
Environmental Enrichment Promotes Transgenerational Programming of Uterine Inflammatory and Stress Markers Comparable to Gestational Chronic Variable Stress

 , , and
, , and
Abstract
:1. Introduction
2. Results
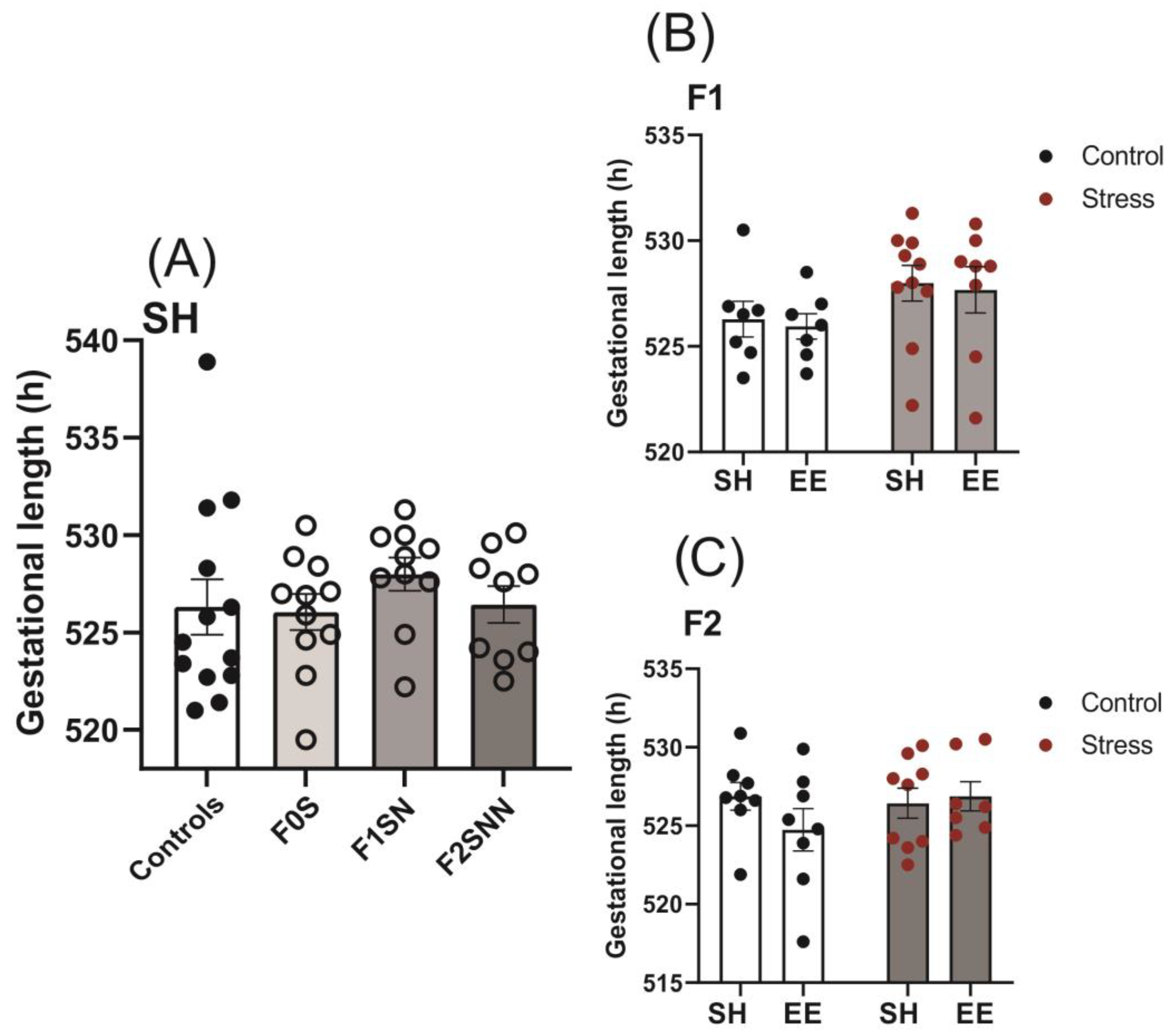
2.1. Gestational Lengths Were Unchanged in the Stress Groups across Two Generations
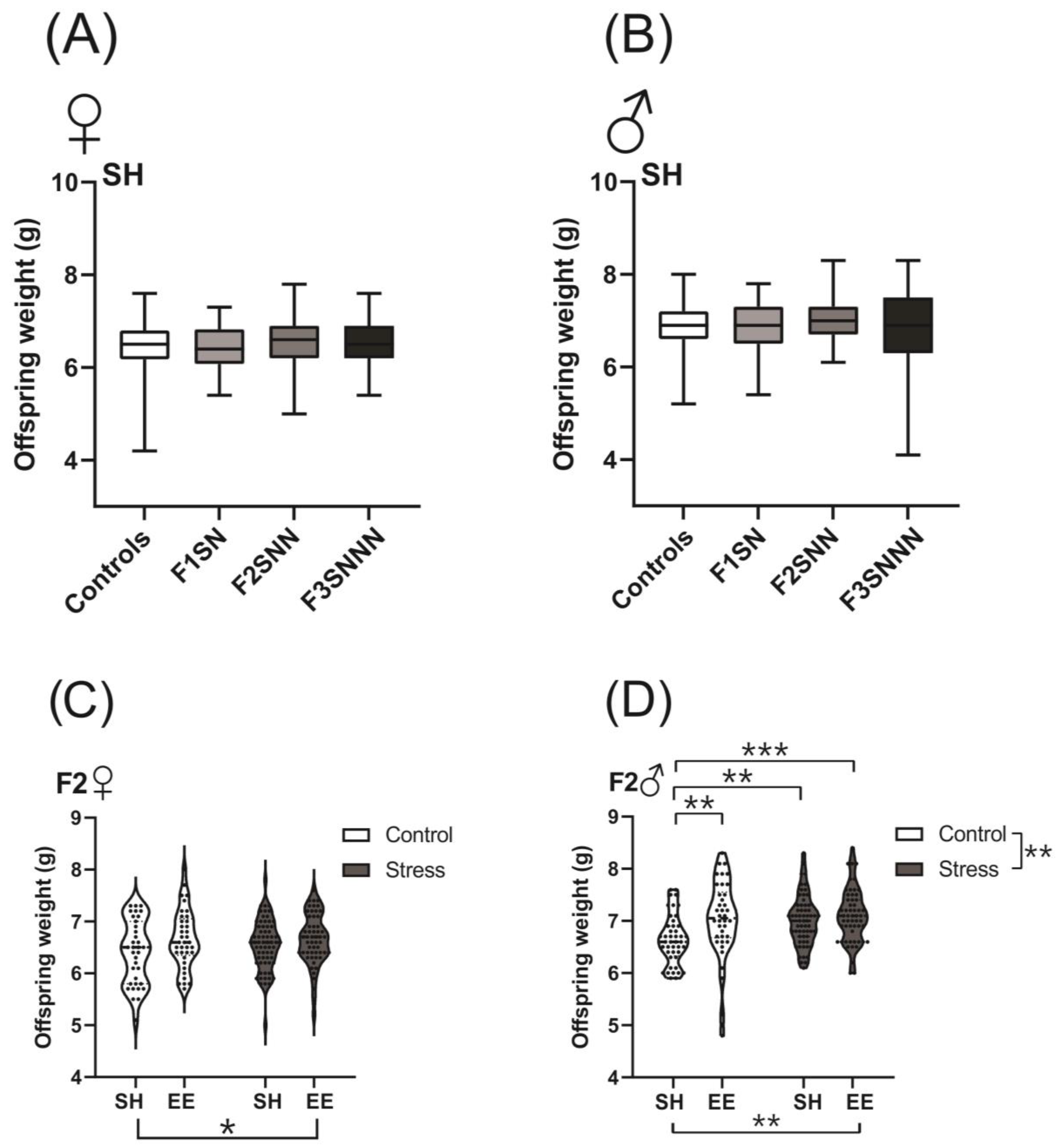
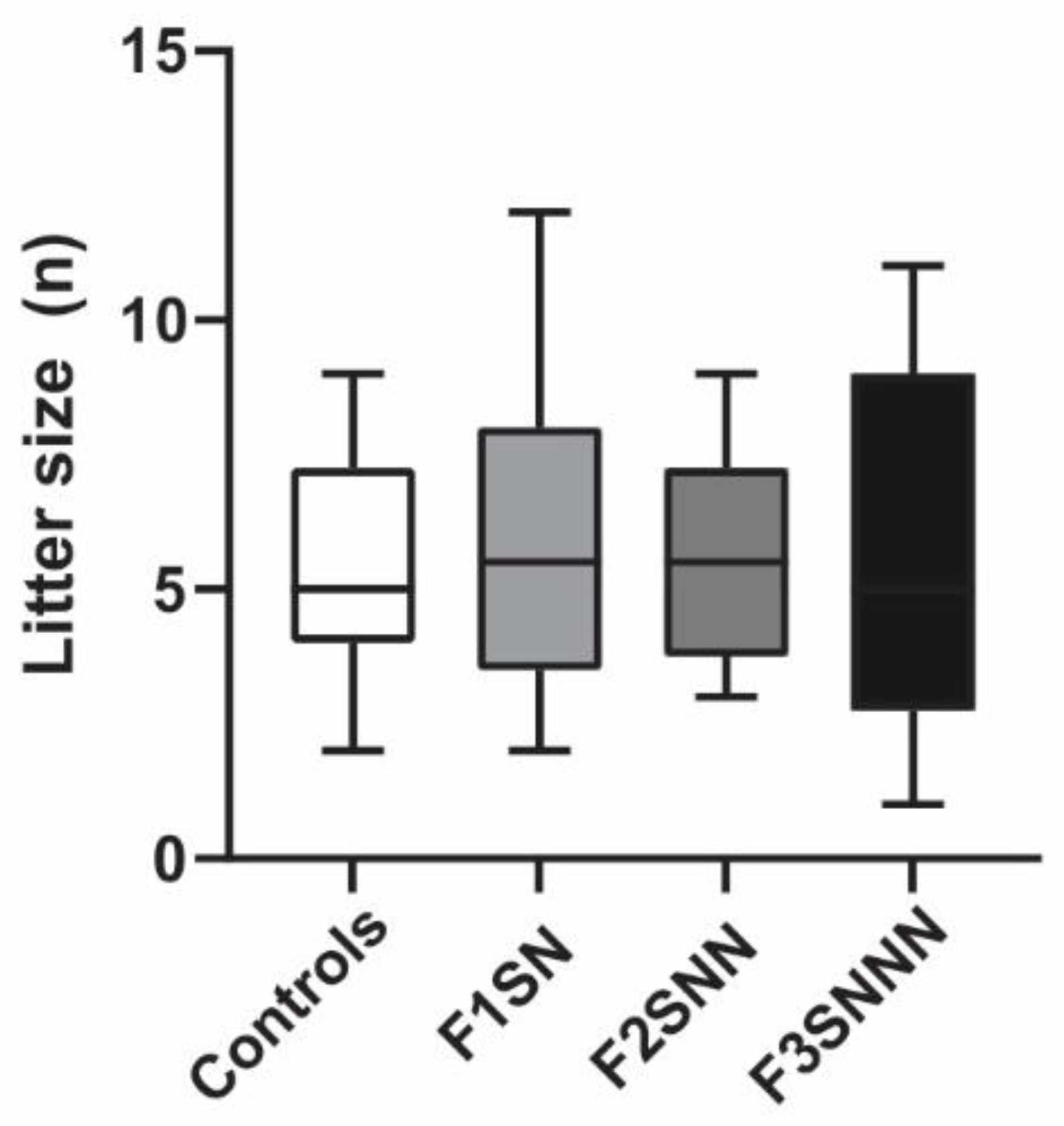
2.2. Offspring Birth Weights Were Unchanged in the Stress Group but Were Increased with Environmental Enrichment
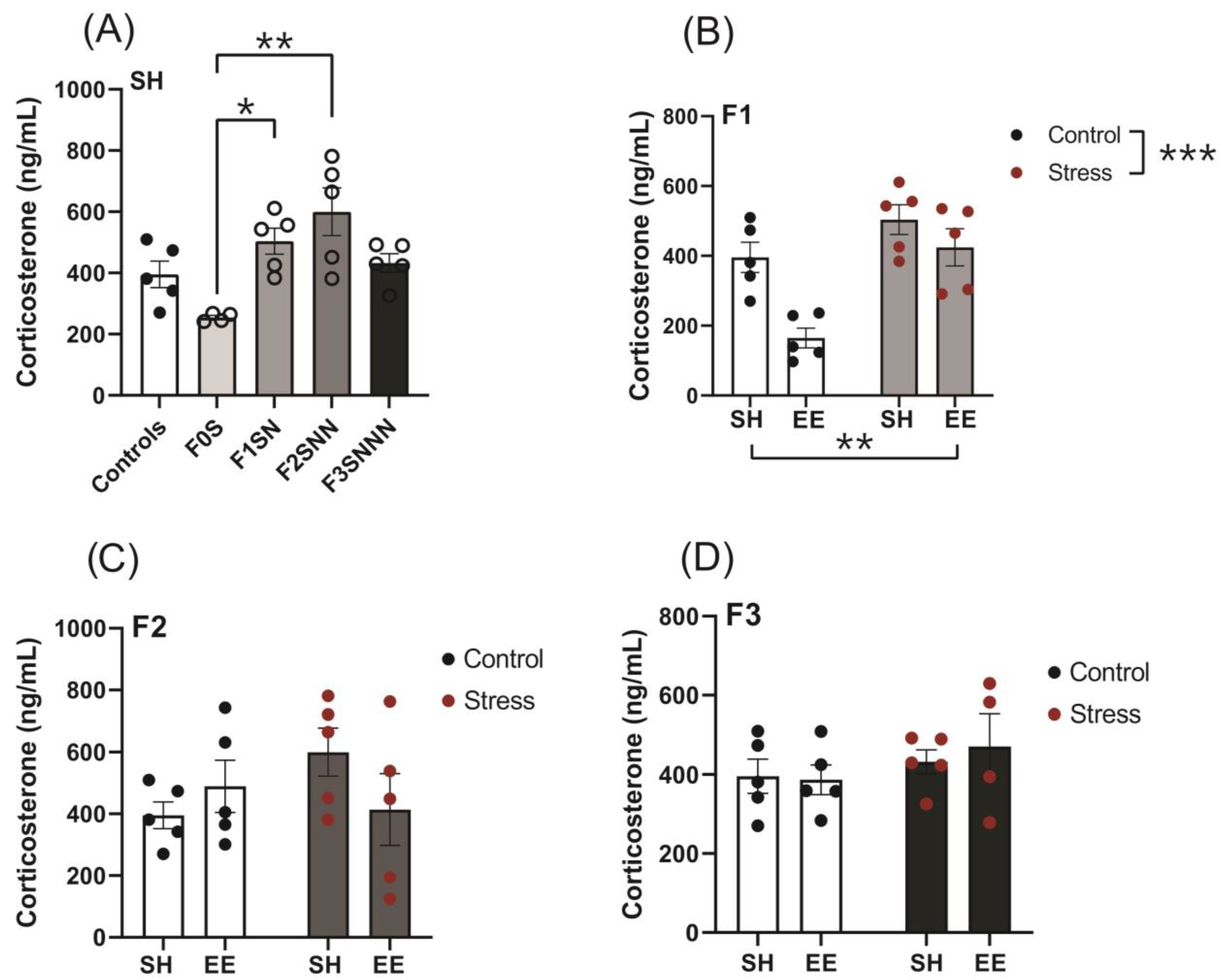
2.3. Concentrations of Plasma CORT Levels Were Increased in F1 and F2 Stressed Animals and Were Not Mitigated by EE Intervention
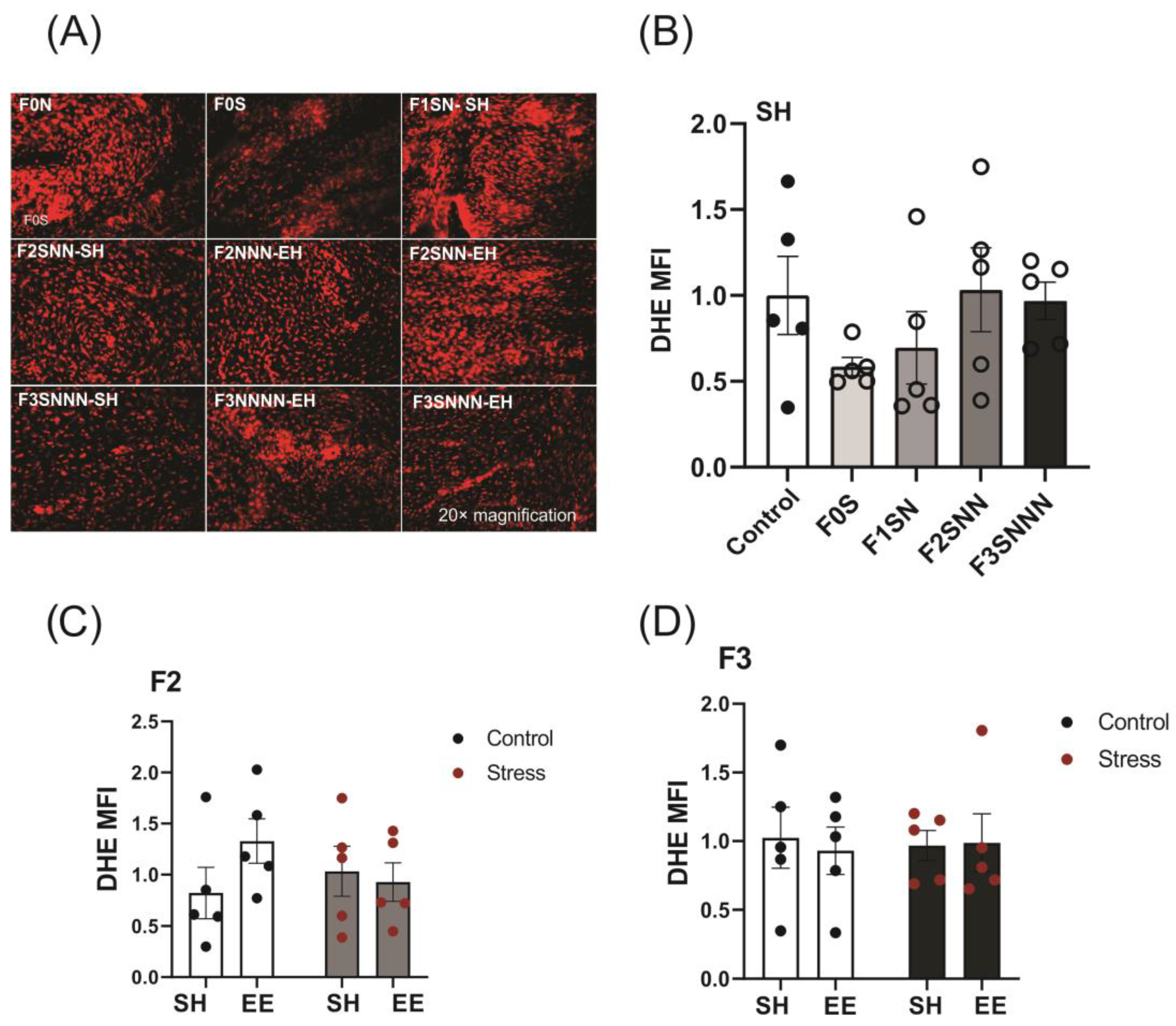
2.4. Uterine ROS Levels Were Unchanged in Animals Exposed to Gestational and Ancestral Stress
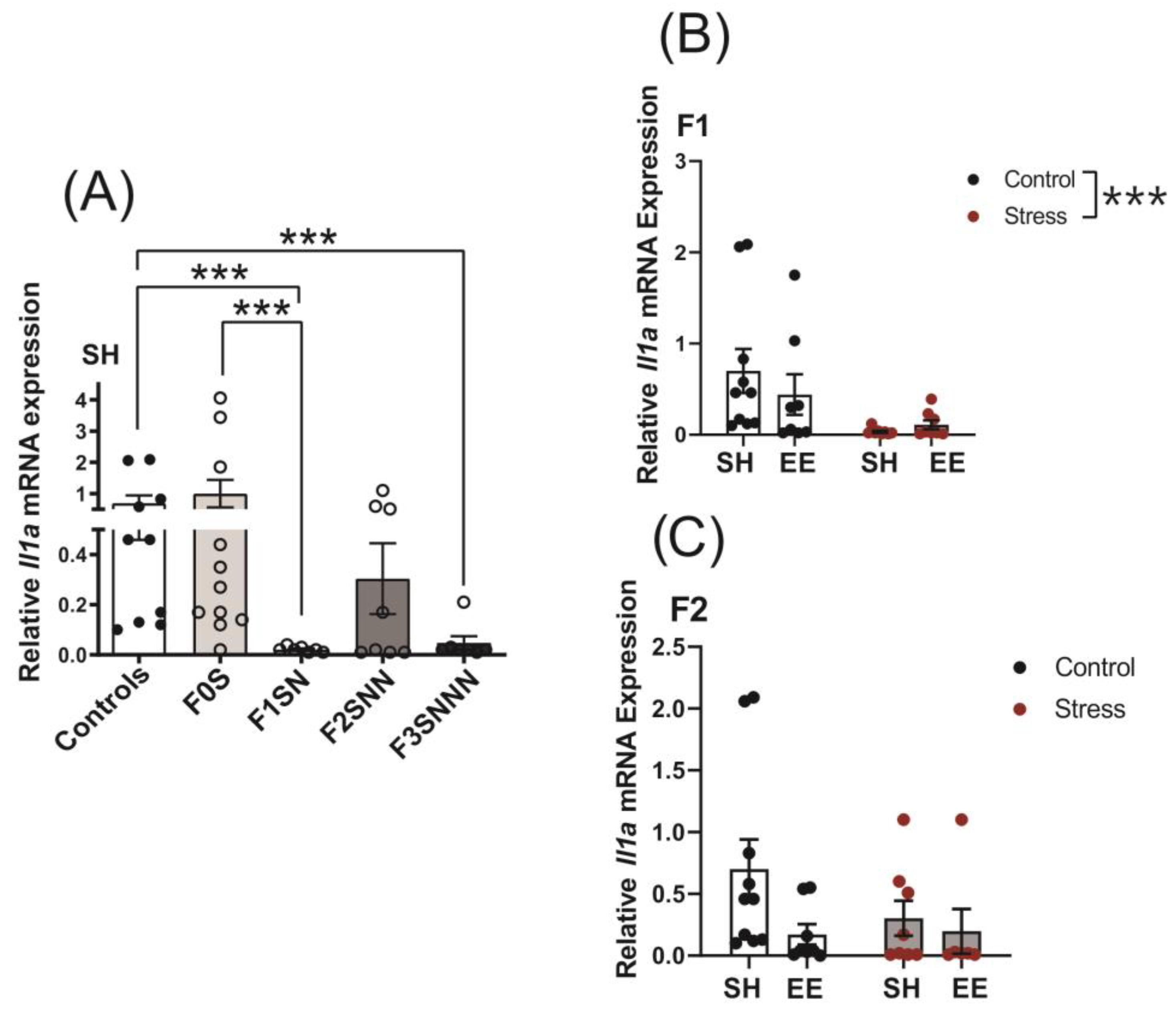
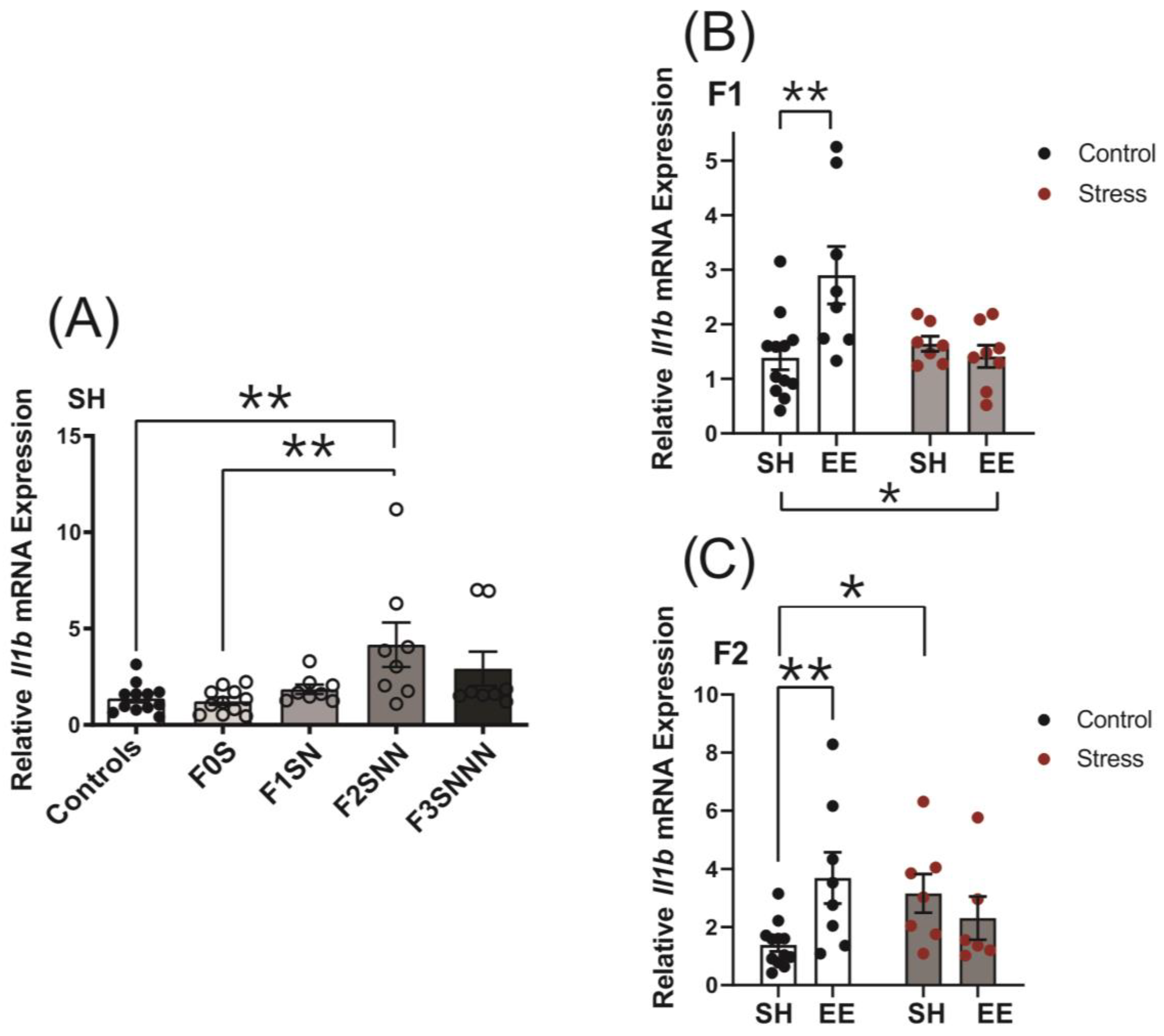
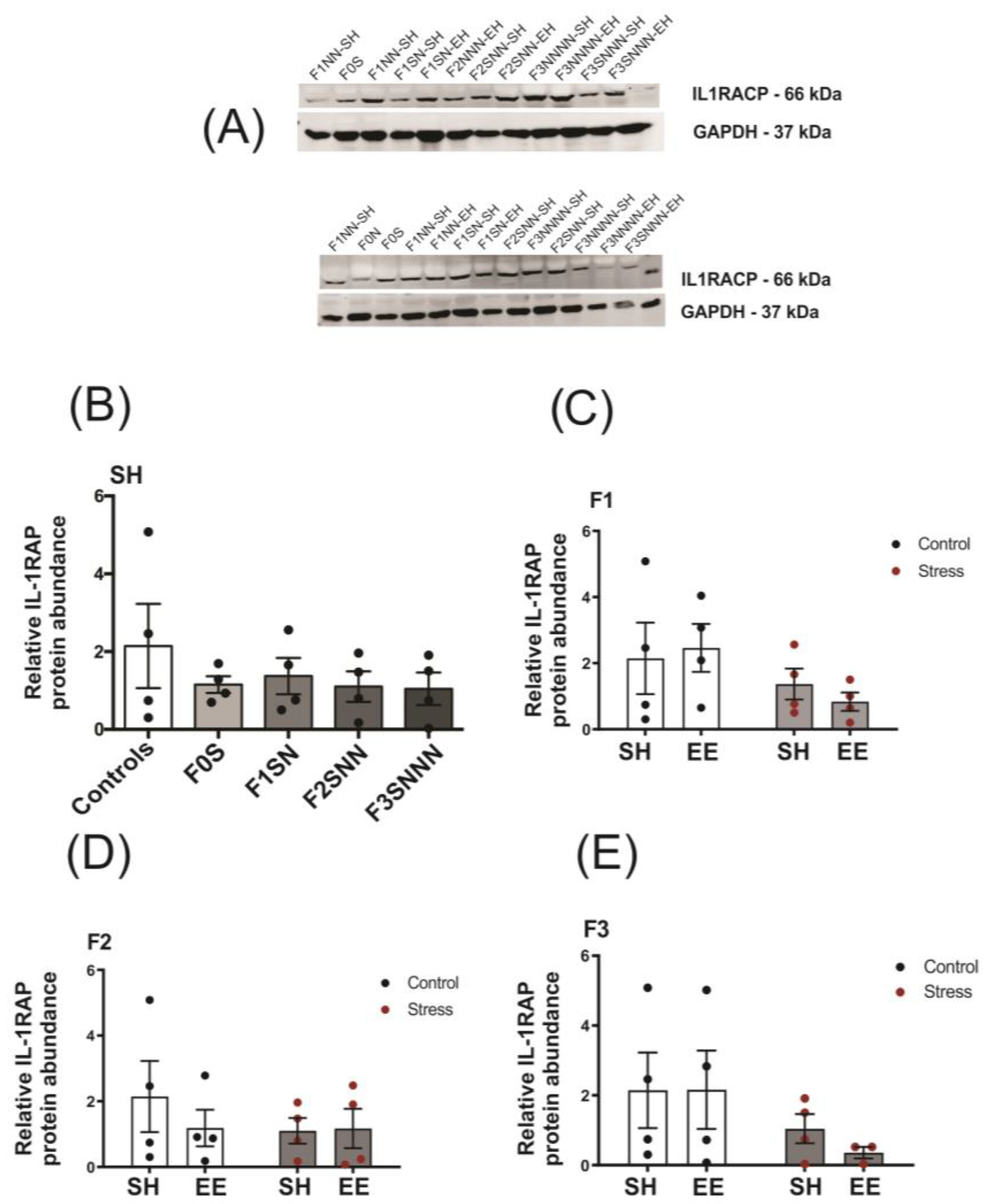
2.5. Uterine Expression of Candidate Genes and Proteins Involved in Local Inflammatory Responses Were Impacted in Rats Exposed to Ancestral Stress
2.5.1. Proinflammatory Cytokines: Il1a and Il1b
2.5.2. Psychological and Psychosocial Stress in Tandem Did Not Induce Changes in IL-1RAP Protein Levels in Rat Uteri
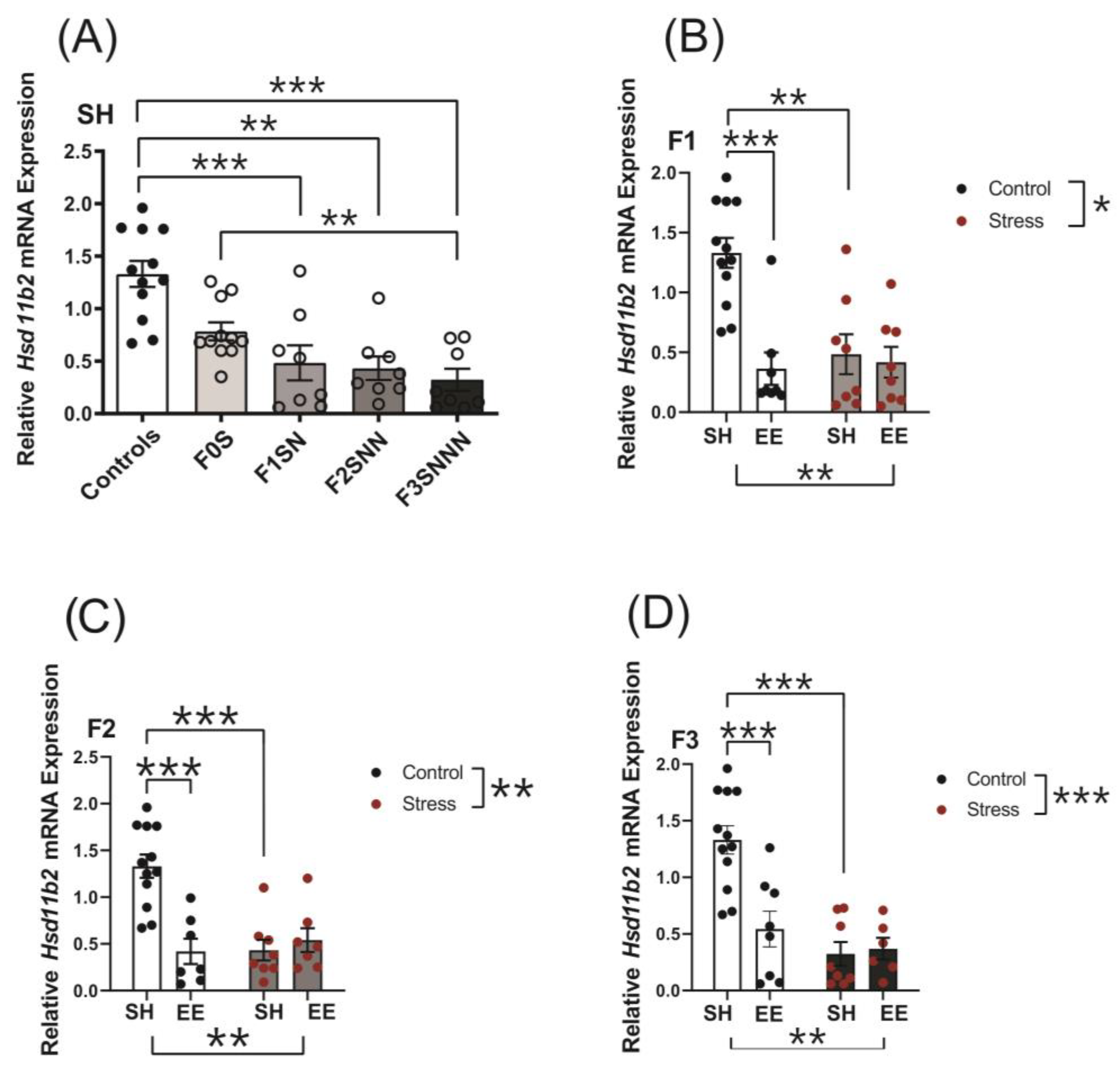
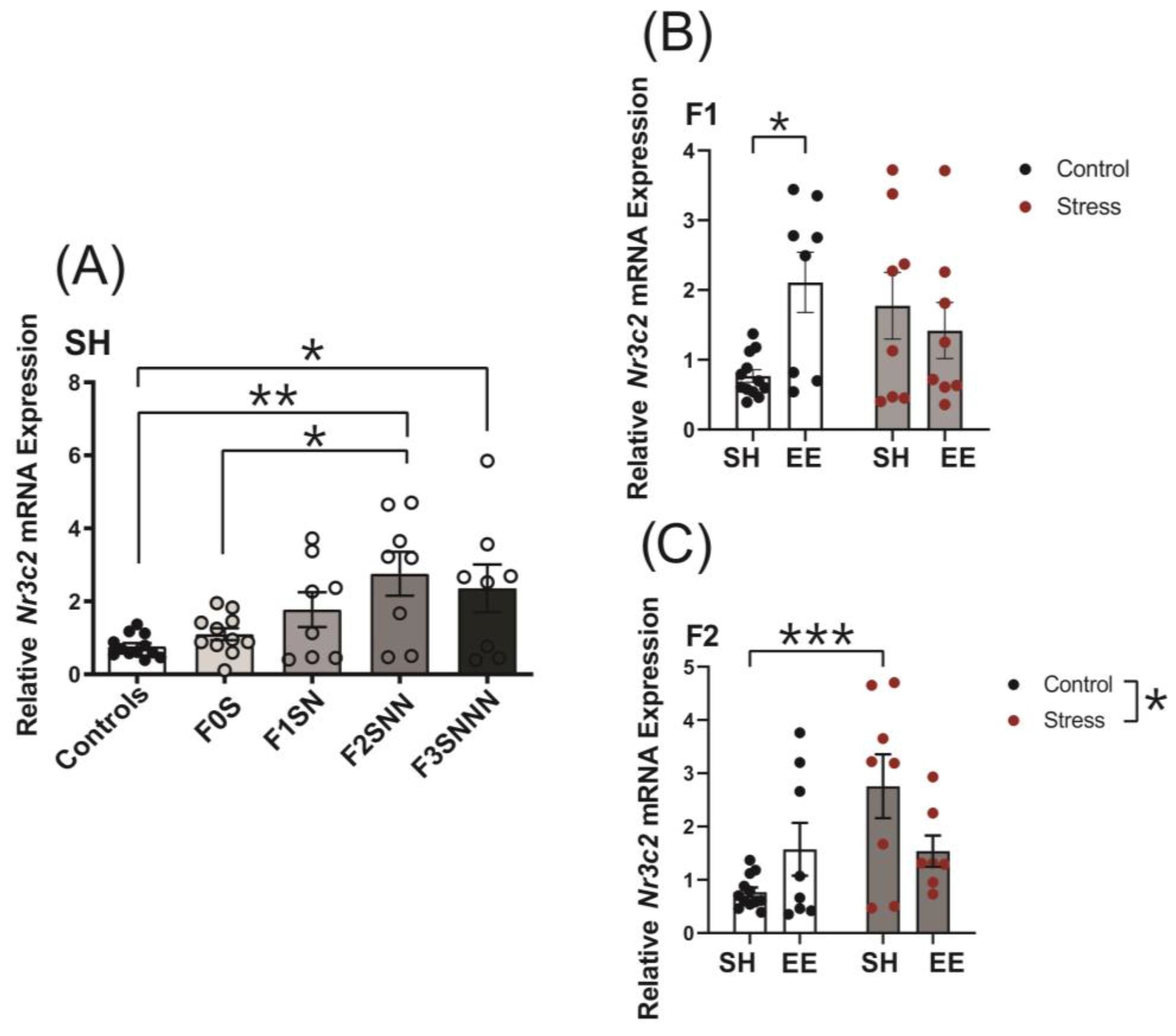
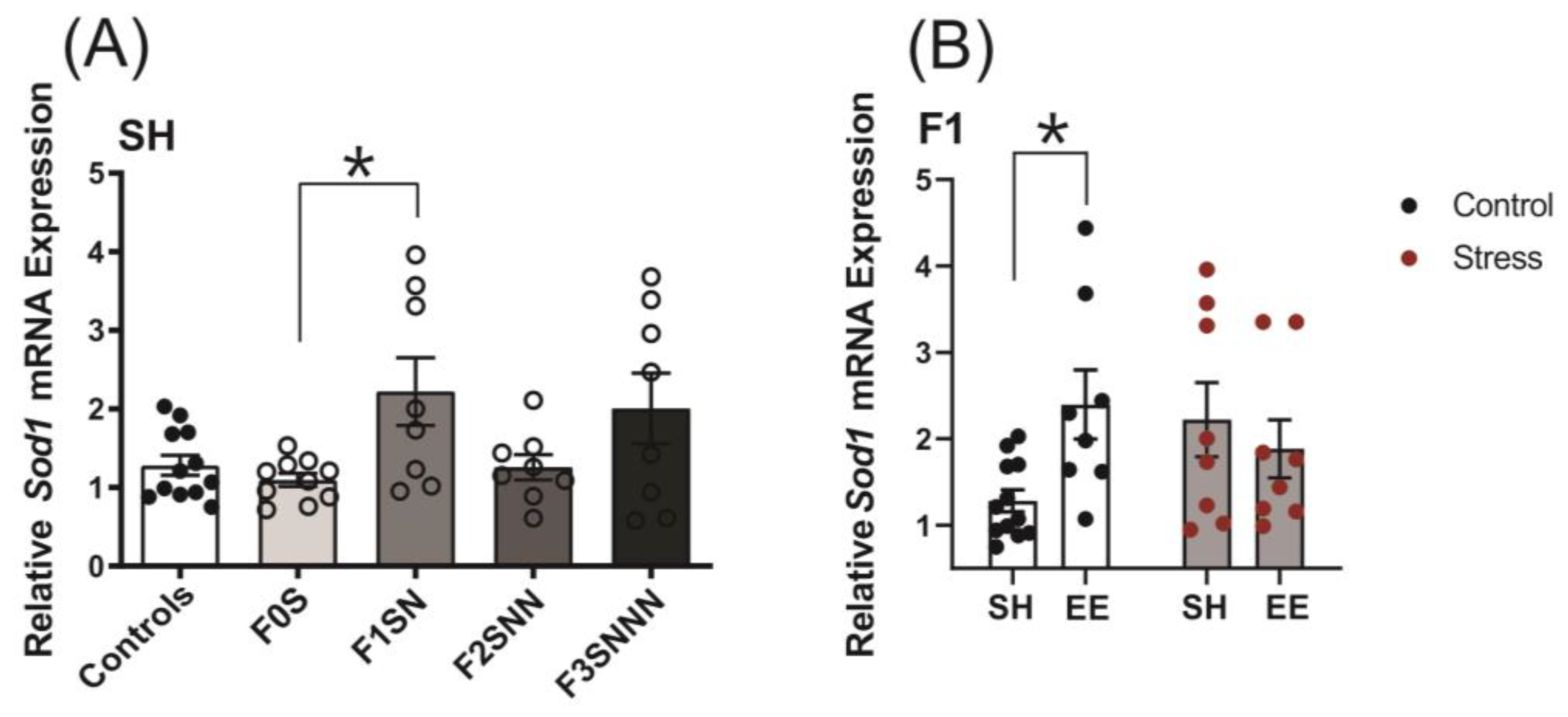
2.6. Uterine Expression of Candidate Genes Involved in Local Stress Responses Were Impacted in Rats Exposed to Ancestral Stress
2.6.1. 11β-Hydroxysteroid Dehydrogenase Type 2 (Hsd11b2)
2.6.2. Mineralocorticoid Receptor (Nr3c2)
2.6.3. Antioxidant Enzyme Gene Expression: Superoxide Dismutase Type 1 (Sod1)
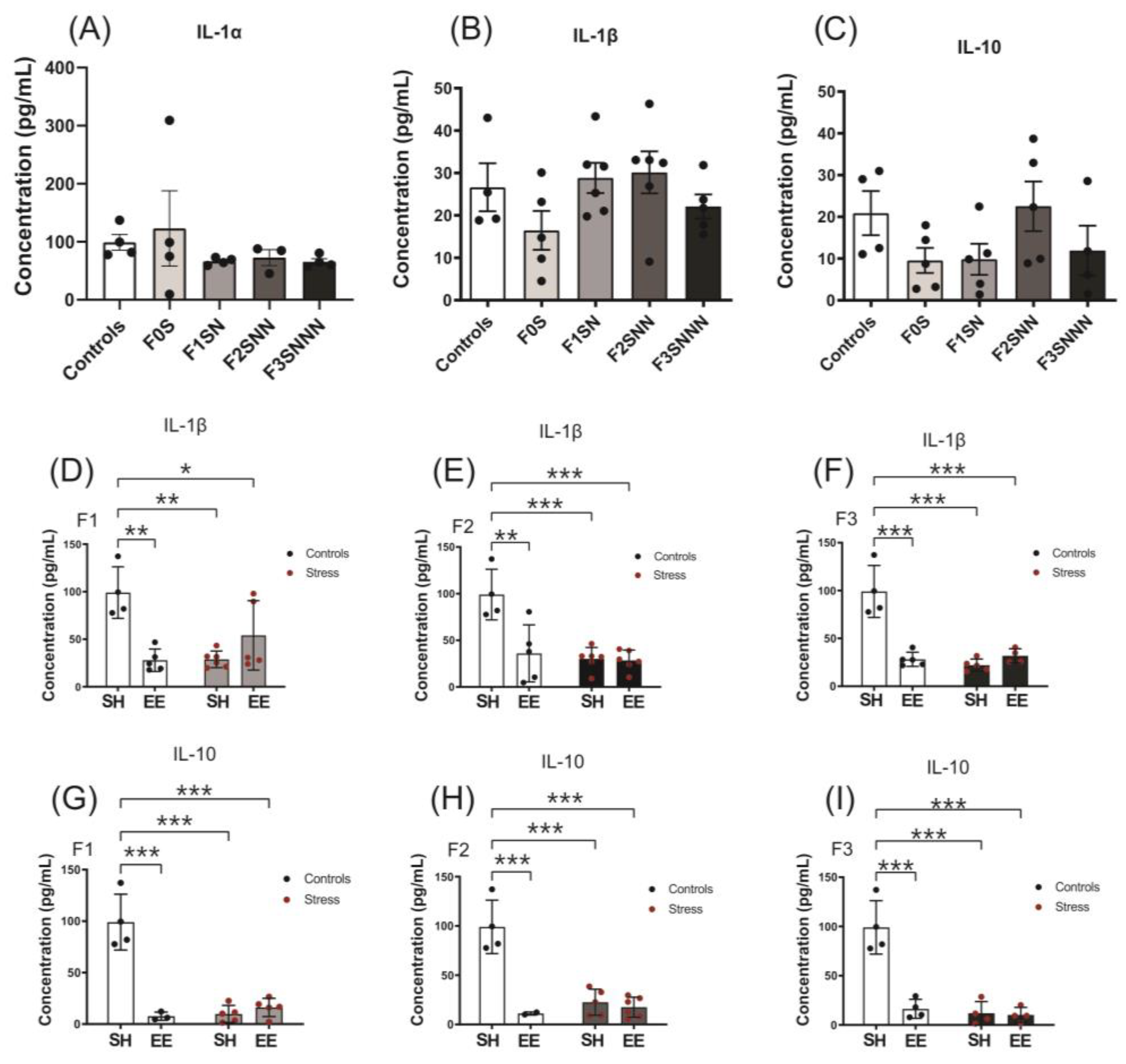
2.7. Uterine Concentration of IL-1α, IL-1β, IL-10, IL-6, and TNF-α Cytokines
3. Discussion
4. Materials and Methods
4.1. Animals
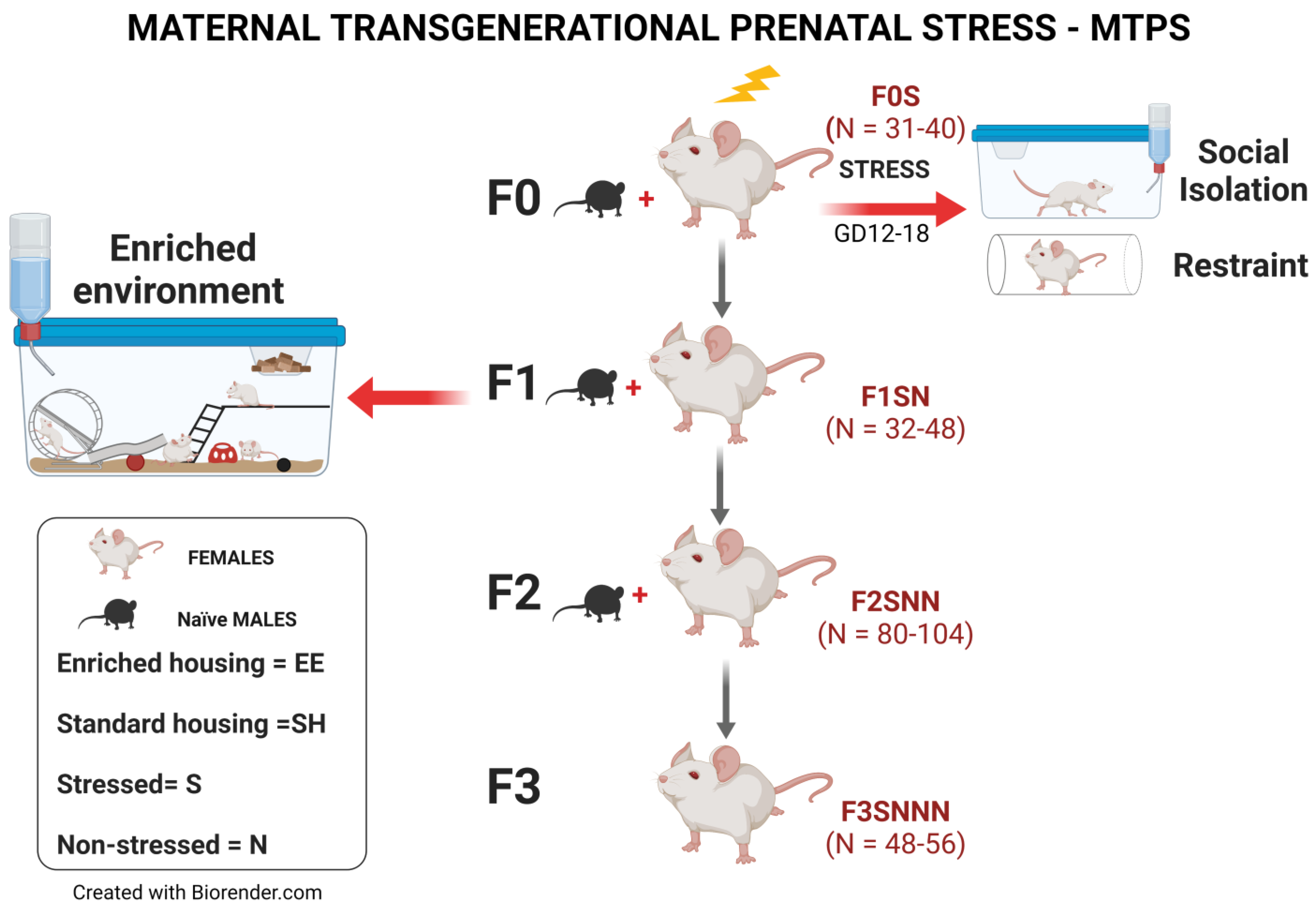
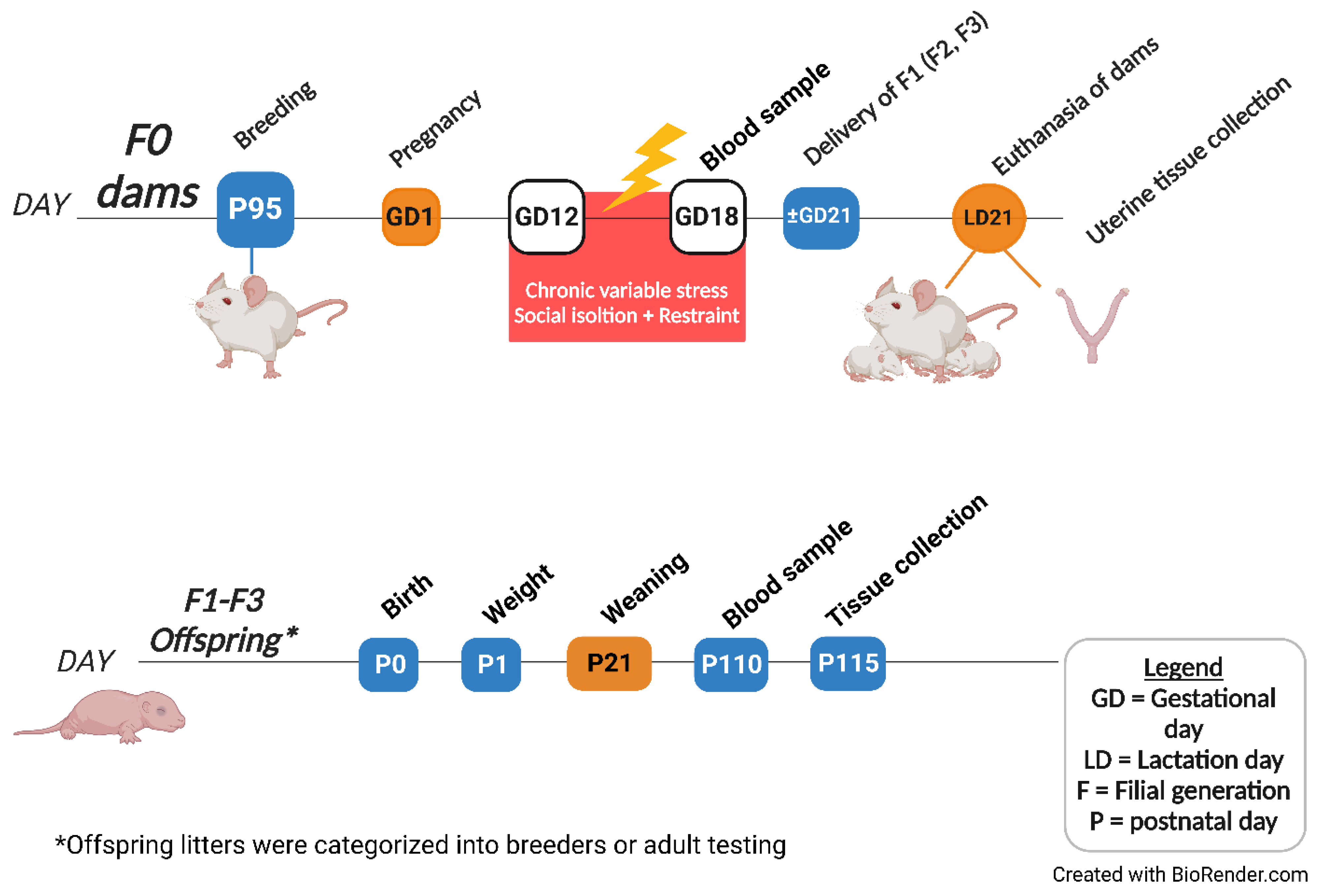
4.2. Experimental Design
4.3. Chronic Variable Stress (CVS) Procedures
4.4. Rearing Environments
4.5. Tissue Collection
4.5.1. Uterine Tissue
4.5.2. Blood
4.6. Molecular Analysis
4.6.1. RNA Extraction
4.6.2. Quantitative Real-Time Polymerase Chain Reaction (RT–qPCR)
4.7. Western Blot
4.8. Superoxide Detection Assay
4.9. Luminex Cytokine Assays
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 11β-hydroxysteroid dehydrogenase type 1 | 11β-HSD1 |
| 11β-hydroxysteroid dehydrogenase type 2 | 11β-HSD2 |
| Adverse pregnancy outcomes | APOs |
| Analysis of variance | ANOVA |
| Chronic variable stress | CVS |
| Complementary deoxyribonucleic acid | cDNA |
| Corticotropin releasing hormone | Crh |
| Corticotropin releasing hormone receptor 1 | Crhr1 |
| Corticotropin releasing hormone receptor 2 | Crhr2 |
| Cortisol/corticosterone | CORT |
| Cyclophilin A | Peptidilprolyl Isomerase A or Ppia |
| Dihydroethidium | DHE |
| Enriched environment | EE |
| Filial | F |
| Genomic deoxyribonucleic acid | DNA |
| Gestational day | GD |
| Glucocorticoid receptor | Nr3c1 |
| Glucocorticoids | GCs |
| Glyceraldehyde 3-phosphate dehydrogenase | GAPDH |
| Hypothalamic-pituitary-adrenal | HPA |
| Interleukin (IL)-1 receptor 1 | IL-1R1 |
| Interleukin (IL)-1 receptor accessory protein | IL-1RAP |
| Interleukin (IL)-10 | IL-10 |
| Interleukin (IL)-1α | IL-1α |
| Interleukin (IL)-1β | IL-1β |
| Interleukin (IL)-6 | IL-6 |
| Lactational day | LD |
| Mean fluorescence intensity | MFI |
| Mineralocorticoid receptor | Nr3c2 |
| Phosphate-buffered saline | PBS |
| Postnatal day | P |
| Prenatal maternal stress | PNMS |
| Quantitative Real-Time Polymerase Chain Reaction | RT–qPCR |
| Radio immunoprecipitation assay | RIPA |
| Reactive oxygen species | ROS |
| Standard housing | SH |
| Superoxide dismutase type 1 | Sod1 |
| Superoxide dismutase type 2 | Sod2 |
| Tumour necrosis factor-alpha | TNF-α |
References
- Hobel, C.J. Stress and preterm birth. Clin. Obstet. Gynecol. 2004, 47, 856–880. [Google Scholar] [CrossRef]
- Gennaro, S.; Hennessy, M.D. Psychological and physiological stress: Impact on preterm birth. J. Obstet. Gynecol. Neonatal. Nurs. 2003, 32, 668–675. [Google Scholar] [CrossRef]
- Yao, Y.; Robinson, A.M.; Zucchi, F.C.; Robbins, J.C.; Babenko, O.; Kovalchuk, O.; Kovalchuk, I.; Olson, D.M.; Metz, G.A. Ancestral exposure to stress epigenetically programs preterm birth risk and adverse maternal and newborn outcomes. BMC Med. 2014, 12, 121. [Google Scholar] [CrossRef]
- Garcia-Flores, V.; Romero, R.; Furcron, A.E.; Levenson, D.; Galaz, J.; Zou, C.; Hassan, S.S.; Hsu, C.D.; Olson, D.; Metz, G.A.S.; et al. Prenatal Maternal Stress Causes Preterm Birth and Affects Neonatal Adaptive Immunity in Mice. Front. Immunol. 2020, 11, 254. [Google Scholar] [CrossRef]
- Chen, H.J.; Antonson, A.M.; Rajasekera, T.A.; Patterson, J.M.; Bailey, M.T.; Gur, T.L. Prenatal stress causes intrauterine inflammation and serotonergic dysfunction, and long-term behavioral deficits through microbe- and CCL2-dependent mechanisms. Transl. Psychiatry 2020, 10, 191. [Google Scholar] [CrossRef]
- Traylor, C.S.; Johnson, J.D.; Kimmel, M.C.; Manuck, T.A. Effects of psychological stress on adverse pregnancy outcomes and nonpharmacologic approaches for reduction: An expert review. Am. J. Obstet. Gynecol. MFM 2020, 2, 100229. [Google Scholar] [CrossRef]
- Skinner, M.K. What is an epigenetic transgenerational phenotype? F3 or F2. Reprod. Toxicol. 2008, 25, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.J.; Kim, Y.J. What is fetal programming?: A lifetime health is under the control of in utero health. Obstet. Gynecol. Sci. 2017, 60, 506–519. [Google Scholar] [CrossRef]
- Barker, D.J. In utero programming of chronic disease. Clin. Sci. 1998, 95, 115–128. [Google Scholar] [CrossRef]
- Barker, D.J.; Gluckman, P.D.; Godfrey, K.M.; Harding, J.E.; Owens, J.A.; Robinson, J.S. Fetal nutrition and cardiovascular disease in adult life. Lancet 1993, 341, 938–941. [Google Scholar] [CrossRef]
- Skinner, M.K.; Manikkam, M.; Guerrero-Bosagna, C. Epigenetic transgenerational actions of environmental factors in disease etiology. Trends Endocrinol. Metab. 2010, 21, 214–222. [Google Scholar] [CrossRef]
- Nilsson, E.E.; Sadler-Riggleman, I.; Skinner, M.K. Environmentally induced epigenetic transgenerational inheritance of disease. Environ. Epigenet. 2018, 4, dvy016. [Google Scholar] [CrossRef]
- Veru, F.; Laplante, D.P.; Luheshi, G.; King, S. Prenatal maternal stress exposure and immune function in the offspring. Stress 2014, 17, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Creutzberg, K.C.; Sanson, A.; Viola, T.W.; Marchisella, F.; Begni, V.; Grassi-Oliveira, R.; Riva, M.A. Long-lasting effects of prenatal stress on HPA axis and inflammation: A systematic review and multilevel meta-analysis in rodent studies. Neurosci. Biobehav. Rev. 2021, 127, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.S.; Lydon, J.; Goulet, L.; Kahn, S.; Dahhou, M.; Platt, R.W.; Sharma, S.; Meaney, M.J.; Séguin, L. Maternal stress/distress, hormonal pathways and spontaneous preterm birth. Paediatr. Perinat. Epidemiol. 2013, 27, 237–246. [Google Scholar] [CrossRef]
- McGowan, P.O.; Matthews, S.G. Prenatal Stress, Glucocorticoids, and Developmental Programming of the Stress Response. Endocrinology 2018, 159, 69–82. [Google Scholar] [CrossRef]
- Lopes, N.A.; Falkenberg, E.A.; Wiley, C.; Patel, V.; Serrano-Lomelin, J.; Fang, X.; Weiler, A.M.; McCreary, J.K.; Metz, G.A.S.; Olson, D.M. Social Isolation Stress Modulates Pregnancy Outcomes and the Inflammatory Profile of Rat Uterus. Int. J. Mol. Sci. 2022, 23, 6169. [Google Scholar] [CrossRef] [PubMed]
- Howland, M.A.; Sandman, C.A.; Glynn, L.M. Developmental origins of the human hypothalamic-pituitary-adrenal axis. Expert Rev. Endocrinol. Metab. 2017, 12, 321–339. [Google Scholar] [CrossRef]
- Kapoor, A.; Dunn, E.; Kostaki, A.; Andrews, M.H.; Matthews, S.G. Fetal programming of hypothalamo-pituitary-adrenal function: Prenatal stress and glucocorticoids. J. Physiol. 2006, 572, 31–44. [Google Scholar] [CrossRef]
- Bollapragada, S.; Bollopragada, S.; Youssef, R.; Jordan, F.; Greer, I.; Norman, J.; Nelson, S. Term labor is associated with a core inflammatory response in human fetal membranes, myometrium, and cervix. Am. J. Obstet. Gynecol. 2009, 200, 104.e1. [Google Scholar] [CrossRef]
- Leimert, K.B.; Xu, W.; Princ, M.M.; Chemtob, S.; Olson, D.M. Inflammatory Amplification: A Central Tenet of Uterine Transition for Labor. Front. Cell. Infect. Microbiol. 2021, 11, 660983. [Google Scholar] [CrossRef]
- Coussons-Read, M.E.; Okun, M.L.; Nettles, C.D. Psychosocial stress increases inflammatory markers and alters cytokine production across pregnancy. Brain Behav. Immun. 2007, 21, 343–350. [Google Scholar] [CrossRef]
- Christian, L.M.; Glaser, R.; Porter, K.; Iams, J.D. Stress-induced inflammatory responses in women: Effects of race and pregnancy. Psychosom. Med. 2013, 75, 658–669. [Google Scholar] [CrossRef]
- Coussons-Read, M.E.; Okun, M.L.; Schmitt, M.P.; Giese, S. Prenatal stress alters cytokine levels in a manner that may endanger human pregnancy. Psychosom. Med. 2005, 67, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Coussons-Read, M.E.; Lobel, M.; Carey, J.C.; Kreither, M.O.; D’Anna, K.; Argys, L.; Ross, R.G.; Brandt, C.; Cole, S. The occurrence of preterm delivery is linked to pregnancy-specific distress and elevated inflammatory markers across gestation. Brain Behav. Immun. 2012, 26, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Dole, N.; Savitz, D.A.; Hertz-Picciotto, I.; Siega-Riz, A.M.; McMahon, M.J.; Buekens, P. Maternal stress and preterm birth. Am. J. Epidemiol. 2003, 157, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Ross, K.M.; Cole, S.W.; Carroll, J.E.; Schetter, C.D. Elevated pro-inflammatory gene expression in the third trimester of pregnancy in mothers who experienced stressful life events. Brain Behav. Immun. 2019, 76, 97–103. [Google Scholar] [CrossRef]
- Verstraeten, B.S.E.; McCreary, J.K.; Weyers, S.; Metz, G.A.S.; Olson, D.M. Prenatal two-hit stress affects maternal and offspring pregnancy outcomes and uterine gene expression in rats: Match or mismatch? Biol. Reprod. 2019, 100, 195–207. [Google Scholar] [CrossRef]
- Dowell, J.; Elser, B.A.; Schroeder, R.E.; Stevens, H.E. Cellular stress mechanisms of prenatal maternal stress: Heat shock factors and oxidative stress. Neurosci. Lett. 2019, 709, 134368. [Google Scholar] [CrossRef]
- Bittle, J.; Menezes, E.C.; McCormick, M.L.; Spitz, D.R.; Dailey, M.; Stevens, H.E. The Role of Redox Dysregulation in the Effects of Prenatal Stress on Embryonic Interneuron Migration. Cereb. Cortex 2019, 29, 5116–5130. [Google Scholar] [CrossRef] [PubMed]
- Bittle, J.; Stevens, H.E. The role of glucocorticoid, interleukin-1β, and antioxidants in prenatal stress effects on embryonic microglia. J. Neuroinflamm. 2018, 15, 44. [Google Scholar] [CrossRef] [PubMed]
- Karanikas, E.; Daskalakis, N.P.; Agorastos, A. Oxidative Dysregulation in Early Life Stress and Posttraumatic Stress Disorder: A Comprehensive Review. Brain Sci. 2021, 11, 723. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.A.; Ahmad, I.M.; Zimmerman, M.C. Oxidative Stress and Preterm Birth: An Integrative Review. Biol. Res. Nurs. 2018, 20, 497–512. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Bocking, A.D.; Unlugedik, E.; Challis, J.R. The effects of chorioamnionitis and betamethasone on 11beta hydroxysteroid dehydrogenase types 1 and 2 and the glucocorticoid receptor in preterm human placenta. J. Soc. Gynecol. Investig. 2005, 12, 238–245. [Google Scholar] [CrossRef]
- Kossintseva, I.; Wong, S.; Johnstone, E.; Guilbert, L.; Olson, D.M.; Mitchell, B.F. Pro-inflammatory cytokines inhibit human placental 11beta-hydroxysteroid dehydrogenase type 2 activity through Ca2+ and cAMP pathways. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E282–E288. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.; Lei, J.; Xu, T.; Zhao, M.; Cai, H.; Qiu, J.; Gao, Q. Effects of prenatal hypoxia on placental glucocorticoid barrier: Mechanistic insight from experiments in rats. Reprod. Toxicol. 2022, 110, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Alfaidy, N.; Gupta, S.; DeMarco, C.; Caniggia, I.; Challis, J.R.G. Oxygen Regulation of Placental 11β-Hydroxysteroid Dehydrogenase 2: Physiological and Pathological Implications. J. Clin. Endocrinol. Metab. 2002, 87, 4797–4805. [Google Scholar] [CrossRef]
- Hantsoo, L.; Kornfield, S.; Anguera, M.C.; Epperson, C.N. Inflammation: A Proposed Intermediary between Maternal Stress and Offspring Neuropsychiatric Risk. Biol. Psychiatry 2019, 85, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Schander, J.A.; Marvaldi, C.; Correa, F.; Wolfson, M.L.; Cella, M.; Aisemberg, J.; Jensen, F.; Franchi, A.M. Maternal environmental enrichment modulates the immune response against an inflammatory challenge during gestation and protects the offspring. J. Reprod. Immunol. 2021, 144, 103273. [Google Scholar] [CrossRef] [PubMed]
- McCreary, J.K.; Erickson, Z.T.; Hao, Y.; Ilnytskyy, Y.; Kovalchuk, I.; Metz, G.A. Environmental Intervention as a Therapy for Adverse Programming by Ancestral Stress. Sci. Rep. 2016, 6, 37814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, B.; Metz, G.A. Animal Models of Brain Plasticity and Behavioral Change. In Handbook of Neuropsychology; Elsevier: Amsterdam, The Netherlands, 2003; Volume 9, pp. 139–152. [Google Scholar]
- Schander, J.A.; Aisemberg, J.; Correa, F.; Wolfson, M.L.; Juriol, L.; Cymeryng, C.; Jensen, F.; Franchi, A.M. The enrichment of maternal environment prevents preterm birth in a mice model. Reproduction 2020, 159, 479–492. [Google Scholar] [CrossRef] [PubMed]
- McCreary, J.K.; Metz, G.A.S. Environmental enrichment as an intervention for adverse health outcomes of prenatal stress. Environ. Epigenet. 2016, 2, dvw013. [Google Scholar] [CrossRef]
- McCreary, J.K.; Erickson, Z.T.; Metz, G.A. Environmental enrichment mitigates the impact of ancestral stress on motor skill and corticospinal tract plasticity. Neurosci. Lett. 2016, 632, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Cooke, C.M.; Shah, A.; Kirschenman, R.D.; Quon, A.L.; Morton, J.S.; Care, A.S.; Davidge, S.T. Increased susceptibility to cardiovascular disease in offspring born from dams of advanced maternal age. J. Physiol. 2018, 596, 5807–5821. [Google Scholar] [CrossRef]
- Entringer, S.; Kumsta, R.; Nelson, E.L.; Hellhammer, D.H.; Wadhwa, P.D.; Wüst, S. Influence of prenatal psychosocial stress on cytokine production in adult women. Dev. Psychobiol. 2008, 50, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, E.B.; Kwee, L.; Nunes, P.; Shuster, D.J.; Ju, G.; McIntyre, K.W.; Chizzonite, R.A.; Labow, M.A. IL-1 receptor accessory protein is an essential component of the IL-1 receptor. J. Immunol. 1998, 161, 5614–5620. [Google Scholar] [CrossRef]
- Sivarajasingam, S.P.; Imami, N.; Johnson, M.R. Myometrial cytokines and their role in the onset of labour. J. Endocrinol. 2016, 231, R101–R119. [Google Scholar] [CrossRef] [PubMed]
- Lilliecreutz, C.; Larén, J.; Sydsjö, G.; Josefsson, A. Effect of maternal stress during pregnancy on the risk for preterm birth. BMC Pregnancy Childbirth 2016, 16, 5. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, P.D.; Entringer, S.; Buss, C.; Lu, M.C. The contribution of maternal stress to preterm birth: Issues and considerations. Clin. Perinatol. 2011, 38, 351–384. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, S.; Wang, G.; Hong, X.; Mallow, E.B.; Walker, S.O.; Pearson, C.; Heffner, L.; Zuckerman, B.; Wang, X. The combined association of psychosocial stress and chronic hypertension with preeclampsia. Am. J. Obstet. Gynecol. 2013, 209, 438.e1. [Google Scholar] [CrossRef] [Green Version]
- Vianna, P.; Bauer, M.E.; Dornfeld, D.; Chies, J.A. Distress conditions during pregnancy may lead to pre-eclampsia by increasing cortisol levels and altering lymphocyte sensitivity to glucocorticoids. Med. Hypotheses 2011, 77, 188–191. [Google Scholar] [CrossRef]
- Gur, T.L.; Shay, L.; Palkar, A.V.; Fisher, S.; Varaljay, V.A.; Dowd, S.; Bailey, M.T. Prenatal stress affects placental cytokines and neurotrophins, commensal microbes, and anxiety-like behavior in adult female offspring. Brain Behav. Immun. 2017, 64, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Bronson, S.L.; Bale, T.L. Prenatal stress-induced increases in placental inflammation and offspring hyperactivity are male-specific and ameliorated by maternal antiinflammatory treatment. Endocrinology 2014, 155, 2635–2646. [Google Scholar] [CrossRef] [PubMed]
- Faraji, J.; Singh, S.; Soltanpour, N.; Sutherland, R.J.; Metz, G.A.S. Environmental determinants of behavioural responses to short-term stress in rats: Evidence for inhibitory effect of ambient landmarks. Behav. Brain Res. 2020, 379, 112332. [Google Scholar] [CrossRef] [PubMed]
- Faraji, J.; Soltanpour, N.; Lotfi, H.; Moeeini, R.; Moharreri, A.R.; Roudaki, S.; Hosseini, S.A.; Olson, D.M.; Abdollahi, A.A.; Mohajerani, M.H.; et al. Lack of Social Support Raises Stress Vulnerability in Rats with a History of Ancestral Stress. Sci. Rep. 2017, 7, 5277. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.H.A.; Kendall, D.A.; Pardon, M.C. Prenatal exposure to chronic mild stress increases corticosterone levels in the amniotic fluid and induces cognitive deficits in female offspring, improved by treatment with the antidepressant drug amitriptyline. Behav. Brain Res. 2012, 231, 29–39. [Google Scholar] [CrossRef]
- Jafari, Z.; Mehla, J.; Afrashteh, N.; Kolb, B.E.; Mohajerani, M.H. Corticosterone response to gestational stress and postpartum memory function in mice. PLoS ONE 2017, 12, e0180306. [Google Scholar] [CrossRef]
- Bateson, P.; Gluckman, P.; Hanson, M. The biology of developmental plasticity and the Predictive Adaptive Response hypothesis. J. Physiol. 2014, 592, 2357–2368. [Google Scholar] [CrossRef] [PubMed]
- Daskalakis, N.P.; Bagot, R.C.; Parker, K.J.; Vinkers, C.H.; de Kloet, E.R. The three-hit concept of vulnerability and resilience: Toward understanding adaptation to early-life adversity outcome. Psychoneuroendocrinology 2013, 38, 1858–1873. [Google Scholar] [CrossRef] [PubMed]
- Gururajan, A.; van de Wouw, M.; Boehme, M.; Becker, T.; O’Connor, R.; Bastiaanssen, T.F.S.; Moloney, G.M.; Lyte, J.M.; Ventura Silva, A.P.; Merckx, B.; et al. Resilience to chronic stress is associated with specific neurobiological, neuroendocrine and immune responses. Brain Behav. Immun. 2019, 80, 583–594. [Google Scholar] [CrossRef]
- Van den Hove, D.L.; Kenis, G.; Brass, A.; Opstelten, R.; Rutten, B.P.; Bruschettini, M.; Blanco, C.E.; Lesch, K.P.; Steinbusch, H.W.; Prickaerts, J. Vulnerability versus resilience to prenatal stress in male and female rats; implications from gene expression profiles in the hippocampus and frontal cortex. Eur. Neuropsychopharmacol. 2013, 23, 1226–1246. [Google Scholar] [CrossRef] [PubMed]
- Boersma, G.J.; Tamashiro, K.L. Individual differences in the effects of prenatal stress exposure in rodents. Neurobiol. Stress 2015, 1, 100–108. [Google Scholar] [CrossRef]
- Franklin, T.B.; Russig, H.; Weiss, I.C.; Gräff, J.; Linder, N.; Michalon, A.; Vizi, S.; Mansuy, I.M. Epigenetic transmission of the impact of early stress across generations. Biol. Psychiatry 2010, 68, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, T.; Takeda, J.; Fang, X.; Bronson, H.; Olson, D.M. Interleukin (IL)-1 in rat parturition: IL-1 receptors 1 and 2 and accessory proteins abundance in pregnant rat uterus at term—Regulation by progesterone. Physiol. Rep. 2016, 4, e12866. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Li, R.; Xue, M. Potential regulatory network in the PSG10P/miR-19a-3p/IL1RAP pathway is possibly involved in preeclampsia pathogenesis. J. Cell. Mol. Med. 2019, 23, 852–864. [Google Scholar] [CrossRef]
- Braun, T.; Challis, J.R.; Newnham, J.P.; Sloboda, D.M. Early-life glucocorticoid exposure: The hypothalamic-pituitary-adrenal axis, placental function, and long-term disease risk. Endocr. Rev. 2013, 34, 885–916. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Zhu, P.; Myatt, L.; Sun, K. Roles of glucocorticoids in human parturition: A controversial fact? Placenta 2014, 35, 291–296. [Google Scholar] [CrossRef]
- Pofi, R.; Tomlinson, J.W. Glucocorticoids in pregnancy. Obstet. Med. 2020, 13, 62–69. [Google Scholar] [CrossRef]
- Muñoz-Durango, N.; Vecchiola, A.; Gonzalez-Gomez, L.M.; Simon, F.; Riedel, C.A.; Fardella, C.E.; Kalergis, A.M. Modulation of Immunity and Inflammation by the Mineralocorticoid Receptor and Aldosterone. BioMed Res. Int. 2015, 2015, 652738. [Google Scholar] [CrossRef]
- Bînă, A.M.; Sturza, A.; Iancu, I.; Mocanu, A.G.; Bernad, E.; Chiriac, D.V.; Borza, C.; Craina, M.L.; Popa, Z.L.; Muntean, D.M.; et al. Placental oxidative stress and monoamine oxidase expression are increased in severe preeclampsia: A pilot study. Mol. Cell. Biochem. 2022, 477, 2851–2861. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Lu, J.; Wang, Z.; Cao, J.; Chen, Y.; Dong, Y. A novel and compact review on the role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2018, 16, 80. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, M.; Ding, S.; Li, C.; Luo, X. Environmental Enrichment during Gestation Improves Behavior Consequences and Synaptic Plasticity in Hippocampus of Prenatal-Stressed Offspring Rats. Acta Histochem. Cytochem. 2012, 45, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.W.; Park, C.H.; Choi, S.H.; Kim, N.J.; Kim, H.S.; Choe, J.C.; Suh, Y.H. The postnatal environment can counteract prenatal effects on cognitive ability, cell proliferation, and synaptic protein expression. FASEB J. 2003, 17, 1556–1558. [Google Scholar] [CrossRef]
- Falkenberg, T.; Mohammed, A.K.; Henriksson, B.; Persson, H.; Winblad, B.; Lindefors, N. Increased expression of brain-derived neurotrophic factor mRNA in rat hippocampus is associated with improved spatial memory and enriched environment. Neurosci. Lett. 1992, 138, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Galani, R.; Berthel, M.C.; Lazarus, C.; Majchrzak, M.; Barbelivien, A.; Kelche, C.; Cassel, J.C. The behavioral effects of enriched housing are not altered by serotonin depletion but enrichment alters hippocampal neurochemistry. Neurobiol. Learn. Mem. 2007, 88, 1–10. [Google Scholar] [CrossRef]
- McCreary, J.K.; Truica, L.S.; Friesen, B.; Yao, Y.; Olson, D.M.; Kovalchuk, I.; Cross, A.R.; Metz, G.A. Altered brain morphology and functional connectivity reflect a vulnerable affective state after cumulative multigenerational stress in rats. Neuroscience 2016, 330, 79–89. [Google Scholar] [CrossRef]
- Leshem, M.; Schulkin, J. Transgenerational effects of infantile adversity and enrichment in male and female rats. Dev. Psychobiol. 2012, 54, 169–186. [Google Scholar] [CrossRef]
- Turschwell, M.P.; White, C.R. The effects of laboratory housing and spatial enrichment on brain size and metabolic rate in the eastern mosquitofish, Gambusia holbrooki. Biol. Open 2016, 5, 205–210. [Google Scholar] [CrossRef]
- Toli, E.A.; Noreikiene, K.; DeFaveri, J.; Merilä, J. Environmental enrichment, sexual dimorphism, and brain size in sticklebacks. Ecol. Evol. 2017, 7, 1691–1698. [Google Scholar] [CrossRef]
- Fairhurst, G.D.; Frey, M.D.; Reichert, J.F.; Szelest, I.; Kelly, D.M.; Bortolotti, G.R. Does environmental enrichment reduce stress? An integrated measure of corticosterone from feathers provides a novel perspective. PLoS ONE 2011, 6, e17663. [Google Scholar] [CrossRef]
- Moncek, F.; Duncko, R.; Johansson, B.B.; Jezova, D. Effect of environmental enrichment on stress related systems in rats. J. Neuroendocrinol. 2004, 16, 423–431. [Google Scholar] [CrossRef]
- Svensson, M.; Rosvall, P.; Boza-Serrano, A.; Andersson, E.; Lexell, J.; Deierborg, T. Forced treadmill exercise can induce stress and increase neuronal damage in a mouse model of global cerebral ischemia. Neurobiol. Stress 2016, 5, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.; Mach, N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: A systematic review for athletes. J. Int. Soc. Sports Nutr. 2016, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- van Praag, H.; Kempermann, G.; Gage, F.H. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat. Neurosci. 1999, 2, 266–270. [Google Scholar] [CrossRef]
- Tzu-Feng Wang, T.F.; Tsai, S.F.; Zhao, Z.W.; Shih, M.M.; Wang, C.Y.; Yang, T.T.; Kuo, Y.M. Exercise-induced increases of corticosterone contribute to exercise-enhanced adult hippocampal neurogenesis in mice. Chin. J. Physiol. 2021, 64, 186–193. [Google Scholar] [CrossRef]
- Akhund-Zade, J.; Ho, S.; O’Leary, C.; de Bivort, B. The effect of environmental enrichment on behavioral variability depends on genotype, behavior, and type of enrichment. J. Exp. Biol. 2019, 222, jeb202234. [Google Scholar] [CrossRef] [PubMed]
- Körholz, J.C.; Zocher, S.; Grzyb, A.N.; Morisse, B.; Poetzsch, A.; Ehret, F.; Schmied, C.; Kempermann, G. Selective increases in inter-individual variability in response to environmental enrichment in female mice. Elife 2018, 7, e35690. [Google Scholar] [CrossRef] [PubMed]
- Lacal, I.; Ventura, R. Epigenetic Inheritance: Concepts, Mechanisms and Perspectives. Front. Mol. Neurosci. 2018, 11, 292. [Google Scholar] [CrossRef]
- Burggren, W. Epigenetic Inheritance and Its Role in Evolutionary Biology: Re-Evaluation and New Perspectives. Biology 2016, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Leimert, K.B.; Messer, A.; Gray, T.; Fang, X.; Chemtob, S.; Olson, D.M. Maternal and fetal intrauterine tissue crosstalk promotes pro-inflammatory amplification and uterine transition †. Biol. Reprod. 2019, 100, 783–797. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gestational Day | G-12 | G-13 | G-14 | G-15 | G-16 | G-17 | G-18 |
|---|---|---|---|---|---|---|---|
| Treatment (am) | Restraint 60 min | N/A | Restraint 45 min | N/A | Restraint 15 min | N/A | Restraint 45 min |
| Treatment (pm) | N/A | Restraint 30 min | Isolation Overnight | Restraint 60 min | N/A | Restraint 30 min | N/A |
| Isolation Overnight |
| Target Gene | Forward Primer (5′ → 3′) | Reverse Primer (5′ → 3′) | Annealing Temperature (°C) | NCBI Reference Sequences * |
|---|---|---|---|---|
| Ppia (Cyclophilin A) | CAC CGT GTT CTT CGA CAT CAC | CCA GTG CTC AGA GCT CGA AAG | 60 | NM_017101.1 |
| Il1a | AAGACAAGCCTGTGTTGCTGAAGG | TCCCAGAAGAAAATGAGGTCGGTC | 55 | NM_017019.1 |
| Il1b | CTCAATGGACAGAACATAAGCC | GGTGTGCCGTCTTTCATCA | 51 | NM_031512.2 |
| Il6 | TCCTACCCCAACTTCCAATGCTC | TTGGATGGTXTTGGTCCTTAGCC | 65 | NM_012589.2 |
| Il1ra | AAGACCTTCTACCTGAGGAACAACC | GCCCAAGAACACATTCCGAAAGTC | 55 | NM_022194.2 |
| Il1rap | GGGCAACATCAACGTCATTTTAG | CAGCTCTTTCACCTTCAAGTCCTT | 68 | NM_012968.1 |
| Hsd11b1 | GAAGAAGCATGGAGGTCAAC | GCAATCAGAGGTTGGGTCAT | 60 | NM_017080.2 |
| Hsd11b2 | CGTCACTCAAGGGGACGTAT | AGGGGTATGGCATGTCTCC | 55 | NM_017081.2 |
| Crh | ATCTCACCTTCCACCTTCTG | GTGTGCTAAATGCAGAATCG | 60 | NM_031019.1 |
| Crhr1 | GGTGACAGCCGCCTACAATT | AAGGTACACCCCAGCCAA | 60 | NM_030999.4 |
| Crhr2 | TGGTGCATACCCTGCCCTAT | GTGGAGGCTCGCAGTTTTGT | 60 | NM_022714.1 |
| Nr3c1 | TGTATCCCACAGACCAAAGCA | AATCCTCATTCGTGTTCCCTTC | 52 | NM_012576.2 |
| Nr3c2 | GGCAAACAGATGATCCAGG | CAACTCAAAGCGAACGATGA | 60 | NM_013131.1 |
| Sod1 | GCAGAAGGCAAGCGGTGA | GGTACAGCCTTGTGTATTGTC CC | 60 | NM_017050.1 |
| Sod2 | GTCTGTGGGAGTCCAAGGTT | GTTCCTTGCAGTGGGTCCTGATTA | 60 | NM_017051.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, N.A.; Ambeskovic, M.; King, S.E.; Faraji, J.; Soltanpour, N.; Falkenberg, E.A.; Scheidl, T.; Patel, M.; Fang, X.; Metz, G.A.S.; et al. Environmental Enrichment Promotes Transgenerational Programming of Uterine Inflammatory and Stress Markers Comparable to Gestational Chronic Variable Stress. Int. J. Mol. Sci. 2023, 24, 3734. https://doi.org/10.3390/ijms24043734
Lopes NA, Ambeskovic M, King SE, Faraji J, Soltanpour N, Falkenberg EA, Scheidl T, Patel M, Fang X, Metz GAS, et al. Environmental Enrichment Promotes Transgenerational Programming of Uterine Inflammatory and Stress Markers Comparable to Gestational Chronic Variable Stress. International Journal of Molecular Sciences. 2023; 24(4):3734. https://doi.org/10.3390/ijms24043734
Chicago/Turabian StyleLopes, Nayara A., Mirela Ambeskovic, Stephanie E. King, Jamshid Faraji, Nasrin Soltanpour, Erin A. Falkenberg, Taylor Scheidl, Mansi Patel, Xin Fang, Gerlinde A. S. Metz, and et al. 2023. "Environmental Enrichment Promotes Transgenerational Programming of Uterine Inflammatory and Stress Markers Comparable to Gestational Chronic Variable Stress" International Journal of Molecular Sciences 24, no. 4: 3734. https://doi.org/10.3390/ijms24043734
APA StyleLopes, N. A., Ambeskovic, M., King, S. E., Faraji, J., Soltanpour, N., Falkenberg, E. A., Scheidl, T., Patel, M., Fang, X., Metz, G. A. S., & Olson, D. M. (2023). Environmental Enrichment Promotes Transgenerational Programming of Uterine Inflammatory and Stress Markers Comparable to Gestational Chronic Variable Stress. International Journal of Molecular Sciences, 24(4), 3734. https://doi.org/10.3390/ijms24043734