
Differential Modulation of Human M1 and M2 Macrophage Activity by ICOS-Mediated ICOSL Triggering

 ,
,  , , , , ,
, , , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
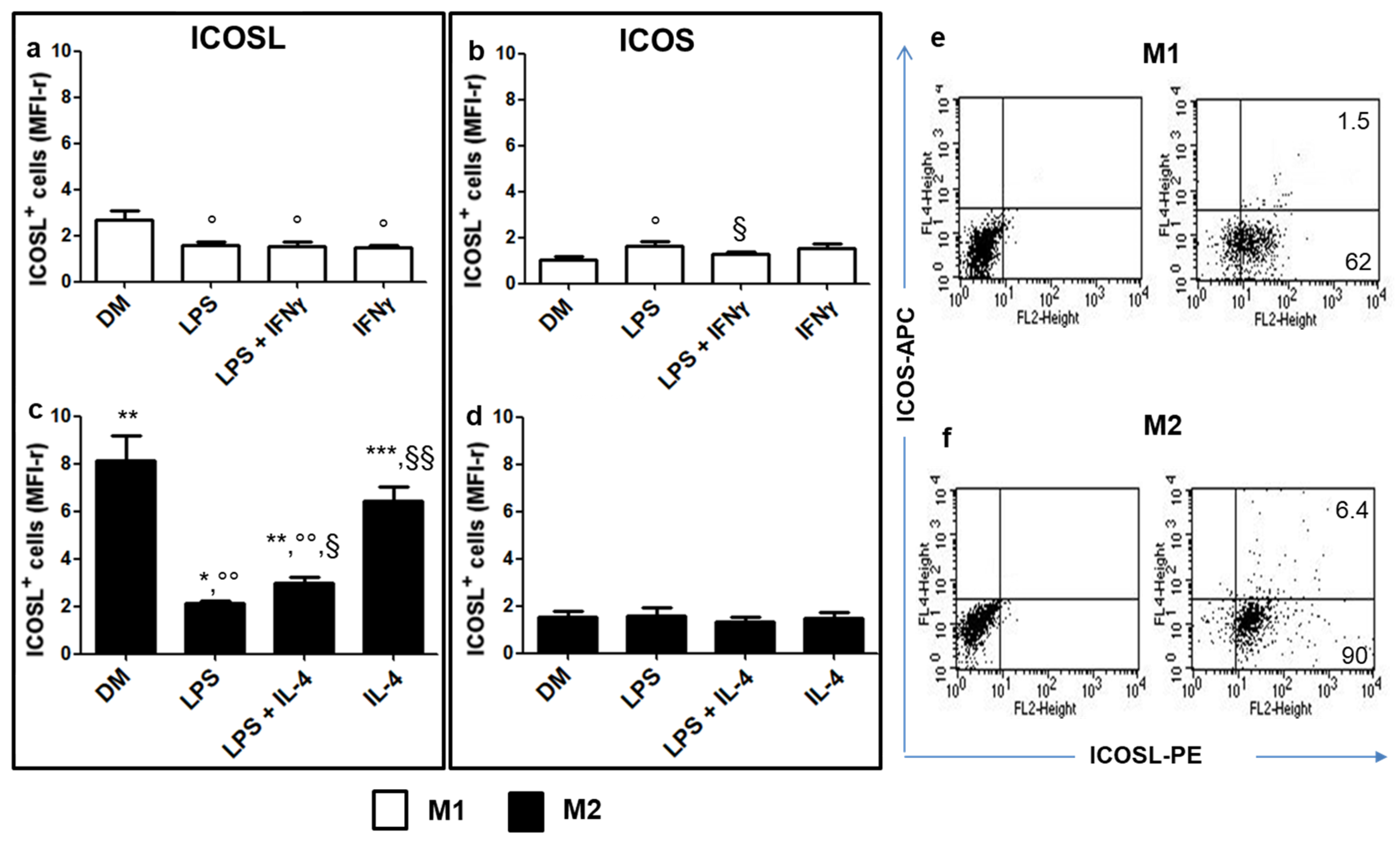
2.1. Analysis of ICOSL and ICOS Expression in M1 vs. M2 Cells
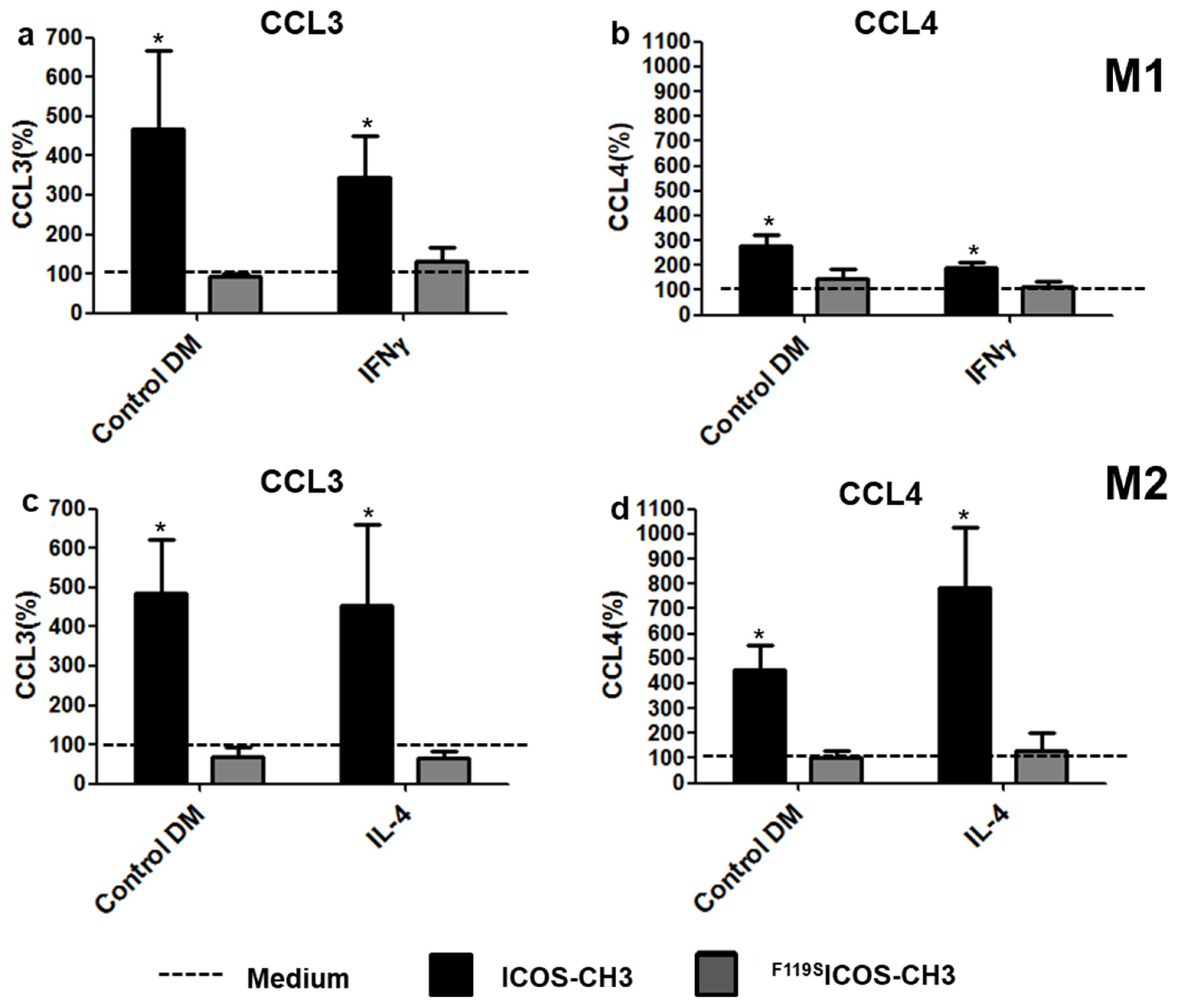
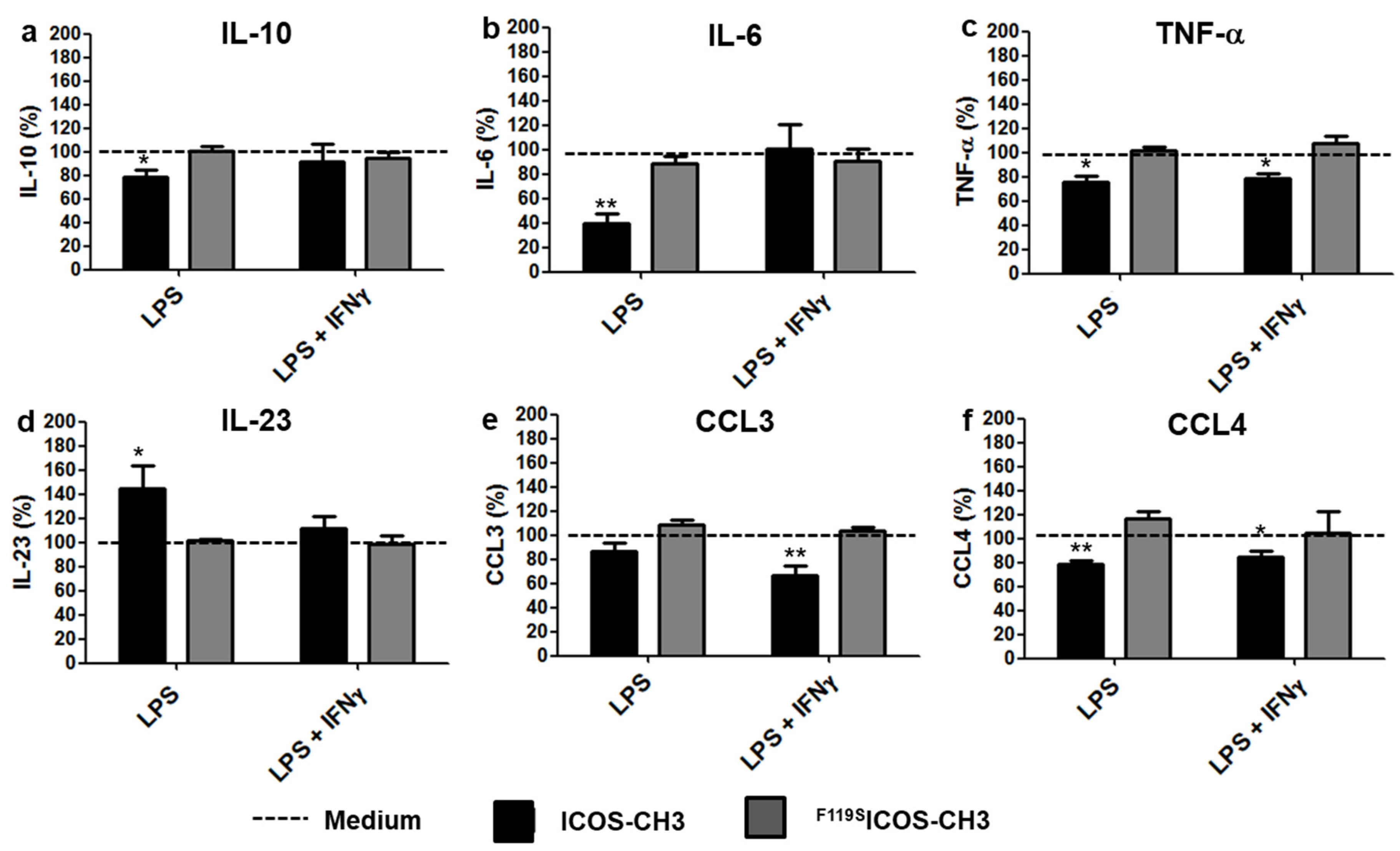
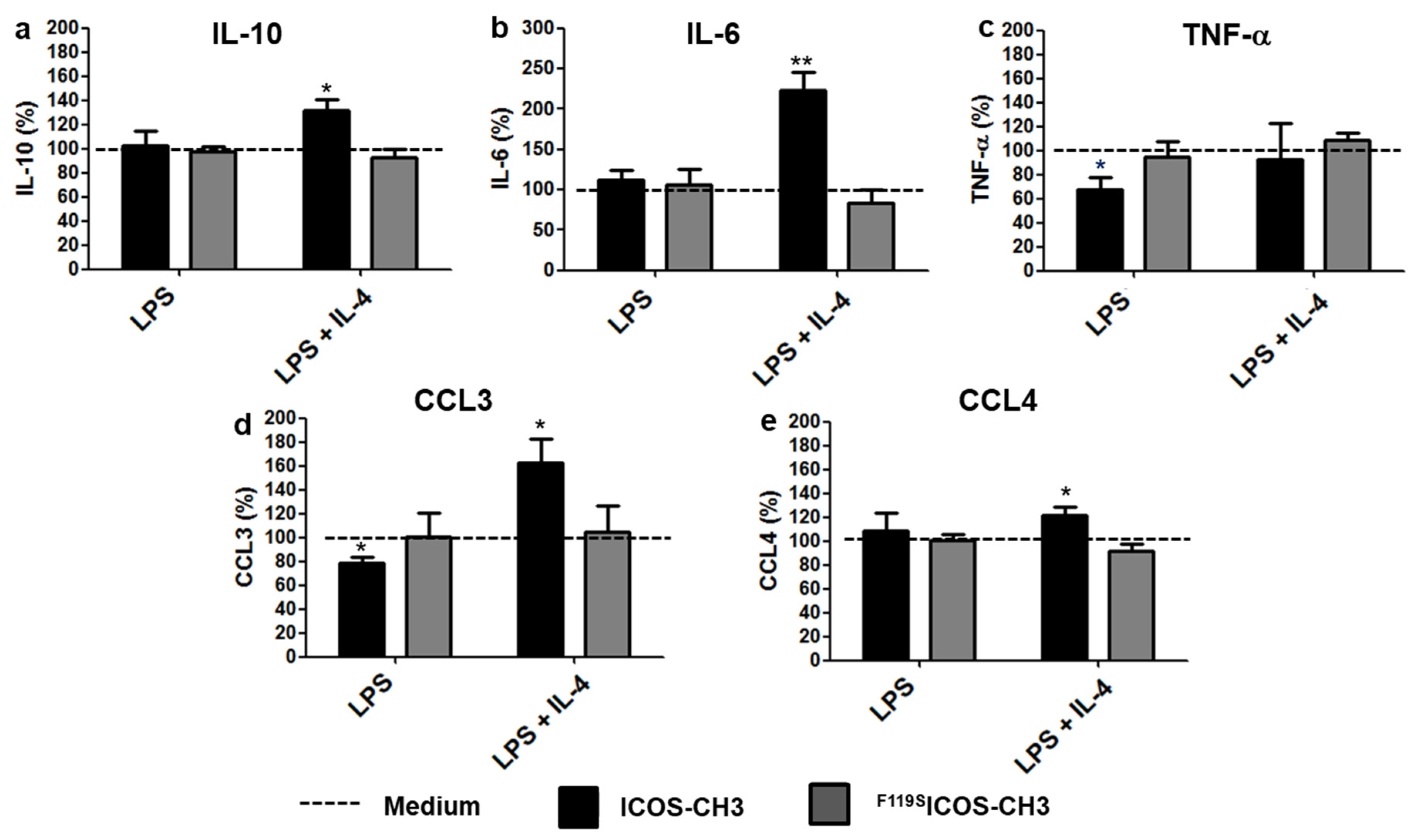
2.2. Effects of ICOSL Triggering on Cytokine Secretion
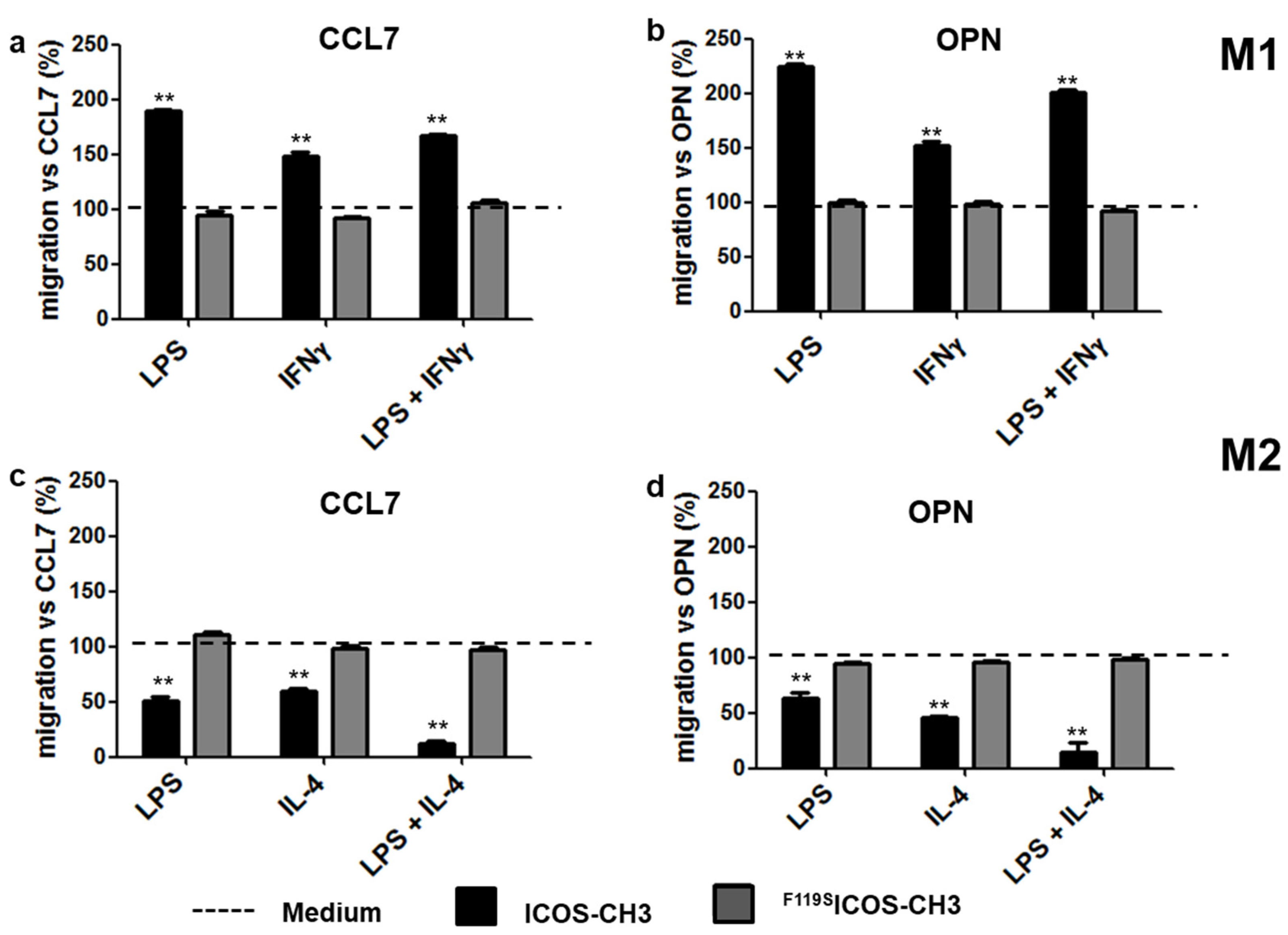
2.3. Effect of ICOSL Stimulation on Cell Migration
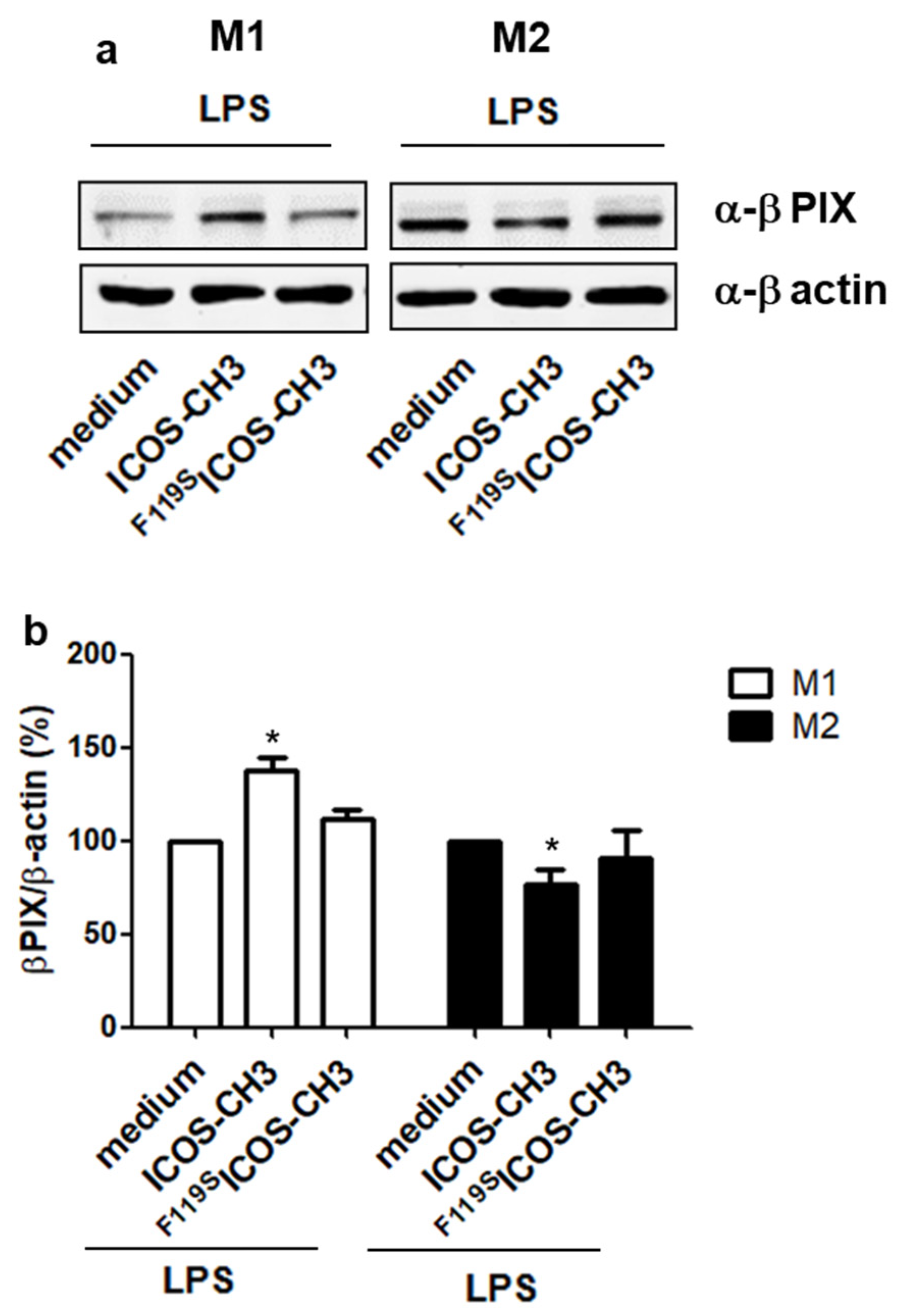
2.4. Effect of ICOSL Stimulation on Downstream Signaling Pathways
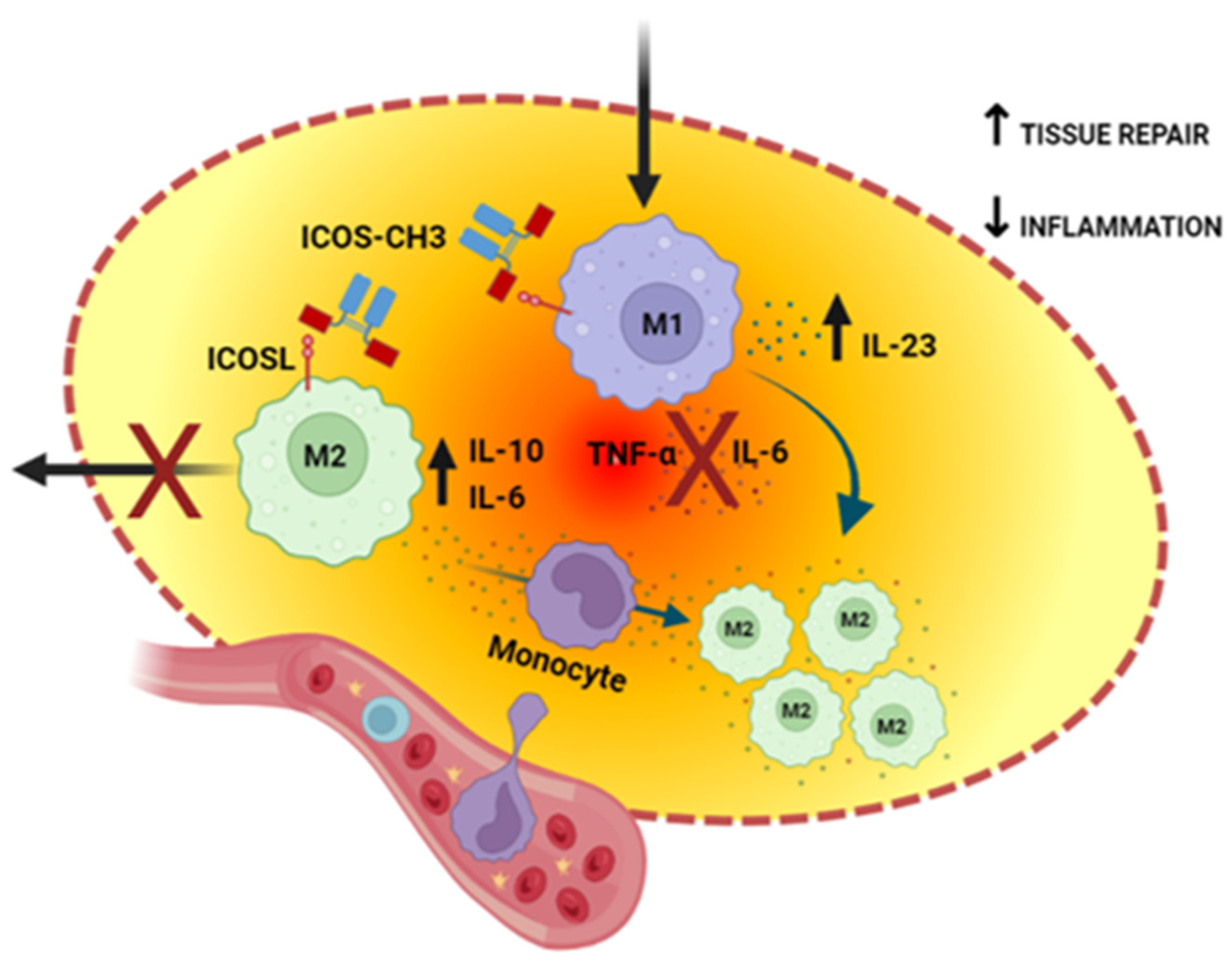
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Immunofluorescence
4.3. Cytokine and Chemokine Analysis
4.4. Migration Assay
4.5. Immunoblotting
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hedl, M.; Lahiri, A.; Ning, K.; Cho, J.H.; Abraham, C. Pattern recognition receptor signaling in human dendritic cells is enhanced by ICOS ligand and modulated by the Crohn’s disease ICOSLG risk allele. Immunity 2014, 40, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, A.H.; Freeman, G.J. The B7-CD28 superfamily. Nat. Rev. Immunol. 2003, 2, 116–126. [Google Scholar] [CrossRef]
- Nurieva, R.I. Regulation of immune and autoimmune responses by ICOS-B7h interaction. Clin. Immunol. 2005, 115, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Bauquet, A.T.; Jin, H.; Paterson, A.M.; Mitsdoerffer, M.; Ho, I.C.; Sharpe, A.H.; Kuchroo, V.K. The costimulatory molecule ICOS regulates the expression of c-Maf and IL-21 in the development of follicular T helper cells and TH-17 cells. Nat. Immunol. 2009, 10, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Yong, P.F.; Salzer, U.; Grimbacher, B. The role of costimulation in antibody deficiencies: ICOS and common variable immunodeficiency. Immunol. Rev. 2009, 229, 101–113. [Google Scholar] [CrossRef]
- Mesturini, R.; Nicola, S.; Chiocchetti, A.; Bernardone, I.S.; Castelli, L.; Bensi, T.; Ferretti, M.; Comi, C.; Dong, C.; Rojo, J.M.; et al. ICOS cooperates with CD28, IL-2, and IFN-gamma and modulates activation of human naïve CD4+ T cells. Eur. J. Immunol. 2006, 36, 2601–2612. [Google Scholar] [CrossRef]
- Mesturini, R.; Gigliotti, C.L.; Orilieri, E.; Cappellano, G.; Soluri, M.F.; Boggio, E.; Woldetsadik, A.; Dianzani, C.; Sblattero, D.; Chiocchetti, A.; et al. Differential induction of IL-17, IL-10, and IL-9 in human T helper cells by B7h and B7.1. Cytokine 2013, 64, 322–330. [Google Scholar] [CrossRef]
- Tang, G.; Qin, Q.; Zhang, P.; Wang, G.; Liu, M.; Ding, Q.; Qin, Y.; Shen, Q. Reverse signaling using an inducible costimulator to enhance immunogenic function of dendritic cells. Cell. Mol. Life Sci. 2008, 66, 3067–3080. [Google Scholar] [CrossRef]
- Dianzani, C.; Minelli, R.; Mesturini, R.; Chiocchetti, A.; Barrera, G.; Boscolo, S.; Sarasso, C.; Gigliotti, C.L.; Sblattero, D.; Yagi, J.; et al. B7h triggering inhibits umbilical vascular endothelial cell adhesiveness to tumor cell lines and polymorphonuclear cells. J. Immunol. 2010, 185, 3970–3979. [Google Scholar] [CrossRef]
- Dianzani, C.; Minelli, R.; Gigliotti, C.L.; Occhipinti, S.; Giovarelli, M.; Conti, L.; Boggio, E.; Shivakumar, Y.; Baldanzi, G.; Malacarne, V.; et al. B7h triggering inhibits the migration of tumor cell lines. J. Immunol. 2014, 192, 4921–4931. [Google Scholar] [CrossRef] [Green Version]
- Occhipinti, S.; Dianzani, C.; Chiocchetti, A.; Boggio, E.; Clemente, N.; Gigliotti, C.L.; Soluri, M.F.; Minelli, R.; Fantozzi, R.; Yagi, J.; et al. Triggering of B7h by the inducible costimulatory modulates maturation and migration of monocyte-derived dendritic cells. J. Immunol. 2013, 190, 1125–1134. [Google Scholar] [CrossRef]
- Gigliotti, C.L.; Boggio, E.; Clemente, N.; Shivakumar, Y.; Toth, E.; Sblattero, D.; D’Amelio, P.; Isaia, G.C.; Dianzani, C.; Yagi, J.; et al. ICOS-Ligand triggering impairs osteoclast differentiation and function in vitro and in vivo. J. Immunol. 2016, 97, 3905–3916. [Google Scholar] [CrossRef] [PubMed]
- Clemente, N.; Boggio, E.; Gigliotti, L.C.; Raineri, D.; Ferrara, B.; Miglio, G.; Argenziano, M.; Chiocchetti, A.; Cappellano, G.; Trotta, F.; et al. Immunotherapy of experimental melanoma with ICOS-Fc loaded in biocompatible and biodegradable nanoparticles. J. Control. Release 2020, 320, 112–124. [Google Scholar] [CrossRef]
- Boggio, E.; Gigliotti, C.L.; Moia, R.; Scotta, A.; Crespi, I.; Boggione, P.; De Paoli, L.; Deambrogi, C.; Garzaro, M.; Vidali, M.; et al. Inducible T-cell co-stimulator (ICOS) and ICOS ligand are novel players in the multiple-myeloma microenvironment. Br. J. Haematol. 2022, 196, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Raineri, D.; Dianzani, C.; Cappellano, G.; Maione, F.; Baldanzi, G.; Iacobucci, I.; Clemente, N.; Baldone, G.; Boggio, E.; Gigliotti, C.L.; et al. Osteopontin binds ICOSL promoting tumor metastasis. Commun. Biol. 2020, 3, 615. [Google Scholar] [CrossRef]
- Stoppa, I.; Gigliotti, C.L.; Clemente, N.; Pantham, D.; Dianzani, C.; Monge, C.; Puricelli, C.; Rolla, R.; Sutti, S.; Renò, F.; et al. ICOSL Stimulation by ICOS-Fc Accelerates Cutaneous Wound Healing In Vivo. Int. J. Mol. Sci. 2022, 23, 7363. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Jaguin, M.; Houlbert, N.; Fardel, O.; Lecureur, V. Polarization profiles of human M-CSF-generated macrophages and comparison of M1-markers in classically activated macrophages from GM-CSF and M-CSF origin. Cell. Immunol. 2013, 281, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Kittan, N.A.; Allen, R.M.; Dhaliwal, A.; Cavassani, K.A.; Schaller, M.; Gallagher, K.A.; Carson, W.F., 4th; Mukherjee, S.; Grembecka, J.; Cierpicki, T.; et al. Cytokine induced phenotypic and epigenetic signatures are key to establishing specific macrophage phenotypes. PLoS ONE 2013, 8, e78045. [Google Scholar] [CrossRef]
- Ramavath, N.N.; Gadipudi, L.L.; Provera, A.; Gigliotti, L.C.; Boggio, E.; Bozzola, C.; Albano, E.; Dianzani, U.; Sutti, S. Inducible T-cell costimulator mediates lymphocyte/macrophage interactions during liver repair. Front. Immunol. 2021, 12, 786680. [Google Scholar] [CrossRef]
- Gigliotti, C.L.; Boggio, E.; Dianzani, U. Department of Health Sciences, Università del Piemonte Orientale, Novara, Italy, 2022. unpublished work.
- Morrison, P.J.; Ballantyne, S.J.; Kullberg, M.C. Interleukin-23 and T helper 17-type responses in intestinal inflammation: From cytokines to T-cell plasticity. Immunology 2011, 133, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Lauridsen, H.M.; Amezquita, R.A.; Pierce, R.W.; Jane-Wit, D.; Fang, C.; Pellowe, A.S.; Kirkiles-Smith, N.C.; Gonzalez, A.L.; Pober, J.S. IL-17 Promotes neutrophil-mediated immunity by activating microvascular pericytes and not endothelium. J. Immunol. 2016, 197, 2400–2408. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef]
- Norris, C.A.; He, M.; Kang, L.I.; Ding, M.Q.; Radder, J.E.; Haynes, M.M.; Yang, Y.; Paranjpe, S.; Bowen, W.C.; Orr, A.; et al. Synthesis of IL-6 by hepatocytes is a normal response to common hepatic stimuli. PLoS ONE 2014, 9, e96053. [Google Scholar] [CrossRef] [PubMed]
- Ferreira Alves, G.; Stoppa, I.; Aimaretti, E.; Monge, C.; Mastrocola, R.; Porchietto, E.; Einaudi, G.; Collotta, D.; Bertocchi, I.; Boggio, E.; et al. ICOS-Fc as innovative immunomodulatory approach to counteract inflammation and organ injury in sepsis. Front. Immunol. 2022, 13, 992614. [Google Scholar]
- Maeda, S.; Fujimoto, M.; Matsushita, T.; Hamaguchi, Y.; Takehara, K.; Hasegawa, M. Inducible costimulator (ICOS) and ICOS ligand signaling has pivotal roles in skin wound healing via cytokine production. Am. J. Pathol. 2011, 179, 2360–2369. [Google Scholar] [CrossRef]
- Cui, K.; Ardell, C.L.; Podolnikova, N.P.; Yakubenko, V.P. Distinct migratory properties of M1, M2, and resident macrophages are regulated by αDβ2 and αMβ2 integrin-mediated adhesion. Front. Immunol. 2018, 9, 2650. [Google Scholar] [CrossRef]
- Vogel, D.Y.; Heijnen, P.D.; Breur, M.; de Vries, H.E.; Tool, A.T.; Amor, S.; Dijkstra, C.D. Macrophages migrate in an activation-dependent manner to chemokines involved in neuroinflammation. J. Neuroinflamm. 2014, 11, 23. [Google Scholar] [CrossRef]
- Bordano, V.; Kinsella, G.K.; Cannito, S.; Dianzani, C.; Gigliotti, C.L.; Stephens, J.C.; Monge, C.; Bocca, C.; Rosa, A.C.; Miglio, G.; et al. G protein-coupled receptor 21 in macrophages: An in vitro study. Eur. J. Pharmacol. 2022, 926, 175018. [Google Scholar] [CrossRef]
- Arimura, Y.; Kato, H.; Dianzani, U.; Okamoto, T.; Kamekura, S.; Buonfiglio, D.; Miyoshi-Akiyama, T.; Uchiyama, T.; Yagi, J. A co-stimulatory molecule on activated T cells, H4/ICOS, delivers specific signals in T(h) cells and regulates their responses. Int. Immunol. 2002, 14, 555–566. [Google Scholar] [CrossRef]
- Roovers, R.C.; van der Linden, E.; de Bruïne, A.P.; Arends, J.W.; Hoogenboom, H.R. In vitro characterisation of a monovalent and bivalent form of a fully human anti Ep-CAM phage antibody. Cancer Immunol. Immunother. 2001, 50, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Boggio, E.; Dianzani, C.; Gigliotti, C.L.; Soluri, M.F.; Clemente, N.; Cappellano, G.; Toth, E.; Raineri, D.; Ferrara, B.; Comi, C.; et al. Thrombin cleavage of osteopontin modulates its activities in human cells in vitro and mouse experimental autoimmune encephalomyelitis in vivo. J. Immunol. Res. 2016, 2016, 9345495. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gigliotti, C.L.; Dianzani, C.; Stoppa, I.; Monge, C.; Sutti, S.; Sblattero, D.; Puricelli, C.; Rolla, R.; Dianzani, U.; Boggio, E. Differential Modulation of Human M1 and M2 Macrophage Activity by ICOS-Mediated ICOSL Triggering. Int. J. Mol. Sci. 2023, 24, 2953. https://doi.org/10.3390/ijms24032953
Gigliotti CL, Dianzani C, Stoppa I, Monge C, Sutti S, Sblattero D, Puricelli C, Rolla R, Dianzani U, Boggio E. Differential Modulation of Human M1 and M2 Macrophage Activity by ICOS-Mediated ICOSL Triggering. International Journal of Molecular Sciences. 2023; 24(3):2953. https://doi.org/10.3390/ijms24032953
Chicago/Turabian StyleGigliotti, Casimiro Luca, Chiara Dianzani, Ian Stoppa, Chiara Monge, Salvatore Sutti, Daniele Sblattero, Chiara Puricelli, Roberta Rolla, Umberto Dianzani, and Elena Boggio. 2023. "Differential Modulation of Human M1 and M2 Macrophage Activity by ICOS-Mediated ICOSL Triggering" International Journal of Molecular Sciences 24, no. 3: 2953. https://doi.org/10.3390/ijms24032953
APA StyleGigliotti, C. L., Dianzani, C., Stoppa, I., Monge, C., Sutti, S., Sblattero, D., Puricelli, C., Rolla, R., Dianzani, U., & Boggio, E. (2023). Differential Modulation of Human M1 and M2 Macrophage Activity by ICOS-Mediated ICOSL Triggering. International Journal of Molecular Sciences, 24(3), 2953. https://doi.org/10.3390/ijms24032953