

The Role of Neutrophils in Spondyloarthritis: A Journey across the Spectrum of Disease Manifestations
 ,
, 
Abstract
:1. Introduction
Neutrophils Overview
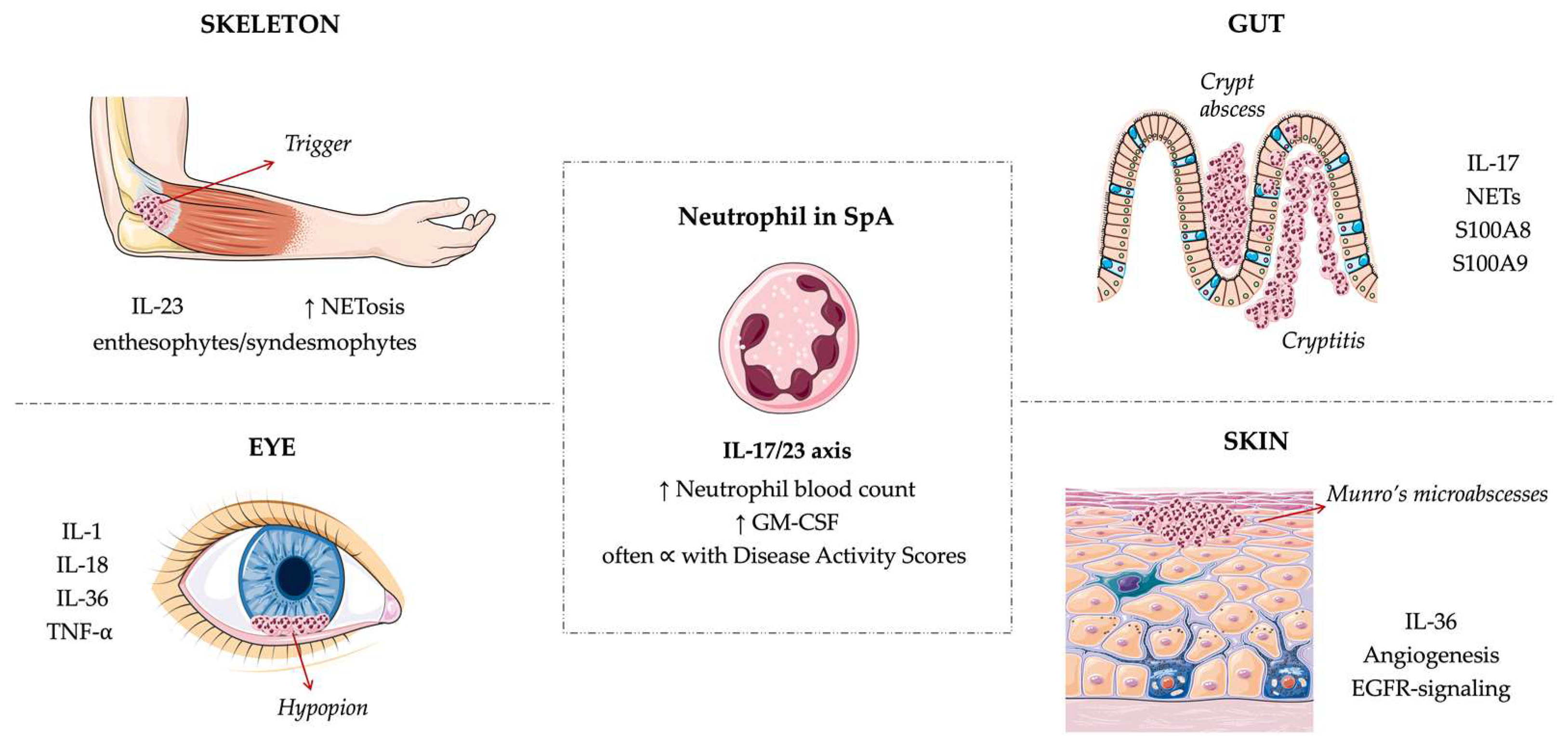
2. Neutrophils across SpA Manifestations
2.1. Articular Involvement
2.1.1. Enthesis
2.1.2. Peripheral and Axial Joints
2.2. Gut Involvement
2.3. Psoriasis
2.4. Uveitis
3. Therapeutics: Neutrophil-Targeted Therapies

{kind=link}
| Strategies | Approaches | Target | Drugs and Diseases |
|---|---|---|---|
| Reducing neutrophil numbers | Targeting production | - GM-CSF receptor | Mavrilimumab in GCA and RA [127,128,129] |
| - GM-CSF | Otilimab in RA [130]; Namilumab in RA [131], PsO [132] and SpA (NCT03622658); Gimsilumab in AS (NCT04205851; NCT04351243) | ||
| - IL-23/IL-17 axis (a regulator of G-CSF production) | IL-17 inhibitors in PsO, PsA, ax-SpA; IL-12/23 inhibitors in PsO, PsA [133], CD, and SLE; IL-23 inhibitors in CD, UC, PsO, and PsA | ||
| Inducing depletion | - Circulating neutrophils | -Extracorporeal granulocytapheresis in RA [134], CD, and RCU [135] | |
| Interfering with neutrophil recruitment and chemotaxis | Selectin and integrin blockers | - α4β1-integrin | -Natalizumab in CD [136] |
| - Selectins | -TBC1269 (and others) in PsO [137] | ||
| Blocking complement | - C5a and C5a receptor | -Eculizumab and Avacopan in AAV [138,139]; NNC0215-0384 in RA (NCT01611688) | |
| Blocking leucotriens | - LTB4 | -CP-195543 in RA (NCT00424294) | |
| Blocking neutrophils activation | Signal transduction blockade (cytokine signaling in neutrophils) | - JAK | -Jak-inhibitors in PsA, RA, UC, AS [140] |
| - SYK | -Fostamatinib in RA [141] and SLE [142] | ||
| - PDE4 | -Apremilast in PsO, PsA [143], SLE (NCT00708916) and AS [144] | ||
| Blocking cytokines whose receptors are also on neutrophils | - TNF-α | -TNF-α inhibitors in RA, PsA, PsO, SpA, AS, CD, UC, uveitis [145] | |
| - IL-6 | -IL-6 inhibitors in RA, AS, SSc, vasculitis, SLE, AOSD [146] | ||
| Blocking neutrophil-derived mediators | Neutrophil granule enzymes | - MMP9 | -andecaliximab in UC and CD [147,148] |
| NETs (blocking activity of enzymes critical for NETs formation) | - NADPH, MPO, PAD4, DNase Inhibitor | Not yet elucidated in humans affected by autoimmune diseases. | |
| Others | Blocking neutrophil function | - Neutrophil inflammasome | -β-hydroxybutyrate in gout flares [149] -IL-1β inhibitors in RA, SpA, PsA, AS, AOSD, uveitis, GCA, vasculitis [150] |
| - Neutrophils alarmins (S100A8/S100A9) | -Paquinimod in SLE (NCT00997100) |
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Dougados, M.; Baeten, D. Spondyloarthritis. Lancet 2011, 377, 2127–2137. [Google Scholar] [CrossRef]
- Benjamin, M.; McGonagle, D. The anatomical basis for disease localisation in seronegative spondyloarthropathy at entheses and related sites. J. Anat. 2001, 199, 503–526. [Google Scholar] [CrossRef]
- Bengtsson, K.; Forsblad-D’Elia, H.; Deminger, A.; Klingberg, E.; Dehlin, M.; Exarchou, S.; Lindström, U.; Askling, J.; Jacobsson, L.T.H. Incidence of extra-articular manifestations in ankylosing spondylitis, psoriatic arthritis and undifferentiated spondyloarthritis: Results from a national register-based cohort study. Rheumatology 2021, 60, 2725–2734. [Google Scholar] [CrossRef]
- Ronneberger, M.; Schett, G. Pathophysiology of Spondyloarthritis. Curr. Rheumatol. Rep. 2011, 13, 416–420. [Google Scholar] [CrossRef]
- Generali, E.; Bose, T.; Selmi, C.; Voncken, J.W.; Damoiseaux, J.G. Nature versus nurture in the spectrum of rheumatic diseases: Classification of spondyloarthritis as autoimmune or autoinflammatory. Autoimmun. Rev. 2018, 17, 935–941. [Google Scholar] [CrossRef]
- Annunziato, F.; Romagnani, C.; Romagnani, S. The 3 major types of innate and adaptive cell-mediated effector immunity. J. Allergy Clin. Immunol. 2015, 135, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Mauro, D.; Simone, D.; Bucci, L.; Ciccia, F. Novel immune cell phenotypes in spondyloarthritis pathogenesis. Semin. Immunopathol. 2021, 43, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Macleod, T.; Bridgewood, C.; McGonagle, D. Role of neutrophil interleukin-23 in spondyloarthropathy spectrum disorders. Lancet Rheumatol. 2023, 5, e47–e57. [Google Scholar] [CrossRef]
- Ng, L.G.; Ostuni, R.; Hidalgo, A. Heterogeneity of neutrophils. Nat. Rev. Immunol. 2019, 19, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Eash, K.J.; Greenbaum, A.; Gopalan, P.K.; Link, D.C. CXCR2 and CXCR4 antagonistically regulate neutrophil trafficking from murine bone marrow. J. Clin. Investig. 2010, 120, 2423–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summers, C.; Rankin, S.M.; Condliffe, A.M.; Singh, N.; Peters, A.M.; Chilvers, E.R. Neutrophil kinetics in health and disease. Trends Immunol. 2010, 31, 318–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. Available online: https://www.frontiersin.org/articles/10.3389/fphys.2018.00113 (accessed on 31 January 2023). [CrossRef] [PubMed]
- Pelletier, M.; Maggi, L.; Micheletti, A.; Lazzeri, E.; Tamassia, N.; Costantini, C.; Cosmi, L.; Lunardi, C.; Annunziato, F.; Romagnani, S.; et al. Evidence for a cross-talk between human neutrophils and Th17 cells. Blood 2010, 115, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Burn, G.L.; Foti, A.; Marsman, G.; Patel, D.F.; Zychlinsky, A. The Neutrophil. Immunity 2021, 54, 1377–1391. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Bevilacqua, D.; Cassatella, M.A.; Scapini, P. Recent advances on the crosstalk between neutrophils and B or T lymphocytes. Immunology 2019, 156, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puga, I.; Cols, M.; Barra, C.M.; He, B.; Cassis, L.; Gentile, M.; Comerma, L.; Chorny, A.; Shan, M.; Xu, W.; et al. B cell–helper neutrophils stimulate the diversification and production of immunoglobulin in the marginal zone of the spleen. Nat. Immunol. 2011, 13, 170–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoyratty, T.E.; Ai, Z.; Ballesteros, I.; Eames, H.L.; Mathie, S.; Martín-Salamanca, S.; Wang, L.; Hemmings, A.; Willemsen, N.; von Werz, V.; et al. Distinct transcription factor networks control neutrophil-driven inflammation. Nat. Immunol. 2021, 22, 1093–1106. [Google Scholar] [CrossRef]
- Montaldo, E.; Lusito, E.; Bianchessi, V.; Caronni, N.; Scala, S.; Basso-Ricci, L.; Cantaffa, C.; Masserdotti, A.; Barilaro, M.; Barresi, S.; et al. Cellular and transcriptional dynamics of human neutrophils at steady state and upon stress. Nat. Immunol. 2022, 23, 1470–1483. [Google Scholar] [CrossRef]
- Enthesitis: From Pathophysiology to Treatment—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/29158573/ (accessed on 2 November 2022).
- Ye, P.; Rodriguez, F.H.; Kanaly, S.; Stocking, K.L.; Schurr, J.; Schwarzenberger, P.; Oliver, P.; Huang, W.; Zhang, P.; Zhang, J.; et al. Requirement of Interleukin 17 Receptor Signaling for Lung Cxc Chemokine and Granulocyte Colony-Stimulating Factor Expression, Neutrophil Recruitment, and Host Defense. J. Exp. Med. 2001, 194, 519–528. [Google Scholar] [CrossRef]
- Yu, J.J.; Ruddy, M.J.; Wong, G.C.; Sfintescu, C.; Baker, P.J.; Smith, J.B.; Evans, R.T.; Gaffen, S.L. An essential role for IL-17 in preventing pathogen-initiated bone destruction: Recruitment of neutrophils to inflamed bone requires IL-17 receptor–dependent signals. Blood 2007, 109, 3794–3802. [Google Scholar] [CrossRef]
- Tamassia, N.; Arruda-Silva, F.; Wright, H.L.; Moots, R.J.; Gardiman, E.; Bianchetto-Aguilera, F.; Gasperini, S.; Capone, M.; Maggi, L.; Annunziato, F.; et al. Human neutrophils activated via TLR8 promote Th17 polarization through IL-23. J. Leukoc. Biol. 2019, 105, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Takahashi, T.; Hata, H.; Nomura, T.; Sakaguchi, N. SKG mice, a new genetic model of rheumatoid arthritis. Arthritis Res. Ther. 2003, 5, 10. [Google Scholar] [CrossRef]
- Stavre, Z.; Bridgewood, C.; Zhou, Q.; Maeda, Y.; Huang, T.-T.; Karman, J.; Khan, A.; Giryes, S.; Sharif, K.; McGonagle, D.; et al. A role for neutrophils in early enthesitis in spondyloarthritis. Arthritis Res. Ther. 2022, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Crowe, L.A.N.; McLean, M.; Kitson, S.M.; Melchor, E.G.; Patommel, K.; Cao, H.M.; Reilly, J.H.; Leach, W.J.; Rooney, B.P.; Spencer, S.J.; et al. S100A8 & S100A9: Alarmin mediated inflammation in tendinopathy. Sci. Rep. 2019, 9, 1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schonthaler, H.B.; Guinea-Viniegra, J.; Wculek, S.K.; Ruppen, I.; Ximénez-Embún, P.; Guío-Carrión, A.; Navarro, R.; Hogg, N.; Ashman, K.; Wagner, E.F. S100A8-S100A9 Protein Complex Mediates Psoriasis by Regulating the Expression of Complement Factor C3. Immunity 2013, 39, 1171–1181. [Google Scholar] [CrossRef] [Green Version]
- Turina, M.C.; Sieper, J.; Yeremenko, N.; Conrad, K.; Haibel, H.; Rudwaleit, M.; Baeten, D.; Poddubnyy, D. Calprotectin serum level is an independent marker for radiographic spinal progression in axial spondyloarthritis. Ann. Rheum. Dis. 2014, 73, 1746–1748. [Google Scholar] [CrossRef]
- Pruenster, M.; Kurz, A.R.M.; Chung, K.-J.; Cao-Ehlker, X.; Bieber, S.; Nussbaum, C.F.; Bierschenk, S.; Eggersmann, T.K.; Rohwedder, I.; Heinig, K.; et al. Extracellular MRP8/14 is a regulator of β2 integrin-dependent neutrophil slow rolling and adhesion. Nat. Commun. 2015, 6, 6915. [Google Scholar] [CrossRef] [Green Version]
- De Wilde, K.; Martens, A.; Lambrecht, S.; Jacques, P.; Drennan, M.B.; Debusschere, K.; Govindarajan, S.; Coudenys, J.; Verheugen, E.; Windels, F.; et al. A20 inhibition of STAT1 expression in myeloid cells: A novel endogenous regulatory mechanism preventing development of enthesitis. Ann. Rheum. Dis. 2017, 76, 585–592. [Google Scholar] [CrossRef]
- McGonagle, D.; Marzo-Ortega, H.; O’Connor, P.; Gibbon, W.; Hawkey, P.; Henshaw, K.; Emery, P. Histological assessment of the early enthesitis lesion in spondyloarthropathy. Ann. Rheum. Dis. 2002, 61, 534–537. [Google Scholar] [CrossRef] [Green Version]
- Pachowsky, M.L.; Raimondo, M.G.; Xu, C.; Rauber, S.; Tascilar, K.; Labinsky, H.; Vogg, M.; Saad, M.S.A.; Simon, D.; Rech, J.; et al. Concise report: A minimal-invasive method to retrieve and identify entheseal tissue from psoriatic arthritis patients. Ann. Rheum. Dis. 2022, 81, 1131–1135. [Google Scholar] [CrossRef]
- Van Tubergen, A.; Weber, U. Diagnosis and classification in spondyloarthritis: Identifying a chameleon. Nat. Rev. Rheumatol. 2012, 8, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Zheng, N.; Chen, S.-B.; Xiao, Z.-Y.; Wu, M.-Y.; Liu, Y.; Zeng, Q.-Y. Ten years’ experience with needle biopsy in the early diagnosis of sacroiliitis. Arthritis Rheum. 2012, 64, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Appel, H.; Maier, R.; Wu, P.; Scheer, R.; Hempfing, A.; Kayser, R.; Thiel, A.; Radbruch, A.; Loddenkemper, C.; Sieper, J. Analysis of IL-17+ cells in facet joints of patients with spondyloarthritis suggests that the innate immune pathway might be of greater relevance than the Th17-mediated adaptive immune response. Arthritis Res. Ther. 2011, 13, R95. [Google Scholar] [CrossRef] [Green Version]
- Papagoras, C.; Tsiami, S.; Chrysanthopoulou, A.; Mitroulis, I.; Baraliakos, X. Serum granulocyte-macrophage colony-stimulating factor (GM-CSF) is increased in patients with active radiographic axial spondyloarthritis and persists despite anti-TNF treatment. Arthritis Res. Ther. 2022, 24, 195. [Google Scholar] [CrossRef] [PubMed]
- Regan-Komito, D.; Swann, J.W.; Demetriou, P.; Cohen, E.S.; Horwood, N.J.; Sansom, S.N.; Griseri, T. GM-CSF drives dysregulated hematopoietic stem cell activity and pathogenic extramedullary myelopoiesis in experimental spondyloarthritis. Nat. Commun. 2020, 11, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Osami, M.H.; Awadh, N.I.; Khalid, K.B.; Awadh, A.I. Neutrophil/lymphocyte and platelet/lymphocyte ratios as potential markers of disease activity in patients with Ankylosing spondylitis: A case-control study. Hortic. Bras. Rheumatol. 2020, 60, 13. [Google Scholar] [CrossRef] [PubMed]
- Karow, F.; Smiljanovic, B.; Grün, J.R.; Poddubnyy, D.; Proft, F.; Talpin, A.; Hue, C.; Boland, A.; Deleuze, J.-F.; Garchon, H.-J.; et al. Monocyte transcriptomes from patients with axial spondyloarthritis reveal dysregulated monocytopoiesis and a distinct inflammatory imprint. Arthritis Res. Ther. 2021, 23, 246. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Limon, P.; Ladehesa-Pineda, M.L.; Castro-Villegas, M.D.C.; Abalos-Aguilera, M.D.C.; Lopez-Medina, C.; Lopez-Pedrera, C.; Barbarroja, N.; Espejo-Peralbo, D.; González-Reyes, J.A.; Villalba, J.M.; et al. Enhanced NETosis generation in radiographic axial spondyloarthritis: Utility as biomarker for disease activity and anti-TNF-α therapy effectiveness. J. Biomed. Sci. 2020, 27, 54. [Google Scholar] [CrossRef] [Green Version]
- Papagoras, C.; Chrysanthopoulou, A.; Mitsios, A.; Ntinopoulou, M.; Tsironidou, V.; Batsali, A.K.; Papadaki, H.A.; Skendros, P.; Ritis, K. IL-17A expressed on neutrophil extracellular traps promotes mesenchymal stem cell differentiation toward bone-forming cells in ankylosing spondylitis. Eur. J. Immunol. 2021, 51, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Mitsios, A.; Arampatzioglou, A.; Arelaki, S.; Mitroulis, I.; Ritis, K. NETopathies? Unraveling the Dark Side of Old Diseases through Neutrophils. Front. Immunol. 2016, 7, 678. [Google Scholar] [CrossRef]
- Jorch, S.K.; Kubes, P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nat. Med. 2017, 23, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Kendall, M.J.; Farr, M.; Meynell, M.J.; Hawkins, C.F. Synovial fluid in ankylosing spondylitis. Ann. Rheum. Dis. 1973, 32, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruithof, E.; Baeten, D.; De Rycke, L.; Vandooren, B.; Foell, D.; Roth, J.; Cañete, J.D.; Boots, A.M.; Veys, E.M.; De Keyser, F. Synovial histopathology of psoriatic arthritis, both oligo- and polyarticular, resembles spondyloarthropathy more than it does rheumatoid arthritis. Arthritis Res. Ther. 2005, 7, R569–R580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baeten, D.; Kruithof, E.; De Rycke, L.; Boots, A.M.; Mielants, H.; Veys, E.M.; De Keyser, F. Infiltration of the synovial membrane with macrophage subsets and polymorphonuclear cells reflects global disease activity in spondyloarthropathy. Arthritis Res. Ther. 2005, 7, R359–R369. [Google Scholar] [CrossRef] [Green Version]
- Kruithof, E.; De Rycke, L.; Vandooren, B.; De Keyser, F.; FitzGerald, O.; McInnes, I.; Tak, P.P.; Bresnihan, B.; Veys, E.M.; Baeten, D.; et al. Identification of synovial biomarkers of response to experimental treatment in early-phase clinical trials in spondylarthritis. Arthritis Rheum. 2006, 54, 1795–1804. [Google Scholar] [CrossRef]
- Li, B.; Li, G.; Yang, X.; Song, Z.; Wang, Y.; Zhang, Z. NETosis in Psoriatic Arthritis: Serum MPO–DNA Complex Level Correlates With Its Disease Activity. Front. Immunol. 2022, 13, 911347. [Google Scholar] [CrossRef]
- Lambert, S.; Hambro, C.A.; Johnston, A.; Stuart, P.E.; Tsoi, L.C.; Nair, R.P.; Elder, J.T. Neutrophil Extracellular Traps Induce Human Th17 Cells: Effect of Psoriasis-Associated TRAF3IP2 Genotype. J. Investig. Dermatol. 2019, 139, 1245–1253. [Google Scholar] [CrossRef] [Green Version]
- De Benedittis, G.; Latini, A.; Conigliaro, P.; Triggianese, P.; Bergamini, A.; Novelli, L.; Ciccacci, C.; Chimenti, M.S.; Borgiani, P. A multilocus genetic study evidences the association of autoimmune-related genes with Psoriatic Arthritis in Italian patients. Immunobiology 2022, 227, 152232. [Google Scholar] [CrossRef]
- Bierkarre, H.; Harder, J.; Cuthbert, R.; Emery, P.; Leuschner, I.; Mrowietz, U.; Hedderich, J.; McGonagle, D.; Gläser, R. Differential expression of antimicrobial peptides in psoriasis and psoriatic arthritis as a novel contributory mechanism for skin and joint disease heterogeneity. Scand. J. Rheumatol. 2016, 45, 188–196. [Google Scholar] [CrossRef]
- Frasca, L.; Palazzo, R.; Chimenti, M.S.; Alivernini, S.; Tolusso, B.; Bui, L.; Botti, E.; Giunta, A.; Bianchi, L.; Petricca, L.; et al. Anti-LL37 Antibodies Are Present in Psoriatic Arthritis (PsA) Patients: New Biomarkers in PsA. Front. Immunol. 2018, 9, 1936. [Google Scholar] [CrossRef] [Green Version]
- Gracey, E.; Vereecke, L.; McGovern, D.; Fröhling, M.; Schett, G.; Danese, S.; De Vos, M.; Bosch, F.V.D.; Elewaut, D. Revisiting the gut–joint axis: Links between gut inflammation and spondyloarthritis. Nat. Rev. Rheumatol. 2020, 16, 415–433. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.M.; Parkos, C.A. The role of neutrophils during intestinal inflammation. Mucosal Immunol. 2012, 5, 354–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielants, H.; Veys, E.M.; Cuvelier, C.; De Vos, M.; Goemaere, S.; De Clercq, L.; Schatteman, L.; Gyselbrecht, L.; Elewaut, D. The evolution of spondyloarthropathies in relation to gut histology. III. Relation between gut and joint. J. Rheumatol. 1995, 22, 2279–2784. [Google Scholar] [PubMed]
- Mielants, H.; Veys, E.M.; Cuvelier, C.; De Vos, M.; Goemaere, S.; De Clercq, L.; Schatteman, L.; Elewaut, D. The evolution of spondyloarthropathies in relation to gut histology. II. Histological aspects. J. Rheumatol. 1995, 22, 2273–2278. [Google Scholar] [PubMed]
- Lin, E.; Lai, H.-J.; Cheng, Y.-K.; Leong, K.-Q.; Cheng, L.-C.; Chou, Y.-C.; Peng, Y.-C.; Hsu, Y.-H.; Chiang, H.-S. Neutrophil Extracellular Traps Impair Intestinal Barrier Function during Experimental Colitis. Biomedicines 2020, 8, 275. [Google Scholar] [CrossRef]
- Spadoni, I.; Zagato, E.; Bertocchi, A.; Paolinelli, R.; Hot, E.; Di Sabatino, A.; Caprioli, F.; Bottiglieri, L.; Oldani, A.; Viale, G.; et al. A gut-vascular barrier controls the systemic dissemination of bacteria. Science 2015, 350, 830–834. [Google Scholar] [CrossRef] [PubMed]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Alessandro, R.; Luchetti, M.M.; Milling, S.; Saieva, L.; Cypers, H.; Stampone, T.; Di Benedetto, P.; et al. Dysbiosis and zonulin upregulation alter gut epithelial and vascular barriers in patients with ankylosing spondylitis. Ann. Rheum. Dis. 2017, 76, 1123–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, A.; Ferrante, A.; Guggino, G.; Ciccia, F. Gut inflammation in spondyloarthritis. Best Pract. Res. Clin. Rheumatol. 2017, 31, 863–876. [Google Scholar] [CrossRef]
- Tester, A.M.; Cox, J.H.; Connor, A.R.; Starr, A.E.; Dean, R.A.; Suarez-Puente, X.; López-Otín, C.; Overall, C.M. LPS Responsiveness and Neutrophil Chemotaxis In Vivo Require PMN MMP-8 Activity. PLoS ONE 2007, 2, e312. [Google Scholar] [CrossRef] [Green Version]
- Van den Steen, P.E.; Proost, P.; Wuyts, A.; Van Damme, J.O.; Opdenakker, G. Neutrophil gelatinase B potentiates interleukin-8 tenfold by aminoterminal processing, whereas it degrades CTAP-III, PF-4, and GRO-alpha and leaves RANTES and MCP-2 intact. Blood 2000, 96, 2673–2681. [Google Scholar] [CrossRef]
- Iking-Konert, C.; Ostendorf, B.; Sander, O.; Jost, M.; Wagner, C.; Joosten, L.; Schneider, M.; Hänsch, G.M. Transdifferentiation of polymorphonuclear neutrophils to dendritic-like cells at the site of inflammation in rheumatoid arthritis: Evidence for activation by T cells. Ann. Rheum. Dis. 2005, 64, 1436–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cypers, H.; Varkas, G.; Beeckman, S.; Debusschere, K.; Vogl, T.; Roth, J.; Drennan, M.; Lavric, M.; Foell, D.; Cuvelier, C.A.; et al. Elevated calprotectin levels reveal bowel inflammation in spondyloarthritis. Ann. Rheum. Dis. 2016, 75, 1357–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, A.; Guggino, G.; Ferrante, A.; Ciccia, F. Role of Subclinical Gut Inflammation in the Pathogenesis of Spondyloarthritis. Front. Med. 2018, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Schatteman, L.; Mielants, H.; Veys, E.M.; Cuvelier, C.; De Vos, M.; Gyselbrecht, L.; Elewaut, D.; Goemaere, S. Gut inflammation in psoriatic arthritis: A prospective ileocolonoscopic study. J. Rheumatol. 1995, 22, 680–683. [Google Scholar]
- Macaluso, F.; Guggino, G.; Rizzo, A.; Ferrante, A.; Ciccia, F. Histopathology of the gut in rheumatic diseases. Reumatismo 2018, 70, 178–186. [Google Scholar] [CrossRef] [Green Version]
- Mielants, H.; Veys, E.M.; De Vos, M.; Cuvelier, C.; Goemaere, S.; De Clercq, L.; Schatteman, L.; Elewaut, D. The evolution of spondyloarthropathies in relation to gut histology. I. Clinical aspects. J. Rheumatol. 1995, 22, 2266–2272. [Google Scholar]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef]
- Serhan, C.N.; Petasis, N.A. Resolvins and Protectins in Inflammation Resolution. Chem. Rev. 2011, 111, 5922–5943. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.D.; Voyich, J.M.; Whitney, A.R.; DeLeo, F.R. Spontaneous neutrophil apoptosis and regulation of cell survival by granulocyte macrophage-colony stimulating factor. J. Leukoc. Biol. 2005, 78, 1408–1418. [Google Scholar] [CrossRef] [Green Version]
- Arosa, L.; Camba-Gómez, M.; Conde-Aranda, J. Neutrophils in Intestinal Inflammation: What We Know and What We Could Expect for the Near Future. Gastrointest. Disord. 2022, 4, 263–276. [Google Scholar] [CrossRef]
- Brannigan, A.E.; O’connell, R.P.; Hurley, H.; O’neill, A.; Brady, H.R.; Fitzpatrick, J.M.; William, R.; Watson, G. Neutrophil apoptosis is delayed in patients with inflammatory bowel disease. Shock 2000, 13, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Ina, K.; Kusugami, K.; Hosokawa, T.; Imada, A.; Shimizu, T.; Yamaguchi, T.; Ohsuga, M.; Kyokane, K.; Sakai, T.; Nishio, Y.; et al. Increased mucosal production of granulocyte colony-stimulating factor is related to a delay in neutrophil apoptosis in Inflammatory Bowel disease. J. Gastroenterol. Hepatol. 1999, 14, 46–53. [Google Scholar] [CrossRef]
- Bassler, K.; Schulte-Schrepping, J.; Warnat-Herresthal, S.; Aschenbrenner, A.C.; Schultze, J.L. The Myeloid Cell Compartment—Cell by Cell. Annu. Rev. Immunol. 2019, 37, 269–293. [Google Scholar] [CrossRef]
- Qaiyum, Z.; Lim, M.; Inman, R.D. The gut-joint axis in spondyloarthritis: Immunological, microbial, and clinical insights. Semin. Immunopathol. 2021, 43, 173–192. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- FitzGerald, O.; Ogdie, A.; Chandran, V.; Coates, L.C.; Kavanaugh, A.; Tillett, W.; Leung, Y.Y.; Dewit, M.; Scher, J.U.; Mease, P.J. Psoriatic arthritis. Nat. Rev. Dis. Prim. 2021, 7, 1–17. [Google Scholar] [CrossRef]
- Czerwińska, J.; Owczarczyk-Saczonek, A. The Role of the Neutrophilic Network in the Pathogenesis of Psoriasis. Int. J. Mol. Sci. 2022, 23, 1840. [Google Scholar] [CrossRef]
- Naik, H.B.; Natarajan, B.; Stansky, E.; Ahlman, M.A.; Teague, H.; Salahuddin, T.; Ng, Q.; Joshi, A.A.; Krishnamoorthy, P.; Dave, J.; et al. Severity of Psoriasis Associates With Aortic Vascular Inflammation Detected by FDG PET/CT and Neutrophil Activation in a Prospective Observational Study. Arter. Thromb. Vasc. Biol. 2015, 35, 2667–2676. [Google Scholar] [CrossRef] [Green Version]
- Polat, M.; Bugdayci, G.; Kaya, H.; Oğuzman, H. Evaluation of neutrophil-to-lymphocyte ratio and platelet-to-lymphocyte ratio in Turkish patients with chronic plaque psoriasis. Acta Dermatovenerol. Alp. Pannonica et Adriat. 2017, 26, 97–100. [Google Scholar] [CrossRef]
- Teague, H.L.; Varghese, N.J.; Tsoi, L.C.; Dey, A.K.; Garshick, M.S.; Silverman, J.I.; Baumer, Y.; Harrington, C.L.; Stempinski, E.; Elnabawi, Y.A.; et al. Neutrophil Subsets, Platelets, and Vascular Disease in Psoriasis. JACC Basic Transl. Sci. 2019, 4, 1–14. [Google Scholar] [CrossRef]
- Toichi, E.; Tachibana, T.; Furukawa, F. Rapid improvement of psoriasis vulgaris during drug-induced agranulocytosis. J. Am. Acad. Dermatol. 2000, 43, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, S.; Takahashi, H.; Suga, Y.; Eto, H.; Etoh, T.; Okuma, K.; Takahashi, K.; Kanbara, T.; Seishima, M.; Morita, A.; et al. Therapeutic depletion of myeloid lineage leukocytes in patients with generalized pustular psoriasis indicates a major role for neutrophils in the immunopathogenesis of psoriasis. J. Am. Acad. Dermatol. 2013, 68, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Umezawa, Y.; Yamagiwa, A.; Saeki, H.; Kondo, M.; Gabazza, E.; Nakagawa, H.; Mizutani, H. Biologic therapy improves psoriasis by decreasing the activity of monocytes and neutrophils. J. Dermatol. 2014, 41, 679–685. [Google Scholar] [CrossRef]
- Hwang, T.-L. Neutrophils in Psoriasis. Front. Immunol. 2019, 10, 12. [Google Scholar]
- Bloomfield, F.J.; Young, M.M. Enhanced chemiluminescence production by phagocytosing neutrophils in psoriasis. Inflammation 1988, 12, 153–159. [Google Scholar] [CrossRef]
- Dilek, N.; Dilek, A.R.; Taşkın, Y.; Erkinüresin, T.; Yalçın, Ö.; Saral, Y. Contribution of myeloperoxidase and inducible nitric oxide synthase to pathogenesis of psoriasis. Postep. Dermatol. Allergol. 2016, 33, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Henry, C.M.; Sullivan, G.P.; Clancy, D.M.; Afonina, I.S.; Kulms, D.; Martin, S.J. Neutrophil-Derived Proteases Escalate Inflammation through Activation of IL-36 Family Cytokines. Cell Rep. 2016, 14, 708–722. [Google Scholar] [CrossRef] [Green Version]
- Xhindoli, D.; Pacor, S.; Benincasa, M.; Scocchi, M.; Gennaro, R.; Tossi, A. The human cathelicidin LL-37—A pore-forming antibacterial peptide and host-cell modulator. Biochim. Biophys. Acta 2016, 1858, 546–566. [Google Scholar] [CrossRef]
- Meyer-Hoffert, U.; Wingertszahn, J.; Wiedow, O. Human Leukocyte Elastase Induces Keratinocyte Proliferation by Epidermal Growth Factor Receptor Activation. J. Investig. Dermatol. 2004, 123, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Loss-of-Function Myeloperoxidase Mutations Are Associated with Increased Neutrophil Counts and Pustular Skin Disease—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/32758448/ (accessed on 2 November 2022).
- Hau, C.S.; Kanda, N.; Tada, Y.; Shibata, S.; Uozaki, H.; Fukusato, T.; Sato, S.; Watanabe, S. Lipocalin-2 exacerbates psoriasiform skin inflammation by augmenting T-helper 17 response. J. Dermatol. 2016, 43, 785–794. [Google Scholar] [CrossRef]
- Schön, M.P.; Broekaert, S.M.C.; Erpenbeck, L. Sexy again: The renaissance of neutrophils in psoriasis. Exp. Dermatol. 2017, 26, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Shao, S.; Fang, H.; Dang, E.; Xue, K.; Zhang, J.; Li, B.; Qiao, H.; Cao, T.; Zhuang, Y.; Shen, S.; et al. Neutrophil Extracellular Traps Promote Inflammatory Responses in Psoriasis via Activating Epidermal TLR4/IL-36R Crosstalk. Front. Immunol. 2019, 10, 746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Kronbichler, A.; Park, D.D.-Y.; Park, Y.; Moon, H.; Kim, H.; Choi, J.H.; Choi, Y.; Shim, S.; Lyu, I.S.; et al. Neutrophil extracellular traps (NETs) in autoimmune diseases: A comprehensive review. Autoimmun. Rev. 2017, 16, 1160–1173. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.C.-S.; Yu, H.-S.; Yen, F.-L.; Lin, C.-L.; Chen, G.-S.; Lan, C.-C.E. Neutrophil extracellular trap formation is increased in psoriasis and induces human β-defensin-2 production in epidermal keratinocytes. Sci. Rep. 2016, 6, 31119. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.-M.; Jin, H.-Z. Role of Neutrophils in Psoriasis. J. Immunol. Res. 2020, 2020, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, D.; Chamilos, G.; Lande, R.; Gregorio, J.; Meller, S.; Facchinetti, V.; Homey, B.; Barrat, F.J.; Zal, T.; Gilliet, M. Self-RNA–antimicrobial peptide complexes activate human dendritic cells through TLR7 and TLR8. J. Exp. Med. 2009, 206, 1983–1994. [Google Scholar] [CrossRef]
- Lande, R.; Gregorio, J.; Facchinetti, V.; Chatterjee, B.; Wang, Y.-H.; Homey, B.; Cao, W.; Wang, Y.-H.; Su, B.; Nestle, F.O.; et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature 2007, 449, 564–569. [Google Scholar] [CrossRef]
- Lin, A.M.; Rubin, C.J.; Khandpur, R.; Wang, J.Y.; Riblett, M.; Yalavarthi, S.; Villanueva, E.C.; Shah, P.; Kaplan, M.J.; Bruce, A.T. Mast Cells and Neutrophils Release IL-17 through Extracellular Trap Formation in Psoriasis. J. Immunol. 2011, 187, 490–500. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Yao, X.; Zhai, Y.; Li, L.; Li, H.; Sun, X.; Yu, P.; Xue, T.; Li, Y.; Hu, Y. Single cell transcriptional zonation of human psoriasis skin identifies an alternative immunoregulatory axis conducted by skin resident cells. Cell Death Dis. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Rosenbaum, J.T. Uveitis in spondyloarthritis including psoriatic arthritis, ankylosing spondylitis, and inflammatory bowel disease. Clin. Rheumatol. 2015, 34, 999–1002. [Google Scholar] [CrossRef]
- Rosenbaum, J.T. The eye in spondyloarthritis✰. Semin. Arthritis Rheum. 2019, 49, S29–S31. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Haroon, M.; Rosenbaum, J.T. Acute Anterior Uveitis and Spondyloarthritis: More Than Meets the Eye. Curr. Rheumatol. Rep. 2015, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, D.; Yates, W.; Amjadi, S.; McCluskey, P. HLA-B27 Anterior Uveitis: Immunology and Immunopathology. Ocul. Immunol. Inflamm. 2016, 24, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.-C.; Li, Q. Immunopathology of uveitis. Br. J. Ophthalmol. 1998, 82, 91–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.; Chen, W.; Jiang, R.; Zhang, R.; Wang, Y.; Wang, L.; Gordon, L.; Chen, L. Expression profile of IL-1 family cytokines in aqueous humor and sera of patients with HLA-B27 associated anterior uveitis and idiopathic anterior uveitis. Exp. Eye Res. 2015, 138, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Repo, H.; Leirisalo-Repo, M.; Koivuranta-Vaara, P. Exaggerated inflammatory responsiveness plays a part in the pathogenesis of HLA-B27 linked diseases—Hypothesis. Ann. Clin. Res. 1984, 16, 47–50. [Google Scholar]
- Yamada, M.; Shichi, H.; Yuasa, T.; Tanouchi, Y.; Mimura, Y. Superoxide in ocular inflammation: Human and experimental uveitis. J. Free. Radicals Biol. Med. 1986, 2, 111–117. [Google Scholar] [CrossRef]
- Sedgwick, J.B.; Bergstresser, P.R.; Hurd, E.R. Increased Granulocyte Adherence in Psoriasis and Psoriatic Arthritis. J. Investig. Dermatol. 1980, 74, 81–84. [Google Scholar] [CrossRef] [Green Version]
- Iijima, S.; Iwata, M.; Otsuka, F. Psoriatic Arthritis and Hypopyon-lridocyclitis. Possible mechanism of the association of psoriasis and anterior uveitis. Dermatology 1996, 193, 295–299. [Google Scholar] [CrossRef]
- Chang, J.H.; Hampartzoumian, T.; Everett, B.; Lloyd, A.; McCluskey, P.J.; Wakefield, D. Changes in Toll-like Receptor (TLR)-2 and TLR4 Expression and Function but Not Polymorphisms Are Associated with Acute Anterior Uveitis. Investig. Opthalmology Vis. Sci. 2007, 48, 1711–1717. [Google Scholar] [CrossRef]
- Erridge, C.; Pridmore, A.; Eley, A.; Stewart, J.; Poxton, I.R. Lipopolysaccharides of Bacteroides fragilis, Chlamydia trachomatis and Pseudomonas aeruginosa signal via Toll-like receptor 2. J. Med Microbiol. 2004, 53, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.H.; McCluskey, P.; Wakefield, D. Toll-like receptors in ocular immunity and the immunopathogenesis of inflammatory eye disease. Br. J. Ophthalmol. 2006, 90, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, T.; Ohashi, S.; Kawa, Y.; Takahama, H.; Ito, M.; Soma, Y.; Mizoguchi, M. Elevated Serum Granulocyte Colony-Stimulating Factor Levels in Patients With Active Phase of Sweet Syndrome and Patients With Active Behçet Disease: Implication in neutrophil apoptosis dysfunction. Arch. Dermatol. 2004, 140, 570–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Antiapoptotic Protein Mcl-1 Is Essential for the Survival of Neutrophils but Not Macrophages—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/17062731/ (accessed on 3 November 2022).
- Increased Production of Pro-Inflammatory Cytokines and Enhanced T Cell Responses after Activation of Human Dendritic Cells with IL-1 and CD40 Ligand—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/12385649/ (accessed on 3 November 2022).
- Goldberg, G.L.; Cornish, A.L.; Murphy, J.; Pang, E.S.; Lim, L.L.; Campbell, I.K.; Scalzo-Inguanti, K.; Chen, X.; McMenamin, P.G.; Maraskovsky, E.; et al. G-CSF and Neutrophils Are Nonredundant Mediators of Murine Experimental Autoimmune Uveoretinitis. Am. J. Pathol. 2016, 186, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Iijima, S.; Iwata, M.; Otsuka, F. Rheological analysis of peripheral blood neutrophils in a patient with cutaneous and arthropathic psoriasis accompanying hypopyon-iridocyclitis. Australas. J. Dermatol. 1996, 37 (Suppl. 1), S40–S41. [Google Scholar] [CrossRef]
- Morton, A.M.; Sefik, E.; Upadhyay, R.; Weissleder, R.; Benoist, C.; Mathis, D. Endoscopic photoconversion reveals unexpectedly broad leukocyte trafficking to and from the gut. Proc. Natl. Acad. Sci. USA 2014, 111, 6696–6701. [Google Scholar] [CrossRef] [Green Version]
- Hysa, E.; Cutolo, C.A.; Gotelli, E.; Pacini, G.; Schenone, C.; Kreps, E.O.; Smith, V.; Cutolo, M. Immunopathophysiology and clinical impact of uveitis in inflammatory rheumatic diseases: An update. Eur. J. Clin. Investig. 2021, 51, e13572. [Google Scholar] [CrossRef]
- Németh, T.; Sperandio, M.; Mócsai, A. Neutrophils as emerging therapeutic targets. Nat. Rev. Drug Discov. 2020, 19, 253–275. [Google Scholar] [CrossRef]
- De Benedetti, F.; Ruperto, N.; Baildam, E.; Burgos-Vargas, R.; Horneff, G.; Huppertz, H.I.; Minden, K.; Myones, B.L.; Onel, K.; Wang, J.; et al. A14: Neutropenia With Tocilizumab Treatment Is Not Associated With Increased Infection Risk in Patients with Systemic Juvenile Idiopathic Arthritis. Arthritis Rheumatol. 2014, 66, S23–S24. [Google Scholar] [CrossRef] [Green Version]
- Shovman, O.; Shoenfeld, Y.; Langevitz, P. Tocilizumab-induced neutropenia in rheumatoid arthritis patients with previous history of neutropenia: Case series and review of literature. Immunol. Res. 2015, 61, 164–168. [Google Scholar] [CrossRef]
- Strand, V.; Dikranian, A.; Beal, J.; Kwok, K.; Krishnaswami, S.; Wood, S.; Nduaka, C. Analysis of early neutropenia, clinical response, and serious infection events in patients receiving tofacitinib for rheumatoid arthritis. In Arthritis & Rheumatology; Wiley-Blackwell 111 River ST: Hoboken, NJ, USA, 2014; pp. S1086–S1087. [Google Scholar]
- Wigerblad, G.; Cao, Q.; Brooks, S.; Naz, F.; Gadkari, M.; Jiang, K.; Gupta, S.; O’Neil, L.; Dell’Orso, S.; Kaplan, M.J.; et al. Single-Cell Analysis Reveals the Range of Transcriptional States of Circulating Human Neutrophils. J. Immunol. 2022, 209, 772–782. [Google Scholar] [CrossRef]
- Burmester, G.R.; McInnes, I.B.; Kremer, J.; Miranda, P.; Korkosz, M.; Vencovsky, J.; Rubbert-Roth, A.; Mysler, E.; Sleeman, M.A.; Godwood, A.; et al. A randomised phase IIb study of mavrilimumab, a novel GM–CSF receptor alpha monoclonal antibody, in the treatment of rheumatoid arthritis. Ann. Rheum. Dis. 2017, 76, 1020–1030. [Google Scholar] [CrossRef]
- Cid, M.C.; Unizony, S.H.; Blockmans, D.; Brouwer, E.; Dagna, L.; Dasgupta, B.; Hellmich, B.; Molloy, E.; Salvarani, C.; Trapnell, B.C.; et al. Efficacy and safety of mavrilimumab in giant cell arteritis: A phase 2, randomised, double-blind, placebo-controlled trial. Ann. Rheum. Dis. 2022, 81, 653–661. [Google Scholar] [CrossRef]
- Weinblatt, M.E.; McInnes, I.B.; Kremer, J.M.; Miranda, P.; Vencovsky, J.; Guo, X.; White, W.I.; Ryan, P.C.; Godwood, A.; Albulescu, M.; et al. A Randomized Phase II b Study of Mavrilimumab and Golimumab in Rheumatoid Arthritis. Arthritis Rheumatol. 2018, 70, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Buckley, C.D.; Simón-Campos, J.A.; Zhdan, V.; Becker, B.; Davy, K.; Fisheleva, E.; Gupta, A.; Hawkes, C.; Inman, D.; Layton, M.; et al. Efficacy, patient-reported outcomes, and safety of the anti-granulocyte macrophage colony-stimulating factor antibody otilimab (GSK3196165) in patients with rheumatoid arthritis: A randomised, phase 2b, dose-ranging study. Lancet Rheumatol. 2020, 2, e677–e688. [Google Scholar] [CrossRef]
- Taylor, P.C.; for the NEXUS Study Group; Saurigny, D.; Vencovsky, J.; Takeuchi, T.; Nakamura, T.; Matsievskaia, G.; Hunt, B.; Wagner, T.; Souberbielle, B. Efficacy and safety of namilumab, a human monoclonal antibody against granulocyte-macrophage colony-stimulating factor (GM-CSF) ligand in patients with rheumatoid arthritis (RA) with either an inadequate response to background methotrexate therapy or an inadequate response or intolerance to an anti-TNF (tumour necrosis factor) biologic therapy: A randomized, controlled trial. Arthritis Res. Ther. 2019, 21, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp, K.; Gooderham, M.; Jenkins, R.; Vender, R.; Szepietowski, J.; Wagner, T.; Hunt, B.; Souberbielle, B.; on behalf of the NEPTUNE investigators; The NEPTUNE investigators. Granulocyte–macrophage colony-stimulating factor (GM-CSF) as a therapeutic target in psoriasis: Randomized, controlled investigation using namilumab, a specific human anti-GM-CSF monoclonal antibody. Br. J. Dermatol. 2019, 180, 1352–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaffen, S.L.; Jain, R.; Garg, A.V.; Cua, D.J. The IL-23–IL-17 immune axis: From mechanisms to therapeutic testing. Nat. Rev. Immunol. 2014, 14, 585–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, A.V.C.; Chiva, L.M.; Villarino, M.R. Granulocyte and monocyte/macrophage apheresis for the treatment of immune-mediated inflammatory arthropathies: Case reports. Drugs Context 2021, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-L.; Mao, J.-W.; Wang, Y.-D. Selective granulocyte and monocyte apheresis in inflammatory bowel disease: Its past, present and future. World J. Gastrointest. Pathophysiol. 2020, 11, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Gubatan, J.; Keyashian, K.; Rubin, S.J.; Wang, J.; Buckman, C.A.; Sinha, S. Anti-Integrins for the Treatment of Inflammatory Bowel Disease: Current Evidence and Perspectives. Clin. Exp. Gastroenterol. 2021, 14, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Schön, M.P.; Zollner, T.M.; Boehncke, W.-H. The Molecular Basis of Lymphocyte Recruitment to the Skin: Clues for Pathogenesis and Selective Therapies of Inflammatory Disorders. J. Investig. Dermatol. 2003, 121, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Jayne, D.R.; Merkel, P.A.; Schall, T.J.; Bekker, P. Avacopan for the Treatment of ANCA-Associated Vasculitis. N. Engl. J. Med. 2021, 384, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Huizenga, N.; Zonozi, R.; Rosenthal, J.; Laliberte, K.; Niles, J.L.; Cortazar, F.B. Treatment of Aggressive Antineutrophil Cytoplasmic Antibody–Associated Vasculitis With Eculizumab. Kidney Int. Rep. 2020, 5, 542–545. [Google Scholar] [CrossRef]
- Shawky, A.M.; Almalki, F.A.; Abdalla, A.N.; Abdelazeem, A.H.; Gouda, A.M. A Comprehensive Overview of Globally Approved JAK Inhibitors. Pharmaceutics 2022, 14, 1001. [Google Scholar] [CrossRef]
- Tanaka, Y.; Millson, D.; Iwata, S.; Nakayamada, S. Safety and efficacy of fostamatinib in rheumatoid arthritis patients with an inadequate response to methotrexate in phase II OSKIRA-ASIA-1 and OSKIRA-ASIA-1X study. Rheumatology 2021, 60, 2884–2895. [Google Scholar] [CrossRef]
- Walker, A.; Erwig, L.; Foster, K.; Nevin, K.; Wenzel, J.; Worm, M.; Williams, N.; Ratia, N.; Hoang, B.; Schneider-Merck, T.; et al. Safety, pharmacokinetics and pharmacodynamics of a topical SYK inhibitor in cutaneous lupus erythematosus: A double-blind Phase Ib study. Exp. Dermatol. 2021, 30, 1686–1692. [Google Scholar] [CrossRef]
- Keating, G.M. Apremilast: A Review in Psoriasis and Psoriatic Arthritis. Drugs 2017, 77, 459–472. [Google Scholar] [CrossRef]
- Taylor, P.C.; van der Heijde, D.; Landewé, R.; McCue, S.; Cheng, S.; Boonen, A. A Phase III Randomized Study of Apremilast, an Oral Phosphodiesterase 4 Inhibitor, for Active Ankylosing Spondylitis. J. Rheumatol. 2021, 48, 1259–1267. [Google Scholar] [CrossRef]
- Gerriets, V.; Goyal, A.; Khaddour, K. Tumor Necrosis Factor Inhibitors. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022; Available online: http://www.ncbi.nlm.nih.gov/books/NBK482425/ (accessed on 26 October 2022).
- Choy, E.H.; De Benedetti, F.; Takeuchi, T.; Hashizume, M.; John, M.R.; Kishimoto, T. Translating IL-6 biology into effective treatments. Nat. Rev. Rheumatol. 2020, 16, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Sandborn, W.J.; Bhandari, B.R.; Randall, C.; Younes, Z.H.; Romanczyk, T.; Xin, Y.; Wendt, E.; Chai, H.; McKevitt, M.; Zhao, S.; et al. Andecaliximab [Anti-matrix Metalloproteinase-9] Induction Therapy for Ulcerative Colitis: A Randomised, Double-Blind, Placebo-Controlled, Phase 2/3 Study in Patients With Moderate to Severe Disease. J. Crohns Colitis 2018, 12, 1021–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, S.; Siegel, C.A.; Friedenberg, K.A.; Younes, Z.H.; Seidler, U.; Bhandari, B.R.; Wang, K.; Wendt, E.; McKevitt, M.; Zhao, S.; et al. A Phase 2, Randomized, Placebo-Controlled Study Evaluating Matrix Metalloproteinase-9 Inhibitor, Andecaliximab, in Patients With Moderately to Severely Active Crohn’s Disease. J. Crohn’s Colitis 2018, 12, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, E.L.; Asher, J.L.; Molony, R.D.; Shaw, A.C.; Zeiss, C.J.; Wang, C.; Morozova-Roche, L.A.; Herzog, R.I.; Iwasaki, A.; Dixit, V.D. β-Hydroxybutyrate Deactivates Neutrophil NLRP3 Inflammasome to Relieve Gout Flares. Cell Rep. 2017, 18, 2077–2087. [Google Scholar] [CrossRef]
- Stefania, S.; Colia, R.; Cinzia, R.; Corrado, A.; Cantatore, F.P. Off-label use of anti-IL-1 drugs in rheumatic diseases. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211006584. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Zeng, F.; Nakamura, S.; Reid, K.T.; Gracey, E.; Lim, M.; Leng, L.; Jo, S.; Park, Y.-S.; Kusuda, M.; et al. Macrophage migration inhibitory factor drives pathology in a mouse model of spondyloarthritis and is associated with human disease. Sci. Transl. Med. 2021, 13, eabg1210. [Google Scholar] [CrossRef]
- Peiseler, M.; Kubes, P. More friend than foe: The emerging role of neutrophils in tissue repair. J. Clin. Investig. 2019, 129, 2629–2639. [Google Scholar] [CrossRef] [Green Version]
- The role of IL-17A in axial spondyloarthritis and psoriatic arthritis: Recent advances and controversies—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/31278139/ (accessed on 2 November 2022).
- Tamassia, N.; e Silva, F.A.; Calzetti, F.; Lonardi, S.; Gasperini, S.; Gardiman, E.; Bianchetto-Aguilera, F.; Gatta, L.B.; Girolomoni, G.; Mantovani, A.; et al. A Reappraisal on the Potential Ability of Human Neutrophils to Express and Produce IL-17 Family Members In Vitro: Failure to Reproducibly Detect It. Front. Immunol. 2018, 9, 795. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coletto, L.A.; Rizzo, C.; Guggino, G.; Caporali, R.; Alivernini, S.; D’Agostino, M.A. The Role of Neutrophils in Spondyloarthritis: A Journey across the Spectrum of Disease Manifestations. Int. J. Mol. Sci. 2023, 24, 4108. https://doi.org/10.3390/ijms24044108
Coletto LA, Rizzo C, Guggino G, Caporali R, Alivernini S, D’Agostino MA. The Role of Neutrophils in Spondyloarthritis: A Journey across the Spectrum of Disease Manifestations. International Journal of Molecular Sciences. 2023; 24(4):4108. https://doi.org/10.3390/ijms24044108
Chicago/Turabian StyleColetto, Lavinia Agra, Chiara Rizzo, Giuliana Guggino, Roberto Caporali, Stefano Alivernini, and Maria Antonietta D’Agostino. 2023. "The Role of Neutrophils in Spondyloarthritis: A Journey across the Spectrum of Disease Manifestations" International Journal of Molecular Sciences 24, no. 4: 4108. https://doi.org/10.3390/ijms24044108
APA StyleColetto, L. A., Rizzo, C., Guggino, G., Caporali, R., Alivernini, S., & D’Agostino, M. A. (2023). The Role of Neutrophils in Spondyloarthritis: A Journey across the Spectrum of Disease Manifestations. International Journal of Molecular Sciences, 24(4), 4108. https://doi.org/10.3390/ijms24044108