewska, M.; Harasym, J.P.; Jałosińska, M.; Gudej, S.; Suchecka, D.; Oczkowski, M.; Gromadzka-Ostrowska, J. The effect of low or high molecular weight oat beta-glucans on the inflammatory and oxidative stress status in the colon of rats with LPS-induced enteritis. Food Funct. 2015, 6, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Cani, P.D. A place for dietary fibre in the management of the metabolic syndrome. Curr. Opin. Clin. Nutr. Metab. Care 2005, 8, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; De Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104, S1–S63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawanishi, K.; Ueda, H.; Moriyasu, M. Aldose reductase inhibitors from the nature. Curr. Med. Chem. 2003, 10, 1353–1374. [Google Scholar] [CrossRef]
- Singh Grewal, A.; Bhardwaj, S.; Pandita, D.; Lather, V.; Singh Sekhon, B. Updates on aldose reductase inhibitors for management of diabetic complications and non-diabetic diseases. Mini Rev. Med. Chem. 2016, 16, 120–162. [Google Scholar] [CrossRef]
- Shukla, K.; Sonowal, H.; Saxena, A.; Ramana, K.V.; Srivastava, S.K. Aldose reductase inhibitor, fidarestat regulates mitochondrial biogenesis via Nrf2/HO-1/AMPK pathway in colon cancer cells. Cancer Lett. 2017, 411, 57–63. [Google Scholar] [CrossRef]
- Jackson, D.N.; Theiss, A.L. Gut bacteria signaling to mitochondria in intestinal inflammation and cancer. Gut Microbes 2020, 11, 285–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgart, D.C.; Sandborn, W.J. Inflammatory bowel disease: Clinical aspects and established and evolving therapies. Lancet 2007, 369, 1641–1657. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Lay, C.; Seksik, P.; Tannock, G.W. Analysis of bacterial bowel communities of IBD patients: What has it revealed? Inflamm. Bowel Dis. 2008, 14, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Hamm, C.M.; Gulati, A.S.; Sartor, R.B.; Chen, H.; Wu, X.; Zhang, T.; Rohlf, F.J.; Zhu, W.; Gu, C.; et al. Inflammatory bowel diseases phenotype, C. difficile and NOD2 genotype are associated with shifts in human ileum associated microbial composition. PLoS ONE 2012, 7, e26284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.; Teft, W.A.; Morse, B.L.; Choi, Y.H.; Woolsey, S.; DeGorter, M.K.; Hegele, R.A.; Tirona, R.G.; Kim, R.B. Trimethylamine-N-oxide: A novel biomarker for the identification of inflammatory bowel disease. Dig. Dis. Sci. 2015, 60, 3620–3630. [Google Scholar] [CrossRef]
- Sun, X.; Jiao, X.; Ma, Y.; Liu, Y.; Zhang, L.; He, Y.; Chen, Y. Trimethylamine N-oxide induces inflammation and endothelial dysfunction in human umbilical vein endothelial cells via activating ROS-TXNIP-NLRP3 inflammasome. Biochem. Biophys. Res Commun. 2016, 481, 63–70. [Google Scholar] [CrossRef]
- Mueller, A.; Raptis, J.; Rice, P.J.; Kalbfleisch, J.H.; Stout, R.D.; Ensley, H.E.; Browder, W.; Williams, D.L. The influence of glucan polymer structure and solution conformation on binding to (1→3)-β-d-glucan receptors in a human monocyte-like cell line. Glycobiology 2000, 10, 339–346. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Xia, Q.; Liu, L.; Wu, Z.; Pan, D. Recent advances of cereal β-glucan on immunity with gut microbiota regulation functions and its intelligent gelling application. Crit. Rev. Food Sci. Nutr. 2021, 8, 1–17. [Google Scholar] [CrossRef]
- Żyła, E.; Dziendzikowska, K.; Gajewska, M.; Wilczak, J.; Harasym, J.; Gromadzka-Ostrowska, J. Beneficial effects of oat beta-glucan dietary supplementation in colitis depend on its molecular weight. Molecules 2019, 24, 3591. [Google Scholar] [CrossRef] [Green Version]
- Moos, W.H.; Faller, D.V.; Harpp, D.N.; Kanara, I.; Pernokas, J.; Powers, W.R.; Steliou, K. Microbiota and neurological disorders: A gut feeling. BioRes. Open Access 2016, 5, 137–145. [Google Scholar] [CrossRef]
- Li, Q.; Chen, H.; Zhang, M.; Wu, T.; Liu, R. Altered short chain fatty acid profiles induced by dietary fiber intervention regulate AMPK levels and intestinal homeostasis. Food Funct. 2019, 10, 7174–7187. [Google Scholar] [CrossRef] [PubMed]
- Lymperopoulos, A.; Suster, M.S.; Borges, J.I. Short-Chain Fatty Acid Receptors and Cardiovascular Function. Int. J. Mol. Sci. 2022, 23, 3303. [Google Scholar] [CrossRef]
- Priyadarshini, M.; Villa, S.R.; Fuller, M.; Wicksteed, B.; Mackay, C.R.; Alquier, T.; Poitout, V.; Mancebo, H.; Mirmira, R.G.; Gilchrist, A.; et al. An acetate-specific GPCR, FFAR2, regulates insulin secretion. Mol. Endocrinol. 2015, 29, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindels, L.B.; Porporato, P.E.; Ducastel, S.; Sboarina, M.; Neyrinck, A.M.; Dewulf, E.M.; Feron, O.; Lestavel, S.; Cani, P.D.; Staels, B.; et al. Ffar2 expression regulates leukaemic cell growth in vivo. Br. J. Cancer 2017, 117, 1336–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masui, R.; Sasaki, M.; Funaki, Y.; Ogasawara, N.; Mizuno, M.; Iida, A.; Izawa, S.; Kondo, Y.; Ito, Y.; Tamura, Y.; et al. G protein-coupled receptor 43 moderates gut inflammation through cytokine regulation from mononuclear cells. Inflamm. Bowel Dis. 2013, 19, 2848–2856. [Google Scholar] [CrossRef]
- Zhou, M.; Pu, C.; Xia, L.; Yu, X.; Zhu, B.; Cheng, R.; Xu, L.; Zhang, J. Salecan diet increases short chain fatty acids and enriches beneficial microbiota in the mouse cecum. Carbohydr. Polym. 2014, 102, 772–779. [Google Scholar] [CrossRef]
- Dong, L.; Qin, C.; Li, Y.; Wu, Z.; Liu, L. Oat phenolic compounds regulate metabolic syndrome in high fat diet-fed mice via gut microbiota. Food Biosci. 2022, 50, 101946. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Ren, X.; Zhang, X.; Wu, Z.; Liu, L. The positive correlation of antioxidant activity and prebiotic effect about oat phenolic compounds. Food Chem. 2023, 402, 134231. [Google Scholar] [CrossRef]
- Li, Y.; Qin, C.; Dong, L.; Zhang, X.; Wu, Z.; Liu, L.; Yang, J.; Liu, L. Whole grain benefit: Synergistic effect of oat phenolic compounds and β-glucan on hyperlipidemia via gut microbiota in high-fat-diet mice. Food Funct. 2022, 13, 12686–12696. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Srivastava, S.K.; Ramana, K.V. A potential therapeutic role for aldose reductase inhibitors in the treatment of endotoxin-related inflammatory diseases. Expert Opin. Investig. Drugs 2012, 21, 329–339. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive oxygen species in metabolic and inflammatory signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Mizuno, K.; Makino, M.; Suzuki, T.; Yagihashi, S. Effects of 15-month aldose reductase inhibition with fidarestat on the experimental diabetic neuropathy in rats. Diabetes Res. Clin. Pract. 2000, 50, 77–85. [Google Scholar] [CrossRef]
- Obrosova, I.G. Anticataract effect as a predictor of drug efficacy against early diabetes-induced retinal changes. Acta Ophthalmol. 2009, 87. [Google Scholar] [CrossRef]
- Obrosova, I.G. How does glucose generate oxidative stress in peripheral nerve? Int. Rev. Neurobiol. 2002, 50, 3–5. [Google Scholar]
- Yao, K.; Zhang, W.W.; Yao, L.; Yang, S.; Nie, W.; Huang, F. Carvedilol promotes mitochondrial biogenesis by regulating the PGC-1/TFAM pathway in human umbilical vein endothelial cells (HUVECs). Biochem. Biophys. Res. Commun. 2016, 470, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Dierckx, A.; Wanrooij, P.H.; Wanrooij, S.; Larsson, N.G.; Wilhelmsson, L.M.; Falkenberg, M.; Gustafsson, C.M. Mammalian transcription factor A is a core component of the mitochondrial transcription machinery. Proc. Natl Acad Sci. USA 2012, 109, 16510–16515. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Shen, J.; Ran, Z. Emerging views of mitophagy in immunity and autoimmune diseases. Autophagy 2020, 16, 3–17. [Google Scholar] [CrossRef]
- Reifen, R.; Levy, E.; Berkovich, Z.; Tirosh, O. Vitamin A exerts its antiinflammatory activities in colitis through preservation of mitochondrial activity. Nutrition 2015, 31, 1402–1407. [Google Scholar] [CrossRef]
- Sonowal, H.; Pal, P.; Shukla, K.; Saxena, A.; Srivastava, S.K.; Ramana, K.V. Aldose reductase inhibitor, fidarestat prevents doxorubicin-induced endothelial cell death and dysfunction. Biochem. Pharmacol. 2018, 150, 181–190. [Google Scholar] [CrossRef]
- Tang, W.H.; Stitham, J.; Jin, Y.; Liu, R.; Lee, S.H.; Du, J.; Atteya, G.; Gleim, S.; Spollett, G.; Martin, K.; et al. Aldose reductase–mediated phosphorylation of p53 leads to mitochondrial dysfunction and damage in diabetic platelets. Circulation 2014, 129, 1598–1609. [Google Scholar] [CrossRef] [Green Version]
- Li, C.X.; Wang, H.W.; Jiang, W.J.; Li, G.C.; Zhang, Y.D.; Luo, C.H.; Li, X.C. The Inhibition of aldose reductase accelerates liver regeneration through regulating energy metabolism. Oxid. Med. Cell. Longev. 2020, 2020, 3076131. [Google Scholar] [CrossRef]
- Ananthakrishnan, R.; Kaneko, M.; Hwang, Y.C.; Quadri, N.; Gomez, T.; Li, Q.; Caspersen, C.; Ramasamy, R. Aldose reductase mediates myocardial ischemia-reperfusion injury in part by opening mitochondrial permeability transition pore. Am. J. Physiol.-Heart Circ. Physiol. 2009, 296, H333–H341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.R.; Wang, Z.; Zhou, W.; Fan, S.R.; Ma, R.; Xue, L.; Yang, L.; Li, Y.S.; Tan, H.L.; Shao, Q.H.; et al. Epalrestat protects against diabetic peripheral neuropathy by alleviating oxidative stress and inhibiting polyol pathway. Neural Regen. Res. 2016, 11, 345. [Google Scholar] [PubMed]
- Wang, X.; Yu, F.; Zheng, W.Q. Aldose reductase inhibitor Epalrestat alleviates high glucose-induced cardiomyocyte apoptosis via ROS. Eur. Rev. Med. Pharmacol. Sci. 2019, 23 (Suppl. 3), 294–303. [Google Scholar]
- Kim, J.H.; Hahm, D.H.; Yang, D.C.; Kim, J.H.; Lee, H.J.; Shim, I. Effect of crude saponin of Korean Red Ginseng on high-fat diet-induced obesity in the rat. J. Pharmacol. Sci. 2005, 97, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boirivant, M.; Fuss, I.J.; Chu, A.; Strober, W. Oxazolone colitis: A murine model of T helper cell type 2 colitis treatable with antibodies to interleukin 4. J. Exp. Med. 1998, 188, 1929–1939. [Google Scholar] [CrossRef] [Green Version]
- Shen, R.L.; Dang, X.Y.; Dong, J.L.; Hu, X.Z. Effects of oat β-glucan and barley β-glucan on fecal characteristics, intestinal microflora, and intestinal bacterial metabolites in rats. J. Agric. Food Chem. 2012, 60, 11301–11308. [Google Scholar] [CrossRef]
- Hattori, T.; Matsubara, A.; Taniguchi, K.; Ogura, Y. Aldose reductase inhibitor fidarestat attenuates leukocyte-endothelial interactions in experimental diabetic rat retina in vivo. Curr. Eye Res. 2010, 35, 146–154. [Google Scholar] [CrossRef]
- Olivera, A.A.; Meigs, R.A. Mitochondria from human term placenta. I. Isolation and assay conditions for oxidative phosphorylation. Biochim. Biophys. Acta (BBA)-Bioenerget. 1975, 376, 426–435. [Google Scholar] [CrossRef]
- Lowry, O.H. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Maity, P.; Bindu, S.; Dey, S.; Goyal, M.; Alam, A.; Pal, C.; Mitra, K.; Bandyopadhyay, U. Indomethacin, a non-steroidal anti-inflammatory drug, develops gastropathy by inducing reactive oxygen species-mediated mitochondrial pathology and associated apoptosis in gastric mucosa: A novel role of mitochondrial aconitase oxidation. J. Biol. Chem. 2009, 284, 3058–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Jin, R.; Hao, J.; Zeng, J.; Yin, D.; Yi, Y.; Zhu, M.; Mandal, A.; Hua, Y.; Ng, C.K.; et al. Consumption of the Fish Oil High-Fat Diet Uncouples Obesity and Mammary Tumor Growth through Induction of Reactive Oxygen Species in Protumor Macrophages. Cancer Res. 2020, 80, 2564–2574. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar] [PubMed]
- Wekell, J.C.; Barnett, H. New method for analysis of trimethylamine oxide using ferrous sulfate and EDTA. J. Food Sci. 1991, 56, 132–135. [Google Scholar] [CrossRef]
- Zhang, Q.; Fan, X.Y.; Guo, W.L.; Cao, Y.J.; Lin, Y.C.; Cheng, W.J.; Chen, L.J.; Rao, P.F.; Ni, L.; Lv, X.C. The protective mechanisms of macroalgae Laminaria japonica consumption against lipid metabolism disorders in high-fat diet-induced hyperlipidemic rats. Food Funct. 2020, 11, 3256–3270. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xu, G.; Sun, Y.; He, H.; Xue, Q.; Liu, Y.; Dong, L. Effect of TrkB-PLC/IP3 pathway on intestinal inflammatory factors and enterocyte apoptosis in mice with colitis. Acta Biochim. Biophys. Sin. 2020, 52, 675–682. [Google Scholar] [CrossRef]
- Zhu, L.; Dai, L.M.; Shen, H.; Gu, P.Q.; Zheng, K.; Liu, Y.J.; Zhang, L.; Cheng, J.F. Qing Chang Hua Shi granule ameliorate inflammation in experimental rats and cell model of ulcerative colitis through MEK/ERK signaling pathway. Biomed. Pharmacother. 2019, 116, 108967. [Google Scholar] [CrossRef]
- Krisnuhoni, E.; Handjari, D.R.; Stephanie, M.; Kencana, L.; Rahadiani, N. Intramucosal Calprotectin Expression in Inflammatory Bowel Disease (IBD) and Non-IBD Colorectal Inflammation. Open Access Maced. J. Med. Sci. 2022, 10, 872–878. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group I | Group II | Group III | Group IV | Group V | |
|---|---|---|---|---|---|
| ATP conc. (nmol/mg protein) | 13.90 ± 0.91 | 4.96 ± 0.94 a | 8.16 ± 0.72 a,b | 8.96 ± 0.80 a,b,c | 10.89 ± 0.98 a,b,c,d |
| Transmembrane potential (ΔΨm) (florescent unit) | 8.48 ± 0.64 | 2.37 ± 0.68 a | 4.46 ± 0.88 a,b | 5.63 ± 0.58 a,b,c | 7.82 ± 0.63 a,b,c,d |
| ROS (pmol/min/mg protein) | 5.42 ± 0.64 | 23.10 ± 2.18 a | 13.74 ± 1.24 a,b | 13.44 ± 1.33 a,b | 8.37 ± 0.93 a,b,c,d |
| Group I | Group II | Group III | Group IV | Group V | |
|---|---|---|---|---|---|
| GSH (μg/mg protein) | 7.99 ± 0.63 | 1.53 ± 0.31 a | 4.39 ± 0.50 a,b | 4.46 ± 0.55 a,b | 6.97 ± 0.52 a,b,c,d |
| SOD (U/gm tissue) | 405.96 ± 11.74 | 119.94 ± 18.45 a | 258.35 ± 28.71 a,b | 259.34 ± 32.64 a,b | 350.86 ± 35.06 a,b,c,d |
| NF-ĸB (ng/mg.tissue protein) | 0.18 ± 0.04 | 0.64 ± 0.05 a | 0.37 ± 0.05 a,b | 0.36 ± 0.07 a,b | 0.24 ± 0.04 a,b,c,d |
| Group I | Group II | Group III | Group IV | Group V | |
|---|---|---|---|---|---|
| Nrf2 DNA binding activity (μg/mg protein) | 2.06 ± 0.16 | 0.72 ± 0.15 a | 1.42 ± 0.21 a,b | 1.42 ± 0.17 a,b | 1.81 ± 0.20 a,b,c,d |
| PGC1-α (ng/mg protein) | 0.95 ± 0.11 | 0.30 ± 0.06 a | 0.56 ± 0.07 a,b | 0.58 ± 0.06 a,b | 0.88 ± 0.10 b,c,d |
| Group I | Group II | Group III | Group IV | Group V | |
|---|---|---|---|---|---|
| TMAO (pmol/mg protein) | 16.99 ± 3.15 | 97.96 ± 7.53 a | 65.12 ± 5.98 a,b | 70.63 ± 8.66 a,b | 35.39 ± 5.08 a,b,c,d |
| SCFAs (µmol/g) | 7.57 ± 0.19 | 1.48 ± 0.20 a | 4.26 ± 0.32 a,b | 4.33 ± 0.51 a,b | 6.00 ± 0.28 a,b,c,d |
| TMAO | SCFAs | FFAR-2 | FFAR-3 | TFAM | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | r | p | |
| Mitochondrial ATP conc. (nmol/mg protein) | −0.84 * | <0.01 | 0.78 * | <0.01 | 0.83* | <0.01 | 0.87 * | <0.01 | 0.84 * | <0.01 |
| Mitochondrial transmembrane potential (ΔΨm) (florescent unit) | −0.88 * | <0.01 | 0.82 * | <0.01 | 0.87 * | <0.01 | 0.90 * | <0.01 | 0.88 * | <0.01 |
| Mitochondrial ROS (pmol/min/mg protein) | 0.81 * | <0.01 | −0.77 * | <0.01 | −0.83 * | <0.01 | −0.87 * | <0.01 | −0.81 * | 0.01 |
| GSH (μg/mg protein) | −0.98 * | <0.01 | 0.95 * | <0.01 | 0.98 * | <0.01 | 0.95 * | <0.01 | 0.94 * | <0.01 |
| SOD (U/gm tissue) | −0.80 * | <0.01 | 0.75 * | 0.01 | 0.78 * | <0.01 | 0.83 * | <0.01 | 0.82 * | <0.01 |
| NF-ĸB (ng/mg.tissue protein) | 0.84 * | <0.01 | −0.81 * | 0.01 | −0.81 * | 0.01 | −0.87 * | <0.01 | −0.85 * | <0.01 |
| Nrf2 DNA binding activity (μg/mg protein) | −0.91 * | <0.01 | 0.86 * | <0.01 | 0.90 * | <0.01 | 0.92 * | <0.01 | 0.90 * | <0.01 |
| PGC1-α (ng/mg protein) | −0.84 * | <0.01 | 0.82 * | <0.01 | 0.85 * | <0.01 | 0.91 * | <0.01 | 0.88 * | <0.01 |
| Forward | Reverse | Accession Number | |
|---|---|---|---|
| FFAR2 | 5′CTACGAGAACTT CACCCAAGAG 3 | 5′ GAAGCGCCAATAA CAGAAGATG 3′ | NM_001005877.2 |
| FFAR3 | 5′ CCGGCGCAAGAGGATAAT-3′ | 5′ CCCACCACATGGG ACATATT 3′ | NM_001108912.1 |
| TFAM | 5′AAGGGAATGGGAAAGGTAGA3 | 5′AACAGGACATGGAAAGCAGAT3′ | NM_0011045 |
| GADPH | 5′AGACAGCCGCATCTTCTTGT 3′ | 5′ CTTGCCGTGGGTAGAGTCAT 3′ | NM_017008.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Deeb, O.S.; Elesawy, R.O.; Eltokhy, A.K.; Al-Shenawy, H.A.; Ghanem, H.B.; Rizk, F.H.; Barhoma, R.A.; Shalaby, R.H.; Abdelsattar, A.M.; Mashal, S.S.; et al. Moderating Gut Microbiome/Mitochondrial Axis in Oxazolone Induced Ulcerative Colitis: The Evolving Role of β-Glucan and/or, Aldose Reductase Inhibitor, Fidarestat. Int. J. Mol. Sci. 2023, 24, 2711. https://doi.org/10.3390/ijms24032711
El-Deeb OS, Elesawy RO, Eltokhy AK, Al-Shenawy HA, Ghanem HB, Rizk FH, Barhoma RA, Shalaby RH, Abdelsattar AM, Mashal SS, et al. Moderating Gut Microbiome/Mitochondrial Axis in Oxazolone Induced Ulcerative Colitis: The Evolving Role of β-Glucan and/or, Aldose Reductase Inhibitor, Fidarestat. International Journal of Molecular Sciences. 2023; 24(3):2711. https://doi.org/10.3390/ijms24032711
Chicago/Turabian StyleEl-Deeb, Omnia Safwat, Rasha Osama Elesawy, Amira K. Eltokhy, Hanan Alsaeid Al-Shenawy, Heba Bassiony Ghanem, Fatma H. Rizk, Ramez AE Barhoma, Rania H. Shalaby, Amal M. Abdelsattar, Shaimaa S. Mashal, and et al. 2023. "Moderating Gut Microbiome/Mitochondrial Axis in Oxazolone Induced Ulcerative Colitis: The Evolving Role of β-Glucan and/or, Aldose Reductase Inhibitor, Fidarestat" International Journal of Molecular Sciences 24, no. 3: 2711. https://doi.org/10.3390/ijms24032711
APA StyleEl-Deeb, O. S., Elesawy, R. O., Eltokhy, A. K., Al-Shenawy, H. A., Ghanem, H. B., Rizk, F. H., Barhoma, R. A., Shalaby, R. H., Abdelsattar, A. M., Mashal, S. S., Eshra, K. A., El-Sharaby, R. M., Ali, D. A., & Ibrahim, R. R. (2023). Moderating Gut Microbiome/Mitochondrial Axis in Oxazolone Induced Ulcerative Colitis: The Evolving Role of β-Glucan and/or, Aldose Reductase Inhibitor, Fidarestat. International Journal of Molecular Sciences, 24(3), 2711. https://doi.org/10.3390/ijms24032711