
Effects of Positive Fighting Experience and Its Subsequent Deprivation on the Expression Profile of Mouse Hippocampal Genes Associated with Neurogenesis

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
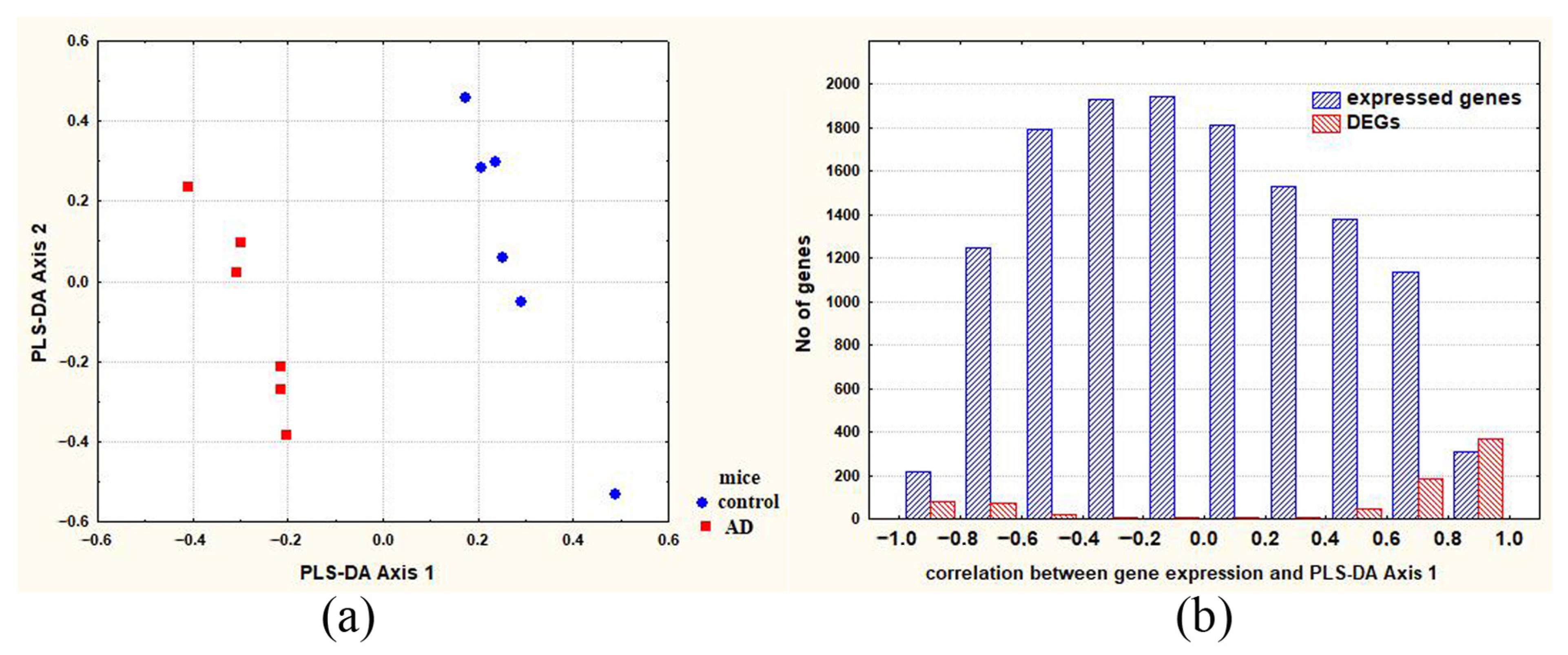
2.1. Differences between Groups of Control and Experimental Mice
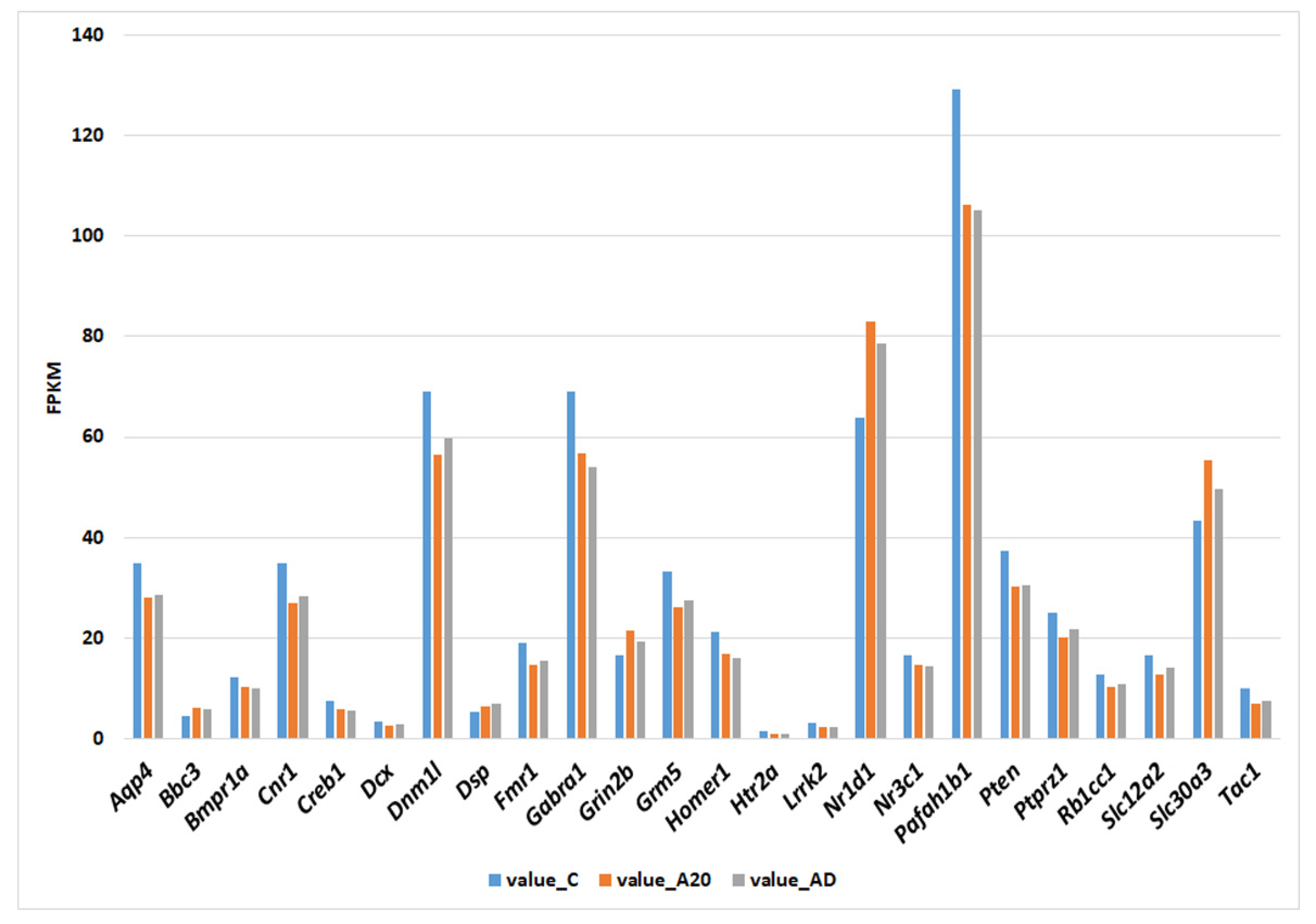
2.2. C_A20 Differentially Expressed Genes (DEGs) Associated with Neurogenesis
Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis of C_A20 DEGs Associated with Neurogenesis
2.3. DEGs Associated with Neurogenesis and Making the Most Significant Contribution to Intergroup Differences (Comparison C_A20)
2.4. C_AD DEGs Associated with Neurogenesis
2.4.1. KEGG Analysis of C_AD DEGs Associated with Neurogenesis
2.4.2. Changes in C_AD DEGs’ Transcription during the Fighting Deprivation as Compared with the Period of Agonistic Interactions
2.4.3. DEGs Associated with Neurogenesis and Making the Most Significant Contribution to C_AD Differences
2.5. Identification of New Candidate Genes Related to Behavior
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Creation of the Repeated Positive Fighting Experience for Male Mice by Agonistic Interactions
4.3. RNA-Seq Data Collection and Processing
4.4. Functional Annotation of the DEGs
4.5. Statistical Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balu, D.T.; Lucki, I. Adult hippocampal neurogenesis: Regulation, functional implications, and contribution to disease pathology. Neurosci. Biobehav. Rev. 2009, 33, 232–252. [Google Scholar] [CrossRef] [PubMed]
- Nakafuku, M.; Del Aguila, A. Developmental dynamics of neurogenesis and gliogenesis in the postnatal mammalian brain in health and disease: Historical and future perspectives. Wiley Interdiscip. Rev. Dev. Biol. 2020, 9, e369. [Google Scholar] [CrossRef] [PubMed]
- Carli, M.; Aringhieri, S.; Kolachalam, S.; Longoni, B.; Grenno, G.; Rossi, M.; Gemignani, A.; Fornai, F.; Maggio, R.; Scarselli, M. Is Adult Hippocampal Neurogenesis Really Relevant for the Treatment of Psychiatric Disorders? Curr. Neuropharmacol. 2021, 19, 1640–1660. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Jimenez, E.P.; Terreros-Roncal, J.; Flor-Garcia, M.; Rabano, A.; Llorens-Martin, M. Evidences for Adult Hippocampal Neurogenesis in Humans. J. Neurosci. 2021, 41, 2541–2553. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.; Eisch, A.J. Epigenetics, hippocampal neurogenesis, and neuropsychiatric disorders: Unraveling the genome to understand the mind. Neurobiol. Dis. 2010, 39, 73–84. [Google Scholar] [CrossRef]
- Snyder, J.S.; Drew, M.R. Functional neurogenesis over the years. Behav. Brain Res. 2020, 382, 112470. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Gage, F.H. Adult hippocampal neurogenesis and its role in Alzheimer’s disease. Mol. Neurodegener. 2011, 6, 85. [Google Scholar] [CrossRef]
- Maruszak, A.; Pilarski, A.; Murphy, T.; Branch, N.; Thuret, S. Hippocampal neurogenesis in Alzheimer’s disease: Is there a role for dietary modulation? J. Alzheimers Dis. 2014, 38, 11–38. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L.; Zusso, M.; Giusti, P. Synaptic Plasticity, Dementia and Alzheimer Disease. CNS Neurol. Disord. Drug Targets 2017, 16, 220–233. [Google Scholar] [CrossRef]
- Teixeira, C.M.; Pallas-Bazarra, N.; Bolos, M.; Terreros-Roncal, J.; Avila, J.; Llorens-Martin, M. Untold New Beginnings: Adult Hippocampal Neurogenesis and Alzheimer’s Disease. J. Alzheimers Dis. 2018, 64, S497–S505. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Ma, Y. Molecular mechanisms of altered adult hippocampal neurogenesis in Alzheimer’s disease. Mech. Ageing Dev. 2021, 195, 111452. [Google Scholar] [CrossRef]
- Babcock, K.R.; Page, J.S.; Fallon, J.R.; Webb, A.E. Adult Hippocampal Neurogenesis in Aging and Alzheimer’s Disease. Stem Cell Rep. 2021, 16, 681–693. [Google Scholar] [CrossRef]
- Goncalves, V.F.; Andreazza, A.C.; Kennedy, J.L. Mitochondrial dysfunction in schizophrenia: An evolutionary perspective. Hum. Genet. 2015, 134, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, F.; Wu, Y.; Zhou, B. New Insights Into the Role of Aberrant Hippocampal Neurogenesis in Epilepsy. Front. Neurol. 2021, 12, 727065. [Google Scholar] [CrossRef]
- Willsey, H.R.; Willsey, A.J.; Wang, B.; State, M.W. Genomics, convergent neuroscience and progress in understanding autism spectrum disorder. Nat. Rev. Neurosci. 2022, 23, 323–341. [Google Scholar] [CrossRef]
- Marxreiter, F.; Regensburger, M.; Winkler, J. Adult neurogenesis in Parkinson’s disease. Cell. Mol. Life Sci. 2013, 70, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, B.; Tirolo, C.; L’Episcopo, F.; Caniglia, S.; Testa, N.; Smith, J.A.; Pluchino, S.; Serapide, M.F. Parkinson’s disease, aging and adult neurogenesis: Wnt/beta-catenin signalling as the key to unlock the mystery of endogenous brain repair. Aging Cell 2020, 19, e13101. [Google Scholar] [CrossRef]
- Paizanis, E.; Hamon, M.; Lanfumey, L. Hippocampal neurogenesis, depressive disorders, and antidepressant therapy. Neural Plast. 2007, 2007, 73754. [Google Scholar] [CrossRef]
- Eisch, A.J.; Petrik, D. Depression and hippocampal neurogenesis: A road to remission? Science 2012, 338, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.S.; Sahay, A.; Hen, R. Increasing Adult Hippocampal Neurogenesis is Sufficient to Reduce Anxiety and Depression-Like Behaviors. Neuropsychopharmacology 2015, 40, 2368–2378. [Google Scholar] [CrossRef] [Green Version]
- Boku, S.; Nakagawa, S.; Toda, H.; Hishimoto, A. Neural basis of major depressive disorder: Beyond monoamine hypothesis. Psychiatry Clin. Neurosci. 2018, 72, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The role of adult hippocampal neurogenesis in brain health and disease. Mol. Psychiatry 2019, 24, 67–87. [Google Scholar] [CrossRef]
- Rocca, M.A.; Barkhof, F.; De Luca, J.; Frisen, J.; Geurts, J.J.G.; Hulst, H.E.; Sastre-Garriga, J.; Filippi, M.; Group, M.S. The hippocampus in multiple sclerosis. Lancet Neurol. 2018, 17, 918–926. [Google Scholar] [CrossRef]
- Jorgensen, C. Adult mammalian neurogenesis and motivated behaviors. Integr. Zool. 2018, 13, 655–672. [Google Scholar] [CrossRef] [PubMed]
- Udo, H.; Yoshida, Y.; Kino, T.; Ohnuki, K.; Mizunoya, W.; Mukuda, T.; Sugiyama, H. Enhanced adult neurogenesis and angiogenesis and altered affective behaviors in mice overexpressing vascular endothelial growth factor 120. J. Neurosci. 2008, 28, 14522–14536. [Google Scholar] [CrossRef]
- Smagin, D.A.; Park, J.H.; Michurina, T.V.; Peunova, N.; Glass, Z.; Sayed, K.; Bondar, N.P.; Kovalenko, I.N.; Kudryavtseva, N.N.; Enikolopov, G. Altered Hippocampal Neurogenesis and Amygdalar Neuronal Activity in Adult Mice with Repeated Experience of Aggression. Front. Neurosci. 2015, 9, 443. [Google Scholar] [CrossRef]
- Mitra, R.; Sundlass, K.; Parker, K.J.; Schatzberg, A.F.; Lyons, D.M. Social stress-related behavior affects hippocampal cell proliferation in mice. Physiol. Behav. 2006, 89, 123–127. [Google Scholar] [CrossRef]
- Leschik, J.; Lutz, B.; Gentile, A. Stress-Related Dysfunction of Adult Hippocampal Neurogenesis-An Attempt for Understanding Resilience? Int. J. Mol. Sci. 2021, 22, 7339. [Google Scholar] [CrossRef]
- Ihunwo, A.O.; Tembo, L.H.; Dzamalala, C. The dynamics of adult neurogenesis in human hippocampus. Neural Regen. Res. 2016, 11, 1869–1883. [Google Scholar] [CrossRef] [PubMed]
- Vivar, C.; van Praag, H. Running Changes the Brain: The Long and the Short of It. Physiology (Bethesda) 2017, 32, 410–424. [Google Scholar] [CrossRef]
- Accogli, A.; Addour-Boudrahem, N.; Srour, M. Neurogenesis, neuronal migration, and axon guidance. Handb. Clin. Neurol. 2020, 173, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Overall, R.W.; Paszkowski-Rogacz, M.; Kempermann, G. The mammalian adult neurogenesis gene ontology (MANGO) provides a structural framework for published information on genes regulating adult hippocampal neurogenesis. PLoS ONE 2012, 7, e48527. [Google Scholar] [CrossRef]
- Kudryavtseva, N.N. Positive fighting experience, addiction-like state, and relapse: Retrospective analysis of experimental studies. Aggress. Viol. Behav. 2020, 52, 101403. [Google Scholar] [CrossRef]
- Smagin, D.A.; Galyamina, A.G.; Kovalenko, I.L.; Kudryavtseva, N.N. Altered Expression of Genes Associated with Major Neurotransmitter Systems in the Reward-Related Brain Regions of Mice with Positive Fighting Experience. Int. J. Mol. Sci. 2022, 23, 13644. [Google Scholar] [CrossRef]
- Kudryavtseva, N.N.; Smagin, D.A.; Kovalenko, I.L.; Vishnivetskaya, G.B. Repeated positive fighting experience in male inbred mice. Nat. Protoc. 2014, 9, 2705–2717. [Google Scholar] [CrossRef] [PubMed]
- Smagin, D.A.; Galyamina, A.G.; Kovalenko, I.L.; Babenko, V.N.; Kudryavtseva, N.N. Aberrant Expression of Collagen Gene Family in the Brain Regions of Male Mice with Behavioral Psychopathologies Induced by Chronic Agonistic Interactions. Biomed. Res. Int. 2019, 2019, 7276389. [Google Scholar] [CrossRef]
- Kudryavtseva, N.N. Lorenz was right! Or does aggressive energy accumulate? Russ. J. Genet. 2004, 40, 656–662. [Google Scholar] [CrossRef]
- Kudryavtseva, N.N. Straub tail, the deprivation effect and addiction to aggression. In Motivation of Health Behavior; O’Neal, P.W., Ed.; NOVA Science Publishers: New York, NY, USA, 2007; pp. 97–110. [Google Scholar]
- Kudryavtseva, N.N.; Smagin, D.A.; Bondar, N.P. Modeling fighting deprivation effect in mouse repeated aggression paradigm. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1472–1478. [Google Scholar] [CrossRef]
- Babenko, V.; Redina, O.; Smagin, D.; Kovalenko, I.; Galyamina, A.; Babenko, R.; Kudryavtseva, N. Dorsal Striatum Transcriptome Profile Profound Shift in Repeated Aggression Mouse Model Converged to Networks of 12 Transcription Factors after Fighting Deprivation. Genes (Basel) 2021, 13, 21. [Google Scholar] [CrossRef]
- Kudryavtseva, N.N.; Bondar, N.P.; Avgustinovich, D.F. Association between experience of aggression and anxiety in male mice. Behav. Brain Res. 2002, 133, 83–93. [Google Scholar] [CrossRef]
- Griesauer, I.; Diao, W.; Ronovsky, M.; Elbau, I.; Sartori, S.; Singewald, N.; Pollak, D.D. Circadian abnormalities in a mouse model of high trait anxiety and depression. Ann. Med. 2014, 46, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Partonen, T. Clock gene variants in mood and anxiety disorders. J. Neural Transm. (Vienna) 2012, 119, 1133–1145. [Google Scholar] [CrossRef]
- Geoffroy, P.A.; Curis, E.; Courtin, C.; Moreira, J.; Morvillers, T.; Etain, B.; Laplanche, J.L.; Bellivier, F.; Marie-Claire, C. Lithium response in bipolar disorders and core clock genes expression. World J. Biol. Psychiatry 2018, 19, 619–632. [Google Scholar] [CrossRef]
- Jones, G.; Rong, C.; Vecera, C.M.; Gurguis, C.I.; Chudal, R.; Khairova, R.; Leung, E.; Ruiz, A.C.; Shahani, L.; Zanetti, M.V.; et al. The role of lithium treatment on comorbid anxiety symptoms in patients with bipolar depression. J. Affect. Disord. 2022, 308, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Smagin, D.A.; Kudryavtseva, N.N. Anxiogenic and anxiolytic effects of lithium chloride under preventive and therapeutic treatments of male mice with repeated experience of aggression. Zhurnal Vyss. Nervn. Deiatelnosti Im. I P Pavlov. 2014, 64, 646–659. [Google Scholar]
- Schnell, A.; Chappuis, S.; Schmutz, I.; Brai, E.; Ripperger, J.A.; Schaad, O.; Welzl, H.; Descombes, P.; Alberi, L.; Albrecht, U. The nuclear receptor REV-ERBalpha regulates Fabp7 and modulates adult hippocampal neurogenesis. PLoS ONE 2014, 9, e99883. [Google Scholar] [CrossRef]
- Smagin, D.A.; Kovalenko, I.L.; Galyamina, A.G.; Orlov, Y.L.; Babenko, V.N.; Kudryavtseva, N.N. Heterogeneity of Brain Ribosomal Genes Expression Following Positive Fighting Experience in Male Mice as Revealed by RNA-Seq. Mol. Neurobiol. 2018, 55, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Valeri, J.; O’Donovan, S.M.; Wang, W.; Sinclair, D.; Bollavarapu, R.; Gisabella, B.; Platt, D.; Stockmeier, C.; Pantazopoulos, H. Altered expression of somatostatin signaling molecules and clock genes in the hippocampus of subjects with substance use disorder. Front. Neurosci. 2022, 16, 903941. [Google Scholar] [CrossRef]
- Piechota, M.; Korostynski, M.; Sikora, M.; Golda, S.; Dzbek, J.; Przewlocki, R. Common transcriptional effects in the mouse striatum following chronic treatment with heroin and methamphetamine. Genes Brain Behav. 2012, 11, 404–414. [Google Scholar] [CrossRef]
- Jurkowski, M.P.; Bettio, L.; Woo, E.; Patten, A.; Yau, S.Y.; Gil-Mohapel, J. Beyond the Hippocampus and the SVZ: Adult Neurogenesis Throughout the Brain. Front. Cell. Neurosci. 2020, 14, 576444. [Google Scholar] [CrossRef]
- Hagerman, R.J.; Berry-Kravis, E.; Hazlett, H.C.; Bailey, D.B., Jr.; Moine, H.; Kooy, R.F.; Tassone, F.; Gantois, I.; Sonenberg, N.; Mandel, J.L.; et al. Fragile X syndrome. Nat. Rev. Dis. Prim. 2017, 3, 17065. [Google Scholar] [CrossRef]
- Saldarriaga, W.; Tassone, F.; Gonzalez-Teshima, L.Y.; Forero-Forero, J.V.; Ayala-Zapata, S.; Hagerman, R. Fragile X syndrome. Colomb. Med. (Cali) 2014, 45, 190–198. [Google Scholar] [CrossRef]
- Eadie, B.D.; Zhang, W.N.; Boehme, F.; Gil-Mohapel, J.; Kainer, L.; Simpson, J.M.; Christie, B.R. Fmr1 knockout mice show reduced anxiety and alterations in neurogenesis that are specific to the ventral dentate gyrus. Neurobiol. Dis. 2009, 36, 361–373. [Google Scholar] [CrossRef]
- Luo, Y.; Shan, G.; Guo, W.; Smrt, R.D.; Johnson, E.B.; Li, X.; Pfeiffer, R.L.; Szulwach, K.E.; Duan, R.; Barkho, B.Z.; et al. Fragile x mental retardation protein regulates proliferation and differentiation of adult neural stem/progenitor cells. PLoS Genet. 2010, 6, e1000898. [Google Scholar] [CrossRef]
- Smagin, D.A.; Bondar, N.P.; Kudryavtseva, N.N. Repeated aggression and implications of deprivation in male mice. Psychopharmacol. Biol. Narcol. 2010, 10, 2636–2648. [Google Scholar] [CrossRef]
- Overall, R.W.; Kempermann, G. The Small World of Adult Hippocampal Neurogenesis. Front. Neurosci. 2018, 12, 641. [Google Scholar] [CrossRef]
- Flati, T.; Gioiosa, S.; Chillemi, G.; Mele, A.; Oliverio, A.; Mannironi, C.; Rinaldi, A.; Castrignano, T. A gene expression atlas for different kinds of stress in the mouse brain. Sci. Data 2020, 7, 437. [Google Scholar] [CrossRef] [PubMed]
- Kudryavtseva, N.N. The sensory contact model for the study of aggressive and submissive behaviors in male mice. Aggress. Behav. 1991, 17, 285–291. [Google Scholar] [CrossRef]
- Allen. Allen Institute for Brain Science. Allen Mouse Brain Atlas 2004. Available online: http://mouse.brain-map.org/static/atlas:2004 (accessed on 24 April 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinform. 2015, 51, 11–14. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. Post-Genome Informatics. Kyoto Encyclopedia of Genes and Genomes; Oxford University Press: Oxford, UK, 2000; p. 148. Available online: http://www.genome.jp/kegg (accessed on 8 October 2022).
- RGD. Neurological Disease Portal, Rat Genome Database Web Site. Medical College of Wisconsin: Milwaukee, WI, USA. Available online: http://rgd.mcw.edu/ (accessed on 16 October 2022).
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Ravasi, T.; Suzuki, H.; Cannistraci, C.V.; Katayama, S.; Bajic, V.B.; Tan, K.; Akalin, A.; Schmeier, S.; Kanamori-Katayama, M.; Bertin, N.; et al. An atlas of combinatorial transcriptional regulation in mouse and man. Cell 2010, 140, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Barker, M.; Rayens, W. Partial least squares for discrimination. J. Chemom. 2003, 17, 166–173. [Google Scholar] [CrossRef]
- Polunin, D.; Shtaiger, I.; Efimov, V. JACOBI4 software for multivariate analysis of biological data. bioRxiv 2019, 803684. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Correlation between DEGs’ Expression and PLS-DA Axis 1 |r| | No. of DEGs | Transcription Factor Genes | Abnormal Aggression- Related Behavior | Increased Aggression | Abnormal Anxiety-Related Response | Abnormal Fear-Related Response | Abnormal Response to Social Novelty | Learning or Memory | Abnormal Learning/ Memory/ Conditioning |
|---|---|---|---|---|---|---|---|---|---|
| 1.00–0.90 | 15 | 4 | 3 | 2 | 4 | 3 | 1 | 1 | |
| 0.89–0.80 | 19 | 2 | 2 | 2 | 5 | 2 | 1 | ||
| 0.79–0.70 | 18 | 6 | 3 | 2 | |||||
| 0.69–0.60 | 8 | 1 | 1 | 3 | |||||
| 0.59–0.50 | 6 | 1 | 2 | 1 | 1 | 1 | |||
| 0.49–0.40 | 4 | 1 | 1 | 1 | |||||
| 0.39–0.30 | |||||||||
| 0.29–0.20 | 1 | ||||||||
| 0.19–0.10 | |||||||||
| 0.09–0.00 | 1 |
| Correlation between DEGs’ Expression and PLS-DA Axis 1 |r| | Abnormal Aggression-Related Behavior | Increased Aggression | Abnormal Anxiety- Related Response | Abnormal Fear-Related Response | Abnormal Response to Social Novelty | Learning or Memory | Abnormal Learning/ Memory/ Conditioning |
|---|---|---|---|---|---|---|---|
| 1.00–0.95 | Nr1d1 * | ||||||
| 0.94–0.90 | Fmr1 * | Fmr1 * | Fmr1 * | Fmr1 * | Fmr1 * | Fmr1 * | |
| Braf | Braf | Atf2 * | Pten | ||||
| Pten | Pten | Fxr2 * |
| Transcription Factor Genes | Gene Expression, FPKM | log2 (Fold Change) A20/C | q_Value | Correlation between Gene Expression and PLS-DA Axis 1 | Gene Name | |
|---|---|---|---|---|---|---|
| Control | A20 | |||||
| Nr1d1 * | 63.81 | 82.94 | 0.38 | 0.001 | −0.957 | Nuclear receptor subfamily 1, group D, member 1 |
| Fmr1 * | 19.02 | 14.66 | −0.38 | 0.001 | 0.944 | Fragile X messenger ribonucleoprotein 1 |
| Atf2 * | 31.49 | 26.75 | −0.24 | 0.001 | 0.931 | Activating transcription factor 2 |
| Braf | 21.25 | 18.12 | −0.23 | 0.0026 | 0.919 | Braf transforming gene |
| Pten | 37.27 | 30.34 | −0.30 | 0.0010 | 0.917 | Phosphatase and tensin homolog |
| Fxr2 * | 46.90 | 53.13 | 0.18 | 0.019 | −0.909 | Fragile X mental retardation, autosomal homolog 2 |
| Correlation between DEGs’ Expression and PLS-DA Axis 1 |r| | No. of DEGs | Transcription Factor Genes | Abnormal Aggression-Related Behavior | Increased Aggression | Abnormal Anxiety- Related Response | Abnormal Fear-Related Response | Abnormal Response to Social Novelty | Learning or Memory | Abnormal Learning/ Memory/ Conditioning |
|---|---|---|---|---|---|---|---|---|---|
| 1.00–0.90 | 6 | 3 | 2 | 1 | 3 | 2 | 1 | 1 | |
| 0.89–0.80 | 7 | 1 | |||||||
| 0.79–0.70 | 7 | 2 | 1 | 1 | 3 | ||||
| 0.69–0.60 | 4 | 1 | 1 | 1 | 1 | ||||
| 0.59–0.50 | 3 | ||||||||
| 0.49–0.40 | 3 | 1 | 1 | ||||||
| 0.39–0.30 | |||||||||
| 0.29–0.20 | 1 |
| Transcription Factor Genes | Gene Expression, FPKM | log2 (Fold Change) AD/C | q_Value | Correlation between Gene Expression and PLS-DA Axis 1 | Gene Name | |
|---|---|---|---|---|---|---|
| Control | AD | |||||
| Nr1d1 | 64.02 | 78.53 | 0.29 | 0.002 | −0.968 | Nuclear receptor subfamily 1, group D, member 1 |
| Fmr1 | 19.09 | 15.37 | −0.31 | 0.002 | 0.956 | Fragile X messenger ribonucleoprotein 1 |
| Creb1 | 7.61 | 5.75 | −0.41 | 0.002 | 0.942 | cAMP responsive element binding protein 1 |
| Correlation between DEGs’ Expression and PLS-DA Axis 1 |r| | Abnormal Aggression- Related Behavior | Increased Aggression | Abnormal Anxiety-Related Response | Abnormal Fear-Related Response | Abnormal Response to Social Novelty | Learning or Memory | Abnormal Learning/ Memory/ Conditioning |
|---|---|---|---|---|---|---|---|
| 1.00–0.95 | Nr1d1 * | ||||||
| Fmr1 * | Fmr1 * | Fmr1 * | Fmr1 * | Fmr1 * | Fmr1 * | ||
| 0.94–0.90 | Pten | Pten | Pten |
| Key C_AD DEGs Related to Neurogenesis and Behavioral Traits | |||||
|---|---|---|---|---|---|
| Nr1d1 | Fmr1 | Pten | |||
| Gene Symbol | r | Gene Symbol | r | Gene Symbol | r |
| Nufip2 | −0.973 | Tmem229a | 0.975 | Eif5a2 | 0.975 |
| Sacm1l | −0.970 | Lztfl1 | 0.962 | Pcm1 | 0.973 |
| Xpo1 | −0.969 | Pcm1 | 0.961 | Dzip3 | 0.973 |
| Pcmtd2 | −0.968 | Abcb7 | 0.960 | Pum2 | 0.972 |
| Hnrnpc | −0.968 | Spred1 * | 0.958 | Lnp | 0.972 |
| 9330159F19Rik | −0.965 | Pten *#§ | 0.955 | Vcpip1 | 0.971 |
| Tmem33 | −0.962 | Azin1 | 0.954 | Rlim | 0.971 |
| Pcnp | −0.961 | Rlim | 0.954 | Elavl2 © | 0.968 |
| Picalm | −0.960 | Far1 | 0.954 | Taok1 | 0.968 |
| Rexo1 | 0.959 | Dzip3 | 0.953 | Fndc3a | 0.968 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Redina, O.E.; Babenko, V.N.; Smagin, D.A.; Kovalenko, I.L.; Galyamina, A.G.; Efimov, V.M.; Kudryavtseva, N.N. Effects of Positive Fighting Experience and Its Subsequent Deprivation on the Expression Profile of Mouse Hippocampal Genes Associated with Neurogenesis. Int. J. Mol. Sci. 2023, 24, 3040. https://doi.org/10.3390/ijms24033040
Redina OE, Babenko VN, Smagin DA, Kovalenko IL, Galyamina AG, Efimov VM, Kudryavtseva NN. Effects of Positive Fighting Experience and Its Subsequent Deprivation on the Expression Profile of Mouse Hippocampal Genes Associated with Neurogenesis. International Journal of Molecular Sciences. 2023; 24(3):3040. https://doi.org/10.3390/ijms24033040
Chicago/Turabian StyleRedina, Olga E., Vladimir N. Babenko, Dmitry A. Smagin, Irina L. Kovalenko, Anna G. Galyamina, Vadim M. Efimov, and Natalia N. Kudryavtseva. 2023. "Effects of Positive Fighting Experience and Its Subsequent Deprivation on the Expression Profile of Mouse Hippocampal Genes Associated with Neurogenesis" International Journal of Molecular Sciences 24, no. 3: 3040. https://doi.org/10.3390/ijms24033040
APA StyleRedina, O. E., Babenko, V. N., Smagin, D. A., Kovalenko, I. L., Galyamina, A. G., Efimov, V. M., & Kudryavtseva, N. N. (2023). Effects of Positive Fighting Experience and Its Subsequent Deprivation on the Expression Profile of Mouse Hippocampal Genes Associated with Neurogenesis. International Journal of Molecular Sciences, 24(3), 3040. https://doi.org/10.3390/ijms24033040