
Comparison of Glyphosate-Degradation Ability of Aldo-Keto Reductase (AKR4) Proteins in Maize, Soybean and Rice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Phylogenetic Analysis of AKR in Plants
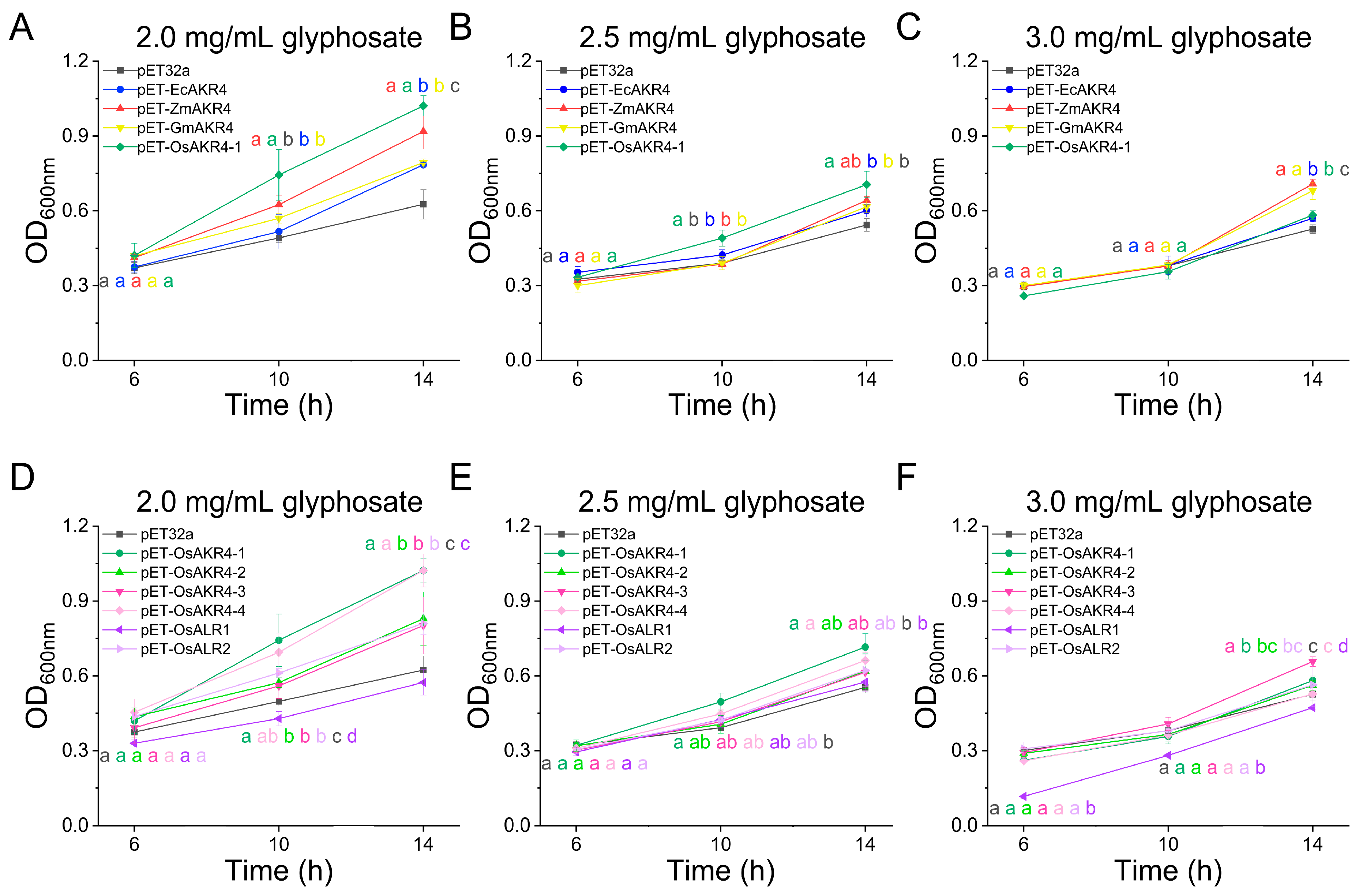
2.2. AKR-Expressing E. coli Cells Acquire Tolerance to Glyphosate
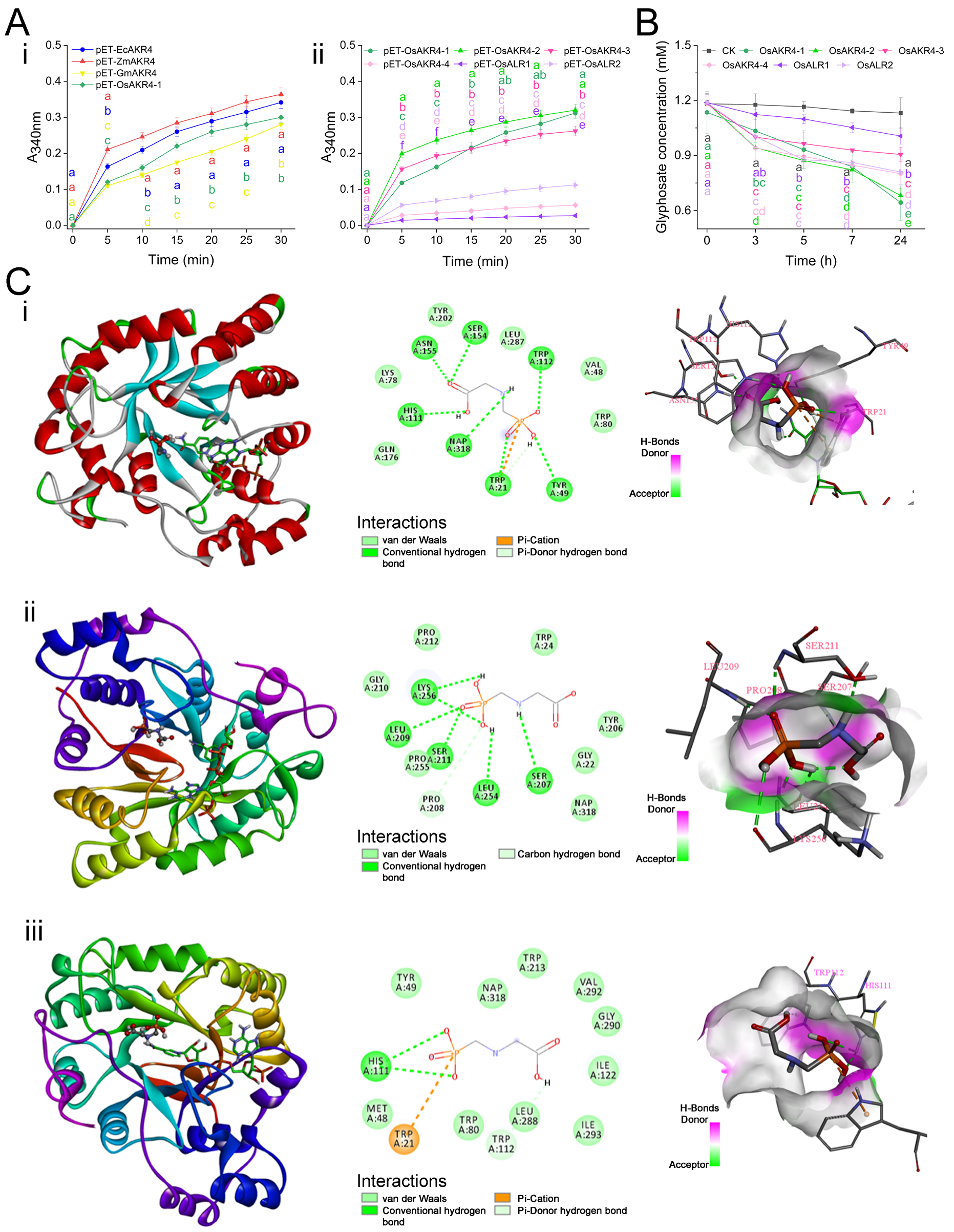
2.3. AKR Protein Expression and Substrate Activity
2.4. OsAKR Proteins Are Involved in Glyphosate Metabolism
2.5. The Response of OsAKRs in Rice Seedlings to Glyphosate Treatment
2.6. Localization of OsAKR4-1 in Rice
3. Materials and Methods
3.1. Sequence Homology among Different Plants
3.2. Cloning Procedure and Plasmid Construction
3.3. Expression and Purification of AKRs in E. coli
3.4. Comparative-Growth Test of Glyphosate in E. coli
3.5. Measurement of Enzyme Activity in E. coli
3.6. Degradation of Glyphosate by E. coli
3.7. Molecular Docking of AKRs with Glyphosate
3.8. Measurement of Glyphosate Using UPLC-MS/MS
3.9. Gene-Expression Analysis Using qRT-PCR
3.10. Glyphosate-Tolerance Seed-Germination Assay
3.11. Subcellular Localization of OsAKR4-1
3.12. Histochemical Determination of GUS Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duke, S.O.; Powles, S.B.; Sammons, R.D. Glyphosate-How it became a once in a hundred year herbicide and its future. Outlooks Pest Manag. 2018, 29, 247–251. [Google Scholar] [CrossRef]
- Padgette, S.R.; Kolacz, K.H.; Delannay, X.; Re, D.B.; LaVallee, B.J.; Tinius, C.N.; Rhodes, W.K.; Otero, Y.I.; Barry, G.F.; Eichholtz, D.A. Development, identification, and characterization of a glyphosate-tolerant soybean line. Crop. Sci. 1995, 35, 1451–1461. [Google Scholar] [CrossRef]
- Tan, S.; Evans, R.; Singh, B. Herbicidal inhibitors of amino acid biosynthesis and herbicide-tolerant crops. Amino Acids 2006, 30, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef]
- Wen, L.; Zhong, J.; Cui, Y.; Duan, Z.; Zhou, F.; Li, C.; Ma, W.; Yin, C.; Chen, H.; Lin, Y. Coexpression of I. variabilis-EPSPS* and WBceGO-B3S1 genes contributes to high glyphosate tolerance and low glyphosate residues in transgenic rice. J. Agric. Food Chem. 2021, 69, 7388–7398. [Google Scholar] [CrossRef]
- Bostamam, Y.; Malone, J.M.; Dolman, F.C.; Boutsalis, P.; Preston, C. Rigid Ryegrass (Lolium rigidum) Populations containing a target site mutation in EPSPS and reduced glyphosate translocation are more resistant to glyphosate. Weed Sci. 2012, 60, 474–479. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef]
- Morran, S.; Moretti, M.L.; Brunharo, C.A.; Fischer, A.J.; Hanson, B.D. Multiple target site resistance to glyphosate in junglerice (Echinochloa colona) lines from California orchards. Pest Manag. Sci. 2018, 74, 2747–2753. [Google Scholar] [CrossRef]
- Pan, L.; Yu, Q.; Wang, J.; Han, H.; Mao, L.; Nyporko, A.; Maguza, A.; Fan, L.; Bai, L.; Powles, S. An ABCC-type transporter endowing glyphosate resistance in plants. Proc. Natl. Acad. Sci. USA 2021, 118, e2100136118. [Google Scholar] [CrossRef]
- Bøhn, T.; Cuhra, M.; Traavik, T.; Sanden, M.; Fagan, J.; Primicerio, R. Compositional differences in soybeans on the market: Glyphosate accumulates in Roundup Ready GM soybeans. Food Chem. 2014, 153, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Barry, G.; Kishore, G.; Padgette, S.; Taylor, M.; Kolacz, K.; Weldon, M.; Re, D.; Eichholtz, D.; Fincher, K.; Hallas, L. Inhibitors of amino acid biosynthesis: Strategies for imparting glyphosate tolerance to crop plants. In Biosynthesis and Molecular Regulation of Amino Acids in Plants; Singh, B.K., Flores, H.E., Shannon, J.C., Eds.; Springer: Rockville, MD, USA, 1992; pp. 139–145. [Google Scholar]
- Zhou, H.; Arrowsmith, J.W.; Fromm, M.E.; Hironaka, C.M.; Taylor, M.L.; Rodriguez, D.; Pajeau, M.E.; Brown, S.M.; Santino, C.G.; FRY, J.E. Glyphosate-tolerant CP4 and GOX genes as a selectable marker in wheat transformation. Plant Cell Rep. 1995, 15, 159–163. [Google Scholar] [CrossRef]
- Correa, E.A.; Dayan, F.E.; Owens, D.K.; Rimando, A.M.; Duke, S.O. Glyphosate-resistant and conventional canola (Brassica napus L.) responses to glyphosate and aminomethylphosphonic acid (AMPA) treatment. J. Agric. Food Chem. 2016, 64, 3508–3513. [Google Scholar] [CrossRef]
- Mörtl, M.; Diederichs, K.; Welte, W.; Molla, G.; Motteran, L.; Andriolo, G.; Pilone, M.S.; Pollegioni, L. Structure-function correlation in glycine oxidase from Bacillus subtilis. J. Biol. Chem. 2004, 279, 29718–29727. [Google Scholar] [CrossRef]
- Pedotti, M.; Rosini, E.; Molla, G.; Moschetti, T.; Savino, C.; Vallone, B.; Pollegioni, L. Glyphosate resistance by engineering the flavoenzyme glycine oxidase. J. Biol. Chem. 2009, 284, 36415–36423. [Google Scholar] [CrossRef]
- Nicolia, A.; Ferradini, N.; Molla, G.; Biagetti, E.; Pollegioni, L.; Veronesi, F.; Rosellini, D. Expression of an evolved engineered variant of a bacterial glycine oxidase leads to glyphosate resistance in alfalfa. J. Biotechnol. 2014, 184, 201–208. [Google Scholar] [CrossRef]
- Han, H.; Zhu, B.; Fu, X.; You, S.; Wang, B.; Li, Z.; Zhao, W.; Peng, R.; Yao, Q. Overexpression of d-amino acid oxidase from Bradyrhizobium japonicum, enhances resistance to glyphosate in Arabidopsis thaliana. Plant Cell Rep. 2015, 34, 2043–2051. [Google Scholar] [CrossRef]
- Reddy, K.N.; Rimando, A.M.; Duke, S.O.; Nandula, V.K. Aminomethylphosphonic acid accumulation in plant species treated with glyphosate. J. Agric. Food Chem. 2008, 56, 2125–2130. [Google Scholar] [CrossRef]
- Duke, S.O. Glyphosate degradation in glyphosate-resistant and-susceptible crops and weeds. J. Agric. Food Chem. 2011, 59, 5835–5841. [Google Scholar] [CrossRef]
- Nandula, V.K.; Riechers, D.E.; Ferhatoglu, Y.; Barrett, M.; Duke, S.O.; Dayan, F.E.; Goldberg-Cavalleri, A.; Tétard-Jones, C.; Wortley, D.J.; Onkokesung, N.; et al. Herbicide metabolism: Crop selectivity, bioactivation, weed resistance, and regulation. Weed Sci. 2019, 67, 149–175. [Google Scholar] [CrossRef]
- Vemanna, R.S.; Vennapusa, A.R.; Easwaran, M.; Chandrashekar, B.K.; Rao, H.; Ghanti, K.; Sudhakar, C.; Mysore, K.S.; Makarla, U. Aldo-keto reductase enzymes detoxify glyphosate and improve herbicide resistance in plants. Plant Biotechnol. J. 2017, 15, 794–804. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Yu, Q.; Han, H.; Mao, L.; Nyporko, A.; Fan, L.; Bai, L.; Powles, S. Aldo-keto reductase metabolizes glyphosate and confers glyphosate resistance in Echinochloa colona. Plant Physiol. 2019, 181, 1519–1534. [Google Scholar] [CrossRef] [PubMed]
- Hyndman, D.; Bauman, D.R.; Heredia, V.V.; Penning, T.M. The aldo-keto reductase superfamily homepage. Chem. Biol. Interact. 2003, 143–144, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Mindnich, R.D.; Penning, T.M. Aldo-keto reductase (AKR) superfamily: Genomics and annotation. Hum. Genom. 2009, 3, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Simpson, P.J.; Tantitadapitak, C.; Reed, A.M.; Mather, O.C.; Bunce, C.M.; White, S.A.; Ride, J.P. Characterization of two novel aldo-keto reductases from Arabidopsis: Expression patterns, broad substrate specificity, and an open active-site structure suggest a role in toxicant metabolism following stress. J. Mol. Biol. 2009, 392, 465–480. [Google Scholar] [CrossRef]
- Penning, T.M. The aldo-keto reductases (AKRs): Overview. Chem.-Biol. Interact. 2015, 234, 236–246. [Google Scholar] [CrossRef]
- Okuda, A.; Okuda, K. Purification and characterization of delta 4-3-ketosteroid 5 beta-reductase. J. Biol. Chem. 1984, 259, 7519–7524. [Google Scholar] [CrossRef]
- Kondo, K.H.; Kai, M.H.; Setoguchi, Y.; Eggertsen, G.; Sjoblom, P.; Setoguchi, T.; Okuda, K.I.; Bjorkhem, I. Cloning and expression of cDNA of human delta4-3-oxosteroid 5 beta-reductase and substrate specificity of the expressed enzyme. Eur. J. Biochem. 1994, 219, 357–363. [Google Scholar] [CrossRef]
- Di Costanzo, L.; Drury, J.E.; Penning, T.M.; Christianson, D.W. Crystal structure of human liver Delta4-3-Ketosteroid 5beta-Reductase (AKR1D1) and implications for substrate binding and catalysis. J. Biol. Chem. 2008, 283, 16830–16839. [Google Scholar] [CrossRef]
- Saito, R.; Shimakawa, G.; Nishi, A.; Iwamoto, T.; Sakamoto, K.; Yamamoto, H.; Amako, K.; Makino, A.; Miyake, C. Functional analysis of the AKR4C subfamily of Arabidopsis thaliana: Model structures, substrate specificity, acrolein toxicity, and responses to light and [CO2]. Biosci. Biotechnol. Biochem. 2013, 77, 2038–2045. [Google Scholar] [CrossRef]
- De Sousa, S.M.; Rosselli, L.K.; Kiyota, E.; Da Silva, J.C.; Souza, G.H.M.F.; Peroni, L.A.; Stach-Machado, D.R.; Eberlin, M.N.; Souza, A.P.; Koch, K.E.; et al. Structural and kinetic characterization of a maize aldose reductase. Plant Physiol. Biochem. 2009, 47, 98–104. [Google Scholar] [CrossRef]
- Turóczy, Z.; Kis, P.; Török, K.; Cserháti, M.; Lendvai, Á.; Dudits, D.; Horváth, G.V. Overproduction of a rice aldo-keto reductase increases oxidative and heat stress tolerance by malondialdehyde and methylglyoxal detoxification. Plant Mol. Biol. 2011, 75, 399–412. [Google Scholar] [CrossRef]
- Agrawal, C.; Sen, S.; Singh, S.; Rai, S.; Singh, P.K.; Singh, V.K.; Rai, L.C. Comparative proteomics reveals association of early accumulated proteins in conferring butachlor tolerance in three N2-fixing Anabaena spp. J. Proteom. 2014, 96, 271–290. [Google Scholar] [CrossRef]
- Agrawal, C.; Sen, S.; Yadav, S.; Rai, S.; Rai, L.C. A novel aldo-keto reductase (AKR17A1) of Anabaena sp. PCC 7120 degrades the rice field herbicide butachlor and confers tolerance to abiotic stresses in E. coli. PLoS ONE 2015, 10, e0137744. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Hu, Y.; Chen, C.; Huang, J.; Min, J.; Dai, L.; Guo, R. Structural analysis and engineering of aldo-keto reductase from glyphosate-resistant Echinochloa colona. J. Hazard. Mater. 2022, 436, 129191. [Google Scholar] [CrossRef]
- Leebens-Mack, J.H.; Barker, M.S.; Carpenter, E.J.; Deyholos, M.K.; Gitzendanner, M.A.; Graham, S.W.; Grosse, I.; Li, Z.; Melkonian, M.; Mirarab, S.; et al. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef]
- Bohren, K.M.; Bullock, B.; Wermuth, B.; Gabbay, K.H. The aldo-keto reductase superfamily: cDNAs and deduced amino acid sequences of human aldehyde and aldose reductases. J. Biol. Chem. 1989, 264, 9547–9551. [Google Scholar] [CrossRef] [PubMed]
- Burczynski, M.E.; Sridhar, G.R.; Palackal, N.T.; Penning, T.M. The reactive oxygen species-and Michael acceptor-inducible human aldo-keto reductase AKR1C1 reduces the alpha, beta-unsaturated aldehyde 4-hydroxy-2-nonenal to 1, 4-dihydroxy-2-nonene. J. Biol. Chem. 2001, 276, 2890–2897. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Naik, D.; Reddy, A.R. Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plant metabolic processes and stress defense: A structure-function update. J. Plant Physiol. 2015, 179, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, K.L.; Klimacek, M.; Nidetzky, B.; Wilson, D.K. The structure of apo and holo forms of xylose reductase, a dimeric aldo-keto reductase from Candida tenuis. Biochemistry 2002, 41, 8785–8795. [Google Scholar] [CrossRef]
- De Giuseppe, P.O.; Dos Santos, M.L.; de Sousa, S.M.; Koch, K.E.; Yunes, J.A.; Aparicio, R.; Murakami, M.T. A comparative structural analysis reveals distinctive features of co-factor binding and substrate specificity in plant aldo-keto reductases. Biochem. Biophys. Res. Commun. 2016, 474, 696–701. [Google Scholar] [CrossRef] [Green Version]
- McElroy, J.S.; Hall, N.D. Echinochloa colona with reported resistance to glyphosate conferred by aldo-keto reductase also contains a Pro-106-Thr EPSPS target site mutation. Plant Physiol. 2020, 183, 447–450. [Google Scholar] [CrossRef]
- Liang, C.; Sun, B.; Meng, Z.; Meng, Z.; Wang, Y.; Sun, G.; Zhu, T.; Lu, W.; Zhang, W.; Malik, W. Co-expression of GR79 EPSPS and GAT yields herbicide-resistant cotton with low glyphosate residues. Plant Biotechnol. J. 2017, 15, 1622–1629. [Google Scholar] [CrossRef]
- Fartyal, D.; Agarwal, A.; James, D.; Borphukan, B.; Ram, B.; Sheri, V.; Yadav, R.; Manna, M.; Varakumar, P.; Achary, V.M.M. Co-expression of P173S mutant rice EPSPS and igrA genes results in higher glyphosate tolerance in transgenic rice. Front. Plant Sci. 2018, 9, 144. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, H.; Li, Z.; Huang, T.; Akihiro, T.; Xu, J.; Xu, H.; Lin, F. An amino acid transporter-like protein (OsATL15) facilitates the systematic distribution of thiamethoxam in rice for controlling the brown planthopper. Plant Biotechnol. J. 2022, 20, 1888–1901. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, R.; Wang, S.; Sun, Y.; Li, H.; Wan, S.; Lin, F.; Xu, H. Comparison of Glyphosate-Degradation Ability of Aldo-Keto Reductase (AKR4) Proteins in Maize, Soybean and Rice. Int. J. Mol. Sci. 2023, 24, 3421. https://doi.org/10.3390/ijms24043421
Chen R, Wang S, Sun Y, Li H, Wan S, Lin F, Xu H. Comparison of Glyphosate-Degradation Ability of Aldo-Keto Reductase (AKR4) Proteins in Maize, Soybean and Rice. International Journal of Molecular Sciences. 2023; 24(4):3421. https://doi.org/10.3390/ijms24043421
Chicago/Turabian StyleChen, Ronghua, Siwei Wang, Yue Sun, Haiqing Li, Shuqing Wan, Fei Lin, and Hanhong Xu. 2023. "Comparison of Glyphosate-Degradation Ability of Aldo-Keto Reductase (AKR4) Proteins in Maize, Soybean and Rice" International Journal of Molecular Sciences 24, no. 4: 3421. https://doi.org/10.3390/ijms24043421
APA StyleChen, R., Wang, S., Sun, Y., Li, H., Wan, S., Lin, F., & Xu, H. (2023). Comparison of Glyphosate-Degradation Ability of Aldo-Keto Reductase (AKR4) Proteins in Maize, Soybean and Rice. International Journal of Molecular Sciences, 24(4), 3421. https://doi.org/10.3390/ijms24043421