
Skin-to-Skin Contact: Crucial for Improving Behavior, Immunity, and Redox State after Short Cohabitation of Chronologically Old Mice and Prematurely Aging Mice with Adult Mice



Abstract
:1. Introduction
2. Results
2.1. Physical Contact Mediates Improvement of Behavioral Responses in Old Mice/PAM after Social Interaction with Adults/E-NPAM
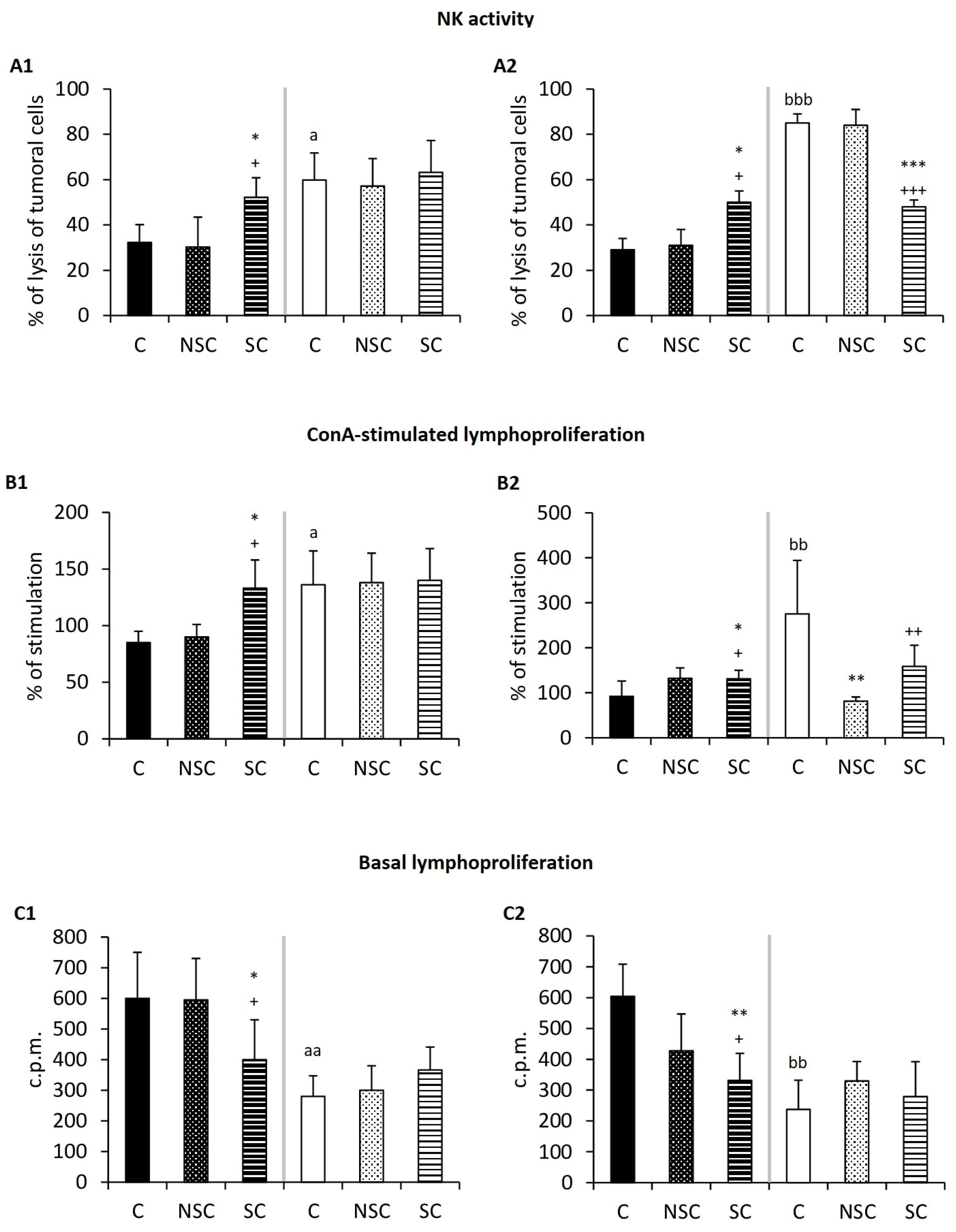
2.2. Physical Contact Mediates Improvement in the Immunity of Old Mice/PAM after Social Interaction with Adults/E-NPAM
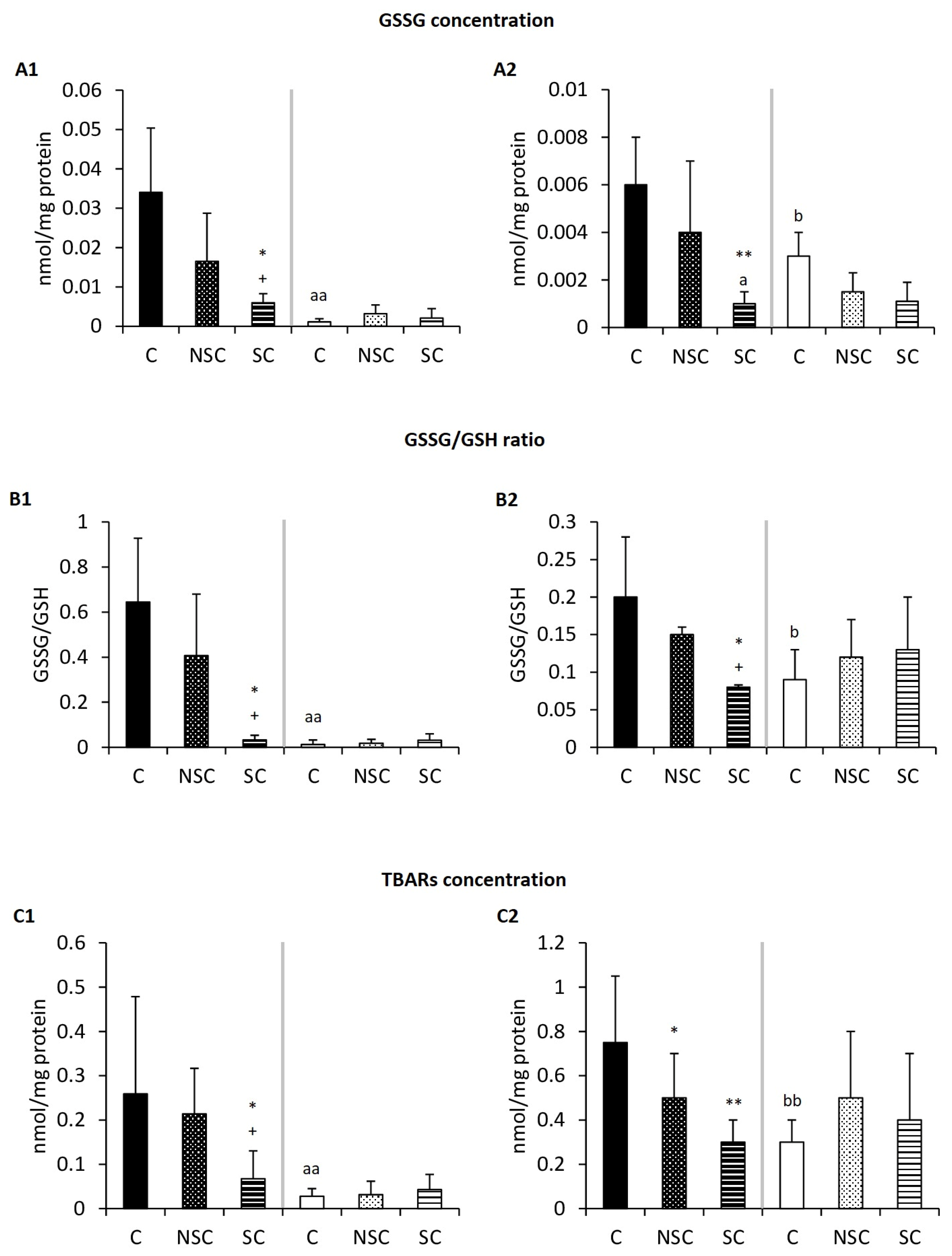
2.3. Physical Contact Mediates Improvement in the Redox Balance of Old Mice/PAM after Social Interaction with Adults/E-NPAM
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Classification of Prematurely Aging Mice (PAM) Using the T-Maze
4.3. Experimental Design
4.4. Behavioral Tests
4.4.1. Wood-Rod Test
4.4.2. Tightrope Test
4.4.3. Elevated-Plus Maze
4.4.4. Hole-Board Test
4.5. Extraction of Peritoneal Leukocytes
4.6. Evaluation of Immune Functions
4.6.1. Phagocytic Activity
4.6.2. Chemotaxis Capacity
4.6.3. Natural killer Cytotoxicity
4.6.4. Lymphoproliferation
4.7. Oxidative Parameters
4.7.1. Glutathione Peroxidase (GPx) Activity
4.7.2. Glutathione Reductase (GR) Activity
4.7.3. Glutathione Concentration
4.7.4. Lipid Peroxidation: TBARS Concentration
4.8. Protein Concentration
4.9. Longevity Study
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De la Fuente, M.; Miquel, J. An update of the oxidation-inflammation theory of aging: The involvement of the immune system in oxi-inflamm-aging. Curr. Pharm. Des. 2009, 15, 3003–3026. [Google Scholar] [CrossRef] [PubMed]
- Martínez de Toda, I.; Ceprián, N.; Díaz-Del Cerro, E.; De la Fuente, M. The role of immune cells in oxi-inflamm-aging. Cells 2021, 10, 2974. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Spencer, R.L.; Hutchison, K.E. Alcohol, aging, and the stress response. Alcohol Res. Health 1999, 23, 272–283. [Google Scholar]
- Aschbacher, K.; O’Donovan, A.; Wolkowitz, O.M.; Dhabhar, F.S.; Su, Y.; Epel, E. Good stress, bad stress and oxidative stress: Insights from anticipatory cortisol reactivity. Psychoneuroendocrinology 2013, 38, 1698–1708. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente, M.; Miñano, M.; Manuel Victor, V.; Del Rio, M.; Ferrández, M.D.; Díez, A.; Miquel, J. Relation between exploratory activity and immune function in aged mice: A preliminary study. Mech. Ageing Dev. 1998, 102, 263–277. [Google Scholar] [CrossRef]
- Guayerbas, N.; Puerto, M.; Víctor, V.M.; Miquel, J.; De la Fuente, M. Leukocyte function and life span in a murine model of premature immunosenescence. Exp. Gerontol. 2002, 37, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Álvarez, L.; Baeza, I.; Arranz, L.; Marco, E.M.; Borcel, E.; Guaza, C.; Viveros, M.P.; De la Fuente, M. Behavioral, endocrine and immunological characteristics of a murine model of premature aging. Dev. Comp. Immunol. 2005, 29, 965–976. [Google Scholar] [CrossRef]
- Vida, C.; Fuente, M. Stress-related Behavioural Responses, Immunity and Ageing in Animal Models. In Immunosenescence; Bosch, J., Phillips, A., Lord, J., Eds.; Springer: New York, NY, USA, 2013; Chapter 8. [Google Scholar] [CrossRef]
- Martínez de Toda, I.; Maté, I.; Vida, C.; Cruces, J.; De la Fuente, M. Immune function parameters as markers of biological age and predictors of longevity. Aging 2016, 8, 3110–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez de Toda, I.; Vida, C.; Garrido, A.; De la Fuente, M. redox parameters as markers of the rate of aging and predictors of life span. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Garrido, A.; Cruces, J.; Ceprián, N.; Vara, E.; De la Fuente, M. Oxidative-inflammatory stress in immune cells from adult mice with premature aging. Int. J. Mol. Sci. 2019, 20, 769. [Google Scholar] [CrossRef] [Green Version]
- Seeman, T.E.; Crimmins, E. Social environment effects on health and aging: Integrating epidemiologic and demographic approaches and perspectives. Ann. N. Y. Acad. Sci. 2001, 954, 88–117. [Google Scholar] [CrossRef]
- Alves, G.J.; Palermo-Neto, J. Odor cues released by Ehrlich tumor-bearing mice are aversive and induce psychological stress. Neuroimmunomodulation 2015, 22, 121–129. [Google Scholar] [CrossRef]
- Deak, T.; Kudinova, A.; Lovelock, D.F.; Gibb, B.E.; Hennessy, M.B. A multispecies approach for understanding neuroimmune mechanisms of stress. Dialogues Clin. Neurosci. 2017, 19, 37–53. [Google Scholar] [CrossRef]
- Hamasato, E.K.; de Lima, A.P.; de Oliveira, A.P.; dos Santos Franco, A.L.; de Lima, W.T.; Palermo-Neto, J. Cohabitation with a sick partner increases allergic lung inflammatory response in mice. Brain Behav. Immun. 2014, 42, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Garrido, A.; Cruces, J.; Ceprián, N.; De la Fuente, M. improvements in behavior and immune function and increased life span of old mice cohabiting with adult animals. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 873–881. [Google Scholar] [CrossRef]
- Garrido, A.; Cruces, J.; Ceprián, N.; Corpas, I.; Tresguerres, J.A.; De la Fuente, M. Social environment improves immune function and redox state in several organs from prematurely aging female mice and increases their lifespan. Biogerontology 2019, 20, 49–69. [Google Scholar] [CrossRef]
- Garrido, A.; Cruces, J.; Ceprián, N.; Díaz-Del Cerro, E.; Félix, J.; De la Fuente, M. The ratio of prematurely aging to non-prematurely aging mice cohabiting, conditions their behavior, immunity and lifespan. J. Neuroimmunol. 2020, 343, 577240. [Google Scholar] [CrossRef]
- Díaz-Del Cerro, E.; Ceprián, N.; Félix, J.; De la Fuente, M. A short social interaction between adult and old mice improves the homeostatic systems and increases healthy longevity. Exp. Gerontol. 2022, 158, 111653. [Google Scholar] [CrossRef]
- Díaz-Del Cerro, E.; Félix, J.; De la Fuente, M. Prematurely aging female mice improve their behavioural response, immunity, redox state, and lifespan after a short social interaction with non-prematurely aging mice. Biogerontology 2022, 23, 307–324. [Google Scholar] [CrossRef]
- McGlone, F.; Wessberg, J.; Olausson, H. Discriminative and affective touch: Sensing and feeling. Neuron 2014, 82, 737–755. [Google Scholar] [CrossRef] [Green Version]
- Field, T. Touch for socioemotional and physical well-being: A review. Dev. Rev. 2010, 30, 367–383. [Google Scholar] [CrossRef]
- Salchner, P.; Lubec, G.; Singewald, N. Decreased social interaction in aged rats may not reflect changes in anxiety-related behaviour. Behav. Brain Res. 2004, 151, 1–8. [Google Scholar] [CrossRef]
- Roosterman, D.; Goerge, T.; Schneider, S.W.; Bunnett, N.W.; Steinhoff, M. Neuronal control of skin function: The skin as a neuroimmunoendocrine organ. Physiol. Rev. 2006, 86, 1309–1379. [Google Scholar] [CrossRef]
- Morrison, I. Keep calm and cuddle on: Social touch as a stress buffer. Adapt. Hum. Behav. Physiol. 2016, 2, 344–362. [Google Scholar] [CrossRef] [Green Version]
- Cannizzo, E.S.; Clement, C.C.; Sahu, R.; Follo, C.; Santambrogio, L. Oxidative stress, inflamm-aging and immunosenescence. J Proteom. 2011, 74, 2313–2323. [Google Scholar] [CrossRef]
- Kard’ar, R. The ratio of oxidized and reduced forms of selected antioxidants as a possible marker of oxidative stress in humans. Biomed. Chromatogr. 2016, 30, 13–28. [Google Scholar]
- Martínez de Toda, I.; Garrido, A.; Vida, C.; Gómez-Cabrera, M.C.; Viña, J.; De la Fuente, M. Frailty quantified by the “valencia score” as a potential predictor of lifespan in mice. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 1323–1329. [Google Scholar] [CrossRef] [Green Version]
- Finkel, J.C.; Besch, V.G.; Hergen, A.; Kakarena, J.; Pohida, T. Effects of aging on current vocalization threshold in mice measured by a novel nociception assay. Anesthesiology 2006, 105, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Osada, K.; Yamazaki, K.; Curran, M.; Bard, J.; Smith, B.P.C.; Beauchamp, G.K. The scent of age. Proc. R. Soc. Lond. B 2003, 270, 929–933. [Google Scholar] [CrossRef]
- Brudzynsky, S.M. Ethotransmission: Communication of emotional states through ultrasonic vocalization in rats. Curr. Op. Neurobiol. 2013, 3, 310–317. [Google Scholar] [CrossRef]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress and the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Klöting, I.; Nitschke, C.; van den Brandt, J. Impact of genetic profiles on experimental studies: Outbred versus wild rats. Toxicol. Appl. Pharmacol. 2003, 189, 68–71. [Google Scholar] [CrossRef]
- Tappa, B.; Amao, H.; Ogasa, A.; Takahashi, K.W. Changes in the estrous cycle and number of ovulated and fertilized ova in aging female IVCS mice. Jikken Dobutsu 1989, 38, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Baeza, I.; De Castro, N.M.; Gimenez-Llort, L.; De la Fuente, M. Ovariectomy, a model of menopause in rodents, causes a premature aging of the nervous and immune systems. J. Neuroimmunol. 2010, 219, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Pellow, S.; Chopin, P.; File, S.E.; Briley, M. Validation of open: Closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods 1985, 14, 149–167. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, C.; Álvarez, P.; Jiménez, L.; De la Fuente, M. Oxidative stress in leukocytes from young prematurely aging mice is reversed by supplementation with biscuits rich in antioxidants. Dev. Comp. Immunol. 2006, 30, 1168–1180. [Google Scholar] [CrossRef]
- Massey, V.; Williams, C. On the reaction mechanism of yeast glutathione reductase. J. Biol. Chem. 1965, 240, 4470–4481. [Google Scholar] [CrossRef] [PubMed]
- Hissin, P.J.; Hilf, R. A fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Behavioral Parameters | Old Mice | Adult Mice | PAM | E-NPAM | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | NSC | SC | C | NSC | SC | C | NSC | SC | C | NSC | SC | |
| Wood-rod test | ||||||||||||
| Latency to leave the starting segment (s) | 6 ± 1 | 4 ± 1 | 3 ± 0.4 * | 3 ± 2 a | 3 ± 2 | 3 ± 2 | 4 ± 2 | 4 ± 2 | 7 ± 4 | 4 ± 2 | 3 ± 2 | 3 ± 2 |
| Tight-rope test | ||||||||||||
| Latency to fall (s) | 14 ± 6 | 4 ± 2 * | 14 ± 8 + | 19 ± 10 | 26 ± 12 | 21 ± 15 | 10 ± 4 | 6 ± 3 | 30 ± 9 * | 48 ± 10 b | 19 ± 8 **++ | 4 ± 2 ***++ |
| Animals that fell | 5/6 | 5/6 | 3/6 | 2/7 a | 2/10 | 3/10 | 5/6 | 6/6 | 3/6 | 4/7 | 4/6 | 4/7 |
| Hole-board test | ||||||||||||
| Total locomotion | 279 ± 24 | 353 ± 105 | 241 ± 27 | 250 ± 115 | 288 ± 64 | 369 ± 50 * | 250 ± 36 | 310 ± 12 ** | 327 ± 29 ** | 281 ± 29 | 274 ± 26 | 324 ± 38 *+ |
| External locomotion | 158 ± 29 | 223 ± 93 | 147 ± 32 | 130 ± 73 | 165 ± 43 | 226 ± 45 *+ | 176 ± 27 | 193 ± 26 | 194 ± 17 | 146 ± 27 | 145 ± 20 | 153 ± 32 |
| N° of rearings | 17 ± 3 | 22 ± 13 | 22 ± 10 | 19 ± 7 | 21 ± 6 | 26 ±8 | 21 ± 6 | 15 ± 3 | 21 ± 7 | 22 ± 8 | 21 ± 8 | 20 ± 6 |
| N° of head-dipping | 19 ± 4 | 12 ± 3 | 15 ± 2 | 24 ± 6 | 20 ± 5 | 22 ± 5 | 16 ± 3 | 20 ± 6 | 18 ± 3 | 16 ± 4 | 20 ± 5 | 19 ± 7 |
| Elevated-plus maze | ||||||||||||
| Time (s) spent in central platform | 100 ± 27 | 75 ± 26 | 94 ± 39 | 105 ± 35 | 101 ± 27 | 96 ± 39 | 74 ± 20 | 81 ± 14 | 74 ± 11 | 64 ± 18 | 81 ± 5 | 76 ± 10 |
| Time (s) spent in closed arms | 101 ± 36 | 99 ± 26 | 130 ± 42 | 155 ± 46 | 149 ± 57 | 126 ± 39 | 179 ± 8 ** | 151 ± 8 ** | 105 ± 39 ** + | 66 ± 32 bb | 118 ± 10 ** | 119 ± 14 ** |
| N° of head-dippings | 41 ± 9 | 43 ± 10 | 27 ± 12 | 21 ± 8 | 21 ± 9 | 29 ± 10 | 33 ± 10 | 44 ± 6 | 49 ± 8 | 55 ± 10 b | 54 ± 10 | 57 ± 8 |
| N° of rearings | 12 ± 3 | 33 ± 8 * | 43 ± 14 * | 52 ± 15 aa | 50 ± 19 | 42 ± 13 | 12 ± 4 | 13 ± 6 | 9 ± 6 | 20 ± 6 b | 13 ± 2 | 14 ± 4 |
| Immune Parameters | Old Mice | Adult Mice | PAM | E-NPAM | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | NSC | SC | C | NSC | SC | C | NSC | SC | C | NSC | SC | |
| Chemotaxis index of macrophages | 320 ± 52 | 566 ± 281 | 445 ± 178 | 585 ± 163 aa | 566 ± 164 | 625 ± 200 | 165 ± 5 | 189 ± 94 | 401 ± 45 | 1069 ± 300 bb | 534 ± 260 | 1051 ± 529 |
| Chemotaxis index of lymphocytes | 207 ± 112 | 542 ± 244 * | 735 ± 190 *+ | 710 ± 144 aa | 619 ± 215 | 611 ± 291 | 306 ± 11 | 300 ± 17 | 268 ± 57 | 1141 ± 162 bb | 295 ± 207 * | 663 ± 249 * |
| Phagocytic efficacy (%) | 31 ± 10 | 57 ± 17 * | 51 ± 6 * | 58 ± 11 a | 54 ± 5 | 69 ± 7 | 184 ± 15 | 442 ± 60 | 322 ± 74 | 445 ± 209 b | 371 ± 144 | 638 ± 252 |
| Phagocytic index | 93 ± 24 | 180 ± 68 * | 146 ± 72 * | 505 ± 185 aa | 747 ± 93 | 695 ± 83 | 309 ± 19 | 408 ± 24 ** | 356 ± 31 ** | 378 ± 43 b | 317 ± 64 | 412 ± 18 *+ |
| LPS-stimulated lymphoproliferation (%) | 90 ± 10 | 92 ± 7 | 100 ± 12 | 150 ± 15 a | 143 ± 13 | 155 ± 12 | 84 ± 38 | 92 ± 24 | 94 ± 20 | 219 ± 62 b | 139 ± 61 | 174 ± 42 |
| Redox Parameters | Old Mice | Adult Mice | PAM | E-NPAM | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | NSC | SC | C | NSC | SC | C | NSC | SC | C | NSC | SC | |
| GR activity (mU/mg protein) | 12 ± 4 | 13 ± 2 | 15 ± 7 | 8 ± 1 | 10 ± 5 | 46 ± 5 bb | 2.6 ± 1 | 1.3 ± 0.8 | 8 ± 2 | 1.6 ± 0.5 | 1.5 ± 1 | 6 ± 3 |
| GPx activity (mU/mg protein) | 5 ± 3 | 6 ± 2 | 20 ± 5 **++ | 40 ± 10 aa | 35 ± 12 | 41 ± 9 | 4 ± 3 | 3.6 ± 2.8 | 5 ± 3 | 0.3 ± 0.3 | 0.5 ± 0.2 | 0.3 ± 0.1 |
| GSH concentration (nmol/mg protein) | 0.13 ± 0.03 | 0.09 ± 0.04 | 0.33 ± 0.1 *+ | 0.36 ± 0.2 a | 0.26 ± 0.04 | 0.33 ± 0.16 | 0.14 ± 0.1 | 0.2 ± 0.05 | 0.24 ± 0.08 | 0.3 ± 0.2 b | 0.28 ± 0.2 | 0.22 ± 0.1 |
| Old Mice | Adult Mice | PAM | E-NPAM | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | NSC | SC | C | NSC | SC | C | NSC | SC | C | NSC | SC | |
| Mean longevity (weeks) | 87 ± 11 | 116 ± 11 | 96 ± 13 | 90 ± 5 | 108 ± 1 | 100 ± 9 | 73 ± 12 | 74 ± 17 | 97 ± 15 *+ | 110 ± 16 b | 93 ± 19 | 103 ± 19 |
| Maximum longevity (weeks) | 112 | 115 | 155 *+ | 118 | 125 | 140 * | 92 | 94 | 113 *+ | 129 b | 125 | 122 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Del Cerro, E.; Félix, J.; De la Fuente, M. Skin-to-Skin Contact: Crucial for Improving Behavior, Immunity, and Redox State after Short Cohabitation of Chronologically Old Mice and Prematurely Aging Mice with Adult Mice. Int. J. Mol. Sci. 2023, 24, 4680. https://doi.org/10.3390/ijms24054680
Díaz-Del Cerro E, Félix J, De la Fuente M. Skin-to-Skin Contact: Crucial for Improving Behavior, Immunity, and Redox State after Short Cohabitation of Chronologically Old Mice and Prematurely Aging Mice with Adult Mice. International Journal of Molecular Sciences. 2023; 24(5):4680. https://doi.org/10.3390/ijms24054680
Chicago/Turabian StyleDíaz-Del Cerro, Estefanía, Judith Félix, and Mónica De la Fuente. 2023. "Skin-to-Skin Contact: Crucial for Improving Behavior, Immunity, and Redox State after Short Cohabitation of Chronologically Old Mice and Prematurely Aging Mice with Adult Mice" International Journal of Molecular Sciences 24, no. 5: 4680. https://doi.org/10.3390/ijms24054680
APA StyleDíaz-Del Cerro, E., Félix, J., & De la Fuente, M. (2023). Skin-to-Skin Contact: Crucial for Improving Behavior, Immunity, and Redox State after Short Cohabitation of Chronologically Old Mice and Prematurely Aging Mice with Adult Mice. International Journal of Molecular Sciences, 24(5), 4680. https://doi.org/10.3390/ijms24054680