


DNA Methylation and Gene Expression of the Cysteinyl Leukotriene Receptors as a Prognostic and Metastatic Factor for Colorectal Cancer Patients



Abstract
:1. Introduction
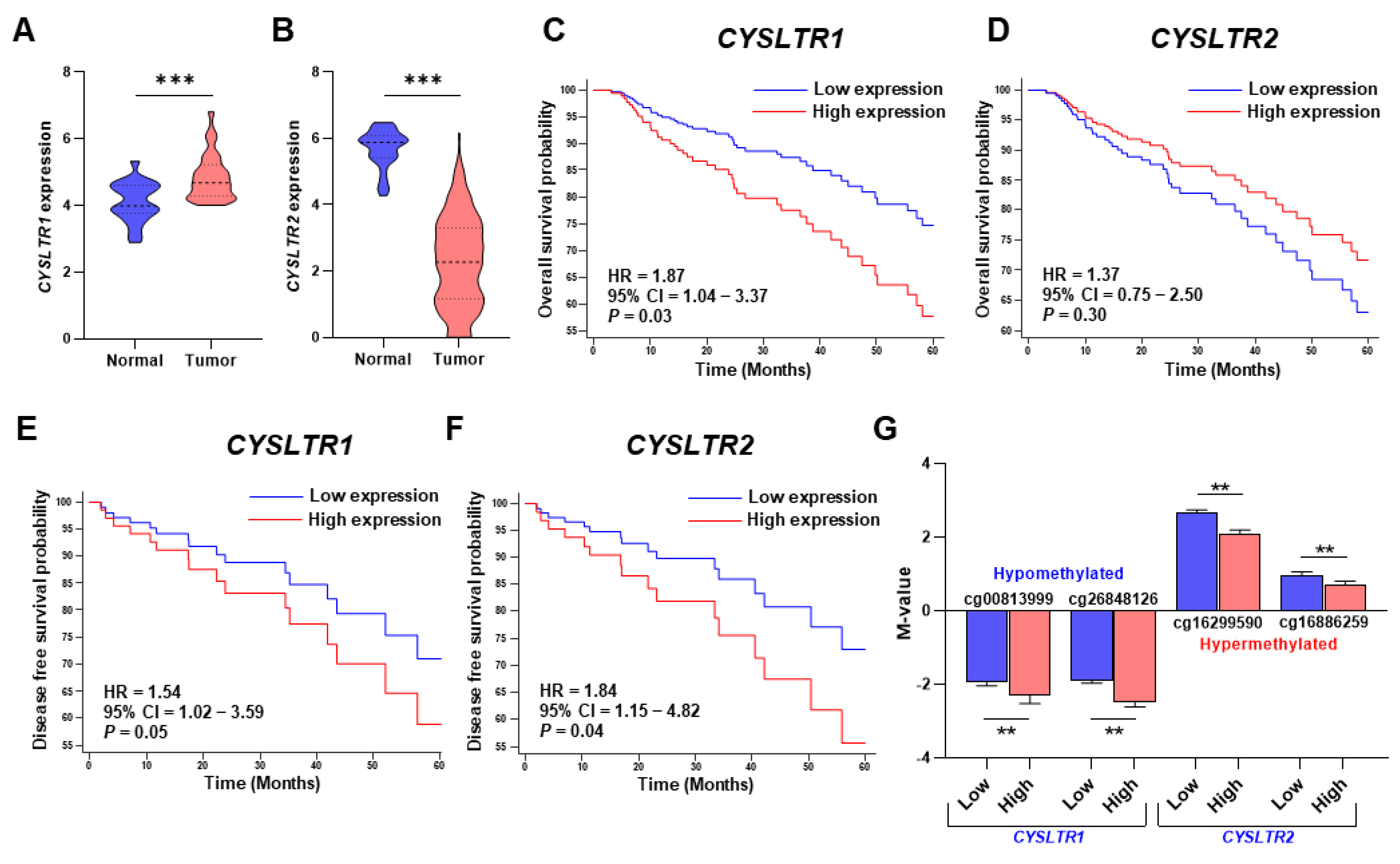
2. Results
3. Discussion
4. Materials and Methods
4.1. Patient Cohorts
4.2. Analysis of DNA Methylation in the Cancer Genome TCGA and GSE Cohort
4.3. Analysis of Gene Expression in the Cancer Genome TCGA and GSE Cohorts
4.4. MSP Primers Designing for DNA Methylation Analysis
4.5. DNA/RNA Extraction from FFPE Tissue and Bisulfite Modification of Extracted DNA
4.6. DNA Methylation by qPCR and Melt Curve Analysis
4.7. qPCR for CYSLTR1, CYSLTR2, CDH1 and VIM Gene Expression
4.8. CRISPR-Cas9 Based Knockdown of CysLT1R
4.9. SW620 Cells Colonosphere Formation and Western Blot Analysis
4.10. Statistical Analysis and Data Visualization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Nogueira, L.; Devasia, T.; Mariotto, A.B.; Yabroff, K.R.; Jemal, A.; Kramer, J.; Siegel, R.L. Cancer treatment and survivorship statistics, 2022. CA Cancer J. Clin. 2022, 72, 409–436. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Lee, S.; Brock, T.G. Leukotriene synthesis by epithelial cells. Histol. Histopathol. 2003, 118, 587–595. [Google Scholar] [CrossRef]
- Tian, W.; Jiang, X.; Kim, D.; Guan, T.; Nicolls, M.R.; Rockson, S.G. Leukotrienes in Tumor-Associated Inflammation. Front. Pharmacol. 2020, 11, 1289. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, T.; Nakamura, M.; Shimizu, T. Leukotriene receptors as potential therapeutic targets. J. Clin. Investig. 2018, 128, 2691–2701. [Google Scholar] [CrossRef] [PubMed]
- Slater, K.; Hoo, P.S.; Buckley, A.M.; Piulats, J.M.; Villanueva, A.; Portela, A.; Kennedy, B.N. Evaluation of oncogenic cysteinyl leukotriene receptor 2 as a therapeutic target for uveal melanoma. Cancer Metastasis Rev. 2018, 37, 335–345. [Google Scholar] [CrossRef]
- Burke, L.; Butler, C.T.; Murphy, A.; Moran, B.; Gallagher, W.M.; O’Sullivan, J.; Kennedy, B.N. Evaluation of Cysteinyl Leukotriene Signaling as a Therapeutic Target for Colorectal Cancer. Front. Cell Dev. Biol. 2016, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Duah, E.; Teegala, L.R.; Kondeti, V.; Adapala, R.K.; Keshamouni, V.G.; Kanaoka, Y.; Austen, K.F.; Thodeti, C.K.; Paruchuri, S. Cysteinyl leukotriene 2 receptor promotes endothelial permeability, tumor angiogenesis, and metastasis. Proc. Natl. Acad. Sci. USA 2019, 116, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-J.; Wu, P.-H.; Sheu, C.-C.; Hsu, Y.-L.; Chang, W.-A.; Hung, J.-Y.; Yang, C.-J.; Yang, Y.-H.; Kuo, P.-L.; Huang, M.-S. Cysteinyl Leukotriene Receptor Antagonists Decrease Cancer Risk in Asthma Patients. Sci. Rep. 2016, 6, 23979. [Google Scholar] [CrossRef]
- Kawahito, Y.; Sano, H.; Nakatani, T.; Yoshimura, R.; Naganuma, T.; Funao, K.; Matsuyama, M. The cysteinylLT1 receptor in human renal cell carcinoma. Mol. Med. Rep. 2008, 1, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, M.; Funao, K.; Kawahito, Y.; Sano, H.; Chargui, J.; Touraine, J.-L.; Nakatani, T.; Yoshimura, R. Expression of cysteinylLT1 receptor in human testicular cancer and growth reduction by its antagonist through apoptosis. Mol. Med. Rep. 2009, 2, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, M.; Funao, K.; Hayama, T.; Tanaka, T.; Kawahito, Y.; Sano, H.; Takemoto, Y.; Nakatani, T.; Yoshimura, R. Relationship Between Cysteinyl-Leukotriene-1 Receptor and Human Transitional Cell Carcinoma in Bladder. Urology 2009, 73, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C.K.; Öhd, J.F.; Wikström, K.; Massoumi, R.; Paruchuri, S.; Juhas, M.; Sjölander, A. The Leukotriene Receptor CYSLT1 And 5- Lipoxygenase Are Upregulated In Colon Cancer. Adv. Exp. Med. Biol. 2003, 525, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, C.; Mezhybovska, M.; Lörinc, E.; Fernebro, E.; Nilbert, M.; Sjölander, A. Low expression of CysLT1R and high expression of CysLT2R mediate good prognosis in colorectal cancer. Eur. J. Cancer 2010, 46, 826–835. [Google Scholar] [CrossRef]
- Mehrabi, S.F.; Ghatak, S.; Mehdawi, L.M.; Topi, G.; Satapathy, S.R.; Sjölander, A. Brain-Derived Neurotrophic Factor, Neutrophils and Cysteinyl Leukotriene Receptor 1 as Potential Prognostic Biomarkers for Patients with Colon Cancer. Cancers 2021, 13, 5520. [Google Scholar] [CrossRef]
- Magnusson, C.; Liu, J.; Ehrnström, R.; Manjer, J.; Jirström, K.; Andersson, T.; Sjölander, A. Cysteinyl leukotriene receptor expression pattern affects migration of breast cancer cells and survival of breast cancer patients. Int. J. Cancer 2011, 129, 9–22. [Google Scholar] [CrossRef]
- Bellamkonda, K.; Satapathy, S.R.; Douglas, D.; Chandrashekar, N.; Selvanesan, B.C.; Liu, M.; Savari, S.; Jonsson, G.; Sjölander, A. Montelukast, a CysLT1 receptor antagonist, reduces colon cancer stemness and tumor burden in a mouse xenograft model of human colon cancer. Cancer Lett. 2018, 437, 13–24. [Google Scholar] [CrossRef]
- Moore, A.R.; Ceraudo, E.; Sher, J.J.; Guan, Y.; Shoushtari, A.N.; Chang, M.T.; Zhang, J.Q.; Walczak, E.G.; Kazmi, M.A.; Taylor, B.S.; et al. Recurrent activating mutations of G-protein-coupled receptor CYSLTR2 in uveal melanoma. Nat. Genet. 2016, 48, 675–680. [Google Scholar] [CrossRef]
- Van de Nes, J.A.; Koelsche, C.; Gessi, M.; Möller, I.; Sucker, A.; Scolyer, R.A.; Buckland, M.E.; Pietsch, T.; Murali, R.; Schadendorf, D.; et al. Activating CYSLTR2 and PLCB4 Mutations in Primary Leptomeningeal Melanocytic Tumors. J. Investig. Dermatol. 2017, 137, 2033–2035. [Google Scholar] [CrossRef]
- Ye, Z.; Zhou, M.; Tian, B.; Wu, B.; Li, J. Expression of lncRNA-CCAT1, E-cadherin and N-cadherin in colorectal cancer and its clinical significance. Int. J. Clin. Exp. Med. 2015, 8, 3707–3715. [Google Scholar]
- Salim, T.; Sand-Dejmek, J.; Sjölander, A. The inflammatory mediator leukotriene D4 induces subcellular β-catenin translocation and migration of colon cancer cells. Exp. Cell Res. 2014, 321, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Vinnakota, K.; Zhang, Y.; Selvanesan, B.C.; Topi, G.; Salim, T.; Sand-Dejmek, J.; Jönsson, G.; Sjölander, A. M2-like macrophages induce colon cancer cell invasion via matrix metalloproteinases. J. Cell. Physiol. 2017, 232, 3468–3480. [Google Scholar] [CrossRef] [PubMed]
- McGovern, T.; Goldberger, M.; Chen, M.; Allard, B.; Hamamoto, Y.; Kanaoka, Y.; Austen, K.F.; Powell, W.S.; Martin, J.G. CysLT1 Receptor Is Protective against Oxidative Stress in a Model of Irritant-Induced Asthma. J. Immunol. 2016, 197, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Liu, Y. DNA methylation in human diseases. Genes Dis. 2018, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Luo, H.; Krawczyk, M.; Wei, W.; Wang, W.; Wang, J.; Flagg, K.; Hou, J.; Zhang, H.; Yi, S.; et al. DNA methylation markers for diagnosis and prognosis of common cancers. Proc. Natl. Acad. Sci. USA 2017, 114, 7414–7419. [Google Scholar] [CrossRef]
- Díez-Villanueva, A.; Sanz-Pamplona, R.; Carreras-Torres, R.; Moratalla-Navarro, F.; Alonso, M.H.; Pare, L.; Aussó, S.; Guinó, E.; Solé, X.; Cordero, D.; et al. DNA methylation events in transcription factors and gene expression changes in colon cancer. Epigenomics 2020, 12, 1593–1610. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, X.; Lu, Y.; Wu, J.; Feng, J. DNA-methylated gene markers for colorectal cancer in TCGA database. Exp. Ther. Med. 2020, 19, 3042–3050. [Google Scholar] [CrossRef]
- Wajed, S.A.; Laird, P.W.; Demeester, T.R. DNA Methylation: An Alternative Pathway to Cancer. Ann. Surg. 2001, 234, 10–20. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, K. Pan-cancer analysis of frequent DNA co-methylation patterns reveals consistent epigenetic landscape changes in multiple cancers. BMC Genom. 2017, 18, 1045. [Google Scholar] [CrossRef]
- Ding, W.; Chen, G.; Shi, T. Integrative analysis identifies potential DNA methylation biomarkers for pan-cancer diagnosis and prognosis. Epigenetics 2019, 14, 67–80. [Google Scholar] [CrossRef]
- Tang, W.; Wan, S.; Yang, Z.; Teschendorff, A.E.; Zou, Q. Tumor origin detection with tissue-specific miRNA and DNA methylation markers. Bioinformatics 2018, 34, 398–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irizarry, R.A.; Ladd-Acosta, C.; Wen, B.; Wu, Z.; Montano, C.; Onyango, P.; Cui, H.; Gabo, K.; Rongione, M.; Webster, M.; et al. The human colon cancer methylome shows similar hypo- and hypermethylation at conserved tissue-specific CpG island shores. Nat. Genet. 2009, 41, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Liu, Z.; Peng, K.; Wu, W. A Prognostic Model Based on Nine DNA Methylation-Driven Genes Predicts Overall Survival for Colorectal Cancer. Front. Genet. 2021, 12, 779383. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-J.; Chang, W.-A.; Chuang, C.-H.; Wu, K.-L.; Cheng, C.-H.; Sheu, C.-C.; Hsu, Y.-L.; Hung, J.-Y. Cysteinyl Leukotriene Pathway and Cancer. Int. J. Mol. Sci. 2022, 23, 120. [Google Scholar] [CrossRef]
- Mehdawi, L.M.; Satapathy, S.R.; Gustafsson, A.; Lundholm, K.; Alvarado-Kristensson, M.; Sjölander, A. A potential anti-tumor effect of leukotriene C4 through the induction of 15-hydroxyprostaglandin dehydrogenase expression in colon cancer cells. Oncotarget 2017, 8, 35033–35047. [Google Scholar] [CrossRef]
- Lande-Diner, L.; Cedar, H. Silence of the genes—mechanisms of long-term repression. Nat. Rev. Genet. 2005, 6, 648–654. [Google Scholar] [CrossRef]
- Zaidi, S.K.; Van Wijnen, A.J.; Lian, J.B.; Stein, J.L.; Stein, G.S. Targeting deregulated epigenetic control in cancer. J. Cell. Physiol. 2013, 228, 2103–2108. [Google Scholar] [CrossRef]
- Lee, K.; Moon, S.; Park, M.-J.; Koh, I.-U.; Choi, N.-H.; Yu, H.-Y.; Kim, Y.J.; Kong, J.; Kang, H.G.; Kim, S.C.; et al. Integrated Analysis of Tissue-Specific Promoter Methylation and Gene Expression Profile in Complex Diseases. Int. J. Mol. Sci. 2020, 21, 5056. [Google Scholar] [CrossRef]
- Rabinovitch, N.; Jones, M.J.; Gladish, N.; Faino, A.V.; Strand, M.; Morin, A.M.; MacIsaac, J.; Lin, D.T.S.; Reynolds, P.R.; Singh, A.; et al. Methylation of cysteinyl leukotriene receptor 1 genes associates with lung function in asthmatics exposed to traffic-related air pollution. Epigenetics 2021, 16, 177–185. [Google Scholar] [CrossRef]
- Öhd, J.F.; Nielsen, C.K.; Campbell, J.; Landberg, G.; Löfberg, H.; Sjölander, A. Expression of the leukotriene D4 receptor CysLT1, COX-2, and other cell survival factors in colorectal adenocarcinomas. Gastroenterology 2003, 124, 57–70. [Google Scholar] [CrossRef]
- Nielsen, C.K.; Campbell, J.I.; Ohd, J.F.; Mörgelin, M.; Riesbeck, K.; Landberg, G.; Sjölander, A. A Novel Localization of the G-Protein-Coupled CysLT1 Receptor in the Nucleus of Colorectal Adenocarcinoma Cells. Cancer Res. 2005, 65, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, M.; Hayama, T.; Funao, K.; Kawahito, Y.; Sano, H.; Takemoto, Y.; Nakatani, T.; Yoshimura, R. Overexpression of cysteinyl LT1 receptor in prostate cancer and CysLT1R antagonist inhibits prostate cancer cell growth through apoptosis. Oncol. Rep. 2007, 18, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Savari, S.; Chandrashekar, N.K.; Osman, J.; Douglas, D.; Bellamkonda, K.; Jönsson, G.; Juhas, M.; Greicius, G.; Pettersson, S.; Sjölander, A. Cysteinyl leukotriene 1 receptor influences intestinal polyp incidence in a gender-specific manner in the ApcMin/+mouse model. Carcinogenesis 2016, 37, 491–499. [Google Scholar] [CrossRef]
- Szabo, E.; Mao, J.T.; Lam, S.; Reid, M.E.; Keith, R.L. Chemoprevention of Lung Cancer: Diagnosis and management of lung cancer, 3rd ed: American College of Chest Physicians evidence-based clinical practice guidelines. Chest 2013, 143, e40S–e60S. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.P.; Peters-Golden, M. Antileukotriene Agents for the Treatment of Lung Disease. Am. J. Respir. Crit. Care Med. 2013, 188, 538–544. [Google Scholar] [CrossRef]
- Wang, D.; DuBois, R.N. Eicosanoids and cancer. Nat. Rev. Cancer 2010, 10, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Sutton, S.S.; Magagnoli, J.; Cummings, T.H.; Hardin, J.W. Leukotriene inhibition and the risk of lung cancer among U.S. veterans with asthma. Pulm. Pharmacol. Ther. 2021, 71, 102084. [Google Scholar] [CrossRef]
- Wijnhoven, B.P.L.; Dinjens, W.N.M.; Pignatelli, M. E-cadherin—Catenin cell—Cell adhesion complex and human cancer. Br. J. Surg. 2000, 87, 992–1005. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Bertrand, F.E.; Davis, N.M.; Sokolosky, M.; Abrams, S.L.; Montalto, G.; D’Assoro, A.B.; Libra, M.; Nicoletti, F.; et al. GSK-3 as potential target for therapeutic intervention in cancer. Oncotarget 2014, 5, 2881–2911. [Google Scholar] [CrossRef]
- Zhou, B.P.; Deng, J.; Xia, W.; Xu, J.; Li, Y.M.; Gunduz, M.C.; Hung, M.-C. Dual regulation of Snail by GSK-3β-mediated phosphorylation in control of epithelial–mesenchymal transition. Nat. Cell Biol. 2004, 6, 931–940. [Google Scholar] [CrossRef]
- Hennig, R.; Ventura, J.; Segersvärd, R.; Ward, E.; Ding, X.-Z.; Rao, S.M.; Jovanovic, B.D.; Iwamura, T.; Talamonti, M.S.; Bell, R.H., Jr.; et al. LY293111 Improves Efficacy of Gemcitabine Therapy on Pancreatic Cancer in a Fluorescent Orthotopic Model in Athymic Mice. Neoplasia 2005, 7, 417–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukic, A.; Wahlund, C.J.; Gómez, C.; Brodin, D.; Samuelsson, B.; Wheelock, C.E.; Gabrielsson, S.; Rådmark, O. Exosomes and cells from lung cancer pleural exudates transform LTC4 to LTD4, promoting cell migration and survival via CysLT1. Cancer Lett. 2019, 444, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lee, S.-J.; Cho, H.J.; Kim, T.W.; Kim, J.-T.; Kim, J.W.; Lee, C.-H.; Kim, B.-Y.; Yeom, Y.I.; Lim, J.-S.; et al. Dehydropeptidase 1 promotes metastasis through regulation of E-cadherin expression in colon cancer. Oncotarget 2016, 7, 9501–9512. [Google Scholar] [CrossRef]
- Satapathy, S.R.; Sjölander, A. Cysteinyl leukotriene receptor 1 promotes 5-fluorouracil resistance and resistance-derived stemness in colon cancer cells. Cancer Lett. 2020, 488, 50–62. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Research Network; Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef]
- Qu, X.; Sandmann, T.; Frierson, H., Jr.; Fu, L.; Fuentes, E.; Walter, K.; Okrah, K.; Rumpel, C.; Moskaluk, C.; Lu, S.; et al. Integrated genomic analysis of colorectal cancer progression reveals activation of EGFR through demethylation of the EREG promoter. Oncogene 2016, 35, 6403–6415. [Google Scholar] [CrossRef]
- Tjärnberg, A.; Mahmood, O.; Jackson, C.A.; Saldi, G.-A.; Cho, K.; Christiaen, L.A.; Bonneau, R.A. Optimal tuning of weighted kNN- and diffusion-based methods for denoising single cell genomics data. PLOS Comput. Biol. 2021, 17, e1008569. [Google Scholar] [CrossRef]
- Li, L.-C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 18, 1427–1431. [Google Scholar] [CrossRef]
- Ghatak, S.; Sanga, Z.; Pautu, J.L.; Kumar, N.S. Coextraction and PCR Based Analysis of Nucleic Acids from Formalin-Fixed Paraffin-Embedded Specimens. J. Clin. Lab. Anal. 2015, 29, 485–492. [Google Scholar] [CrossRef]
- Smith, E.; Jones, M.E.; Drew, P.A. Quantitation of DNA methylation by melt curve analysis. BMC Cancer 2009, 9, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satapathy, S.R.; Topi, G.; Osman, J.; Hellman, K.; Ek, F.; Olsson, R.; Sime, W.; Mehdawi, L.M.; Sjölander, A. Tumour suppressor 15-hydroxyprostaglandin dehydrogenase induces differentiation in colon cancer via GLI1 inhibition. Oncogenesis 2020, 9, 74. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Datasets | Sample Type | Tissue Types | Data Types | Platform | Age (Mean ± SD) | Gender | Anatomical Location | TNM Stage | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Male (n) | Female (n) | Missing (n) | Left (n) | Right (n) | Missing (n) | Stage I | Stage II | Stage III | Stage IV | ||||||
| TCGA COADREAD cohort | N (n = 41) | FF | Tissue mRNA and DNA methylation | RNA-seq and HumanMethylation450k | 63.64 ± 13.83 | 22 | 19 | - | 20 | 16 | 5 | - | - | - | - |
| PT (n = 375) | 63.56 ± 13.92 | 207 | 168 | - | 187 | 163 | 25 | 57 | 143 | 123 | 52 | ||||
| GSE77955 cohort | N (n = 13) | FF | Tissue mRNA and DNA methylation | Gene expression Microarray and HuanMethylation450k | 52.46 ± 10.64 | 5 | 5 | 2 | 5 | 5 | 3 | - | - | - | - |
| PT (n = 17) | 64.55 ± 11.38 | 11 | 6 | - | 11 | 6 | - | - | - | - | 17 | ||||
| AD (n = 17) | |||||||||||||||
| 53.26 ± 8.53 | 7 | 8 | 2 | 5 | 7 | 5 | - | - | - | - | |||||
| ME (Liver, n = 10 Ovary, n = 1) | 66.28 ± 11.24 | 6 | 5 | - | - | - | - | - | - | - | - | ||||
| E-MTAB-8148 cohort | N (n = 32) | FF | Tissue mRNA and DNA methylation | RNA-seq and HumanMethylation450k | 68.92 ± 16.55 | 6 | 26 | Missing | Missing | ||||||
| PT (n = 216) | 66.24 ± 13.10 | 100 | 116 | ||||||||||||
| Malmö-CC cohort | N (n = 20) | FFPE | Tissue mRNA and DNA methylation | qRT-PCR and quantitative methylation-specific PCR | 65.73 ± 12.73 | 14 | 6 | - | 9 | 8 | 3 | - | - | - | - |
| PT (n = 20) | 65.73 ± 12.73 | 14 | 6 | - | 9 | 8 | 3 | - | 5 | 13 | 2 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghatak, S.; Satapathy, S.R.; Sjölander, A. DNA Methylation and Gene Expression of the Cysteinyl Leukotriene Receptors as a Prognostic and Metastatic Factor for Colorectal Cancer Patients. Int. J. Mol. Sci. 2023, 24, 3409. https://doi.org/10.3390/ijms24043409
Ghatak S, Satapathy SR, Sjölander A. DNA Methylation and Gene Expression of the Cysteinyl Leukotriene Receptors as a Prognostic and Metastatic Factor for Colorectal Cancer Patients. International Journal of Molecular Sciences. 2023; 24(4):3409. https://doi.org/10.3390/ijms24043409
Chicago/Turabian StyleGhatak, Souvik, Shakti Ranjan Satapathy, and Anita Sjölander. 2023. "DNA Methylation and Gene Expression of the Cysteinyl Leukotriene Receptors as a Prognostic and Metastatic Factor for Colorectal Cancer Patients" International Journal of Molecular Sciences 24, no. 4: 3409. https://doi.org/10.3390/ijms24043409
APA StyleGhatak, S., Satapathy, S. R., & Sjölander, A. (2023). DNA Methylation and Gene Expression of the Cysteinyl Leukotriene Receptors as a Prognostic and Metastatic Factor for Colorectal Cancer Patients. International Journal of Molecular Sciences, 24(4), 3409. https://doi.org/10.3390/ijms24043409