
The Impact of Permethrin and Cypermethrin on Plants, Soil Enzyme Activity, and Microbial Communities





Abstract
:1. Introduction
2. Results
2.1. Effect of Cypermethrin and Permethrin on Zea mays
2.2. Effect of Cypermethrin and Permethrin on the Activity of Soil Enzymes
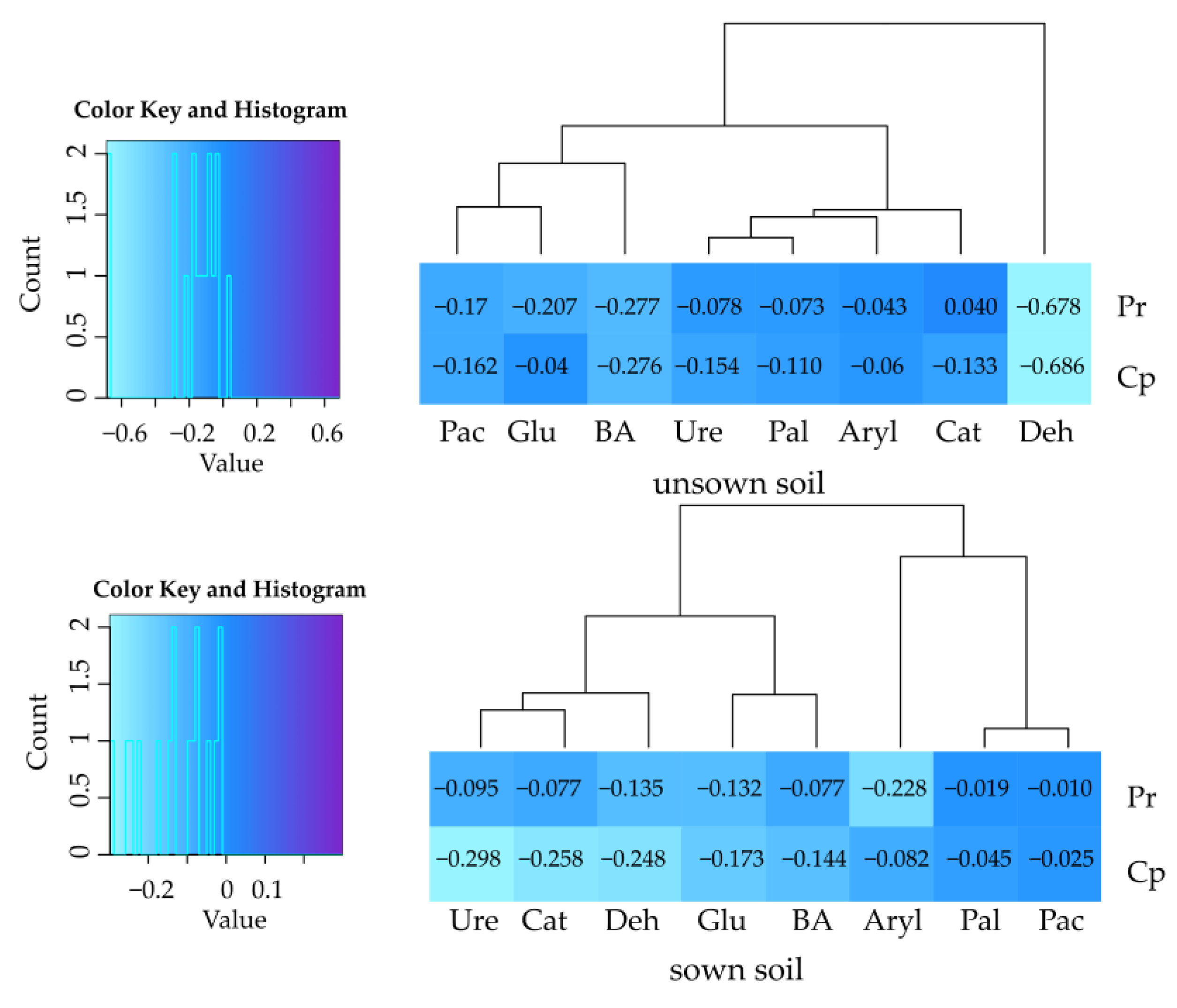
2.3. Effect of Cypermethrin and Permethrin on Soil Microorganisms
2.3.1. Cultured Microorganisms
2.3.2. Bacteria Determined with the NGS Method
2.3.3. Fungi Determined with the NGS Method
3. Discussion
3.1. Effect of Pesticides on Plants
3.2. Effect of Pesticides on the Activity of Soil Enzymes
3.3. Effect of Pesticides on Soil Microorganisms
4. Materials and Methods
4.1. Soil
4.2. Insecticides
4.3. Study Design
4.4. Soil Microbiological Analysis
4.4.1. Culture Microbes
4.4.2. Isolation of DNA and Identification of Bacteria and Fungi with the NGS Method
4.5. Biochemical Analyses of Soil
4.6. Data and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide Pesticide Usage and Its Impacts on Ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef]
- Wdowenko, N.; Tomilin, O.; Kowalenko, L.; Badri, G.; Konczakowski, E. Global Trends and Development Prospects of The Market of Plant Protection Products. J. Agric. Resour. Econ. 2022, 8, 179–205. [Google Scholar] [CrossRef]
- Badgujar, C.; Mansur, H.; Flippo, D. Pest-Infested Soybean Leaf Image Classification with Deep Learning Techniques for Integrated Pest Management (IPM). In Proceedings of the 2022 ASABE Annual International Meeting, Houston, TX, USA, 17–20 July 2022; p. 2201096. [Google Scholar] [CrossRef]
- FAOSTAT, Pesticides Use. Available online: https://www.fao.org/faostat/en/#data/RP/visualize (accessed on 5 December 2022).
- Tang, W.; Wang, D.; Wang, J.; Wu, Z.; Li, L.; Huang, M.; Xu, S.; Yan, D. Pyrethroid Pesticide Residues in the Global Environment: An Overview. Chemosphere 2018, 191, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Burns, C.J.; Pastoor, T.P. Pyrethroid Epidemiology: A Quality-Based Review. Crit. Rev. Toxicol. 2018, 48, 297–311. [Google Scholar] [CrossRef]
- Bhatt, P.; Rene, E.R.; Huang, Y.; Wu, X.; Zhou, Z.; Li, J.; Kumar, A.J.; Sharma, A.; Chen, S. Indigenous Bacterial Consortium-Mediated Cypermethrin Degradation in the Presence of Organic Amendments and Zea Mays Plants. Environ. Res. 2022, 212, 113137. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Rene, E.R.; Huang, Y.; Lin, Z.; Pang, S.; Zhang, W.; Chen, S. Systems Biology Analysis of Pyrethroid Biodegradation in Bacteria and Its Effect on the Cellular Environment of Pests and Humans. J. Environ. Chem. Eng. 2021, 9, 106582. [Google Scholar] [CrossRef]
- Ndlovu, N.N.; Little, K.; Baillie, B.; Rolando, C. An Evaluation of the Environmental Behaviour, Fate and Risk of Key Pesticides Used in South African Forest Plantations. South J. For. Sc. 2022, 84, 83–92. [Google Scholar] [CrossRef]
- Rivera-Dávila, O.L.; Sánchez-Martínez, G.; Rico-Martínez, R. Toxicity Tests, Bioaccumulation and Residuality of Pyrethroid Insecticides Commonly Used to Control Conifer Bark Beetles in Mexico. Ecotoxicology 2022, 31, 782–796. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Statement on the Available Outcomes of the Human Health Assessment in the Context of the Pesticides Peer Review of the Active Substance Chlorpyrifos. EFSA J. 2019, 17, e05809. [Google Scholar] [CrossRef]
- Li, H.; Cheng, F.; Wei, Y.; Lydy, M.J.; You, J. Global Occurrence of Pyrethroid Insecticides in Sediment and the Associated Toxicological Effects on Benthic Invertebrates: An Overview. J. Hazard. Mater. 2017, 324, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.R.; David, A.; Freire, C.; Fernández, M.F.; D’Cruz, S.C.; Reina-Pérez, I.; Fini, J.-B.; Blaha, L. Pyrethroids and Developmental Neurotoxicity—A Critical Review of Epidemiological Studies and Supporting Mechanistic Evidence. Environ. Res. 2022, 214, 113935. [Google Scholar] [CrossRef] [PubMed]
- U.S. EPA. Environmental Protection Agency. Permethrin, Resmethrin, d-Phenothrin (Sumithrin®): Synthetic Pyrethroids for Mosquito Control. 2022. Available online: https://www.epa.gov/mosquitocontrol/permethrin-resmethrin-d-phenothrin-sumithrinr-synthetic-pyrethroids-mosquito (accessed on 14 January 2023).
- Bragança, I.; Mucha, A.P.; Tomasino, M.P.; Santos, F.; Lemos, P.C.; Delerue-Matos, C.; Domingues, V.F. Deltamethrin Impact in a Cabbage Planted Soil: Degradation and Effect on Microbial Community Structure. Chemosphere 2019, 220, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.; Yang, X.; Fleskens, L.; Ritsema, C.J.; Geissen, V. Environmental and Human Health at Risk—Scenarios to Achieve the Farm to Fork 50% Pesticide Reduction Goals. Environ. Inter. 2022, 165, 107296. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Zhao, J.; Zhu, X.; Chen, S.; Yang, X. Pyrethroid Bioaccumulation in Wild Fish Linked to Geographic Distribution and Feeding Habit. J. Hazard. Mater. 2022, 430, 128470. [Google Scholar] [CrossRef]
- Zhan, H.; Huang, Y.; Lin, Z.; Bhatt, P.; Chen, S. New Insights into the Microbial Degradation and Catalytic Mechanism of Synthetic Pyrethroids. Environ. Res. 2020, 182, 109138. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Jiang, Y.; Gong, L.; Chen, X.; Xie, Q.; Jin, Y.; Du, J.; Wang, S.; Liu, G. Mechanism of β-Cypermethrin Metabolism by Bacillus Cereus GW-01. Chem. Eng. J. 2022, 430, 132961. [Google Scholar] [CrossRef]
- Morgan, M.K.; Sheldon, L.S.; Croghan, C.W.; Jones, P.A.; Chuang, J.C.; Wilson, N.K. An Observational Study of 127 Preschool Children at Their Homes and Daycare Centers in Ohio: Environmental Pathways to Cis- and Trans-Permethrin Exposure. Environ. Res. 2007, 104, 266–274. [Google Scholar] [CrossRef]
- Bravo, N.; Garí, M.; Grimalt, J.O. Occupational and Residential Exposures to Organophosphate and Pyrethroid Pesticides in a Rural Setting. Environ. Res. 2022, 214, 114186. [Google Scholar] [CrossRef]
- Akelma, H.; Kilic, E.T.; Salik, F.; Bicak, E.A.; Yektas, A. Pyrethroid Intoxication: A Rare Case Report and Literature Review. Niger. J. Clin. Pr. 2019, 22, 442–444. [Google Scholar] [CrossRef]
- Shahid, M.; Khan, M.S.; Syed, A.; Marraiki, N.; Elgorban, A.M. Mesorhizobium Ciceri as Biological Tool for Improving Physiological, Biochemical and Antioxidant State of Cicer Aritienum (L.) Under Fungicide Stress. Sci. Rep. 2021, 11, 9655. [Google Scholar] [CrossRef]
- Ajermoun, N.; Aghris, S.; Ettadili, F.; Alaoui, O.T.; Laghrib, F.; Farahi, A.; Lahrich, S.; Bakasse, M.; Saqrane, S.; El Mhammedi, M.A. Phytotoxic Effect of The Insecticide Imidacloprid in Phaseolus Vulgaris L. Plant and Evaluation of Its Bioaccumulation and Translocation by Electrochemical Methods. Environ. Res. 2022, 214, 113794. [Google Scholar] [CrossRef]
- Sheng, Y.; Benmati, M.; Guendouzi, S.; Benmati, H.; Yuan, Y.; Song, J.; Xia, C.; Berkani, M. Latest Eco-Friendly Approaches for Pesticides Decontamination Using Microorganisms and Consortia Microalgae: A Comprehensive Insights, Challenges, and Perspectives. Chemosphere 2022, 308, 136183. [Google Scholar] [CrossRef]
- Ortiz-Hernández, M.L.; Sánchez-Salinas, E.; Dantán-González, E.; Castrejón-Godínez, M.L.; Ortiz-Hernández, M.L.; Sánchez-Salinas, E.; Dantán-González, E.; Castrejón-Godínez, M.L. Pesticide Biodegradation: Mechanisms, Genetics and Strategies to Enhance the Process. In Biodegradation-Life of Science; IntechOpen: London, UK, 2013; Volume 10, pp. 251–287. ISBN 978-953-51-1154-2. [Google Scholar]
- Das, A.C.; Mukherjee, D. Soil Application of Insecticides Influences Microorganisms and Plant Nutrients. Appl. Soil Ecol. 2000, 14, 55–62. [Google Scholar] [CrossRef]
- Sethi, S.; Mathur, N.; Bhatnagar, P. Effects of Synthetic Pyrethroids on Phosphate Solubilizing Activity of Microorganisms. Int. J. Curr. Microbiol. App. Sci. 2013, 2, 240–246. [Google Scholar]
- Tefera, T.; Kanampiu, F.; De Groote, H.; Hellin, J.; Mugo, S.; Kimenju, S.; Beyene, Y.; Boddupalli, P.M.; Shiferaw, B.; Banziger, M. The metal silo: An Effective Grain Storage Technology for Reducing Post-harvest Insect and Pathogen Losses in Maize While Improving Smallholder Farmers’ Food Security in Developing Countries. Crop Prot. 2011, 30, 240–245. [Google Scholar] [CrossRef]
- Awika, J.M. Major Cereal Grains Production and Use around the World. Advances in Cereal Science: Implications to Food Processing and Health Promotion. In Advances in Cereal Science: Implications to Food Processing and Health Promotion; American Chemical Society: Washington, DC, USA, 2011; pp. 1–13. [Google Scholar] [CrossRef]
- Galani, Y.J.H.; Ligowe, I.S.; Kieffer, M.; Kamalongo, D.; Kambwiri, A.M.; Kuwali, P.; Thierfelder, C.; Dougill, A.J.; Gong, Y.Y.; Orfila, C. Conservation Agriculture Affects Grain and Nutrient Yields of Maize (Zea Mays L.) and Can Impact Food and Nutrition Security in Sub-Saharan Africa. Front. Nutr. 2022, 8, 804663. [Google Scholar] [CrossRef]
- Kumar, P.; Choudhary, M.; Halder, T.; Prakash, N.R.; Singh, V.; Vineeth, T.V.; Sheoran, S.; Ravikiran, K.T.; Longmei, N.; Rakshit, S.; et al. Salinity Stress Tolerance and Omics Approaches: Revisiting the Progress and Achievements in Major Cereal Crops. Heredity 2022, 128, 497–518. [Google Scholar] [CrossRef] [PubMed]
- Purewal, S.S.; Kaur, P.; Bangar, S.P.; Sandhu, K.S.; Singh, S.K.; Kaur, M. Maize. Nutritional Composition, Processing, and Industrial Uses, 1st ed.; CRC Press: Boca Raton, FL, USA, 2022; Volume 32, p. 383. [Google Scholar] [CrossRef]
- Piperno, D.R.; Ranere, A.J.; Holst, I.; Iriarte, J.; Dickau, L. Starch Grain and Phytolith Evidence for Early Ninth Millennium B.P. Maize from The Central Balsas River Valley, Mexico. PNAS 2009, 109, 5019–5024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strada, J.; Bruno, C.; Nassetta, M.M.; Balzarini, M.G.; Conles, M.Y.; Rojas, D.; Ricca, A.P.; Cristos, D.; Martinez, M.J. Evaluation of Degradation of Organophosphate Insecticide Residues in Stored Corn (Zea mays L.) Grains. Agriscientia 2021, 38, 53–61. [Google Scholar] [CrossRef]
- Hasnaki, R.; Ziaee, M.; Mahdavi, V. Pesticide Residues in Corn and Soil of Corn fields of Khuzestan, Iran, and Potential Health Risk Assessment. J. Food Comp. Anal. 2023, 115, 104972. [Google Scholar] [CrossRef]
- Yusà, V.; Fernández, S.F.; Dualde, P.; López, A.; Lacomba, I.; Coscollà, C. Exposure to Non-Persistent Pesticides in the Spanish Population Using Biomonitoring: A Review. Envioron. Res. 2022, 205, 112437. [Google Scholar] [CrossRef] [PubMed]
- Tarazona, J.V.; Cattaneo, I.; Niemann, L.; Pedraza-Diaz, S.; González-Caballero, M.C.; de Alba-Gonzalez, M.; Cañas, A.; Dominguez-Morueco, N.; Esteban-López, M.; Castaño, A.; et al. A Tiered Approach for Assessing Individual and Combined Risk of Pyrethroids Using Human Biomonitoring Data. Toxics 2022, 10, 451. [Google Scholar] [CrossRef]
- Fama, F.; Feltracco, M.; Moro, G.; Barbaro, E.; Bassanello, M.; Gambaro, A.; Zanardi, C. Pesticides Monitoring in Biological Fluids: Mapping the Gaps in Analytical Strategies. Talanta 2023, 253, 123969. [Google Scholar] [CrossRef]
- Calvo-Agudo, M.; Tooker, J.F.; Dicke, M.; Tena, A. Insecticide-Contaminated Honeydew: Risks for Beneficial Insects. Biol. Rev. 2022, 97, 664–678. [Google Scholar] [CrossRef]
- Zhang, J.; Sheng, X.; Cao, J.; Fang, S.; Liu, X.; Liu, X.; Peng, L.; Sun, H.; Weng, R. Occurrence and Risk Exposure Assessment of Multiple Pesticide Residues in Edible Mint in China. J. Food Compost. Anal. 2023, 116, 105071. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, T.; Liu, X.; Wang, S.; Wang, K.; Xiao, R.; Chen, X.; Zhang, T. Ecological Risk Assessment and Environment Carrying Capacity of Soil Pesticide Residues in Vegetable Ecosystem in the Three Gorges Reservoir Area. J. Hazard. Mater. 2022, 435, 128987. [Google Scholar] [CrossRef]
- Sabzevari, S.; Hofman, J. Currently Used Pesticides’ Occurrence in Soils: Recent Results and Advances in Soil-Monitoring and Survey Studies. In Pesticides in Soils: Occurrence, Fate, Control and Remediation; Rodríguez-Cruz, M.S., Sánchez-Martín, M.J., Eds.; The Handbook of Environmental Chemistry; Springer International Publishing: Cham, Switzerland, 2022; pp. 1–33. ISBN 978-3-030-90546-0. [Google Scholar]
- Martínez Galera, M.; Martínez Vidal, J.L.; Garrido Frenich, A.; García, M.D.G. Determination of Cypermethrin, Fenvalerate and Cis- and Trans-Permethrin in Soil and Groundwater by High-Performance Liquid Chromatography Using Partial Least-Squares Regression. J. Chromatogr. A 1996, 727, 39–46. [Google Scholar] [CrossRef]
- Bronshtein, A.; Chuang, J.C.; Van Emon, J.M.; Altstein, M. Development of a Multianalyte Enzyme-Linked Immunosorbent Assay for Permethrin and Aroclors and Its Implementation for Analysis of Soil/Sediment and House Dust Extracts. J. Agric. Food Chem. 2012, 60, 4235–4242. [Google Scholar] [CrossRef]
- Chuang, J.C.; Van Emon, J.M.; Tefft, M.E.; Wilson, N.K. Application of a Permethrin Immunosorbent Assay Method to Residential Soil and Dust Samples. J. Environ. Sci. Health B 2010, 45, 516–523. [Google Scholar] [CrossRef]
- Bokade, P.; Gaur, V.K.; Tripathi, V.; Bobate, S.; Manickam, N.; Bajaj, A. Bacterial Remediation of Pesticide Polluted Soils: Exploring the Feasibility of Site Restoration. J. Hazard. Mater. 2023, 441, 129906. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-H.; Ji, G.-D. Stereoselective Degradation and Molecular Ecological Mechanism of Chiral Pesticides Beta-Cypermethrin in Soils with Different PH Values. Environ. Sci. Technol. 2015, 49, 14166–14175. [Google Scholar] [CrossRef] [PubMed]
- Tejada, M.; García, C.; Hernández, T.; Gómez, I. Response of Soil Microbial Activity and Biodiversity in Soils Polluted with Different Concentrations of Cypermethrin Insecticide. Arch. Environ. Contam. Toxicol. 2015, 69, 8–19. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Borowik, A.; Kucharski, J. The Role of Grass Compost and Zea Mays in Alleviating Toxic Effects of Tetracycline on the Soil Bacteria Community. Int. J. Environ. Res. Public Health 2022, 19, 7357. [Google Scholar] [CrossRef] [PubMed]
- Zaborowska, M.; Wyszkowska, J.; Borowik, A.; Kucharski, J. Effect of Separate and Combined Toxicity of Bisphenol A and Zinc on the Soil Microbiome. Int. J. Mol. Sci. 2022, 23, 5937. [Google Scholar] [CrossRef] [PubMed]
- Bashir, F.; Mahmooduzzafar; Siddiqi, T.O.; Iqbal, M. The Antioxidative Response System in Glycine Max (L.) Merr. Exposed to Deltamethrin, a Synthetic Pyrethroid Insecticide. Environ. Pollut. 2007, 147, 94–100. [Google Scholar] [CrossRef]
- Birolli, W.G.; da Silva, B.F.; Rodrigues Filho, E. Biodegradation of the Pyrethroid Cypermethrin by Bacterial Consortia Collected from Orange Crops. Environ. Res. 2022, 215, 114388. [Google Scholar] [CrossRef]
- Suresh Kumar, R.S.; Shiny, P.J.; Anjali, C.H.; Jerobin, J.; Goshen, K.M.; Magdassi, S.; Mukherjee, A.; Chandrasekaran, N. Distinctive Effects of Nano-Sized Permethrin in the Environment. Environ. Sci. Pollut. Res. 2013, 20, 2593–2602. [Google Scholar] [CrossRef]
- Duran, R.E.; Kilic, S.; Coskun, Y. Response of Maize (Zea Mays L. Saccharata Sturt) to Different Concentration Treatments of Deltamethrin. Pestic. Biochem. Phys. 2015, 124, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, Z.H.; Abbas, Z.K.; Ansari, A.A.; Khan, M.N.; Ansari, W.A. Pesticides and Their Effects on Plants: A Case Study of Deltamethrin. In Agrochemicals in Soil and Environment: Impacts and Remediation; Naeem, M., Bremont, J.F.J., Ansari, A.A., Gill, S.S., Eds.; Springer Nature: Singapore, 2022; pp. 183–193. ISBN 9789811693106. [Google Scholar]
- Pandey, J.K.; Dubey, G.; Gopal, R. Prolonged Use of Insecticide Dimethoate Inhibits Growth and Photosynthetic Activity of Wheat Seedlings: A Study by Laser-Induced Chlorophyll Fluorescence Spectroscopy. J. Fluoresc. 2022, 32, 2159–2172. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Khan, M.S. Ecotoxicological Implications of Residual Pesticides to Beneficial Soil Bacteria: A Review. Pestic. Biochem. Phys. 2022, 188, 105272. [Google Scholar] [CrossRef]
- Wołejko, E.; Jabłońska-Trypuć, A.; Wydro, U.; Butarewicz, A.; Łozowicka, B. Soil Biological Activity as an Indicator of Soil Pollution with Pesticides—A Review. Appl. Soil Ecol. 2020, 147, 103356. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Borowik, A.; Kucharski, M.; Kucharski, J. Effect of Cadmium, Copper and Zinc on Plants, Soil Microorganisms and Soil Enzymes. J. Elem. 2013, 18, 769–796. [Google Scholar] [CrossRef]
- Filimon, M.; Voia, S.; Popescu, R.; Dumitrescu, G.; Ciochină, L.; Mitulețu, M.; Vlad, D. The Effect of Some Insecticides on Soil Microorganisms Based on Enzymatic and Bacteriological Analyses. Rom. Biotechnol. Lett. 2015, 20, 10439–10447. [Google Scholar]
- Zhuang, R.; Chen, H.; Yao, J.; Li, Z.; Burnet, J.E.; Choi, M.M.F. Impact of Beta-Cypermethrin on Soil Microbial Community Associated with Its Bioavailability: A Combined Study by Isothermal Microcalorimetry and Enzyme Assay Techniques. J. Hazard. Mater. 2011, 189, 323–328. [Google Scholar] [CrossRef]
- Gianfreda, L.; Rao, M.A. Interactions Between Xenobiotics and Microbial and Enzymatic Soil Activity. Crit. Rev. Environ. Sci. Technol. 2008, 38, 269–310. [Google Scholar] [CrossRef]
- Muñoz-Leoz, B.; Garbisu, C.; Charcosset, J.-Y.; Sánchez-Pérez, J.M.; Antigüedad, I.; Ruiz-Romera, E. Non-Target Effects of Three Formulated Pesticides on Microbially-Mediated Processes in a Clay-Loam Soil. Sci. Total Environ. 2013, 449, 345–354. [Google Scholar] [CrossRef]
- Ismail, B.S.; Kalithasan, K. Adsorption, Desorption, and Mobility of Permethrin in Malaysian Soils. J. Environ. Sci. Health B 2004, 39, 419–429. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, L.; Li, F.; Xiao, M.; Lin, D.; Long, X.; Wu, Z. Microbial Degradation of Pesticide Residues and an Emphasis on the Degradation of Cypermethrin and 3-Phenoxy Benzoic Acid: A Review. Molecules 2018, 23, 2313. [Google Scholar] [CrossRef]
- Mandal, A.; Sarkar, B.; Mandal, S.; Vithanage, M.; Patra, A.K.; Manna, M.C. Impact of Agrochemicals on Soil Health. In Agrochemicals Detection, Treatment and Remediation; Prasad, M.N.V., Ed.; Butterworth-Heinemann: Oxford, UK, 2020; pp. 161–187. ISBN 978-0-08-103017-2. [Google Scholar]
- Das, R.; Das, S.J.; Das, A.C. Effect of Synthetic Pyrethroid Insecticides on N2-Fixation and Its Mineralization in Tea Soil. Eur. J. Soil Sci. 2016, 74, 9–15. [Google Scholar] [CrossRef]
- Cycoń, M.; Piotrowska-Seget, Z. Pyrethroid-Degrading Microorganisms and Their Potential for the Bioremediation of Contaminated Soils: A Review. Front. Microbiol. 2016, 7, 1463. [Google Scholar] [CrossRef]
- Kaviraj, A.; Gupta, A. Biomarkers of Type II Synthetic Pyrethroid Pesticides in Freshwater Fish. BioMed Res. Int. 2014, 2014, 928063. [Google Scholar] [CrossRef]
- Ray, P.; Lakshmanan, V.; Labbé, J.L.; Craven, K.D. Microbe to Microbiome: A Paradigm Shift in the Application of Microorganisms for Sustainable Agriculture. Front. Microbiol. 2020, 11, 622926. [Google Scholar] [CrossRef]
- Zhang, B.; Bai, Z.; Hoefel, D.; Tang, L.; Wang, X.; Li, B.; Li, Z.; Zhuang, G. The Impacts of Cypermethrin Pesticide Application on the Non-Target Microbial Community of the Pepper Plant Phyllosphere. Sci. Total Environ. 2009, 407, 1915–1922. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Kumar, P.S.; Vo, D.-V.N.; Rajamohan, N.; Saravanan, R. Microbial Degradation of Recalcitrant Pesticides: A Review. Environ. Chem. Lett. 2021, 19, 3209–3228. [Google Scholar] [CrossRef]
- Fang, Y.; Xu, W.; Zhang, W.; Guang, C.; Mu, W. Microbial Elimination of Pyrethroids: Specific Strains and Involved Enzymes. Appl. Microbiol. Biotechnol. 2022, 106, 6915–6932. [Google Scholar] [CrossRef]
- Bhatt, P.; Huang, Y.; Zhang, W.; Sharma, A.; Chen, S. Enhanced Cypermethrin Degradation Kinetics and Metabolic Pathway in Bacillus Thuringiensis Strain SG4. Microorganisms 2020, 8, 223. [Google Scholar] [CrossRef] [PubMed]
- Imade, E.E.; Babalola, O.O. Biotechnological Utilization: The Role of Zea Mays Rhizospheric Bacteria in Ecosystem Sustainability. Appl. Microbiol. Biotechnol. 2021, 105, 4487–4500. [Google Scholar] [CrossRef] [PubMed]
- Erguven, G.Ö.; Koçak, E. Determining the Detoxification Potential of Some Soil Bacteria and Plants on Bioremediation of Deltamethrin, Fenvalerate and Permethrin Pesticides. Eurasian J. Agric. Res. 2019, 3, 36–47. [Google Scholar]
- IUSS Working Group. WRB World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Wyszkowska, J.; Borowik, A.; Zaborowska, M.; Kucharski, J. Evaluation of the Usefulness of Sorbents in the Remediation of Soil Exposed to the Pressure of Cadmium and Cobalt. Materials 2022, 15, 5738. [Google Scholar] [CrossRef]
- Günal, A.Ç.; Tunca, S.K.; Arslan, P.; Gül, G.; Dinçel, A.S. How Does Sublethal Permethrin Effect Non-Target Aquatic Organisms? Environ. Sci. Pollut. Res. 2021, 28, 52405–52417. [Google Scholar] [CrossRef]
- Bulgarella, M.; Knutie, S.A.; Voss, M.A.; Cunninghame, F.; Florence-Bennett, B.J.; Robson, G.; Keyzers, R.A.; Taylor, L.M.; Lester, P.J.; Heimpel, G.E.; et al. Sub-Lethal Effects of Permethrin Exposure on a Passerine: Implications for Managing Ectoparasites in Wild Bird Nests. Conserv. Physiol. 2020, 8, coaa076. [Google Scholar] [CrossRef]
- Akbar, S.; Sultan, S.; Kertesz, M. Bacterial Community Analysis of Cypermethrin Enrichment Cultures and Bioremediation of Cypermethrin Contaminated Soils. J. Basic Microbiol. 2015, 55, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Morales-Máximo, C.N.; López-Sosa, L.B.; Rutiaga-Quiñones, J.G.; Corral-Huacuz, J.C.; Aguilera-Mandujano, A.; Pintor-Ibarra, L.F.; López-Miranda, A.; Delgado-Domínguez, S.N.; del Rodríguez-Magallón, M.C.; Morales-Máximo, M. Characterization of Agricultural Residues of Zea Mays for Their Application as Solid Biofuel: Case Study in San Francisco Pichátaro, Michoacán, Mexico. Energies 2022, 15, 6870. [Google Scholar] [CrossRef]
- OECD-FAO. OECD-FAO Agricultural Outlook 2021–2030; OECD Publishing: Paris, France, 2021; ISBN 978-92-64-43607-7. [Google Scholar]
- Erenstein, O.; Chamberlin, J.; Sonder, K. Estimating the Global Number and Distribution of Maize and Wheat Farms. Glob. Food Sec. 2021, 30, 100558. [Google Scholar] [CrossRef]
- Ten Berge, H.F.M.; Hijbeek, R.; van Loon, M.P.; Rurinda, J.; Tesfaye, K.; Zingore, S.; Craufurd, P.; van Heerwaarden, J.; Brentrup, F.; Schröder, J.J.; et al. Maize Crop Nutrient Input Requirements for Food Security in Sub-Saharan Africa. Glob. Food Sec. 2019, 23, 9–21. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Borowik, A.; Zaborowska, M.; Kucharski, J. Mitigation of the Adverse Impact of Copper, Nickel, and Zinc on Soil Microorganisms and Enzymes by Mineral Sorbents. Materials 2022, 15, 5198. [Google Scholar] [CrossRef]
- Bunt, J.S.; Rovira, A.D. Microbiological Studies of Some Subantarctic Soils. J. Soil Sci. 1955, 6, 119–128. [Google Scholar] [CrossRef]
- Parkinson, D.; Gray, T.R.G.; Williams, S.T. Methods for Studying the Ecology of Soil Microorganisms; IBP Handbook; Blackwell Scientific Publications [for the] International Biological Programme: Oxford, UK, 1971; ISBN 978-0-632-08260-5. [Google Scholar]
- Martin, J.P. Use of Acid, Rose Bengal, and Streptomycin in the Plate Method for Estimating Soil Fungi. Soil. Sci. 1950, 69, 215–232. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Wyszkowski, M. Resistance of Aerobic Microorganisms and Soil Enzyme Response to Soil Contamination with Ekodiesel Ultra Fuel. Environ. Sci. Pollut. Res. 2017, 24, 24346–24363. [Google Scholar] [CrossRef] [PubMed]
- Zaborowska, M.; Wyszkowska, J.; Kucharski, J. Role of Chlorella Sp. and Rhamnolipid 90 in Maintaining Homeostasis in Soil Contaminated with Bisphenol A. J. Soils Sediments 2021, 21, 27–41. [Google Scholar] [CrossRef]
- Zaborowska, M.; Wyszkowska, J.; Borowik, A.; Kucharski, J. Bisphenol A—A Dangerous Pollutant Distorting the Biological Properties of Soil. Int. J. Mol. Sci. 2021, 22, 12753. [Google Scholar] [CrossRef] [PubMed]
- De Leij, F.A.A.M.; Whipps, J.M.; Lynch, J.M. The Use of Colony Development for the Characterization of Bacterial Communities in Soil and on Roots. Microb. Ecol. 1993, 27, 81–97. [Google Scholar] [CrossRef] [PubMed]
- TIBCO Software Inc Statistica (Data Analysis Software System), Version 13. 2017. Available online: https://www.tibco.com/products/data-science (accessed on 8 December 2022).
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical Analysis of Taxonomic and Functional Profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R. RStudio, Inc., Boston, MA, USA. 2019. Available online: Http://Www.Rstudio.com/ (accessed on 8 December 2022).
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, M.; Moeller, S.; et al. Gplots: Various R Programming Tools for Plotting Data. R Package Version 2.17.0. 2020. Available online: https://CRAN.R-Project.Org/Package=gplots (accessed on 8 December 2022).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; Vienna, Austria. 2019. Available online: https://www.R-project.org (accessed on 8 December 2022).
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A Web-Based Tool for the Analysis of Sets through Venn Diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for Inferring Very Large Phylogenies by Using the Neighbor-joining Method. Proc. Nat. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Molec. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Object | Aerial Parts | Roots |
|---|---|---|
| ZmC | 53.411 a | 6.176 a |
| ZmPr | 33.173 c | 4.082 c |
| ZmCp | 44.411 b | 5.913 b |
| Object | Development Phase | |
|---|---|---|
| Four Leaves | Six Leaves | |
| ZmC | 45.313 b | 39.681 a |
| ZmPr | 44.081 c | 37.644 ab |
| ZmCp | 46.981 a | 36.628 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borowik, A.; Wyszkowska, J.; Zaborowska, M.; Kucharski, J. The Impact of Permethrin and Cypermethrin on Plants, Soil Enzyme Activity, and Microbial Communities. Int. J. Mol. Sci. 2023, 24, 2892. https://doi.org/10.3390/ijms24032892
Borowik A, Wyszkowska J, Zaborowska M, Kucharski J. The Impact of Permethrin and Cypermethrin on Plants, Soil Enzyme Activity, and Microbial Communities. International Journal of Molecular Sciences. 2023; 24(3):2892. https://doi.org/10.3390/ijms24032892
Chicago/Turabian StyleBorowik, Agata, Jadwiga Wyszkowska, Magdalena Zaborowska, and Jan Kucharski. 2023. "The Impact of Permethrin and Cypermethrin on Plants, Soil Enzyme Activity, and Microbial Communities" International Journal of Molecular Sciences 24, no. 3: 2892. https://doi.org/10.3390/ijms24032892
APA StyleBorowik, A., Wyszkowska, J., Zaborowska, M., & Kucharski, J. (2023). The Impact of Permethrin and Cypermethrin on Plants, Soil Enzyme Activity, and Microbial Communities. International Journal of Molecular Sciences, 24(3), 2892. https://doi.org/10.3390/ijms24032892