
A Phytoprostane from Gracilaria longissima Increases Platelet Activation, Platelet Adhesion to Leukocytes and Endothelial Cell Migration by Potential Binding to EP3 Prostaglandin Receptor

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Individual PhytoPs and PhytoFs Composition in Gracilaria Longissima Extract
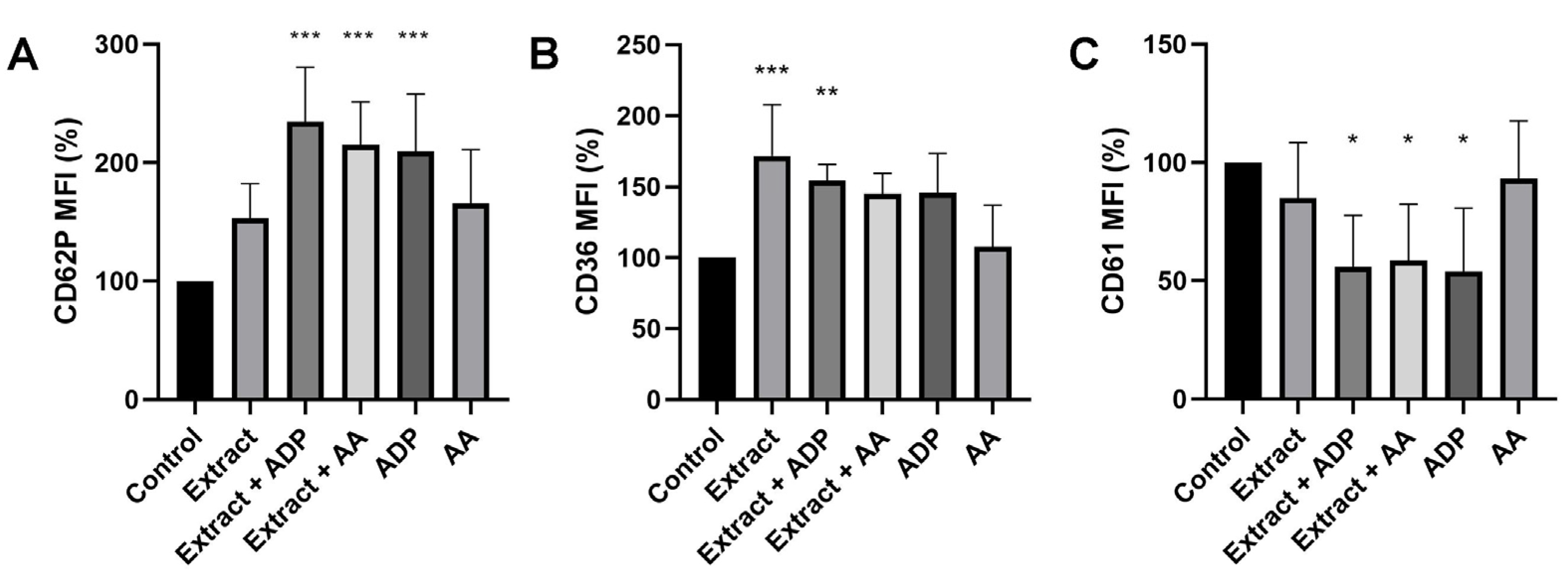
2.2. PhytoPs Extract Stimulates Platelet Activation and Aggregation
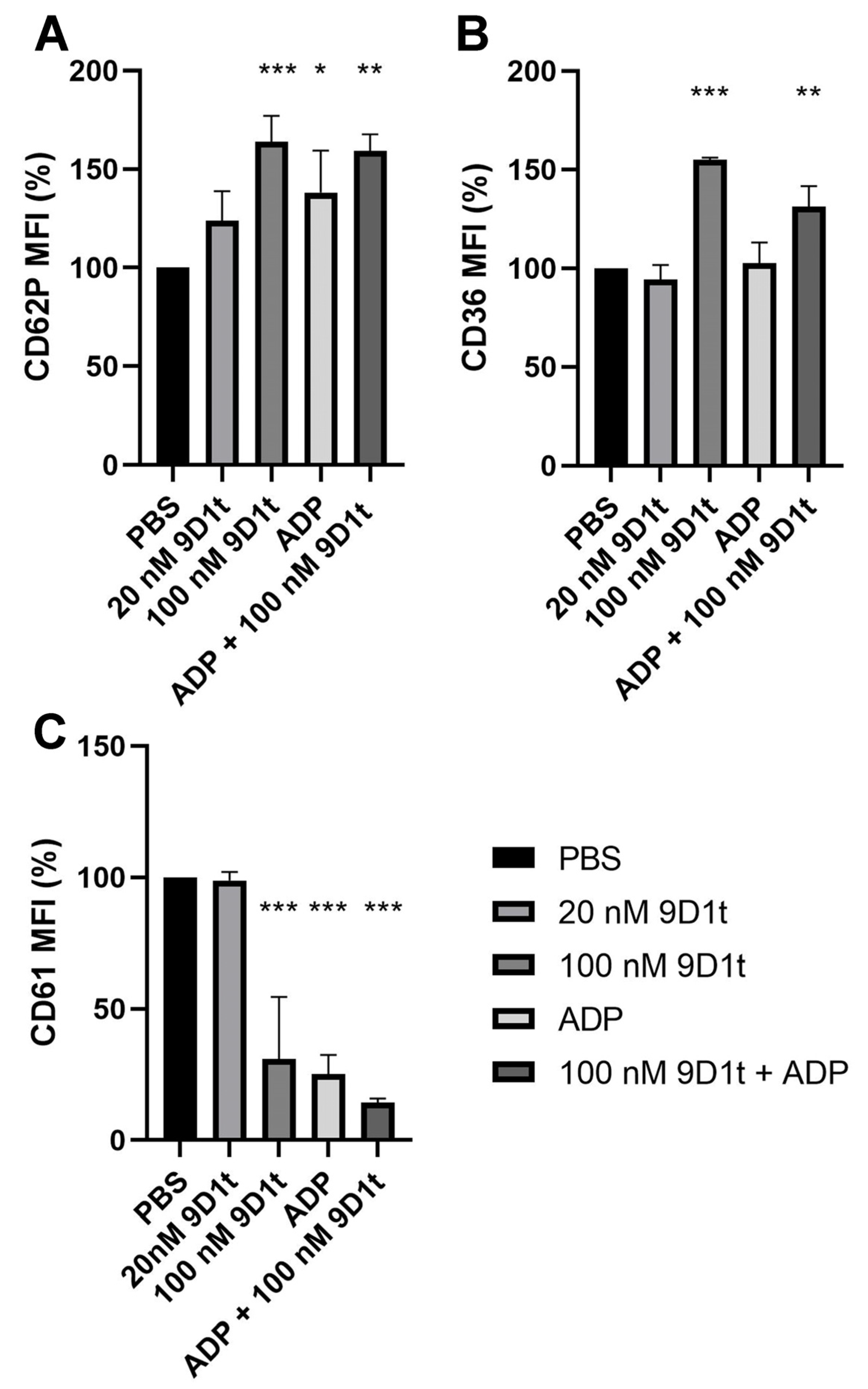
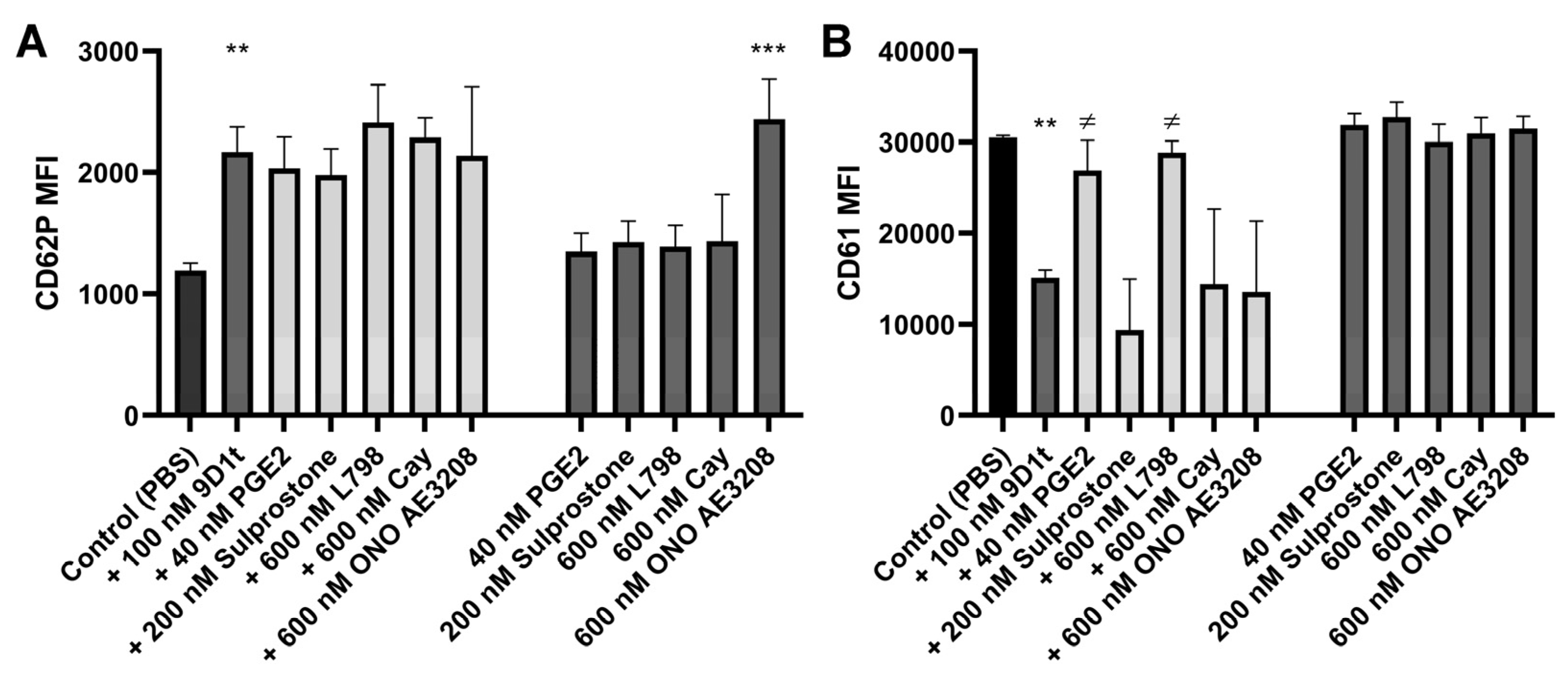
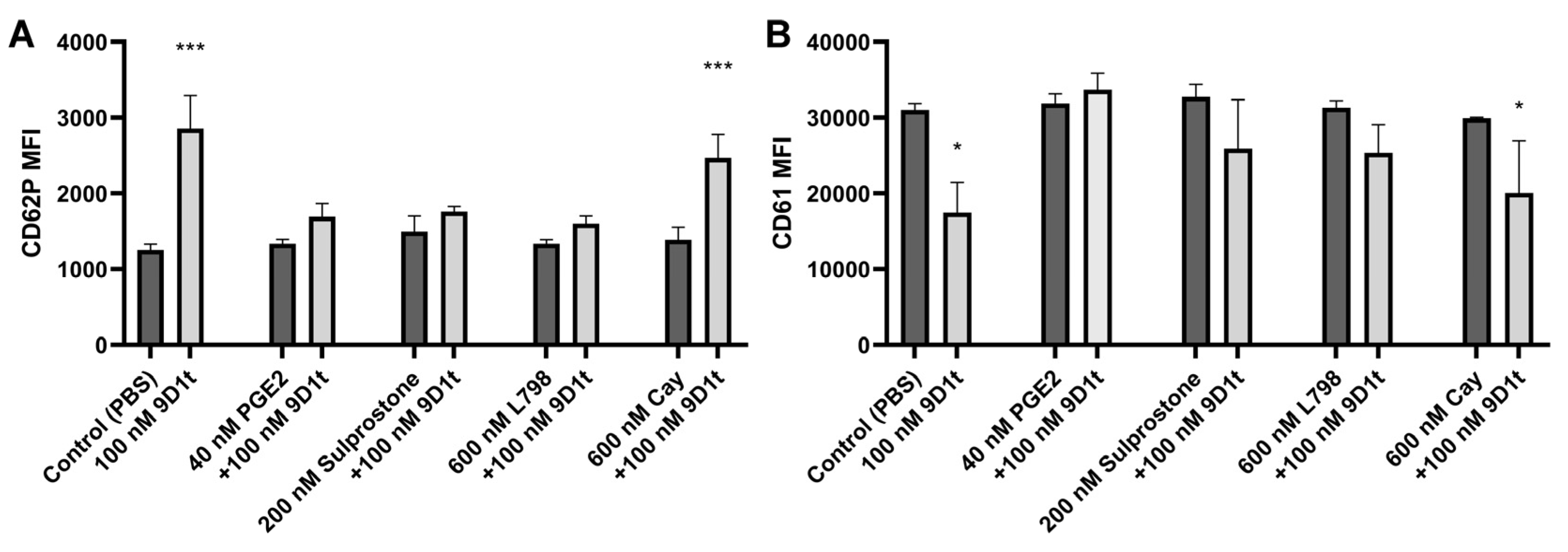
2.3. 9-D1t-PhytoP Affects Platelet Aggregation through EP Receptors
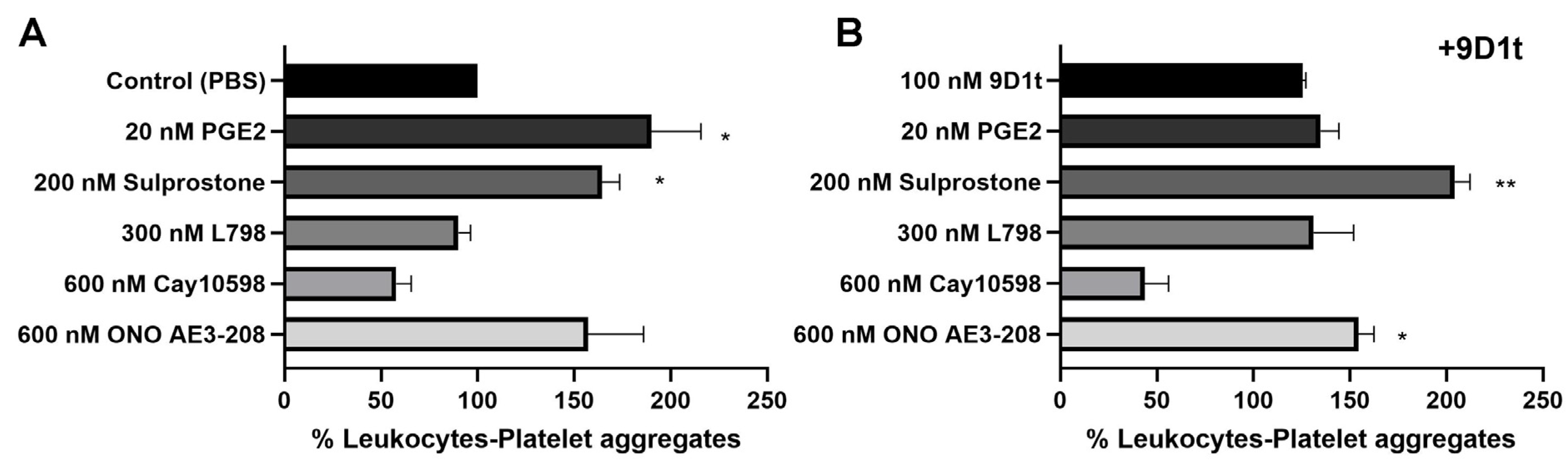
2.4. 9-D1t-PhytoP Alone Does Not Increase Platelet Adhesion to Leukocytes
2.5. 9-D1t-PhytoP Does Not Compromise Endothelial Cell Viability
2.6. 9-D1t-PhytoP Bioactivity over the Endothelium
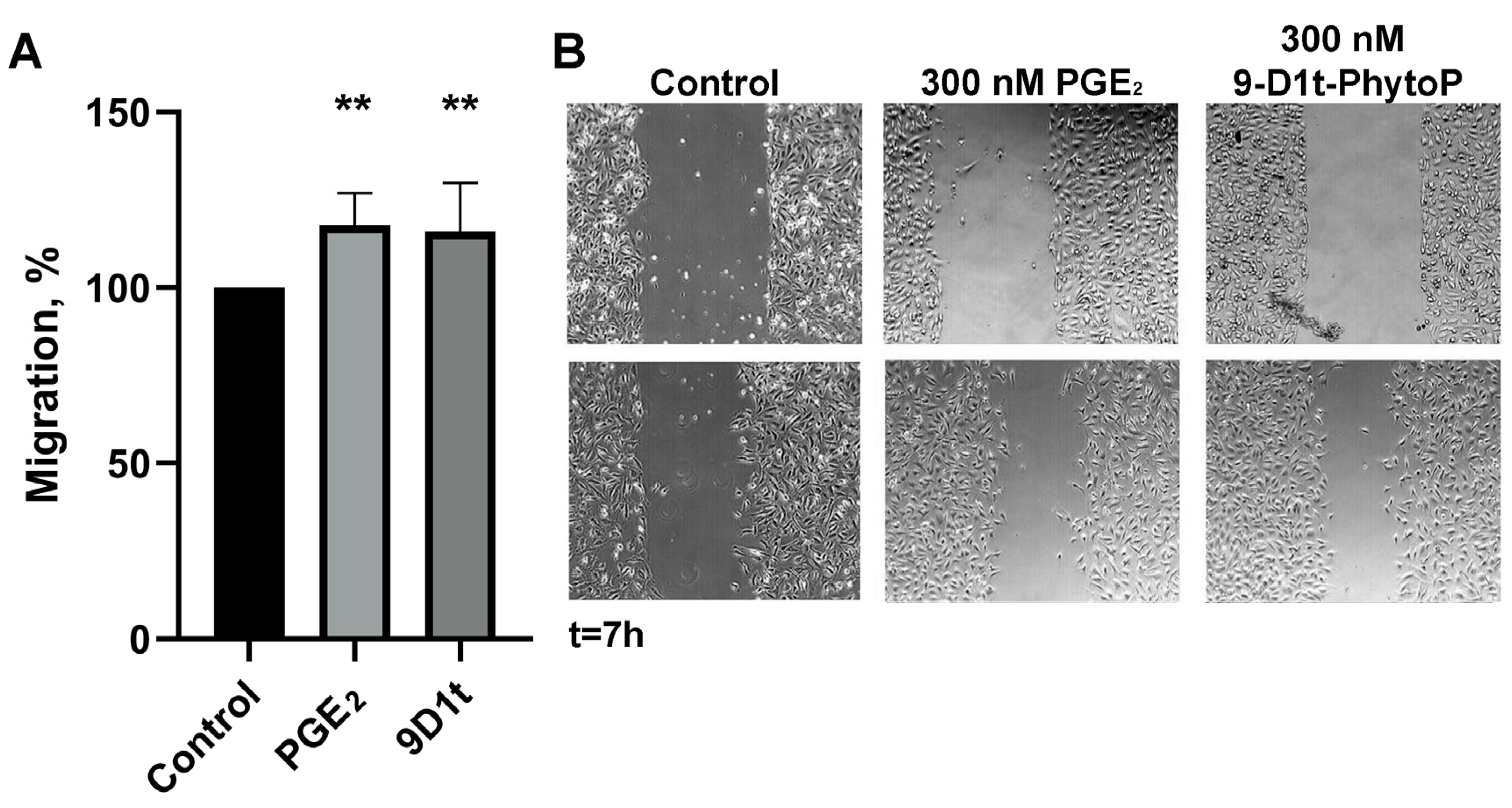
2.6.1. 9-D1t-PhytoP Contributes to In Vitro Migration
2.6.2. 9-D1t-PhytoP Induces an Endothelial Dysfunctional Status
2.7. Computational Methods for Discovering the Biological Receptors of 9-D1t-PhytoP
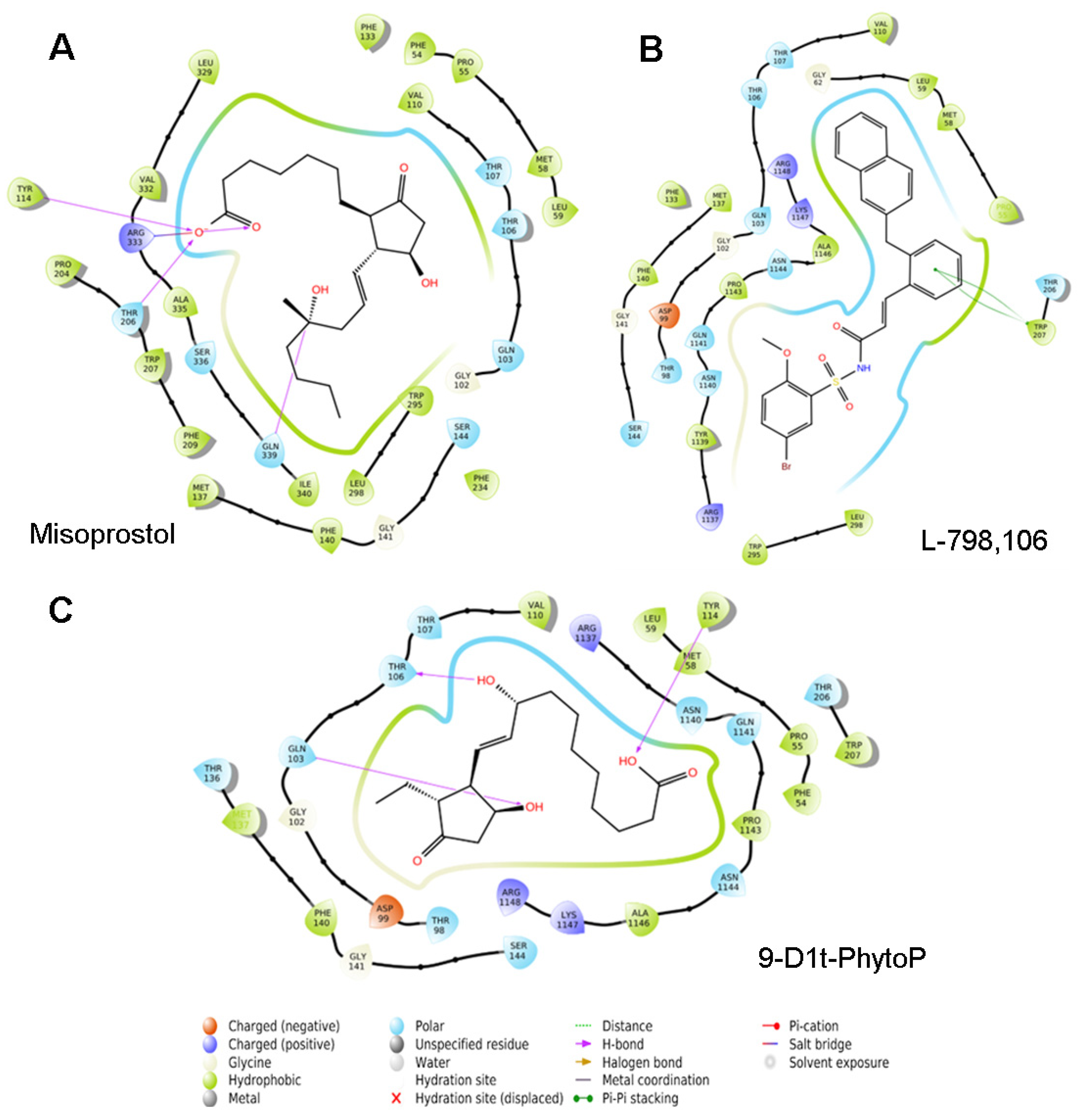
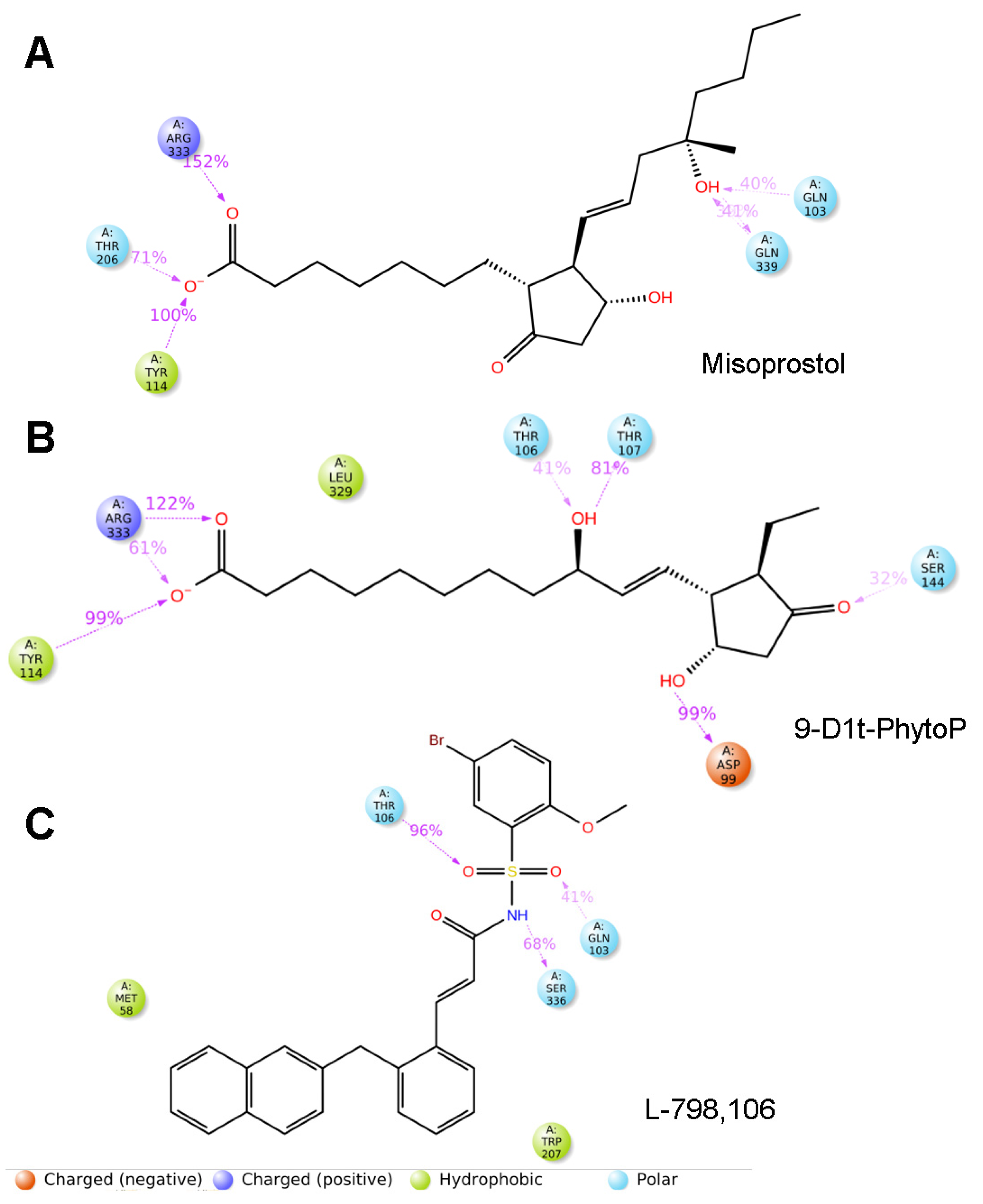
2.7.1. 9-D1t-PhytoP Binds to the Hydrophobic Pocket of EP3
2.7.2. 9-D1t-PhytoP Shares Hydrophobic Interactions with EP3 Agonists
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Analytical Extract Rich in Oxylipins
4.3. In Vitro Assessment of Platelet Activation and Polymorphonuclear Leukocytes (PMNs)-Platelet Aggregates
4.4. Cell Line and Culture Conditions
4.5. Cell Viability Assay
4.6. Cell Migration Assay
4.7. Biological Activity in an Endothelial Dysfunction Model
4.8. Blind Docking
4.9. Molecular Dynamics
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Medina, S.; Gil-Izquierdo, A.; Durand, T.; Ferreres, F.; Dominguez-Perles, R. Structural/Functional Matches and Divergences of Phytoprostanes and Phytofurans with Bioactive Human Oxylipins. Antioxidants 2018, 7, 165. [Google Scholar] [CrossRef] [PubMed]
- Collado-Gonzalez, J.; Cano-Lamadrid, M.; Perez-Lopez, D.; Carbonell-Barrachina, A.A.; Centeno, A.; Medina, S.; Grinan, I.; Guy, A.; Galano, J.M.; Durand, T.; et al. Effects of Deficit Irrigation, Rootstock, and Roasting on the Contents of Fatty Acids, Phytoprostanes, and Phytofurans in Pistachio Kernels. J. Agric. Food Chem. 2020, 68, 8915–8924. [Google Scholar] [CrossRef] [PubMed]
- Collado-Gonzalez, J.; Medina, S.; Durand, T.; Guy, A.; Galano, J.M.; Torrecillas, A.; Ferreres, F.; Gil-Izquierdo, A. New UHPLC-QqQ-MS/MS method for quantitative and qualitative determination of free phytoprostanes in foodstuffs of commercial olive and sunflower oils. Food Chem. 2015, 178, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Medina, S.; Collado-Gonzalez, J.; Ferreres, F.; Londono-Londono, J.; Jimenez-Cartagena, C.; Guy, A.; Durand, T.; Galano, J.M.; Gil-Izquierdo, A. Quantification of phytoprostanes—bioactive oxylipins—and phenolic compounds of Passiflora edulis Sims shell using UHPLC-QqQ-MS/MS and LC-IT-DAD-MS/MS. Food Chem. 2017, 229, 1–8. [Google Scholar] [CrossRef]
- Pinciroli, M.; Dominguez-Perles, R.; Abellan, A.; Guy, A.; Durand, T.; Oger, C.; Galano, J.M.; Ferreres, F.; Gil-Izquierdo, A. Comparative Study of the Phytoprostane and Phytofuran Content of indica and japonica Rice (Oryza sativa L.) Flours. J. Agric. Food Chem. 2017, 65, 8938–8947. [Google Scholar] [CrossRef]
- Pinciroli, M.; Dominguez-Perles, R.; Garbi, M.; Abellan, A.; Oger, C.; Durand, T.; Galano, J.M.; Ferreres, F.; Gil-Izquierdo, A. Impact of Salicylic Acid Content and Growing Environment on Phytoprostane and Phytofuran (Stress Biomarkers) in Oryza sativa L. J. Agric. Food Chem. 2018, 66, 12561–12570. [Google Scholar] [CrossRef] [PubMed]
- Lipan, L.; Collado-Gonzalez, J.; Dominguez-Perles, R.; Corell, M.; Bultel-Ponce, V.; Galano, J.M.; Durand, T.; Medina, S.; Gil-Izquierdo, A.; Carbonell-Barrachina, A. Phytoprostanes and Phytofurans-Oxidative Stress and Bioactive Compounds-in Almonds are Affected by Deficit Irrigation in Almond Trees. J. Agric. Food Chem. 2020, 68, 7214–7225. [Google Scholar] [CrossRef]
- Dominguez-Perles, R.; Abellan, A.; Leon, D.; Ferreres, F.; Guy, A.; Oger, C.; Galano, J.M.; Durand, T.; Gil-Izquierdo, A. Sorting out the phytoprostane and phytofuran profile in vegetable oils. Food Res. Int. 2018, 107, 619–628. [Google Scholar] [CrossRef]
- Carrasco-Del Amor, A.M.; Aguayo, E.; Collado-Gonzalez, J.; Guy, A.; Galano, J.M.; Durand, T.; Gil-Izquierdo, A. Impact of packaging atmosphere, storage and processing conditions on the generation of phytoprostanes as quality processing compounds in almond kernels. Food Chem. 2016, 211, 869–875. [Google Scholar] [CrossRef]
- Carrasco-Del Amor, A.M.; Aguayo, E.; Collado-Gonzalez, J.; Guy, A.; Galano, J.M.; Durand, T.; Gil-Izquierdo, A. Impact of processing conditions on the phytoprostanes profile of three types of nut kernels. Free Radic. Res. 2017, 51, 141–147. [Google Scholar] [CrossRef]
- Martinez Sanchez, S.; Dominguez-Perles, R.; Montoro-Garcia, S.; Gabaldon, J.A.; Guy, A.; Durand, T.; Oger, C.; Ferreres, F.; Gil-Izquierdo, A. Bioavailable phytoprostanes and phytofurans from Gracilaria longissima have anti-inflammatory effects in endothelial cells. Food Funct. 2020, 11, 5166–5178. [Google Scholar] [CrossRef] [PubMed]
- Campillo, M.; Medina, S.; Fanti, F.; Gallego-Gomez, J.I.; Simonelli-Munoz, A.; Bultel-Ponce, V.; Durand, T.; Galano, J.M.; Tomas-Barberan, F.A.; Gil-Izquierdo, A.; et al. Phytoprostanes and phytofurans modulate COX-2-linked inflammation markers in LPS-stimulated THP-1 monocytes by lipidomics workflow. Free Radic. Biol. Med. 2021, 167, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Lara-Guzman, O.J.; Gil-Izquierdo, A.; Medina, S.; Osorio, E.; Alvarez-Quintero, R.; Zuluaga, N.; Oger, C.; Galano, J.M.; Durand, T.; Munoz-Durango, K. Oxidized LDL triggers changes in oxidative stress and inflammatory biomarkers in human macrophages. Redox Biol. 2018, 15, 1–11. [Google Scholar] [CrossRef]
- Markovic, T.; Jakopin, Z.; Dolenc, M.S.; Mlinaric-Rascan, I. Structural features of subtype-selective EP receptor modulators. Drug Discov. Today 2017, 22, 57–71. [Google Scholar] [CrossRef]
- Petrucci, G.; De Cristofaro, R.; Rutella, S.; Ranelletti, F.O.; Pocaterra, D.; Lancellotti, S.; Habib, A.; Patrono, C.; Rocca, B. Prostaglandin E2 differentially modulates human platelet function through the prostanoid EP2 and EP3 receptors. J. Pharmacol. Exp. Ther. 2011, 336, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.Z.; Ashby, B.; Sheth, S.B. Distribution of prostaglandin IP and EP receptor subtypes and isoforms in platelets and human umbilical artery smooth muscle cells. Br. J. Haematol. 1998, 102, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Iyu, D.; Juttner, M.; Glenn, J.R.; White, A.E.; Johnson, A.J.; Fox, S.C.; Heptinstall, S. PGE1 and PGE2 modify platelet function through different prostanoid receptors. Prostaglandins Other Lipid Mediat. 2011, 94, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Philipose, S.; Konya, V.; Sreckovic, I.; Marsche, G.; Lippe, I.T.; Peskar, B.A.; Heinemann, A.; Schuligoi, R. The prostaglandin E2 receptor EP4 is expressed by human platelets and potently inhibits platelet aggregation and thrombus formation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2416–2423. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.S.; Jones, R.L. Potentiation of aggregation and inhibition of adenylate cyclase in human platelets by prostaglandin E analogues. Br. J. Pharmacol. 1993, 108, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Heptinstall, S.; Espinosa, D.I.; Manolopoulos, P.; Glenn, J.R.; White, A.E.; Johnson, A.; Dovlatova, N.; Fox, S.C.; May, J.A.; Hermann, D.; et al. DG-041 inhibits the EP3 prostanoid receptor--a new target for inhibition of platelet function in atherothrombotic disease. Platelets 2008, 19, 605–613. [Google Scholar] [CrossRef]
- Kuriyama, S.; Kashiwagi, H.; Yuhki, K.; Kojima, F.; Yamada, T.; Fujino, T.; Hara, A.; Takayama, K.; Maruyama, T.; Yoshida, A.; et al. Selective activation of the prostaglandin E2 receptor subtype EP2 or EP4 leads to inhibition of platelet aggregation. Thromb. Haemost. 2010, 104, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Schober, L.J.; Khandoga, A.L.; Dwivedi, S.; Penz, S.M.; Maruyama, T.; Brandl, R.; Siess, W. The role of PGE(2) in human atherosclerotic plaque on platelet EP(3) and EP(4) receptor activation and platelet function in whole blood. J. Thromb. Thrombolysis 2011, 32, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Schober, L.J.; Khandoga, A.L.; Penz, S.M.; Siess, W. The EP3-agonist sulprostone, but not prostaglandin E2 potentiates platelet aggregation in human blood. Thromb. Haemost. 2010, 103, 1268–1269. [Google Scholar] [CrossRef] [PubMed]
- Rinder, C.S.; Student, L.A.; Bonan, J.L.; Rinder, H.M.; Smith, B.R. Aspirin does not inhibit adenosine diphosphate-induced platelet alpha-granule release. Blood 1993, 82, 505–512. [Google Scholar] [CrossRef]
- Lagarde, M.; Chen, P.; Vericel, E.; Guichardant, M. Fatty acid-derived lipid mediators and blood platelet aggregation. Prostaglandins Leukot. Essent. Fatty Acids 2010, 82, 227–230. [Google Scholar] [CrossRef]
- Jadhav, V.; Jabre, A.; Lin, S.Z.; Lee, T.J. EP1- and EP3-receptors mediate prostaglandin E2-induced constriction of porcine large cerebral arteries. J. Cereb. Blood Flow Metab. 2004, 24, 1305–1316. [Google Scholar] [CrossRef]
- Shantsila, E.; Montoro-Garcia, S.; Lip, G.Y. Monocytes circulate in constant reversible interaction with platelets in a [Ca2+]-dependent manner. Platelets 2014, 25, 197–201. [Google Scholar] [CrossRef]
- Montoro-Garcia, S.; Shantsila, E.; Hernandez-Romero, D.; Jover, E.; Valdes, M.; Marin, F.; Lip, G.Y. Small-size platelet microparticles trigger platelet and monocyte functionality and modulate thrombogenesis via P-selectin. Br. J. Haematol. 2014, 166, 571–580. [Google Scholar] [CrossRef]
- Rao, R.; Redha, R.; Macias-Perez, I.; Su, Y.; Hao, C.; Zent, R.; Breyer, M.D.; Pozzi, A. Prostaglandin E2-EP4 receptor promotes endothelial cell migration via ERK activation and angiogenesis in vivo. J. Biol. Chem. 2007, 282, 16959–16968. [Google Scholar] [CrossRef]
- Lau, S.; Gossen, M.; Lendlein, A.; Jung, F. Venous and Arterial Endothelial Cells from Human Umbilical Cords: Potential Cell Sources for Cardiovascular Research. Int. J. Mol. Sci. 2021, 22, 978. [Google Scholar] [CrossRef]
- Geenen, I.L.; Molin, D.G.; van den Akker, N.M.; Jeukens, F.; Spronk, H.M.; Schurink, G.W.; Post, M.J. Endothelial cells (ECs) for vascular tissue engineering: Venous ECs are less thrombogenic than arterial ECs. J. Tissue Eng. Regen. Med. 2015, 9, 564–576. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Sanchez, S.M.; Perez-Sanchez, H.; Antonio Gabaldon, J.; Abellan-Aleman, J.; Montoro-Garcia, S. Multifunctional Peptides from Spanish Dry-Cured Pork Ham: Endothelial Responses and Molecular Modeling Studies. Int. J. Mol. Sci. 2019, 20, 4204. [Google Scholar] [CrossRef] [PubMed]
- Hollenstein, K. Structures shed light on prostanoid signaling. Nat. Chem. Biol. 2019, 15, 3–5. [Google Scholar] [CrossRef]
- Morimoto, K.; Suno, R.; Hotta, Y.; Yamashita, K.; Hirata, K.; Yamamoto, M.; Narumiya, S.; Iwata, S.; Kobayashi, T. Crystal structure of the endogenous agonist-bound prostanoid receptor EP3. Nat. Chem. Biol. 2019, 15, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Vigor, C.; Balas, L.; Guy, A.; Bultel-Poncé, V.; Reversat, G.; Galano, J.M.; Durand, T.; Oger, C. Isoprostanoids, Isofuranoids and Isoketals—From Synthesis to Lipidomics. Eur. J. Org. Chem. 2022, 2022, 137–149. [Google Scholar] [CrossRef]
- van Velzen, J.F.; Laros-van Gorkom, B.A.; Pop, G.A.; van Heerde, W.L. Multicolor flow cytometry for evaluation of platelet surface antigens and activation markers. Thromb. Res. 2012, 130, 92–98. [Google Scholar] [CrossRef]
- Hechler, B.; Cattaneo, M.; Gachet, C. The P2 receptors in platelet function. Semin. Thromb. Hemost. 2005, 31, 150–161. [Google Scholar] [CrossRef]
- Alburquerque-Gonzalez, B.; Bernabe-Garcia, A.; Bernabe-Garcia, M.; Ruiz-Sanz, J.; Lopez-Calderon, F.F.; Gonnelli, L.; Banci, L.; Pena-Garcia, J.; Luque, I.; Nicolas, F.J.; et al. The FDA-Approved Antiviral Raltegravir Inhibits Fascin1-Dependent Invasion of Colorectal Tumor Cells In Vitro and In Vivo. Cancers 2021, 13, 861. [Google Scholar] [CrossRef]
- Alburquerque-Gonzalez, B.; Bernabe-Garcia, M.; Montoro-Garcia, S.; Bernabe-Garcia, A.; Rodrigues, P.C.; Ruiz Sanz, J.; Lopez-Calderon, F.F.; Luque, I.; Nicolas, F.J.; Cayuela, M.L.; et al. New role of the antidepressant imipramine as a Fascin1 inhibitor in colorectal cancer cells. Exp. Mol. Med. 2020, 52, 281–292. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tapia-Abellan, A.; Angosto-Bazarra, D.; Martinez-Banaclocha, H.; de Torre-Minguela, C.; Ceron-Carrasco, J.P.; Perez-Sanchez, H.; Arostegui, J.I.; Pelegrin, P. MCC950 closes the active conformation of NLRP3 to an inactive state. Nat. Chem. Biol. 2019, 15, 560–564. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Roos, K.; Wu, C.; Damm, W.; Reboul, M.; Stevenson, J.M.; Lu, C.; Dahlgren, M.K.; Mondal, S.; Chen, W.; Wang, L.; et al. OPLS3e: Extending Force Field Coverage for Drug-Like Small Molecules. J. Chem. Theory Comput. 2019, 15, 1863–1874. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Stroganov, O.V.; Novikov, F.N.; Stroylov, V.S.; Kulkov, V.; Chilov, G.G. Lead finder: An approach to improve accuracy of protein-ligand docking, binding energy estimation, and virtual screening. J. Chem. Inf. Model 2008, 48, 2371–2385. [Google Scholar] [CrossRef]
- Gasteiger, J.; Marsili, M. Iterative partial equalization of orbital electronegativity—A rapid access to atomic charges. Tetrahedron 1980, 36, 3219–3228. [Google Scholar] [CrossRef]
- Koynova, R.; Caffrey, M. Phases and phase transitions of the sphingolipids. Biochim. Biophys. Acta 1995, 1255, 213–236. [Google Scholar] [CrossRef]
- Tapias, D.; Sanders, D.P.; Bravetti, A. Geometric integrator for simulations in the canonical ensemble. J. Chem. Phys. 2016, 145, 084113. [Google Scholar] [CrossRef]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant pressure molecular dynamics algorithms. J. Chem. Phys. 1994, 101, 4177–4189. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PhytoPs Z | PhytoFs Y | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample | FP-1 | FP-2 | FP-3 | FP-4 | FP-5 | FP-6 | FP-7 | Total | FF-1 | FF-2 | FF-3 | Total |
| 1 | 38.5 | 1591 | 55.1 | 0.2 | 3.1 | 9.0 | 13.6 | 1717.9 | 171.9 | 23.1 | 13.6 | 208.7 |
| 2 | 25.2 | 752 | 26.7 | <0.1 | 1.1 | 3.6 | 7.0 | 815.7 | 298.2 | 31.7 | 6.7 | 336.6 |
| 3 | 52.8 | 1967 | 44.8 | 0.2 | 3.4 | 10.0 | 12.1 | 2100.6 | 307.6 | 7.7 | 12.1 | 326.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montoro-García, S.; Martínez-Sánchez, S.; Carmena-Bargueño, M.; Pérez-Sánchez, H.; Campillo, M.; Oger, C.; Galano, J.-M.; Durand, T.; Gil-Izquierdo, Á.; Gabaldón, J.A. A Phytoprostane from Gracilaria longissima Increases Platelet Activation, Platelet Adhesion to Leukocytes and Endothelial Cell Migration by Potential Binding to EP3 Prostaglandin Receptor. Int. J. Mol. Sci. 2023, 24, 2730. https://doi.org/10.3390/ijms24032730
Montoro-García S, Martínez-Sánchez S, Carmena-Bargueño M, Pérez-Sánchez H, Campillo M, Oger C, Galano J-M, Durand T, Gil-Izquierdo Á, Gabaldón JA. A Phytoprostane from Gracilaria longissima Increases Platelet Activation, Platelet Adhesion to Leukocytes and Endothelial Cell Migration by Potential Binding to EP3 Prostaglandin Receptor. International Journal of Molecular Sciences. 2023; 24(3):2730. https://doi.org/10.3390/ijms24032730
Chicago/Turabian StyleMontoro-García, Silvia, Sara Martínez-Sánchez, Miguel Carmena-Bargueño, Horacio Pérez-Sánchez, María Campillo, Camille Oger, Jean-Marie Galano, Thierry Durand, Ángel Gil-Izquierdo, and José Antonio Gabaldón. 2023. "A Phytoprostane from Gracilaria longissima Increases Platelet Activation, Platelet Adhesion to Leukocytes and Endothelial Cell Migration by Potential Binding to EP3 Prostaglandin Receptor" International Journal of Molecular Sciences 24, no. 3: 2730. https://doi.org/10.3390/ijms24032730
APA StyleMontoro-García, S., Martínez-Sánchez, S., Carmena-Bargueño, M., Pérez-Sánchez, H., Campillo, M., Oger, C., Galano, J. -M., Durand, T., Gil-Izquierdo, Á., & Gabaldón, J. A. (2023). A Phytoprostane from Gracilaria longissima Increases Platelet Activation, Platelet Adhesion to Leukocytes and Endothelial Cell Migration by Potential Binding to EP3 Prostaglandin Receptor. International Journal of Molecular Sciences, 24(3), 2730. https://doi.org/10.3390/ijms24032730