
Genomic Analysis of Corynebacterium diphtheriae Strains Isolated in the Years 2007–2022 with a Report on the Identification of the First Non-Toxigenic tox Gene-Bearing Strain in Poland


Abstract
:1. Introduction
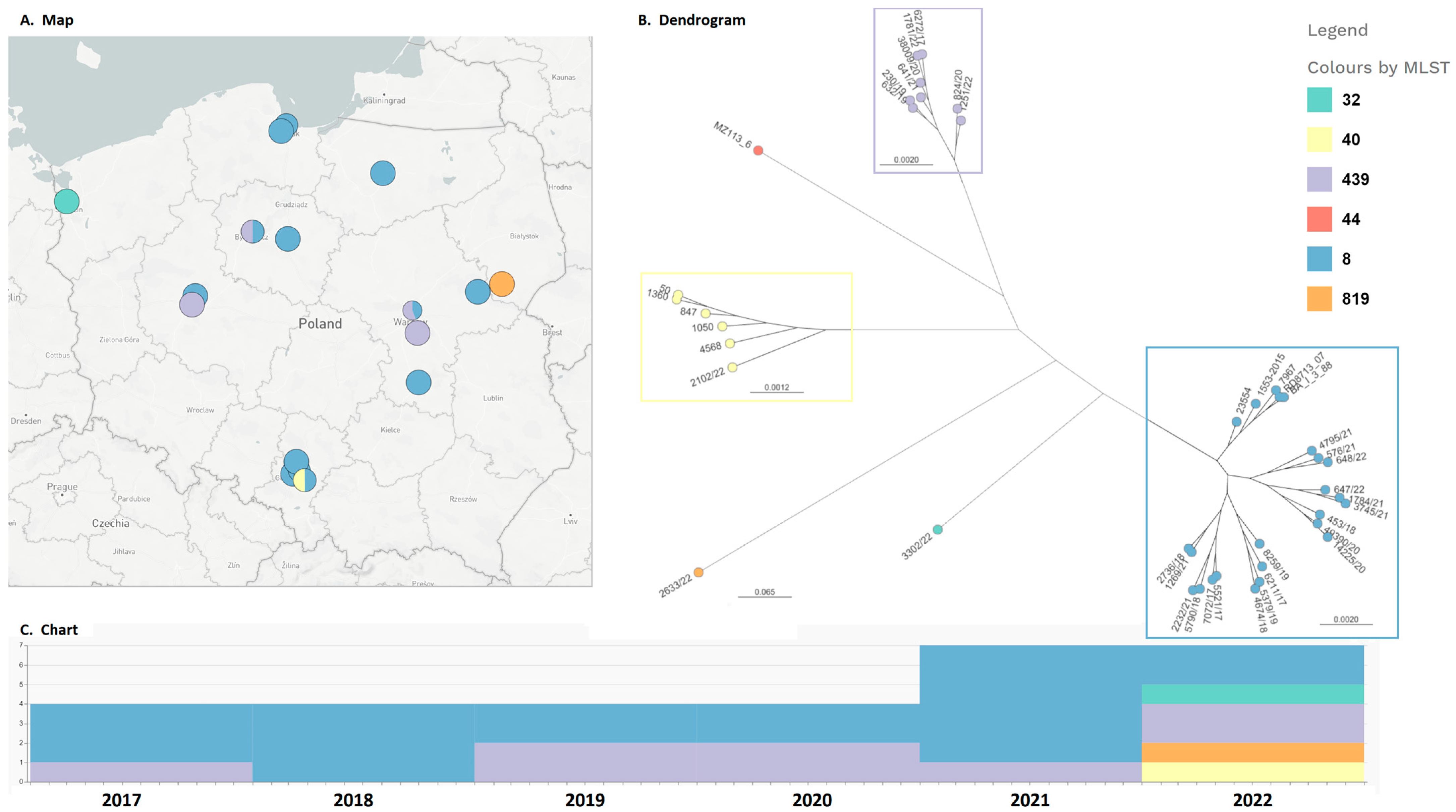
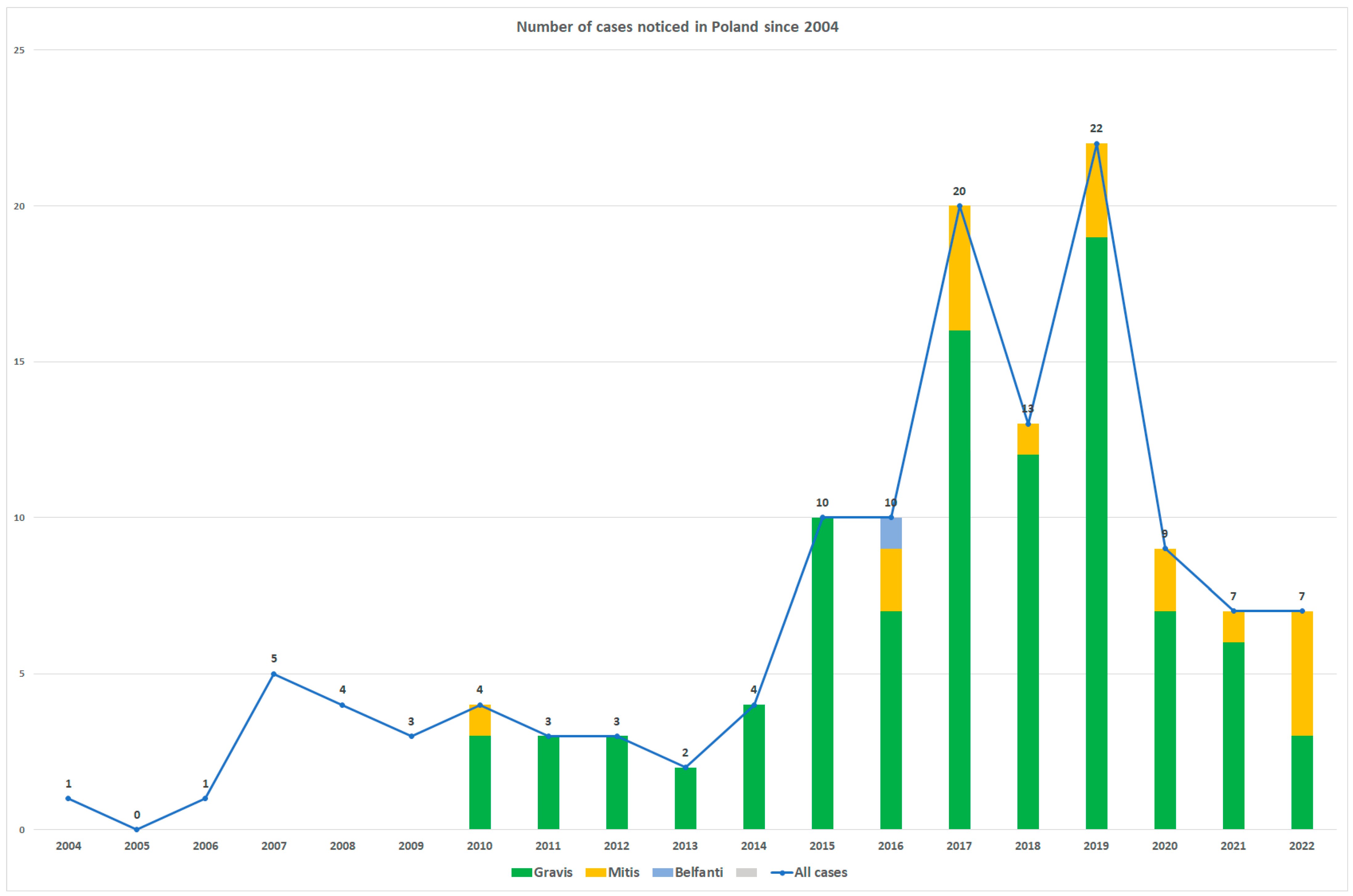
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Species Verification and Toxin Identification
4.3. Whole Genome Sequencing
4.4. WGS Data Analysis
4.5. Phylogenetic Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zasada, A.A. Nontoxigenic Highly Pathogenic Clone of Corynebacterium diphtheriae, Poland, 2004–2012. Emerg. Infect. Dis. 2013, 19, 1870–1872. [Google Scholar] [CrossRef] [PubMed]
- Zakikhany, K.; Neal, S.; Efstratiou, A. Emergence and Molecular Characterisation of Non-Toxigenic Tox Gene-Bearing Corynebacterium diphtheriae Biovar Mitis in the United Kingdom, 2003–2012. Euro Surveill. Bull. Eur. Sur. Mal. Transm. Eur. Commun. Dis. Bull. 2014, 19, 20819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangel, A.; Berger, A.; Konrad, R.; Bischoff, H.; Sing, A. Geographically Diverse Clusters of Nontoxigenic Corynebacterium diphtheriae Infection, Germany, 2016–2017. Emerg. Infect. Dis. 2018, 24, 1239–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zasada, A.A.; Baczewska-Rej, M.; Wardak, S. An Increase in Non-Toxigenic Corynebacterium diphtheriae Infections in Poland—Molecular Epidemiology and Antimicrobial Susceptibility of Strains Isolated from Past Outbreaks and Those Currently Circulating in Poland. Int. J. Infect. Dis. 2010, 14, e907–e912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubler, J.; Huber-Schneider, C.; Gruner, E.; Altwegg, M. An Outbreak of Nontoxigenic Corynebacterium diphtheriae Infection: Single Bacterial Clone Causing Invasive Infection among Swiss Drug Users. Clin. Infect. Dis. 1998, 27, 1295–1298. [Google Scholar] [CrossRef] [Green Version]
- Farfour, E.; Badell, E.; Zasada, A.; Hotzel, H.; Tomaso, H.; Guillot, S.; Guiso, N. Characterization and Comparison of Invasive Corynebacterium diphtheriae Isolates from France and Poland. J. Clin. Microbiol. 2012, 50, 173–175. [Google Scholar] [CrossRef] [Green Version]
- Fricchione, M.J.; Deyro, H.J.; Jensen, C.Y.; Hoffman, J.F.; Singh, K.; Logan, L.K. Non-Toxigenic Penicillin and Cephalosporin-Resistant Corynebacterium diphtheriae Endocarditis in a Child: A Case Report and Review of the Literature. J. Pediatr. Infect. Dis. Soc. 2014, 3, 251–254. [Google Scholar] [CrossRef] [Green Version]
- Zasada, A.A.; Zaleska, M.; Podlasin, R.B.; Seferyńska, I. The First Case of Septicemia Due to Nontoxigenic Corynebacterium diphtheriae in Poland: Case Report. Ann. Clin. Microbiol. Antimicrob. 2005, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Walory, J.; Grzesiowski, J.; Hryniewicz, W. The Prevalence of Diphtheria Immunity in Healthy Population in Poland. Epidemiol. Infect. 2001, 126, 225–230. [Google Scholar] [CrossRef]
- Sangal, V.; Hoskisson, P.A. Corynephages: Infections of the Infectors. In Corynebacterium diphtheriae and Related Toxigenic Species; Springer: Dordrecht, The Netherlands, 2014; pp. 67–81. ISBN 978-94-007-7623-4. [Google Scholar]
- Argimón, S.; Abudahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.T.G.; Yeats, C.A.; Grundmann, H.; et al. Microreact: Visualizing and Sharing Data for Genomic Epidemiology and Phylogeography. Microb. Genom. 2016, 2, e000093. [Google Scholar] [CrossRef] [Green Version]
- Czajka, U.; Wiatrzyk, A.; Mosiej, E.; Formińska, K.; Zasada, A.A. Changes in MLST Profiles and Biotypes of Corynebacterium diphtheriae Isolates from the Diphtheria Outbreak Period to the Period of Invasive Infections Caused by Nontoxigenic Strains in Poland (1950–2016). BMC Infect. Dis. 2018, 18, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeffer, J.; Huhulescu, S.; Stoeger, A.; Allerberger, F.; Ruppitsch, W. Assessing the Genetic Diversity of Austrian Corynebacterium diphtheriae Clinical Isolates, 2011 to 2019. J. Clin. Microbiol. 2021, 59, e02529-20. [Google Scholar] [CrossRef] [PubMed]
- Grosse-Kock, S.; Kolodkina, V.; Schwalbe, E.C.; Blom, J.; Burkovski, A.; Hoskisson, P.A.; Brisse, S.; Smith, D.; Sutcliffe, I.C.; Titov, L.; et al. Genomic Analysis of Endemic Clones of Toxigenic and Non-Toxigenic Corynebacterium diphtheriae in Belarus during and after the Major Epidemic in 1990s. BMC Genom. 2017, 18, 873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borisova, O.I.; Mazurova, I.K.; Chagina, I.A.; Pimenova, A.S.; Donskikh, E.E.; Aleshkin, V.A. Multilocus sequencing of Corynebacterium diphtheriae strains isolated in Russia in 2002–2012. Zh. Mikrobiol. Epidemiol. Immunobiol. 2013, 4, 17–23. [Google Scholar]
- Zasada, A.A.; Rzeczkowska, M. Nontoxigenic Corynebacterium diphtheriae Infections, Europe. Emerg. Infect. Dis. 2019, 25, 1437–1438. [Google Scholar] [CrossRef] [Green Version]
- Mandlik, A.; Swierczynski, A.; Das, A.; Ton-That, H. Corynebacterium diphtheriae Employs Specific Minor Pilins to Target Human Pharyngeal Epithelial Cells. Mol. Microbiol. 2007, 64, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Reardon-Robinson, M.E.; Ton-That, H. Assembly and Function of Corynebacterium diphtheriae Pili. In Corynebacterium diphtheriae and Related Toxigenic Species: Genomics, Pathogenicity and Applications; Burkovski, A., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 123–141. ISBN 978-94-007-7624-1. [Google Scholar]
- Sangal, V.; Blom, J.; Sutcliffe, I.C.; von Hunolstein, C.; Burkovski, A.; Hoskisson, P.A. Adherence and Invasive Properties of Corynebacterium diphtheriae Strains Correlates with the Predicted Membrane-Associated and Secreted Proteome. BMC Genom. 2015, 16, 765. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, R.S.; Antunes, C.A.; Lourêdo, L.S.; Viana, V.G.; Santos, C.S.D.; Fuentes Ribeiro da Silva, J.; Hirata, R.; Hacker, E.; Mattos-Guaraldi, A.L.; Burkovski, A. Functional Characterization of the Collagen-Binding Protein DIP2093 and Its Influence on Host-Pathogen Interaction and Arthritogenic Potential of Corynebacterium diphtheriae. Microbiol. Read. Engl. 2017, 163, 692–701. [Google Scholar] [CrossRef]
- Kolodkina, V.; Denisevich, T.; Titov, L. Identification of Corynebacterium Diphtheriae Gene Involved in Adherence to Epithelial Cells. Infect. Genet. Evol. 2011, 11, 518–521. [Google Scholar] [CrossRef]
- Ott, L.; Höller, M.; Gerlach, R.G.; Hensel, M.; Rheinlaender, J.; Schäffer, T.E.; Burkovski, A. Corynebacterium diphtheriae Invasion-Associated Protein (DIP1281) Is Involved in Cell Surface Organization, Adhesion and Internalization in Epithelial Cells. BMC Microbiol. 2010, 10, 2. [Google Scholar] [CrossRef] [Green Version]
- Sangal, V.; Hoskisson, P.A. Evolution, Epidemiology and Diversity of Corynebacterium diphtheriae: New Perspectives on an Old Foe. Infect. Genet. Evol. 2016, 43, 364–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farfour, E.; Badell, E.; Dinu, S.; Guillot, S.; Guiso, N. Microbiological Changes and Diversity in Autochthonous Non-Toxigenic Corynebacterium diphtheriae Isolated in France. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2013, 19, 980–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoefer, A.; Pampaka, D.; Herrera-León, S.; Peiró, S.; Varona, S.; López-Perea, N.; Masa-Calles, J.; Herrera-León, L. Molecular and Epidemiological Characterization of Toxigenic and Nontoxigenic Corynebacterium diphtheriae, Corynebacterium Belfantii, Corynebacterium Rouxii, and Corynebacterium Ulcerans Isolates Identified in Spain from 2014 to 2019. J. Clin. Microbiol. 2021, 59, e02410-20. [Google Scholar] [CrossRef] [PubMed]
- Chorlton, S.D.; Ritchie, G.; Lawson, T.; Romney, M.G.; Lowe, C.F. Whole-Genome Sequencing of Corynebacterium diphtheriae Isolates Recovered from an Inner-City Population Demonstrates the Predominance of a Single Molecular Strain. J. Clin. Microbiol. 2020, 58, e01651-19. [Google Scholar] [CrossRef]
- Timms, V.J.; Nguyen, T.; Crighton, T.; Yuen, M.; Sintchenko, V. Genome-Wide Comparison of Corynebacterium diphtheriae Isolates from Australia Identifies Differences in the Pan-Genomes between Respiratory and Cutaneous Strains. BMC Genom. 2018, 19, 869. [Google Scholar] [CrossRef]
- Doyle, C.J.; Mazins, A.; Graham, R.M.A.; Fang, N.-X.; Smith, H.V.; Jennison, A.V. Sequence Analysis of Toxin Gene-Bearing Corynebacterium diphtheriae Strains, Australia. Emerg. Infect. Dis. 2017, 23, 105–107. [Google Scholar] [CrossRef] [Green Version]
- Mel’nikov, V.G.; Kombarova, S.I.; Borisova, O.I.; Volozhantsev, N.V.; Verevkin, V.V.; Volkovoĭ, K.I.; Mazurova, I.K. Corynebacterium diphtheriae nontoxigenic strain carrying the gene of diphtheria toxin. Zh. Mikrobiol. Epidemiol. Immunobiol. 2004, 1, 3–7. [Google Scholar]
- World Health Organization. WHO Laboratory Manual for the Diagnosis of Diphtheria and Other Related Infections; World Health Organization: Geneva, Switzerland, 2021; ISBN 978-92-4-003805-9. [Google Scholar]
- Pallen, M.J.; Hay, A.J.; Puckey, L.H.; Efstratiou, A. Polymerase Chain Reaction for Screening Clinical Isolates of Corynebacteria for the Production of Diphtheria Toxin. J. Clin. Pathol. 1994, 47, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Hauser, D.; Popoff, M.R.; Kiredjian, M.; Boquet, P.; Bimet, F. Polymerase Chain Reaction Assay for Diagnosis of Potentially Toxinogenic Corynebacterium diphtheriae Strains: Correlation with ADP-Ribosylation Activity Assay. J. Clin. Microbiol. 1993, 31, 2720–2723. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.-H.; Liao, Y.-C. CISA: Contig Integrator for Sequence Assembly of Bacterial Genomes. PLoS ONE 2013, 8, e60843. [Google Scholar] [CrossRef] [PubMed]
- Czmil, A.; Wronski, M.; Czmil, S.; Sochacka-Pietal, M.; Cmil, M.; Gawor, J.; Wołkowicz, T.; Plewczynski, D.; Strzalka, D.; Pietal, M. NanoForms: An Integrated Server for Processing, Analysis and Assembly of Raw Sequencing Data of Microbial Genomes, from Oxford Nanopore Technology. PeerJ 2022, 10, e13056. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of Methods for Genomic Taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasman, H.; Saputra, D.; Sicheritz-Ponten, T.; Lund, O.; Svendsen, C.A.; Frimodt-Møller, N.; Aarestrup, F.M. Rapid Whole-Genome Sequencing for Detection and Characterization of Microorganisms Directly from Clinical Samples. J. Clin. Microbiol. 2014, 52, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Cosentino, S.; Larsen, M.V.; Aarestrup, F.M.; Lund, O. PathogenFinder—Distinguishing Friend from Foe Using Bacterial Whole Genome Sequence Data. PLoS ONE 2013, 8, e77302. [Google Scholar] [CrossRef]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Strain ID | Biotype | Year of Isolation | Patient’s Age | Voivodeship | Source | Other Information |
|---|---|---|---|---|---|---|
| 3302/22 | gravis | 2022 | 21 | Zachodniopomorskie | respiratory tract | |
| 2633/22 | mitis | 2022 | 90 | Podlaskie | wound | |
| 2102/22 | mitis | 2022 | 51 | Śląskie | blood | |
| 1781/22 | mitis | 2022 | 58 | Mazowieckie | unk | |
| 1251/22 | mitis | 2022 | 35 | Mazowieckie | wound | homeless |
| 648/22 | gravis | 2022 | 42 | Śląskie | blood | |
| 647/22 | gravis | 2022 | 40 | Śląskie | blood | |
| 4795/21 | gravis | 2021 | 60 | Śląskie | unk | |
| 3745/21 | gravis | 2021 | 39 | Śląskie | blood | |
| 2232/21 | gravis | 2021 | 17 | Mazowieckie | wound | |
| 1784/21 | gravis | 2021 | 41 | Śląskie | wound | homeless |
| 1269/21 | gravis | 2021 | 55 | Warmińsko-mazurskie | blood | |
| 641/21 | mitis | 2021 | 39 | Mazowieckie | respiratory tract | |
| 576/21 | gravis | 2021 | 65 | Mazowieckie | blood | |
| 14225/20 | gravis | 2020 | 42 | Pomorskie | blood | |
| 49390/20 | gravis | 2020 | 72 | Kujawsko-pomorskie | wound | |
| 38009/20 | mitis | 2020 | 62 | Kujawsko-pomorskie | eye | homeless * |
| 824/20 | mitis | 2020 | 35 | Wielkopolskie | blood | |
| 8259/19 | gravis | 2019 | 66 | Kujawsko-pomorskie | respiratory tract | |
| 5379/19 | gravis | 2019 | 59 | Pomorskie | blood | homeless |
| 632/19 | mitis | 2019 | 66 | Mazowieckie | respiratory tract | |
| 230/19 | mitis | 2019 | 53 | Mazowieckie | wound | homeless * |
| 4674/18 | gravis | 2018 | 54 | Wielkopolskie | blood | homeless |
| 2736/18 | gravis | 2018 | 45 | Pomorskie | blood | |
| 5790/18 | gravis | 2018 | 59 | Mazowieckie | blood | |
| 453/18 | gravis | 2018 | 64 | Śląskie | wound | homeless * |
| 7072/17 | gravis | 2017 | 63 | Mazowieckie | unk | homeless |
| 5521/17 | gravis | 2017 | 31 | Mazowieckie | wound | homeless * |
| 6272/17 | mitis | 2017 | 57 | Mazowieckie | wound | homeless * |
| 6211/17 | gravis | 2017 | 18 | Mazowieckie | blood | |
| 54/E | gravis | 2015 | 45 | Mazowieckie | wound | homeless |
| 25/E | gravis | 2009 | unk | Małopolskie | blood | |
| 42/E | gravis | 2007 | unk | Pomorskie | blood | |
| 17/E | gravis | 2007 | unk | Kujawsko-pomorskie | wound | |
| 18/E | gravis | 2007 | unk | Pomorskie | blood | |
| 7/B | mitis | 1990s | unk | unk | throat | Diphtheria (tox+) |
| Strain ID | MLST | Resistance Genes | tox | dtxR | SpaA-Type Pili (spaA, spaB, spaC, srtA) | SpaD-Type Pili (spaD, spaE, spaF, srtB, srtC) | SpaH-Type Pili (spaG, spaH, spaI, srtD, srtE) | Surface-Anchored Pilus (sapA, sapD) | ABC Transporter (fagA, fagB, fagC, fagD) | ABC-Type Haem Transporter (hmuT, hmuU, hmuV) | Siderophore-Dependent Iron Uptake System (irp6A, irp6B, irp6C) | Ciu Iron Uptake and Siderophore Biosynthesis System (ciuA, ciuB, ciuC, ciuD, ciuE) | RS 14030 | RS 14950 | RS 17590 | RS 19245 | mdbA | RS 23695 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gravis | ||||||||||||||||||
| 3302/22 | 32 | - | - | + | + | - | + | + | + | + | + | + | - | + | + | + | + | - |
| 648/22 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 647/22 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 4795/21 | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 3745/21 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 2232/21 | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 1784/21 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 641/21 | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 576/21 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 14225/20 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 49390/20 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 8259/19 | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 5379/19 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 4674/18 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 2736/18 | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 5790/18 | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 453/18 | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 7072/17 | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 5521/17 | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 6211/17 | 8 | - | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 54/E | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 42/E | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 25/E | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 18/E | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 17/E | 8 | sul1 | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Mitis | ||||||||||||||||||
| 2633/22 | 819 | - | - | + | +/- | - | - | +/- | + | + | + | + | - | + | + | + | + | - |
| 2102/22 | 40 | - | + | + | +/- | - | - | +/- | + | + | + | + | - | + | + | + | + | - |
| 1781/22 | 439 | - | - | + | +/- | +/- | - | +/- | + | + | + | + | + | + | + | + | + | - |
| 1251/22 | 439 | - | - | + | +/- | +/- | - | +/- | + | + | + | + | + | + | + | + | + | - |
| 1269/21 | 439 | - | - | + | +/- | +/- | - | +/- | + | + | + | + | + | + | + | + | + | - |
| 38009/20 | 439 | - | - | + | +/- | +/- | - | +/- | + | + | + | + | + | + | + | + | + | - |
| 824/20 | 439 | - | - | + | +/- | +/- | - | +/- | + | + | + | + | + | + | + | + | + | - |
| 632/19 | 439 | - | - | + | +/- | +/- | - | +/- | + | + | + | + | + | + | + | + | + | - |
| 230/19 | 439 | - | - | + | +/- | +/- | - | +/- | + | + | + | + | + | + | + | + | + | - |
| 6272/17 | 439 | - | - | + | +/- | +/- | - | +/- | + | + | + | + | + | + | + | + | + | - |
| 7/B | 44 | - | + | + | +/- | - | - | +/- | + | + | + | + | - | + | + | + | + | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wołkowicz, T.; Zacharczuk, K.; Zasada, A.A. Genomic Analysis of Corynebacterium diphtheriae Strains Isolated in the Years 2007–2022 with a Report on the Identification of the First Non-Toxigenic tox Gene-Bearing Strain in Poland. Int. J. Mol. Sci. 2023, 24, 4612. https://doi.org/10.3390/ijms24054612
Wołkowicz T, Zacharczuk K, Zasada AA. Genomic Analysis of Corynebacterium diphtheriae Strains Isolated in the Years 2007–2022 with a Report on the Identification of the First Non-Toxigenic tox Gene-Bearing Strain in Poland. International Journal of Molecular Sciences. 2023; 24(5):4612. https://doi.org/10.3390/ijms24054612
Chicago/Turabian StyleWołkowicz, Tomasz, Katarzyna Zacharczuk, and Aleksandra Anna Zasada. 2023. "Genomic Analysis of Corynebacterium diphtheriae Strains Isolated in the Years 2007–2022 with a Report on the Identification of the First Non-Toxigenic tox Gene-Bearing Strain in Poland" International Journal of Molecular Sciences 24, no. 5: 4612. https://doi.org/10.3390/ijms24054612
APA StyleWołkowicz, T., Zacharczuk, K., & Zasada, A. A. (2023). Genomic Analysis of Corynebacterium diphtheriae Strains Isolated in the Years 2007–2022 with a Report on the Identification of the First Non-Toxigenic tox Gene-Bearing Strain in Poland. International Journal of Molecular Sciences, 24(5), 4612. https://doi.org/10.3390/ijms24054612