
Regulation of De Novo Lipid Synthesis by the Small GTPase Rac1 in the Adipogenic Differentiation of Progenitor Cells from Mouse White Adipose Tissue

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Establishment and Characterization of an In Vitro Differentiation Assay Using Adipose Progenitor Cells Obtained from Mouse WAT
2.2. The Expression of Enzymes for De Novo Synthesis of Fatty Acids and Triacylglycerol during Differentiation of Adipose Progenitor Cells into Adipocytes In Vitro
2.3. The Expression and Phosphorylation of Transcription Factors during Differentiation of Adipose Progenitor Cells into Adipocytes In Vitro
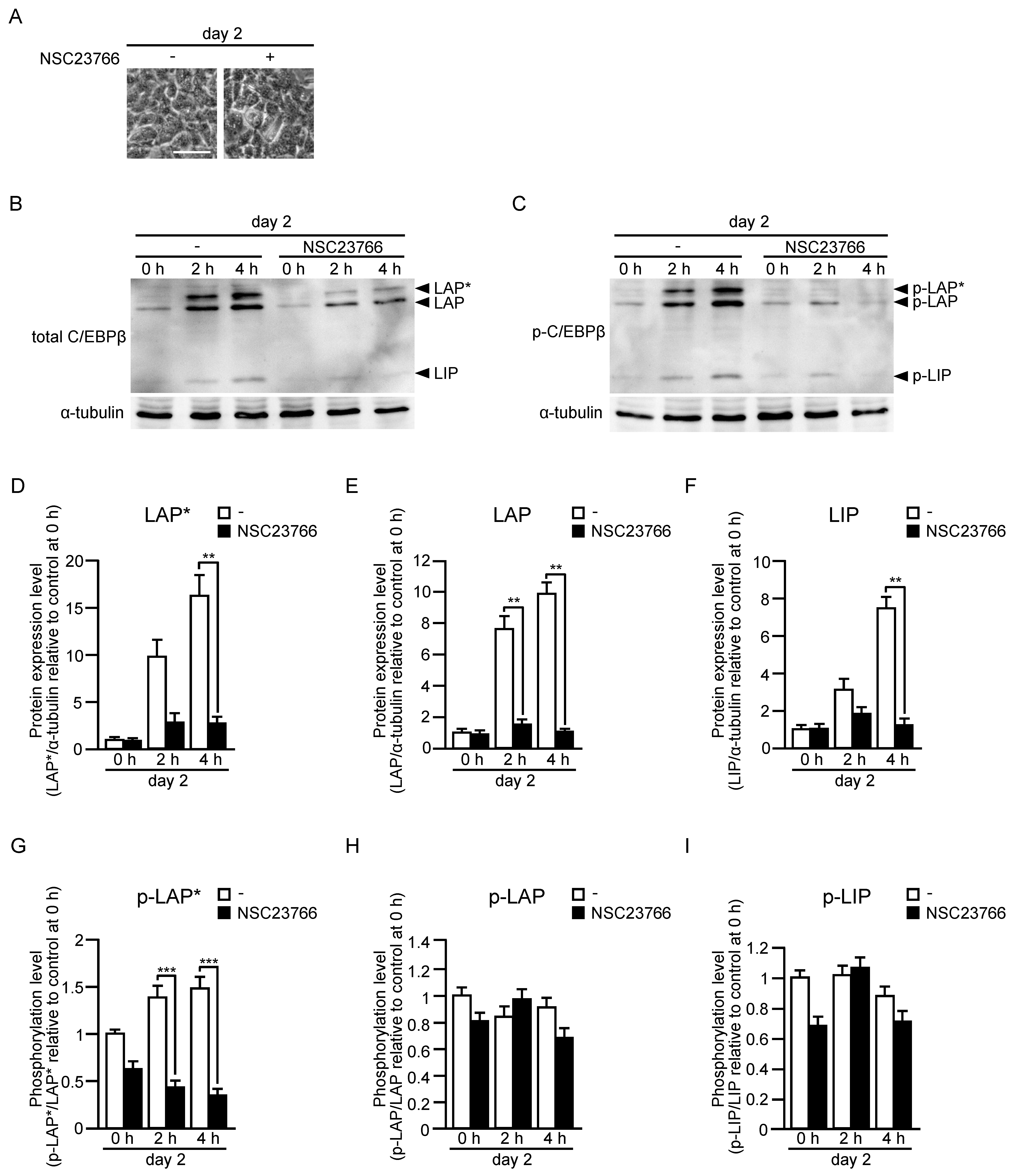
2.4. Protein and Phosphorylation Levels of C/EBPβ in the Early Stage of Differentiation of Adipose Progenitor Cells into Adipocytes In Vitro
2.5. Role of Rac1 in Differentiation of 3T3-L1 Cells into Adipocytes In Vitro
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animal Experiments
4.3. Conventional RT-PCR Analysis
4.4. Preparation of the SVF from Subcutaneous WAT
4.5. Induction of Differentiation of Adipose Progenitor Cells in the SVF into Adipocytes In Vitro
4.6. Quantitative RT-PCR Analysis
4.7. Immunofluorescent Microscopy
4.8. Immunoblot Analysis
4.9. Induction of Differentiation of 3T3-L1 Cells into Adipocytes In Vitro
4.10. shRNA-Mediated Knockdown of Rac1 in 3T3-L1 Cells
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACC | acetyl-CoA carboxylase |
| ACLY | ATP citrate lyase |
| adipo-rac1-KO | adipocyte-specific rac1 knockout |
| bZIP | basic leucine zipper |
| C/EBP | CCAAT/enhancer-binding protein |
| ChREBP | carbohydrate response element-binding protein |
| Dex | dexamethasone |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FASN | fatty acid synthase |
| FBS | fetal bovine serum |
| GPAT1 | glycerol-3-phosphate acyltransferase 1 |
| IBMX | 3-isobutyl-1-methylxanthine |
| PBS | phosphate-buffered saline |
| PI3K | phosphoinositide 3-kinase |
| p-LAP* | phosphorylated LAP* |
| p-LAP | phosphorylated LAP |
| p-LIP | phosphorylated LIP |
| PPARγ | peroxisome proliferator-activated receptor γ |
| RT-PCR | reverse transcriptase-polymerase chain reaction |
| SCD1 | stearoyl-CoA desaturase 1 |
| shRNA | small hairpin RNA |
| SREBP-1c | sterol regulatory element-binding protein 1c |
| SVF | stromal-vascular fraction |
| WAT | white adipose tissue |
References
- Rosen, E.D.; Spiegelman, B.M. What we talk about when we talk about fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Czech, M.P. The GLUT4 glucose transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, T. Molecular mechanisms for the regulation of insulin-stimulated glucose uptake by small guanosine triphosphatases in skeletal muscle and adipocytes. Int. J. Mol. Sci. 2014, 15, 18677–18692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaldin-Fincati, J.R.; Pavarotti, M.; Frendo-Cumbo, S.; Bilan, P.J.; Klip, A. Update on GLUT4 vesicle traffic: A cornerstone of insulin action. Trends Endocrinol. Metab. 2017, 28, 597–611. [Google Scholar] [CrossRef]
- Kane, S.; Sano, H.; Liu, S.C.; Asara, J.M.; Lane, W.S.; Garner, C.C.; Lienhard, G.E. A method to identify serine kinase substrates. Akt phosphorylates a novel adipocyte protein with a Rab GTPase-activating protein (GAP) domain. J. Biol. Chem. 2002, 277, 22115–22118. [Google Scholar] [CrossRef] [Green Version]
- Sano, H.; Eguez, L.; Teruel, M.N.; Fukuda, M.; Chuang, T.D.; Chavez, J.A.; Lienhard, G.E.; McGraw, T.E. Rab10, a target of the AS160 Rab GAP, is required for insulin-stimulated translocation of GLUT4 to the adipocyte plasma membrane. Cell Metab. 2007, 5, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, Y.; Zhang, J.; Deng, Y.; Jiang, L.; Song, E.; Wu, X.S.; Hammer, J.A.; Xu, T.; Lippincott-Schwartz, J. Rab10 and myosin-Va mediate insulin-stimulated GLUT4 storage vesicle translocation in adipocytes. J. Cell Biol. 2012, 198, 545–560. [Google Scholar] [CrossRef] [Green Version]
- Chiang, S.H.; Baumann, C.A.; Kanzaki, M.; Thurmond, D.C.; Watson, R.T.; Neudauer, C.L.; Macara, I.G.; Pessin, J.E.; Saltiel, A.R. Insulin-stimulated GLUT4 translocation requires the CAP-dependent activation of TC10. Nature 2001, 410, 944–948. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, N.; Nihata, Y.; Ueda, S.; Satoh, T. In situ detection of the activation of Rac1 and RalA small GTPases in mouse ad-ipocytes by immunofluorescent microscopy following in vivo and ex vivo insulin stimulation. Cell. Signal. 2017, 39, 108–117. [Google Scholar] [CrossRef]
- Takenaka, N.; Nakao, M.; Matsui, S.; Satoh, T. A crucial role for the small GTPase Rac1 downstream of the protein kinase Akt2 in insulin signaling that regulates glucose uptake in mouse adipocytes. Int. J. Mol. Sci. 2019, 2, 5443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenaka, N.; Nakao, M.; Hasegawa, K.; Chan, M.P.; Satoh, T. The guanine nucleotide exchange factor FLJ00068 activates Rac1 in adipocyte insulin signaling. FEBS Lett. 2020, 594, 4370–4380. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Takenaka, N.; Tanida, K.; Chan, M.P.; Sakata, M.; Aiba, A.; Satoh, T. Atrophy of white adipose tissue accompanied with decreased insulin-stimulated glucose uptake in mice lacking the small GTPase Rac1 specifically in adipocytes. Int. J. Mol. Sci. 2021, 22, 10753. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Kataoka, T.; Satoh, T. Activation of the small GTPase Rac1 by a specific guanine nucleotide exchange factor suffices to induce glucose uptake into skeletal muscle cells. Biol. Cell 2008, 100, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, N.; Nihata, Y.; Satoh, T. Rac1 activation caused by membrane translocation of a guanine nucleotide exchange factor in Akt2-mediated insulin signaling in mouse skeletal muscle. PLoS ONE 2016, 11, e0155292. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.W.; Leto, D.; Chiang, S.H.; Wang, Q.; Saltiel, A.R. Activation of RalA is required for insulin-stimulated Glut4 traf-ficking to the plasma membrane via the exocyst and the motor protein Myo1c. Dev. Cell 2007, 13, 391–404. [Google Scholar] [CrossRef]
- Moskalenko, S.; Henry, D.O.; Rosse, C.; Mirey, G.; Camonis, J.H.; White, M.A. The exocyst is a Ral effector complex. Nat. Cell Biol. 2002, 4, 66–72. [Google Scholar] [CrossRef]
- Sugihara, K.; Asano, S.; Tanaka, K.; Iwamatsu, A.; Okawa, K.; Ohta, Y. The exocyst complex binds the small GTPase RalA to mediate filopodia formation. Nat. Cell Biol. 2002, 4, 73–78. [Google Scholar] [CrossRef]
- Bose, A.; Guilherme, A.; Robida, S.I.; Nicoloro, S.M.; Zhou, Q.L.; Jiang, Z.Y.; Pomerleau, D.P.; Czech, M.P. Glucose transporter recycling in response to insulin is facilitated by myosin Myo1c. Nature 2002, 420, 821–824. [Google Scholar] [CrossRef]
- Nozaki, S.; Ueda, S.; Takenaka, N.; Kataoka, T.; Satoh, T. Role of RalA downstream of Rac1 in insulin-dependent glucose uptake in muscle cells. Cell. Signal. 2012, 24, 2111–2117. [Google Scholar] [CrossRef]
- Takenaka, N.; Sumi, Y.; Matsuda, K.; Fujita, J.; Hosooka, T.; Noguchi, T.; Aiba, A.; Satoh, T. Role for RalA downstream of Rac1 in skeletal muscle insulin signalling. Biochem. J. 2015, 469, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Morigny, P.; Boucher, J.; Arner, P.; Langin, D. Lipid and glucose metabolism in white adipocytes: Pathways, dysfunction and therapeutics. Nat. Rev. Endocrinol. 2021, 17, 276–295. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Xiaoli, A.M.; Yang, F. Regulation and metabolic significance of de novo lipogenesis in adipose tissues. Nutrients 2018, 10, 1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramji, D.P.; Foka, P. CCAAT/enhancer-binding proteins: Structure, function and regulation. Biochem. J. 2002, 365, 561–575. [Google Scholar] [CrossRef] [Green Version]
- Rodeheffer, M.S.; Birsoy, K.; Friedman, J.M. Identification of white adipocyte progenitor cells in vivo. Cell 2008, 135, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Berry, R.; Jeffery, E.; Rodeheffer, M.S. Weighing in on adipocyte precursors. Cell Metab. 2014, 19, 8–20. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, J.; Wang, X.; Yu, S.; Kershaw, E.E.; Chiu, P.C.; Dushay, J.; Estall, J.L.; Klein, U.; Maratos-Flier, E.; Rosen, E.D. Tran-scriptional control of adipose lipid handling by IRF4. Cell Metab. 2011, 13, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef]
- Guo, L.; Li, X.; Tang, Q.Q. Transcriptional regulation of adipocyte differentiation: A central role for CCAAT/enhancer-binding protein (C/EBP) β. J. Biol. Chem. 2015, 290, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Thompson, B.R.; Lobo, S.; Bernlohr, D.A. Fatty acid flux in adipocytes: The in’s and out’s of fat cell lipid trafficking. Mol. Cell. Endocrinol. 2010, 318, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Kunitomi, H.; Oki, Y.; Onishi, N.; Kano, K.; Banno, K.; Aoki, D.; Saya, H.; Nobusue, H. The insulin-PI3K-Rac1 axis contributes to terminal adipocyte differentiation through regulation of actin cytoskeleton dynamics. Genes Cells 2020, 25, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; DeYoung, S.M.; Zhang, M.; Zhang, M.; Cheng, A.; Saltiel, A.R. Changes in integrin expression during adipocyte differentiation. Cell Metab. 2005, 2, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhao, L.; Smas, C.; Sul, H.S. Pref-1 interacts with fibronectin to inhibit adipocyte differentiation. Mol. Cell. Biol. 2010, 30, 3480–3492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassai, H.; Terashima, T.; Fukaya, M.; Nakao, K.; Sakahara, M.; Watanabe, M.; Aiba, A. Rac1 in cortical projection neurons is selectively required for midline crossing of commissural axonal formation. Eur. J. Neurosci. 2008, 28, 257–267. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasegawa, K.; Takenaka, N.; Yamamoto, M.; Sakoda, Y.; Aiba, A.; Satoh, T. Regulation of De Novo Lipid Synthesis by the Small GTPase Rac1 in the Adipogenic Differentiation of Progenitor Cells from Mouse White Adipose Tissue. Int. J. Mol. Sci. 2023, 24, 4608. https://doi.org/10.3390/ijms24054608
Hasegawa K, Takenaka N, Yamamoto M, Sakoda Y, Aiba A, Satoh T. Regulation of De Novo Lipid Synthesis by the Small GTPase Rac1 in the Adipogenic Differentiation of Progenitor Cells from Mouse White Adipose Tissue. International Journal of Molecular Sciences. 2023; 24(5):4608. https://doi.org/10.3390/ijms24054608
Chicago/Turabian StyleHasegawa, Kiko, Nobuyuki Takenaka, Maaya Yamamoto, Yoshiki Sakoda, Atsu Aiba, and Takaya Satoh. 2023. "Regulation of De Novo Lipid Synthesis by the Small GTPase Rac1 in the Adipogenic Differentiation of Progenitor Cells from Mouse White Adipose Tissue" International Journal of Molecular Sciences 24, no. 5: 4608. https://doi.org/10.3390/ijms24054608
APA StyleHasegawa, K., Takenaka, N., Yamamoto, M., Sakoda, Y., Aiba, A., & Satoh, T. (2023). Regulation of De Novo Lipid Synthesis by the Small GTPase Rac1 in the Adipogenic Differentiation of Progenitor Cells from Mouse White Adipose Tissue. International Journal of Molecular Sciences, 24(5), 4608. https://doi.org/10.3390/ijms24054608