
Vitamin D3 Metabolism and Its Role in Temporomandibular Joint Osteoarthritis and Autoimmune Thyroid Diseases


Abstract
:1. Introduction
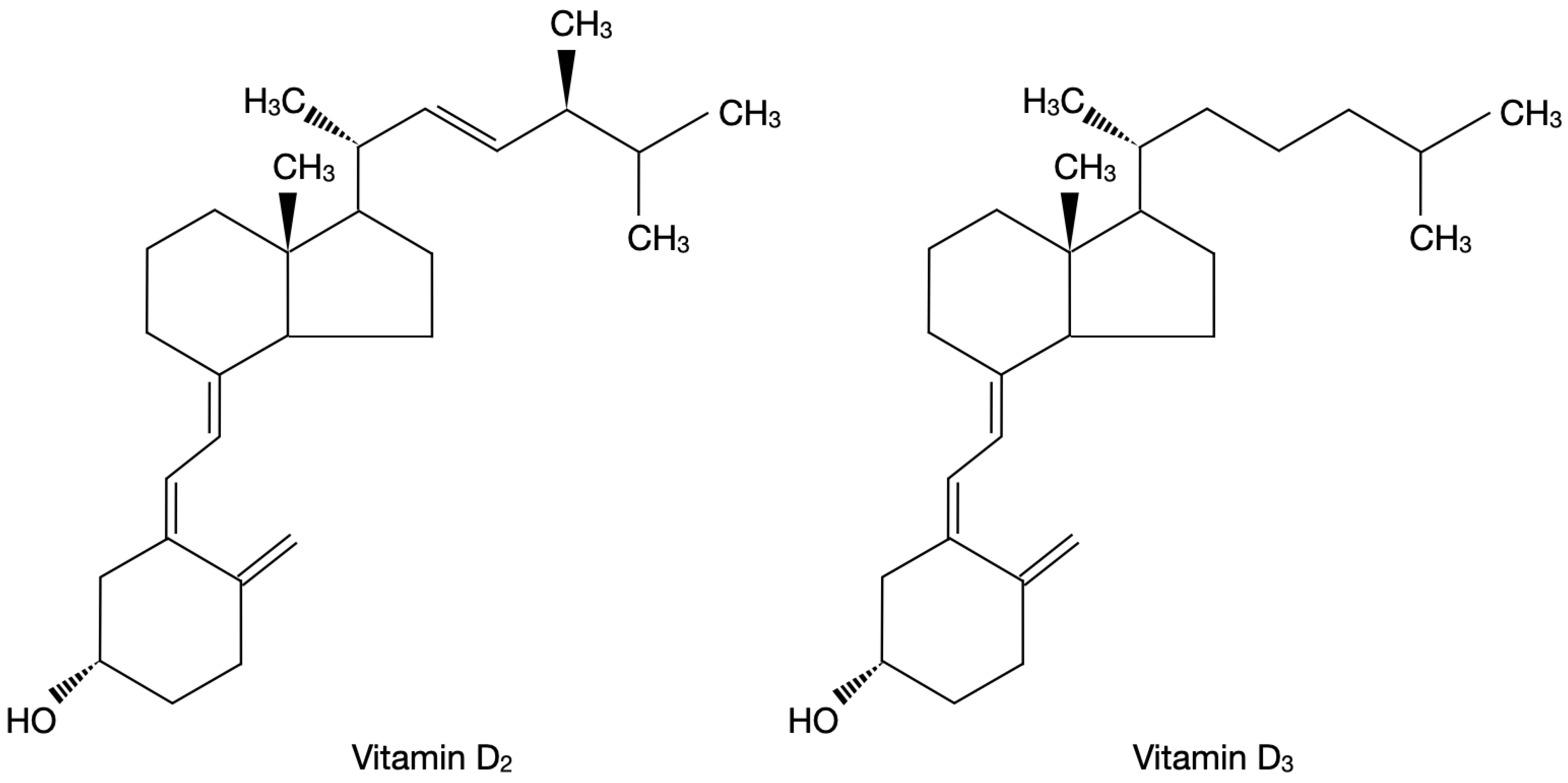
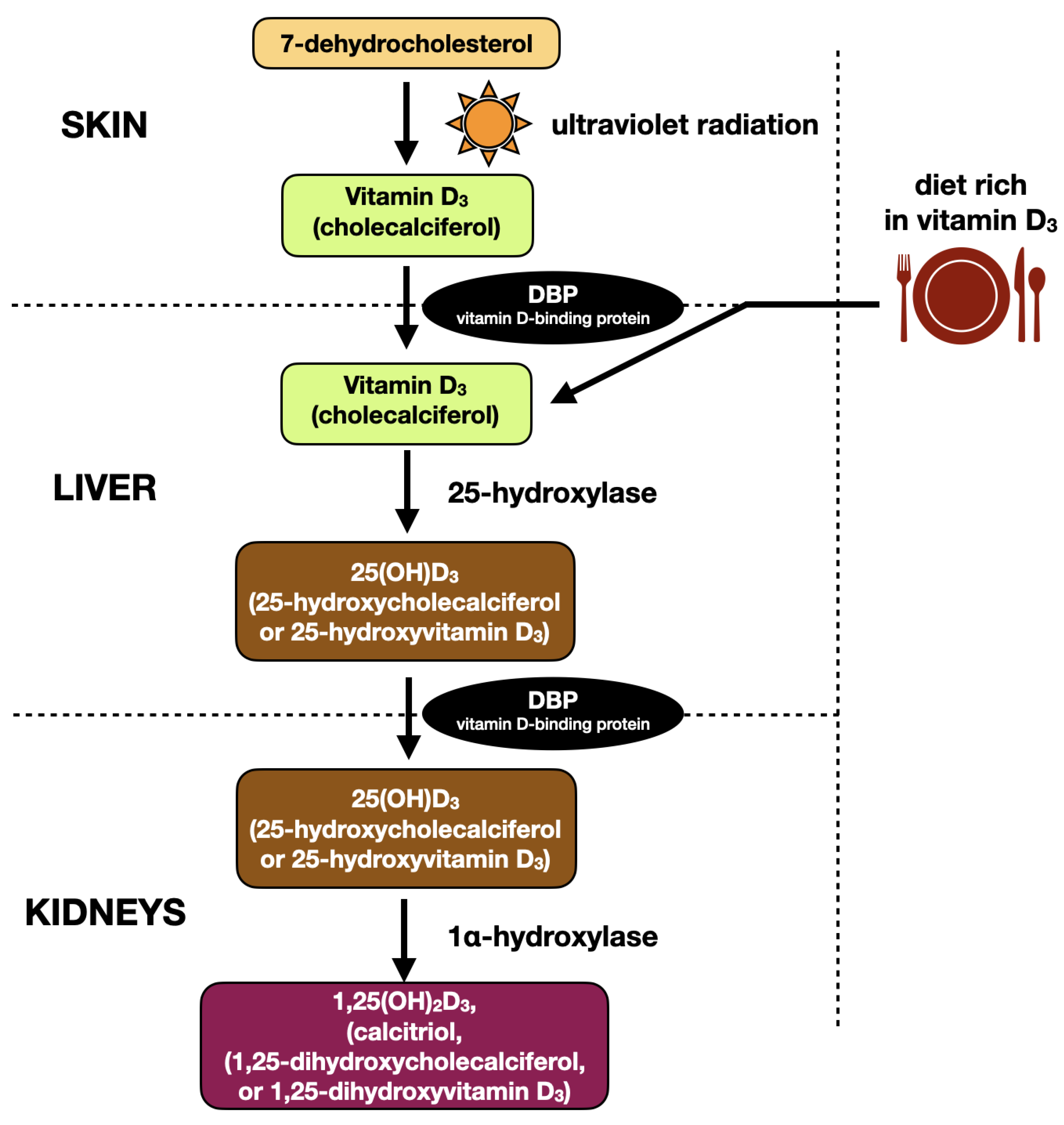
2. Vitamin D3—Biochemical Structure and Metabolism
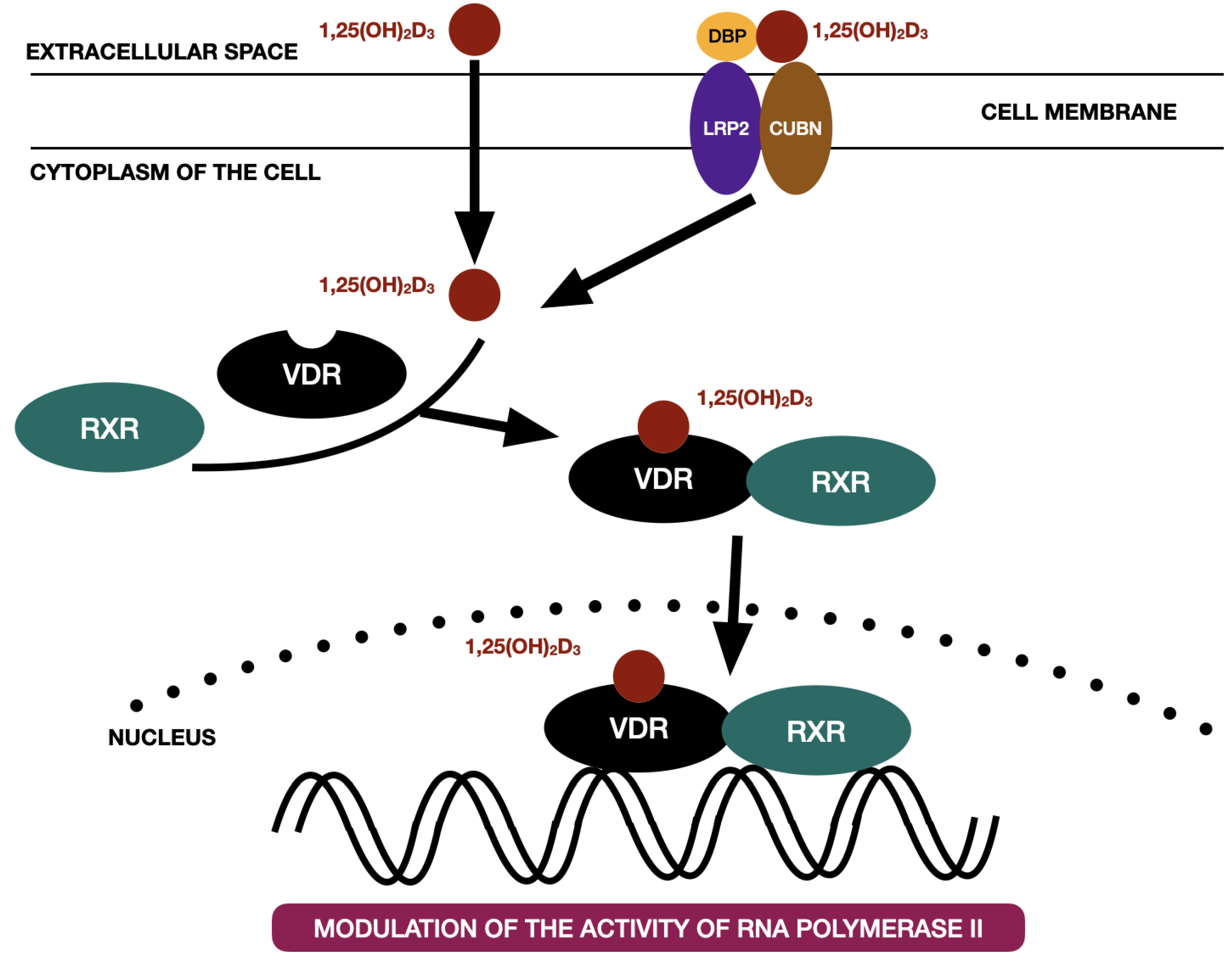
3. Vitamin D3 Receptor and Mechanisms of Action
4. Vitamin D3 and Immune System
5. Role of Vitamin D3 in Autoimmune Thyroid Diseases
6. Vitamin D3 and Bone Metabolism
7. Vitamin D3 and Osteoarthritis
8. Vitamin D3 and Temporomandibular Joint Osteoarthritis
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lumachi, F.; Motta, R.; Cecchin, D.; Ave, S.; Camozzi, V.; Basso, S.M.; Luisetto, G. Calcium metabolism & hypercalcemia in adults. Curr. Med. Chem. 2011, 18, 3529–3536. [Google Scholar] [CrossRef] [PubMed]
- Creo, A.L.; Thacher, T.; Pettifor, J.; Strand, M.A.; Fischer, P.R. Nutritional rickets around the world: An update. Ann. Trop. Paediatr. 2016, 37, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Bhattoa, H.P.; Konstantynowicz, J.; Laszcz, N.; Wojcik, M.; Pludowski, P. Vitamin D: Musculoskeletal health. Rev. Endocr. Metab. Disord. 2017, 18, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Watts, N.B.; Camacho, P.M.; Lewiecki, E.M.; Petak, S.M. AACE/ACE Postmenopausal Osteoporosis Guidelines Task Force. American Association of Clinical Endocrinologists/American College of Endocrinology Clinical Practice Guidelines for the Diagnosis and Treatment of Postmenopausal Osteoporosis-2020 Update. Endocr. Pract. 2021, 27, 379–380. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wu, W.; Gu, J.; Zhang, X.; Dang, J.; Wang, J.; Zheng, Y.; Huang, F.; Yuan, J.; Xue, Y.; et al. Human gingival tissue-derived MSC suppress osteoclastogenesis and bone erosion via CD39-adenosine signal pathway in autoimmune arthritis. Ebiomedicine 2019, 43, 620–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, N.; Lan, Q.; Chen, M.; Zheng, T.; Su, W.; Wang, J.; Yang, Z.; Park, R.; Dagliyan, G.; Conti, P.S.; et al. Induced T regulatory cells suppress osteoclastogenesis and bone erosion in collagen-induced arthritis better than natural T regulatory cells. Ann. Rheum. Dis. 2012, 71, 1567–1572. [Google Scholar] [CrossRef] [Green Version]
- Bellan, M.; Andreoli, L.; Mele, C.; Sainaghi, P.P.; Rigamonti, C.; Piantoni, S.; De Benedittis, C.; Aimaretti, G.; Pirisi, M.; Marzullo, P. Pathophysiological Role and Therapeutic Implications of Vitamin D in Autoimmunity: Focus on Chronic Autoimmune Diseases. Nutrients 2020, 12, 789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchemal, M.; Hakem, D.; Azzouz, M.; Touil-Boukoffa, C.; Mezioug, D. Vitamin D Levels Correlate with Metabolic Syndrome Criteria in Algerian Patients: The Ex-vivo Immunomodulatory Effect of α, 25 Dihydroxyvitamin D3. Endocr. Metab. Immune Disord Drug Targets 2020, 20, 1282–1294. [Google Scholar] [CrossRef]
- Song, L. Calcium and Bone Metabolism Indices. Adv. Clin. Chem. 2017, 82, 1–46. [Google Scholar] [CrossRef]
- Zakeri, M.; Parsian, H.; Bijani, A.; Shirzad, A.; Neamati, N. Serum levels of vitamin D in patients with recurrent aphthous stomatitis. Dent. Med. Probl. 2021, 58, 27–30. [Google Scholar] [CrossRef]
- Krawiec, M.; Dominiak, M. Prospective evaluation of vitamin D levels in dental treated patients: A screening study. Dent. Med. Probl. 2021, 58, 321–326. [Google Scholar] [CrossRef]
- Zhao, R.; Zhang, W.; Ma, C.; Zhao, Y.; Xiong, R.; Wang, H.; Chen, W.; Zheng, S.G. Immunomodulatory Function of Vitamin D and Its Role in Autoimmune Thyroid Disease. Front. Immunol. 2021, 12, 574967. [Google Scholar] [CrossRef] [PubMed]
- Kim, D. The Role of Vitamin D in Thyroid Diseases. Int. J. Mol. Sci. 2017, 18, 1949. [Google Scholar] [CrossRef] [Green Version]
- Vieira, I.H.; Rodrigues, D.; Paiva, I. Vitamin D and Autoimmune Thyroid Disease-Cause, Consequence, or a Vicious Cycle? Nutrients 2020, 12, 2791. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Ferrari, S.M.; Corrado, A.; Di Domenicantonio, A.; Fallahi, P. Autoimmune thyroid disorders. Autoimmun. Rev. 2015, 14, 174–180. [Google Scholar] [CrossRef]
- Saravanan, P.; Dayan, C.M. Thyroid autoantibodies. Endocrinol. Metab. Clin. N. Am. 2001, 30, 315–337. [Google Scholar] [CrossRef]
- Takedani, K.; Notsu, M.; Yamauchi, M.; Nawata, K.; Sugimoto, T.; Kanasaki, K. Graves’ disease and vertebral fracture: Possible pathogenic link in postmenopausal women. Clin. Endocrinol. 2020, 93, 204–211. [Google Scholar] [CrossRef]
- Wu, J.; Huang, H.; Yu, X. How does Hashimoto’s thyroiditis affect bone metabolism? Rev. Endocr. Metab. Disord. 2022. Epub ahead of print. [Google Scholar]
- Charoenngam, N. Vitamin D and Rheumatic Diseases: A Review of Clinical Evidence. Int. J. Mol. Sci. 2021, 22, 10659. [Google Scholar] [CrossRef]
- Li, H.-M.; Liu, Y.; Zhang, R.-J.; Ding, J.-Y.; Shen, C.-L. Vitamin D receptor gene polymorphisms and osteoarthritis: A meta-analysis. Rheumatology 2021, 60, 538–548. [Google Scholar] [CrossRef]
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Bergink, A.P.; Zillikens, M.C.; Van Leeuwen, J.P.; Hofman, A.; Uitterlinden, A.G.; van Meurs, J.B. 25-Hydroxyvitamin D and osteoarthritis: A meta-analysis including new data. Semin. Arthritis Rheum. 2016, 45, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, S.K.; Gantaguru, A.; Nanda, S.N.; Velagada, S.; Srinivasan, A.; Mangaraj, M. Association of vitamin D and knee osteoarthritis in younger individuals. World J. Orthop. 2020, 11, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Derwich, M.; Mitus-Kenig, M.; Pawlowska, E. Interdisciplinary Approach to the Temporomandibular Joint Osteoarthritis—Review of the Literature. Medicina 2020, 56, 225. [Google Scholar] [CrossRef] [PubMed]
- Derwich, M.; Mitus-Kenig, M.; Pawlowska, E. Morphology of the Temporomandibular Joints Regarding the Presence of Osteoarthritic Changes. Int. J. Environ. Res. Public Health 2020, 17, 2923. [Google Scholar] [CrossRef] [Green Version]
- Ferrillo, M.; Lippi, L.; Giudice, A.; Calafiore, D.; Paolucci, T.; Renò, F.; Migliario, M.; Fortunato, L.; Invernizzi, M.; de Sire, A. Temporomandibular Disorders and Vitamin D Deficiency: What Is the Linkage between These Conditions? A Systematic Review. J. Clin. Med. 2022, 11, 6231. [Google Scholar] [CrossRef]
- Tuckey, R.C.; Cheng, C.; Slominski, A.T. The serum vitamin D metabolome: What we know and what is still to discover. J. Steroid Biochem. Mol. Biol. 2019, 186, 4–21. [Google Scholar] [CrossRef]
- Żmijewski, M.A. Nongenomic Activities of Vitamin, D. Nutrients 2022, 14, 5104. [Google Scholar] [CrossRef]
- Zhu, J.; DeLuca, H.F. Vitamin D 25-hydroxylase-Four decades of searching, are we there yet? Arch Biochem. Biophys. 2012, 523, 30–36. [Google Scholar] [CrossRef]
- Jones, G.; Prosser, D.E.; Kaufmann, M. Cytochrome P450-mediated metabolism of vitamin D. J. Lipid Res. 2014, 55, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Laird, E.; Ward, M.; McSorley, E.; Strain, J.; Wallace, J. Vitamin D and Bone Health; Potential Mechanisms. Nutrients 2010, 2, 693–724. [Google Scholar] [CrossRef] [Green Version]
- Bouillon, R.; Schuit, F.; Antonio, L.; Rastinejad, F. Vitamin D Binding Protein: A Historic Overview. Front. Endocrinol. 2020, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Farrell, C.-J.L.; Pusceddu, I.; Fabregat-Cabello, N.; Cavalier, E. Assessment of vitamin D status–a changing landscape. Clin. Chem. Lab. Med. 2017, 55, 3–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad, J.G.; Hillman, L.; Rojanasathit, S. Human Serum Binding Capacity and Affinity for 25-Hydroxyergocalciferol and 25-Hydroxycholecalciferol. J. Clin. Endocrinol. Metab. 1976, 43, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.S.; Hewison, M. Extrarenal expression of the 25-hydroxyvitamin D-1-hydroxylase. Arch. Biochem. Biophys. 2012, 523, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikle, D.D. Vitamin D metabolism, mechanism of action, and clinical applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [Green Version]
- Zmijewski, M.A.; Carlberg, C. Vitamin D receptor(s): In the nucleus but also at membranes? Exp Dermatol. 2020, 29, 876–884. [Google Scholar] [CrossRef]
- Bourguet, W.; Germain, P.; Gronemeyer, H. Nuclear receptor ligand-binding domains: Three-dimensional structures, molecular interactions and pharmacological implications. Trends Pharmacol. Sci. 2000, 21, 381–388. [Google Scholar] [CrossRef]
- Rochel, N.; Wurtz, J.; Mitschler, A.; Klaholz, B.; Moras, D. The Crystal Structure of the Nuclear Receptor for Vitamin D Bound to Its Natural Ligand. Mol. Cell 2000, 5, 173–179. [Google Scholar] [CrossRef]
- Van de Peppel, J.; van Leeuwen, J.P. Vitamin D and gene networks in human osteoblasts. Front. Physiol. 2014, 5, 137. [Google Scholar] [CrossRef] [Green Version]
- Carlberg, C. Vitamin D and Its Target Genes. Nutrients 2022, 14, 1354. [Google Scholar] [CrossRef] [PubMed]
- Civitelli, R.; Kim, Y.S.; Gunsten, S.L.; Fujimori, A.; Huskey, M.; Avioli, L.V.; Hruska, K.A. Nongenomic Activation of the Calcium Message System by Vitamin D Metabolites in Osteoblast-like Cells. Endocrinology 1990, 127, 2253–2262. [Google Scholar] [CrossRef]
- Nemere, I.; Norman, A.W. The rapid, hormonally stimulated transport of calcium (transcaltachia). J. Bone Miner. Res. 1987, 2, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Selles, J.; Boland, R. Evidence on the participation of the 3’,5’-cyclic AMP pathway in the non-genomic action of 1,25-dihydroxy-vitamin D3 in cardiac muscle. Mol. Cell. Endocrinol. 1991, 82, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Mendel, C.M. The Free Hormone Hypothesis: A Physiologically Based Mathematical Model. Endocr. Rev. 1989, 10, 232–274. [Google Scholar] [CrossRef]
- Provvedini, D.M.; Tsoukas, C.D.; Deftos, L.J.; Manolagas, S.C. 1,25-dihydroxyvitamin D3 receptors in human leukocytes. Science 1983, 221, 1181–1183. [Google Scholar] [CrossRef] [PubMed]
- Medrano, M.; Carrillo-Cruz, E.; Montero, I.; Perez-Simon, J.A. Vitamin D: Effect on Haematopoiesis and Immune System and Clinical Applications. Int. J. Mol. Sci. 2018, 19, 2663. [Google Scholar] [CrossRef] [Green Version]
- Zmijewski, M.A. Vitamin D and Human Health. Int. J. Mol. Sci. 2019, 20, 145. [Google Scholar] [CrossRef] [Green Version]
- Ramalingam, R.; Larmonier, C.B.; Thurston, R.D.; Midura-Kiela, M.T.; Zheng, S.G.; Ghishan, F.K.; Kiela, P.R. Dendritic Cell-Specific Disruption of TGF-β Receptor II Leads to Altered Regulatory T Cell Phenotype and Spontaneous Multiorgan Autoimmunity. J. Immunol. 2012, 189, 3878–3893. [Google Scholar] [CrossRef] [Green Version]
- Kundu, R.; Theodoraki, A.; Haas, C.T.; Zhang, Y.; Chain, B.; Kriston-Vizi, J.; Noursadeghi, M.; Khoo, B. Cell-type-specific modulation of innate immune signalling by vitamin D in human mononuclear phagocytes. Immunology 2016, 150, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Lan, Q.; Zhou, X.; Fan, H.; Chen, M.; Wang, J.; Ryffel, B.; Brand, D.; Ramalingam, R.; Kiela, P.R.; Horwitz, D.A.; et al. Polyclonal CD4+Foxp3+ Treg cells induce TGFβ-dependent tolerogenic dendritic cells that suppress the murine lupus-like syndrome. J. Mol. Cell Biol. 2012, 4, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanherwegen, A.-S.; Gysemans, C.; Mathieu, C. Vitamin D endocrinology on the cross-road between immunity and metabolism. Mol. Cell. Endocrinol. 2017, 453, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Weaver, C.T.; Harrington, L.E.; Mangan, P.R.; Gavrieli, M.; Murphy, K.M. Th17: An Effector CD4 T Cell Lineage with Regulatory T Cell Ties. Immunity 2006, 24, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Schnell, A.; Littman, D.R.; Kuchroo, V.K. TH17 cell heterogeneity and its role in tissue inflammation. Nat. Immunol. 2023, 24, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Zhou, R.; Luger, D.; Zhu, W.; Silver, P.B.; Grajewski, R.S.; Su, S.-B.; Chan, C.-C.; Adorini, L.; Caspi, R.R. Calcitriol Suppresses Antiretinal Autoimmunity through Inhibitory Effects on the Th17 Effector Response. J. Immunol. 2009, 182, 4624–4632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, S.A.; Rahimzadeh, M.; Brierley, C.; Gration, B.; Doree, C.; Kimber, C.E.; Cajide, A.P.; Lamikanra, A.A.; Roberts, D.J. The role of vitamin D in increasing circulating T regulatory cell numbers and modulating T regulatory cell phenotypes in patients with inflammatory disease or in healthy volunteers: A systematic review. PLoS ONE 2019, 14, e0222313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Sims, G.P.; Chen, X.X.; Gu, Y.Y.; Chen, S.; Lipsky, P.E. Modulatory Effects of 1,25-Dihydroxyvitamin D3 on Human B Cell Differentiation. J. Immunol. 2007, 179, 1634–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, M.; Okabe, T.; Ozawa, K.; Urabe, A.; Takaku, F. 1 alpha,25-Dihydroxyvitamin D3 (calcitriol) stimulates proliferation of human circulating monocytes in vitro. FEBS Lett. 1985, 185, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Skrobot, A.; Demkow, U.; Wachowska, M. Immunomodulatory Role of Vitamin D: A Review. Adv. Exp. Med. Biol. 2018, 1108, 13–23. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Krysiak, R.; Szkróbka, W.; Okopień, B. The Effect of Vitamin D on Thyroid Autoimmunity in Levothyroxine-Treated Women with Hashimoto’s Thyroiditis and Normal Vitamin D Status. Exp. Clin. Endocrinol. Diabetes 2017, 125, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Tamer, G.; Arik, S.; Tamer, I.; Coksert, D. Relative vitamin D insufficiency in Hashimoto’s thyroiditis. Thyroid 2011, 21, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Klecha, A.J.; Arcos, M.L.B.; Frick, L.; Genaro, A.M.; Cremaschi, G. Immune-Endocrine Interactions in Autoimmune Thyroid Diseases. Neuroimmunomodulation 2008, 15, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Kivity, S.; Agmon-Levin, N.; Zisappl, M.; Shapira, Y.; Nagy, E.V.; Dankó, K.; Szekanecz, Z.; Langevitz, P.; Shoenfeld, Y. Vitamin D and autoimmune thyroid diseases. Cell. Mol. Immunol. 2011, 8, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Bozkurt, N.C.; Karbek, B.; Ucan, B.; Sahin, M.; Cakal, E.; Ozbek, M.; Delibasi, T. The association between severity of vitamin D deficiency and Hashimoto’s thyroiditis. Endocr Pract. 2013, 19, 479–484. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Wu, D.; Li, C.; Fan, C.; Chao, N.; Liu, J.; Li, Y.; Wang, R.; Miao, W.; Guan, H.; et al. Lower Serum 25-Hydroxyvitamin D Level is Associated With 3 Types of Autoimmune Thyroid Diseases. Medicine 2015, 94, e1639. [Google Scholar] [CrossRef]
- Botelho, I.M.B.; Neto, A.M.; Silva, C.A.; Tambascia, M.A.; Alegre, S.M.; Zantut-Wittmann, D.E. Vitamin D in Hashimoto’s thyroiditis and its relationship with thyroid function and inflammatory status. Endocr. J. 2018, 65, 1029–1037. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Chai, Y.; Wei, H.; Wang, K.; Tan, L.; Zhang, W.; Fan, Y.; Li, F.; Shan, Z.; Zhu, M. Vitamin D deficiency is associated with thyroid autoimmunity: Results from an epidemiological survey in Tianjin, China. Endocrine 2021, 73, 447–454. [Google Scholar] [CrossRef]
- Chao, G.; Zhu, Y.; Fang, L. Correlation Between Hashimoto’s Thyroiditis-Related Thyroid Hormone Levels and 25-Hydroxyvitamin, D. Front. Endocrinol. 2020, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Effraimidis, G.; Badenhoop, K.; Tijssen, J.G.P.; Wiersinga, W.M. Vitamin D deficiency is not associated with early stages of thyroid autoimmunity. Eur. J. Endocrinol. 2012, 167, 43–48. [Google Scholar] [CrossRef] [Green Version]
- D’Aurizio, F.; Villalta, D.; Metus, P.; Doretto, P.; Tozzoli, R. Is vitamin D a player or not in the pathophysiology of autoimmune thyroid diseases? Autoimmun Rev. 2015, 14, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; Sun, T.; Zhang, Y.; He, L.; Wu, Q.; Liu, J.; Zha, B. 25-Hydroxyvitamin D serum level in Hashimoto’s thyroiditis, but not Graves’ disease is relatively deficient. Endocr. J. 2017, 64, 581–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duntas, L.H.; Alexandraki, K.I. On the Centennial of Vitamin D—Vitamin D, Inflammation, and Autoimmune Thyroiditis: A Web of Links and Implications. Nutrients 2022, 14, 5032. [Google Scholar] [CrossRef]
- Zhong, Z.; Ethen, N.J.; Williams, B.O. WNT signaling in bone development and homeostasis. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 489–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, K.; Irie, N. Osteoclast-osteoblast communication. Arch. Biochem. Biophys. 2008, 473, 201–209. [Google Scholar] [CrossRef] [PubMed]
- van Driel, M.; van Leeuwen, J.P. Vitamin D endocrine system and osteoblasts. Bonekey Rep. 2014, 3, 493. [Google Scholar] [CrossRef] [Green Version]
- van Driel, M.; Koedam, M.; Buurman, C.; Roelse, M.; Weyts, F.; Chiba, H.; Uitterlinden, A.; Pols, H.; van Leeuwen, J. Evidence that both 1α,25-dihydroxyvitamin D3 and 24-hydroxylated D3 enhance human osteoblast differentiation and mineralization. J. Cell. Biochem. 2006, 99, 922–935. [Google Scholar] [CrossRef]
- Xu, D.; Gao, H.-J.; Lu, C.-Y.; Tian, H.-M.; Yu, X.-J. Vitamin D inhibits bone loss in mice with thyrotoxicosis by activating the OPG/RANKL and Wnt/β-catenin signaling pathways. Front. Endocrinol. 2022, 13, 1066089. [Google Scholar] [CrossRef]
- Hannan, F.M.; Kallay, E.; Chang, W.; Brandi, M.L.; Thakker, R.V. The calcium-sensing receptor in physiology and in calcitropic and noncalcitropic diseases. Nat. Rev. Endocrinol. 2018, 15, 33–51. [Google Scholar] [CrossRef]
- Chu, H.; Qin, Z.; Ma, J.; Xie, Y.; Shi, H.; Gu, J.; Shi, B. Calcium-Sensing Receptor (CaSR)-Mediated Intracellular Communication in Cardiovascular Diseases. Cells 2022, 11, 3075. [Google Scholar] [CrossRef]
- Rendina-Ruedy, E.; Rosen, C.J. Parathyroid hormone (PTH) regulation of metabolic homeostasis: An old dog teaches us new tricks. Mol. Metab. 2022, 60, 101480. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Southgate, R.D.; Farhat, Y.M.; Loiselle, A.E.; Hammert, W.C.; Awad, H.A.; O’Keefe, R.J. Parathyroid hormone 1-34 enhances extracellular matrix deposition and organization during flexor tendon repair. J. Orthop. Res. 2014, 33, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kryśkiewicz, E.; Lorenc, R.S. Mechanizmy działania leków antykatabolicznych stosowanych w osteoporozie [Mechanisms of action of anticatabolic drugs used in osteoporosis therapy]. Endokrynol. Pol. 2009, 60, 134–144. [Google Scholar] [PubMed]
- Blaine, J.; Chonchol, M.; Levi, M. Renal control of calcium, phosphate, and magnesium homeostasis. Clin. J. Am. Soc. Nephrol. 2015, 10, 1257–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.; Lee, K.; Ju, J.H. Recent Updates of Diagnosis, Pathophysiology, and Treatment on Osteoarthritis of the Knee. Int. J. Mol. Sci. 2021, 22, 2619. [Google Scholar] [CrossRef]
- Zhang, Y.; Jordan, J.M. Epidemiology of Osteoarthritis. Clin. Geriatr. Med. 2010, 26, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Mobasheri, A.; Batt, M. An update on the pathophysiology of osteoarthritis. Ann. Phys. Rehabil. Med. 2016, 59, 333–339. [Google Scholar] [CrossRef]
- Hannan, M.T.; Felson, D.T.; Pincus, T. Analysis of the discordance between radiographic changes and knee pain in osteoarthritis of the knee. J. Rheumatol. 2000, 27, 1513–1517. [Google Scholar]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.P. Osteoarthritis. Nat. Rev. Dis. Primers. 2016, 2, 16072. [Google Scholar] [CrossRef] [Green Version]
- Liu-Bryan, R.; Terkeltaub, R. Emerging regulators of the inflammatory process in osteoarthritis. Nat. Rev. Rheumatol. 2015, 11, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Troeberg, L.; Nagase, H. Proteases involved in cartilage matrix degradation in osteoarthritis. Biochim. Biophys. Acta. 2012, 1824, 133–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orfanidou, T.; Malizos, K.N.; Varitimidis, S.; Tsezou, A. 1,25-Dihydroxyvitamin D3 and extracellular inorganic phosphate activate mitogen-activated protein kinase pathway through fibroblast growth factor 23 contributing to hypertrophy and mineralization in osteoarthritic chondrocytes. Exp. Biol. Med. 2012, 237, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.F.; Driban, J.B.; Lo, G.H.; Price, L.L.; Booth, S.; Eaton, C.B.; Lu, B.; Nevitt, M.; Jackson, B.; Garganta, C.; et al. Vitamin D Deficiency Is Associated with Progression of Knee Osteoarthritis. J. Nutr. 2014, 144, 2002–2008. [Google Scholar] [CrossRef] [Green Version]
- Heidari, B.; Heidari, P.; Hajian-Tilaki, K. Association between serum vitamin D deficiency and knee osteoarthritis. Int. Orthop. 2011, 35, 1627–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Jones, G.; Cicuttini, F.; Wluka, A.; Zhu, Z.; Han, W.; Antony, B.; Wang, X.; Winzenberg, T.; Blizzard, L.; et al. Effect of Vitamin D Supplementation on Tibial Cartilage Volume and Knee Pain Among Patients With Symptomatic Knee Osteoarthritis: A Randomized Clinical Trial. JAMA 2016, 315, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.-R.; Chen, Y.-S.; Deng, W. The effect of vitamin D supplementation on knee osteoarthritis: A meta-analysis of randomized controlled trials. Int. J. Surg. 2017, 46, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Divjak, A.; Jovanovic, I.; Matic, A.; Lucic, A.T.; Gajovic, N.; Jurisevic, M.; Skevin, A.J.; Veselinovic, M. The influence of vitamin D supplementation on the expression of mediators of inflammation in knee osteoarthritis. Immunol. Res. 2022. Epub ahead of print. [Google Scholar]
- Muresan, G.C.; Hedesiu, M.; Lucaciu, O.; Boca, S.; Petrescu, N. Effect of Vitamin D on BoneRegeneration: A Review. Medicina 2022, 58, 1337. [Google Scholar] [CrossRef]
- Werny, J.G.; Sagheb, K.; Diaz, L.; Kämmerer, P.W.; Al-Nawas, B.; Schiegnitz, E. Does vitamin D have an effect on osseointegration of dental implants? A systematic review. Int. J. Implant. Dent. 2022, 8, 16. [Google Scholar] [CrossRef]
- Makke, A. Vitamin D Supplementation for Prevention of Dental Implant Failure: A Systematic Review. Int. J. Dent. 2022, 2022, 2845902. [Google Scholar] [CrossRef]
- Salomó-Coll, O.; Maté-Sánchez de Val, J.E.; Ramírez-Fernandez, M.P.; Hernández-Alfaro, F.; Gargallo-Albiol, J.; Calvo-Guirado, J.L. Topical applications of vitamin D on implant surface for bone-to-implant contact enhance: A pilot study in dogs part II. Clin. Oral. Implant. Res. 2016, 27, 896–903. [Google Scholar] [CrossRef]
- Dvorak, G.; Fügl, A.; Watzek, G.; Tangl, S.; Pokorny, P.; Gruber, R. Impact of dietary vitamin D on osseointegration in the ovariec-tomized rat. Clin. Oral. Implant. Res. 2012, 23, 1308–1313. [Google Scholar] [CrossRef] [PubMed]
- Kwiatek, J.; Jaroń, A.; Trybek, G. Impact of the 25-Hydroxycholecalciferol Concentration and Vitamin D Deficiency Treatment on Changes in the Bone Level at the Implant Site during the Process of Osseointegration: A Prospective, Randomized, Controlled Clinical Trial. J. Clin. Med. 2021, 10, 526. [Google Scholar] [CrossRef] [PubMed]
- Grønborg, I.M.; Tetens, I.; Andersen, E.W.; Kristensen, M.; Larsen, R.E.K.; Tran, T.L.L.; Andersen, R. Effect of vitamin D fortified foods on bone markers and muscle strength in women of Pakistani and Danish origin living in Denmark: A randomised controlled trial. Nutr. J. 2019, 18, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohrbach, R.; Bair, E.; Fillingim, R.B.; Gonzalez, Y.; Gordon, S.M.; Lim, P.-F.; Ribeiro-Dasilva, M.; Diatchenko, L.; Dubner, R.; Greenspan, J.D.; et al. Clinical Orofacial Characteristics Associated With Risk of First-Onset TMD: The OPPERA Prospective Cohort Study. J. Pain 2013, 14 (Suppl. 12), T33–T50. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.D.; Zhang, J.N.; Gan, Y.H.; Zhou, Y.H. Current Understanding of Pathogenesis and Treatment of TMJ Osteoarthritis. J. Dent. Res. 2015, 94, 666–673. [Google Scholar] [CrossRef]
- Tanaka, E.; Detamore, M.; Mercuri, L. Degenerative Disorders of the Temporomandibular Joint: Etiology, Diagnosis, and Treatment. J. Dent. Res. 2008, 87, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Roth, R.H. Temporomandibular pain-dysfunction and occlusal relationships. Angle Orthod. 1973, 43, 136–153. [Google Scholar] [CrossRef]
- Cordray, F.E. Three-dimensional analysis of models articulated in the seated condylar position from a deprogrammed asymptomatic population: A prospective study. Part 1. Am. J. Orthod. Dentofac. Orthop. 2006, 129, 619–630. [Google Scholar] [CrossRef]
- Manfredini, D.; Lombardo, L.; Siciliani, G. Temporomandibular disorders and dental occlusion. A systematic review of association studies: End of an era? J. Oral. Rehabil. 2017, 44, 908–923. [Google Scholar] [CrossRef]
- Michelotti, A.; Iodice, G. The role of orthodontics in temporomandibular disorders. J. Oral Rehabil. 2010, 37, 411–429. [Google Scholar] [CrossRef]
- Derwich, M.; Mitus-Kenig, M.; Pawlowska, E. Is the Temporomandibular Joints’ Reciprocal Clicking Related to the Morphology and Position of the Mandible, as Well as to the Sagittal Position of Lower Incisors?-A Case-Control Study. Int. J. Environ. Res. Public Health 2021, 18, 4994. [Google Scholar] [CrossRef] [PubMed]
- Wiȩckiewicz, M.; Paradowska, A.; Kawala, B.; Wiȩckiewicz, W. SAPHO syndrome as a possible cause of masticatory system anomalies-A review of the literature. Adv. Clin. Exp. Med. 2011, 20, 521–525. [Google Scholar]
- Smardz, J.; Martynowicz, H.; Michalek-Zrabkowska, M.; Wojakowska, A.; Mazur, G.; Winocur, E.; Wieckiewicz, M. Sleep Bruxism and Occurrence of Temporomandibular Disorders-Related Pain: A Polysomnographic Study. Front. Neurol. 2019, 10, 168. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, E.; Ohrbach, R.; Truelove, E.; Look, J.; Anderson, G.; Goulet, J.P.; List, T.; Svensson, P.; Gonzalez, Y.; Lobbezoo, F.; et al. International RDC/TMD Consortium Network, International association for Dental Research; Orofacial Pain Special Interest Group, International Association for the Study of Pain. Diagnostic Criteria for Temporomandibular Disorders (DC/TMD) for Clinical and Research Applications: Recommendations of the International RDC/TMD Consortium Network* and Orofacial Pain Special Interest Group†. J. Oral Facial Pain Headache 2014, 28, 6–27. [Google Scholar] [PubMed]
- Shen, M.; Luo, Y.; Niu, Y.; Chen, L.; Yuan, X.; Goltzman, D.; Chen, N.; Miao, D. 1,25(OH)2D deficiency induces temporomandibular joint osteoarthritis via secretion of senescence-associated inflammatory cytokines. Bone 2013, 55, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.W.; Kang, J.-H. Bone mineral density, bone microstructure, and bone turnover markers in females with temporomandibular joint osteoarthritis. Clin. Oral Investig. 2021, 25, 6435–6448. [Google Scholar] [CrossRef] [PubMed]
- Jagur, O.; Kull, M.; Leibur, E.; Kallikorm, R.; Loorits, D.; Lember, M.; Voog-Oras, U. Relationship between radiographic changes in the temporomandibular joint and bone mineral density: A population based study. Stomatologija 2011, 13, 42–48. [Google Scholar]
- Gupta, A.K.; Gupta, R.; Gill, S. Effectiveness of Vitamin D along with Splint therapy in the Vit D deficient patients with Temporomandibular disorder-A Randomized, double-blind, placebo-controlled clinical trial. J. Indian Prosthodont. Soc. 2022, 22, 65–73. [Google Scholar] [CrossRef]
- Demir, C.Y.; Ersoz, M.E. Biochemical changes associated with temporomandibular disorders. J. Int. Med. Res. 2018, 47, 765–771. [Google Scholar] [CrossRef] [Green Version]
- Madani, A.; Shamsian, S.A.; Layegh, P.; Abrisham, S.M.; Ravaghi, A.; Tayarani Najjaran, N. Are certain factors involved in calcium metabolism associated with temporomandibular disorders? Cranio 2021, 39, 202–208. [Google Scholar] [CrossRef]
- Kui, A.; Buduru, S.; Labunet, A.; Balhuc, S.; Negucioiu, M. Vitamin D and Temporomandibular Disorders: What Do We Know So Far? Nutrients 2021, 13, 1286. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Reference | Study Design | Participants and Intervention | Results |
|---|---|---|---|
| Kivity et al. (2021) [65] | Case control study | 92 patients (71 women, 21 men, mean age—group AITD: 45 ± 16 years; group non-AITD: 52 ± 16 years): - AITD (50 patients) - non-AITD (42 patients) - control group: healthy participants (98 patients) - blood tests were taken in all of the participants | The prevalence of vitamin D3 deficiency was: - significantly higher in patients with AITD compared to healthy individuals (72% versus 30.6%; p < 0.001), - in patients with HT compared to patients with non-AITDs (79% versus 52%; p < 0.05) - correlated to the presence of antithyroid antibodies (p = 0.01) |
| Bozkurt et al. (2013) [66] | Case control study | 560 patients: - group 1: euthyroid patients with ongoing HT (180 patients: 123 women, 57 men) - group 2: euthyroid subjects with newly diagnosed HT (180 patients, sex-, age-, and BMI-matched) - control group: healthy volunteers (180 patients) - blood tests were taken in all of the participants | Group 1 presented the lowest concentrations of vitamin D3 (11.4 ± 5.2 ng/mL) having compared to newly diagnosed HT subjects (Group 2) (13.1 ± 5.9 ng/mL, p = 0.002), as well as to control subjects (15.4 ± 6.8 ng/mL, p < 0.001) Serum vitamin D3 concentrations correlated positively with thyroid volume (r = 0.145, p < 0.001) and negatively with anti-TPO (r = −0.361, p < 0.001), as well as with anti-Tg levels (r = −0.335, p < 0.001). |
| Ma et al. (2015) [67] | Case control study | 210 patients: - GD (70 patients: 48 women, 22 men, mean age: 40.04 ± 15.24 years) - HT (70 patients: 51 women, 19 men, mean age: 40.11 ± 14.60 years) - Control (70 patients: 49 women, 21 men, mean age: 41.99 ± 13.31 years) - blood tests were taken in all of the participants | AITD patients presented significantly lower levels of 25(OH)D3 comparing to the control group (p < 0.001). Every 5 nmol/L increase in serum 25(OH)D3 concentrations led to reduction in GD and HT morbidity by 1.55-, and 1.62-fold. There were no significant relationships between serum 25(OH)D3 concentration and any of the below listed: titer of anti-TPO, anti-Tg, and TSH serum level. |
| Botelho et al. (2018) [68] | Case control study | 159 patients: - HT (88 patients: 82 women, 6 men, mean age: 42.2 years, age range: 20–66 years) - euthyroid healthy subjects (71 patients: 61 women, 10 men, mean age: 47.4, age range: 19–77 years) - blood tests were taken in all of the participants | Vitamin D3 concentrations did not differ significantly between HT patients and the control group (p = 0.1024). In HT group there were found positive correlations between vitamin D3 concentration and the below listed: free T4 (p = 0.0224), TNF-α (p = 0.0004), IL-5 (p = 0.0144), IL-17 (p = 0.0011). |
| Fang et al. (2021) [69] | Case control study | 1812 patients: - anti-TPO positive (237 patients: 171 women, 66 men, mean age: 45.76 ± 15.06 years) - anti-TPO negative (1575 patients: 880 women, 695 men, mean age: 42.39 ± 14.97 years) - anti Tg positive (254 patients: 196 women, 58 men, mean age: 45.11 ± 15.09 years) - anti Tg negative (1558 patients: 855 women, 703 men, mean age: 42.46 ± 14.98 years) - blood tests were taken in all of the participants | Vitamin D3 deficiency was associated with increased likelihood of positive anti-TPO ([OR]: 2.428, 95%, [CI]: 1.383–4.261), as well as with positive anti-TG was (OR: 2.366, 95% CI: 1.366–4.099). HT patients, compared to healthy group, presented a significantly higher proportions of Th1 and Th17 cells, as well as significantly higher level of related cytokines. |
| Chao et al. (2020) [70] | Case control study | 5230 patients: - non-HT (4889 patients: 1851 women, 3038 men, mean age: 48.99 ± 9.04 years) - HT (373 patients: 248 women, 125 men, mean age: 48.51 ± 9.36 years) - blood tests were taken in all of the participants | The concentration of vitamin D3 was significantly lower in the HT group comparing to the non-HT group (p = 0.014). 25(OH)D3 deficiency and insufficiency groups presented significantly higher concentrations of TSH comparing to the 25(OH)D3 sufficiency group (p < 0.001). The concentrations of FT3 and FT4 were significantly lower in the 25(OH)D3 deficiency and insufficiency groups (p < 0.001) comparing to 25(OH)D3 sufficiency group. |
| Effraimidis et al. (2012) [71] | Two case control studies | Study A: 156 patients (156 women): - healthy euthyroid thyroid antibody-1 negative female relatives of AITD patients (78 women, mean age: 42.1 ± 13.2 years) - healthy controls (78 women, mean age: 42.3 ± 13.1 years) Study B: - healthy euthyroid thyroid antibodies-negative women who developed anti-TPO during 5-year follow-up (67 women, mean age: 38.3 ± 11.5 years) - healthy women who did not develop anti-TPO during 5-year follow-up (67 women, mean age: 38.1 ± 11.3 years) - blood tests were taken in all of the participants in both studies | Healthy euthyroid thyroid antibody-1 negative female relatives of AITD patients presented significantly higher serum 25(OH)D3 concentration compared to the control group (p = 0.01). The prevalence of 25(OH)D3 deficiency (<20 ng/mL) occurred less often in female relatives of AITD patients comparing to healthy controls (p = 0.05). The concentrations of the 1,25(OH)2D3 and 25(OH)D3 did not differ significantly between patients who developed anti-TPO and healthy controls. There was no association between low concentration of vitamin D3 and early stages of thyroid autoimmunity. |
| Ke et al. (2017) [73] | Case control study | 226 patients: - patients diagnosed with AITD (175 patients) * GD (51 patients, 30 women, 21 men, mean age 39.79 ± 1.73) * euthyroid HT, mild HT (61 patients, 34 women, 27 men, mean age 40.88 ± 1.61) * euthyroid HT patients with hypothyroidism receiving, treated HT (63 patients, 35 women, 28 men, mean age 42.41 ± 1.49) - healthy controls (51 patients, 31 women, 20 men, mean age 36.48 ± 1.68) - blood tests were taken in all of the participants | Patients from groups: treated HT and mild HT presented significantly lower vitamin D3 levels (p < 0.001) comparing to the controls. The concentration of vitamin D3 did not differ significantly between GD patients and healthy controls. Within the AITD group there were no correlations between vitamin D3 serum levels and thyroid hormones, antithyroid antibodies, as well as serum cytokines TNF-α, IL-4 and IL-17. |
| Reference | Study Design | Participants and Intervention | Results |
|---|---|---|---|
| Orfanidou et al. (2012) [93] | Case control study | 50 patients: - patients with end-stage primary OA undergoing knee replacement surgery (40 patients, 30 women, 10 men, mean age: 64.18 + 14.24 years) - control group, healthy cartilage samples obtained during fracture repair surgery (10 patients, 7 women, 3 men, mean age: 44.60 + 7.6) - blood tests were taken in all of the participants | The expression levels of FGF23, FGFR1c, VDR, PiT-1, and PiT-2 were significantly higher in patients diagnosed with OA comparing to the control group. |
| Zhang et al. (2014) [94] | Case control study | 418 patients: - patients with at least one knee with radiographic symptoms of OA and frequent pain/aching or stiffness in the area of knee (418 patients, 195 women, 223 men, mean age: 61.0 ± 9.2) - blood tests were taken in all of the participants | Low serum vitamin D3 concentration was associated with an increased risk of OA progression. |
| Heidari et al. (2011) [95] | Case control study | 298 patients: - knee OA group (148 patients, mean age: 60.2 ± 12.9 years) - control group (150 patients, mean age: 60.1 ± 10.2 years) - blood tests were taken in all of the participants | Mean serum vitamin D3 concentration did not differ significantly bwteen OA patients and the control group (p = 0.28). Mean serum vitamin D3 in OA patients aged below 60 years old was significantly lower comparing to the control group (p = 0.01). In patients younger than 60 years old the knee OA was significantly correlated with vitamin D3 deficiency (p = 0.018). |
| Jin et al. (2016) [96] | Randomized clinical trial | 413 patients with symptomatic knee OA for at least 6 months: - oral vitamin D3 group (209 patients, 106 women, 103 men, mean age: 63.5 + 6.9 years)—monthly intake of 50 000 IU of vitamin D3 for 24 months - placebo group (204 patients, 102 women, 102 men, mean age: 62.9 + 7.2 years) - all patients underwent MRI examination of the knee - blood tests were taken in all of the participants | The serum level of 25(OH)D3 increased significantly more in the vitamin D3 group comparing to the placebo (p < 0.001). Neither tibial cartilage volume nor WOMAC pain score changed significantly. Supplementation of vitamin D3 does not prevent tibial cartilage loss nor improves WOMAC knee pain. |
| Divjak et al. (2022) [98] | Randomized clinical trial | 80 patients diagnosed with knee OA: - oral vitamin D3 solution group (60 patients, 35 women, 25 men, mean age: 57.4 + 4.2 years)— 4 000 IU of vitamin D3 per day for 3 months - control group—without supplementation (20 patients, 12 women, 8 men, mean age: 56.1 + 4.3 years) - blood tests were taken in all of the participants | Vitamin D3 supplementation led to a significant pain reduction, both in VAS and WOMAC pain scores. The serum concentrations of IL-1β (p < 0.01), IL-23 (p < 0.01), and IL-33 (p < 0.01) significantly increased in patients who received vitamin D3 supplementation comparing to control group. The serum concentrations of IL-13 (p < 0.01), IL-17 (p < 0.01) and TNF-α (p < 0.01) significantly decreased in patients who received vitamin D3 supplementation comparing to control group. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szulc, M.; Świątkowska-Stodulska, R.; Pawłowska, E.; Derwich, M. Vitamin D3 Metabolism and Its Role in Temporomandibular Joint Osteoarthritis and Autoimmune Thyroid Diseases. Int. J. Mol. Sci. 2023, 24, 4080. https://doi.org/10.3390/ijms24044080
Szulc M, Świątkowska-Stodulska R, Pawłowska E, Derwich M. Vitamin D3 Metabolism and Its Role in Temporomandibular Joint Osteoarthritis and Autoimmune Thyroid Diseases. International Journal of Molecular Sciences. 2023; 24(4):4080. https://doi.org/10.3390/ijms24044080
Chicago/Turabian StyleSzulc, Michał, Renata Świątkowska-Stodulska, Elżbieta Pawłowska, and Marcin Derwich. 2023. "Vitamin D3 Metabolism and Its Role in Temporomandibular Joint Osteoarthritis and Autoimmune Thyroid Diseases" International Journal of Molecular Sciences 24, no. 4: 4080. https://doi.org/10.3390/ijms24044080
APA StyleSzulc, M., Świątkowska-Stodulska, R., Pawłowska, E., & Derwich, M. (2023). Vitamin D3 Metabolism and Its Role in Temporomandibular Joint Osteoarthritis and Autoimmune Thyroid Diseases. International Journal of Molecular Sciences, 24(4), 4080. https://doi.org/10.3390/ijms24044080