
Salt-Tolerant Plants, Halophytes, as Renewable Natural Resources for Cancer Prevention and Treatment: Roles of Phenolics and Flavonoids in Immunomodulation and Suppression of Oxidative Stress towards Cancer Management


Abstract
:1. Introduction
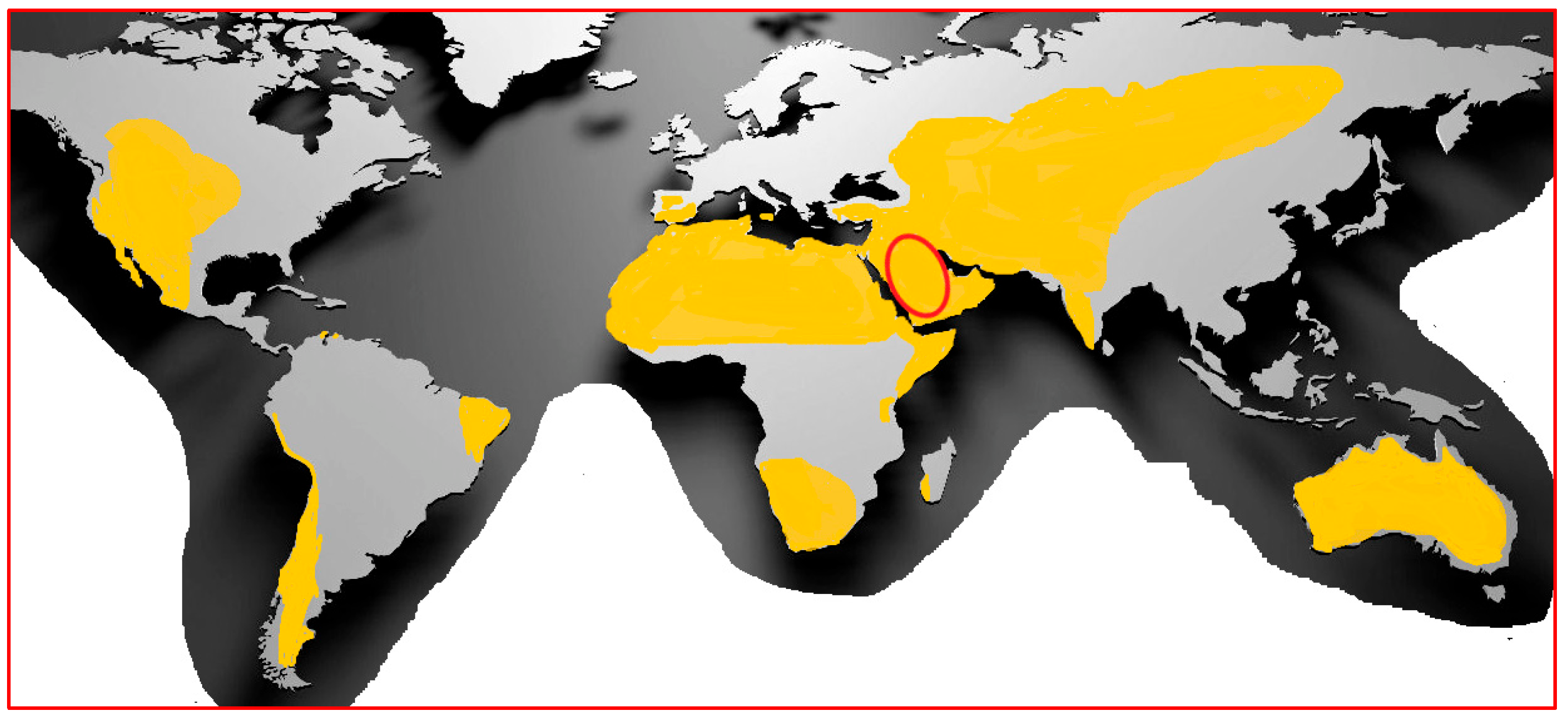
2. Salt-Tolerant Plants: Halophytes
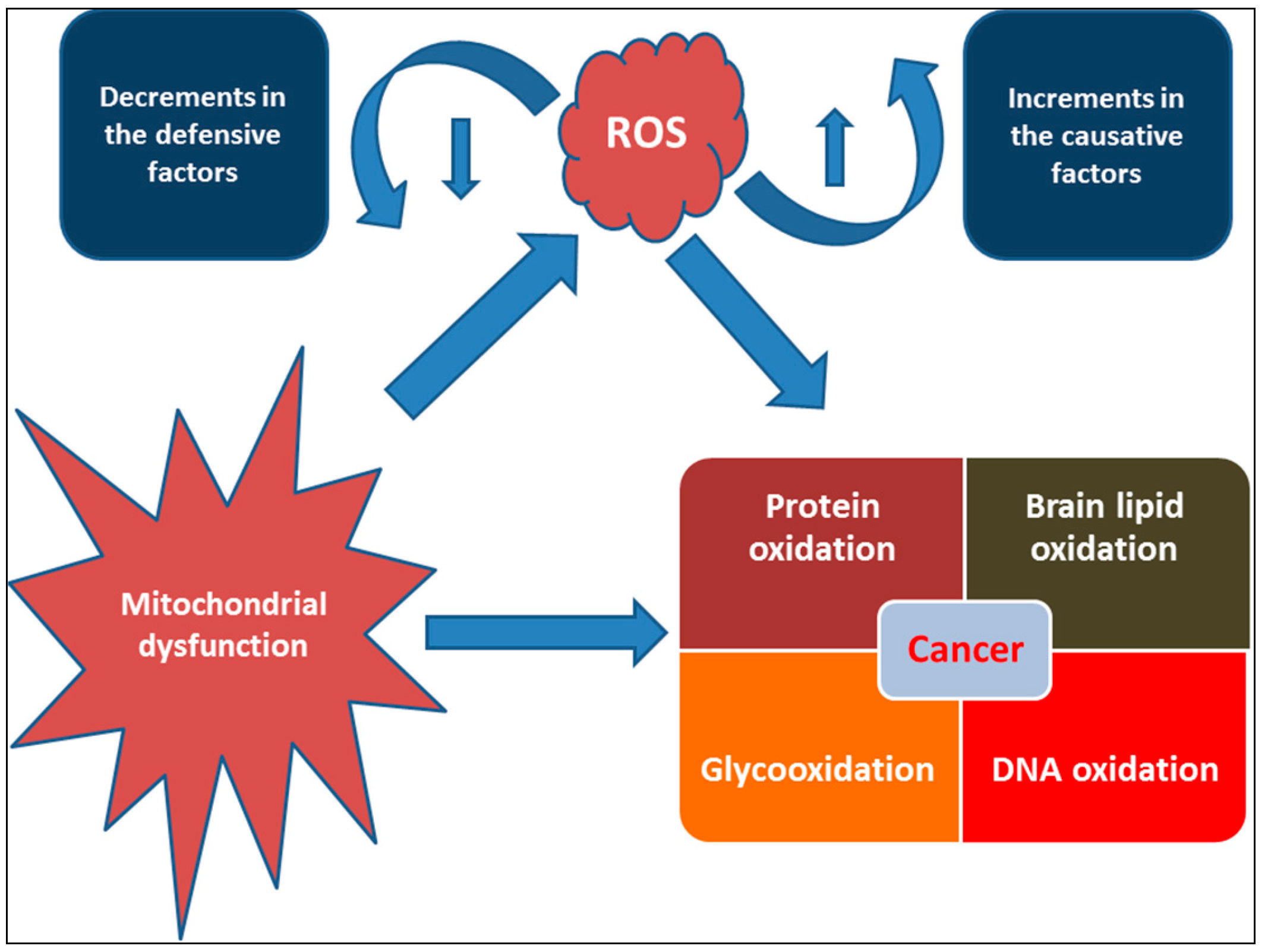
3. Oxidative Stress and Antioxidants: Halophytes Perspective
4. Roles of Phenolics and Flavonoids in Cancers Preventions and Treatments
5. Traditional Uses of Different Halophytes in Cancers and Cancer-Related Symptoms
6. Evaluating Anti-Cancer Activities of Halophytic Plants Extracts
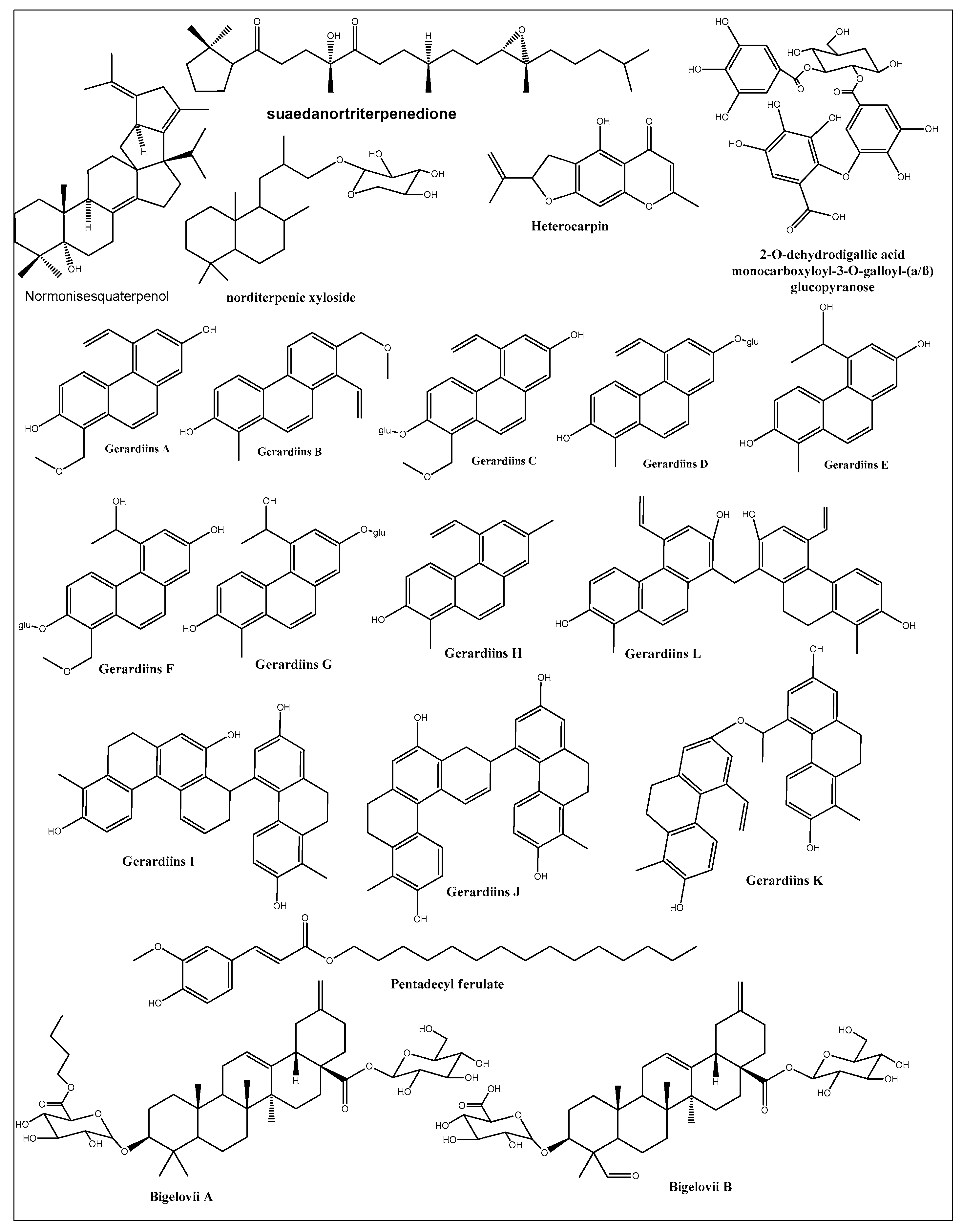
7. Isolated-Purified Anti-Cancer Agents from Different Halophytes
8. New Anticancer Agents from Halophytes
9. Immune System, Immunity, and Cancer Immunotherapy: The Flavonoids and Polyphenols Perspective
10. Molecular Basis of Immunology, and Major Cancer Immunotherapy Mechanisms: The Flavonoids and Polyphenols Stand-Point
11. Summary and Future Prospects
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Geiger, T.R.; Peeper, D.S. Metastasis Mechanisms. Biochim. Biophys. Acta-Rev. Cancer 2009, 1796, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, R.A. How Cancer Arises. Sci. Am. 1996, 275, 62–70. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. What Is Cancer? Available online: https://www.cancer.org/content/dam/CRC/PDF/Public/6041.00.pdf (accessed on 1 January 2023).
- Thrift-Perry, M.; Cabanes, A.; Cardoso, F.; Hunt, K.M.; Cruz, T.A.; Faircloth, K. Global Analysis of Metastatic Breast Cancer Policy Gaps and Advocacy Efforts across the Patient Journey. Breast 2018, 41, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Kunnumakara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer Is a Preventable Disease That Requires Major Lifestyle Changes. Pharm. Res. 2008, 25, 2097–2116. [Google Scholar] [CrossRef]
- World Health Organization. National Cancer Control Programmes (NCCP). 2022. Available online: https://apps.who.int/iris/handle/10665/42494 (accessed on 1 January 2023).
- Street, W. Cancer Facts & Figures 2019; American Cancer Socicety: Atlanta, GA, USA, 2019. [Google Scholar]
- Wu, S.; Zhu, W.; Thompson, P.; Hannun, Y.A. Evaluating Intrinsic and Non-Intrinsic Cancer Risk Factors. Nat. Commun. 2018, 9, 3490. [Google Scholar] [CrossRef] [Green Version]
- Dhahri, M.; Alghrably, M.; Mohammed, H.A.; Badshah, S.L.; Noreen, N.; Mouffouk, F.; Rayyan, S.; Qureshi, K.A.; Mahmood, D.; Lachowicz, J.I. Natural Polysaccharides as Preventive and Therapeutic Horizon for Neurodegenerative Diseases. Pharmaceutics 2021, 14, 1. [Google Scholar] [CrossRef]
- O’Rourke, A.; Kremb, S.; Duggan, B.M.; Sioud, S.; Kharbatia, N.; Raji, M.; Emwas, A.-H.; Gerwick, W.H.; Voolstra, C.R. Identification of a 3-Alkylpyridinium Compound from the Red Sea Sponge Amphimedon Chloros with in Vitro Inhibitory Activity against the West Nile Virus NS3 Protease. Molecules 2018, 23, 1472. [Google Scholar] [CrossRef] [Green Version]
- Katz, L.; Baltz, R.H. Natural Product Discovery: Past, Present, and Future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef]
- Uzoigwe, J.; Sauter, E.R. Cancer Prevention and Treatment Using Combination Therapy with Plant-and Animal-Derived Compounds. Expert Rev. Clin. Pharmacol. 2012, 5, 701–709. [Google Scholar] [CrossRef]
- Anwar, S.; Almatroudi, A.; Alsahli, M.A.; Khan, M.A.; Khan, A.A.; Rahmani, A.H. Natural Products: Implication in Cancer Prevention and Treatment through Modulating Various Biological Activities. Anti-Cancer Agents Med. Chem. 2020, 20, 2025–2040. [Google Scholar] [CrossRef] [PubMed]
- Nobili, S.; Lippi, D.; Witort, E.; Donnini, M.; Bausi, L.; Mini, E.; Capaccioli, S. Natural Compounds for Cancer Treatment and Prevention. Pharmacol. Res. 2009, 59, 365–378. [Google Scholar] [CrossRef]
- Vlaykova, T.; Dimitrova, I.; Pavlov, I.; Tacheva, T. Cancer Prevention—Dietary Anticarcinogens. Medicine 2013, 3, 381–392. [Google Scholar]
- Nirmala, M.J.; Samundeeswari, A.; Sankar, P.D. Natural Plant Resources in Anti-Cancer Therapy-A Review. Res. Plant Biol. 2011, 1, 1–14. [Google Scholar]
- Cragg, G.M.; Newman, D.J. Plants as a Source of Anti-Cancer Agents. J. Ethnopharmacol. 2005, 100, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, H.A.; Abdulkarim, A.K.; Alamami, A.D.; Elshibani, F.A. Phytochemical Constituents and Biological Activities of Jasonia Glutinosa L. : The First Report for the Plant Growing in North Africa. J. Chem. 2022, 2022, 1–11. [Google Scholar]
- Wangkheirakpam, S. Traditional and Folk Medicine as a Target for Drug Discovery. In Natural Products and Drug Discovery; Elsevier: Amsterdam, The Netherlands, 2018; pp. 29–56. [Google Scholar]
- Mann, J. Natural Products in Cancer Chemotherapy: Past, Present and Future. Nat. Rev. Cancer 2002, 2, 143–148. [Google Scholar] [CrossRef]
- Srivastava, V.; Negi, A.S.; Kumar, J.K.; Gupta, M.M.; Khanuja, S.P.S. Plant-Based Anticancer Molecules: A Chemical and Biological Profile of Some Important Leads. Bioorg. Med. Chem. 2005, 13, 5892–5908. [Google Scholar] [CrossRef]
- Pezzuto, J.M. Plant-Derived Anticancer Agents. Biochem. Pharmacol. 1997, 53, 121–133. [Google Scholar] [CrossRef]
- Wang, R.; Chen, H.; Yan, W.; Zheng, M.; Zhang, T.; Zhang, Y. Ferrocene-Containing Hybrids as Potential Anticancer Agents: Current Developments, Mechanisms of Action and Structure-Activity Relationships. Eur. J. Med. Chem. 2020, 190, 112109. [Google Scholar] [CrossRef]
- Rasool, I.; Ahmad, M.; Khan, Z.A.; Mansha, A.; Maqbool, T.; Zahoor, A.F.; Aslam, S. Recent Advancements in Oxadiazole-Based Anticancer Agents. Trop. J. Pharm. Res. 2017, 16, 723–733. [Google Scholar] [CrossRef] [Green Version]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS Homeostasis in Halophytes in the Context of Salinity Stress Tolerance. J. Exp. Bot. 2014, 65, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Ben Hamed, K.; Ellouzi, H.; Talbi, O.Z.; Hessini, K.; Slama, I.; Ghnaya, T.; Bosch, S.M.; Savouré, A.; Abdelly, C. Physiological Response of Halophytes to Multiple Stresses. Funct. Plant Biol. 2013, 40, 883–896. [Google Scholar] [CrossRef]
- Mishra, A.; Tanna, B. Halophytes: Potential Resources for Salt Stress Tolerance Genes and Promoters. Front. Plant Sci. 2017, 8, 829. [Google Scholar] [CrossRef]
- Shabala, S. Learning from Halophytes: Physiological Basis and Strategies to Improve Abiotic Stress Tolerance in Crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- He, M.; He, C.-Q.; Ding, N.-Z. Abiotic Stresses: General Defenses of Land Plants and Chances for Engineering Multistress Tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef] [Green Version]
- Waqas, M.A.; Kaya, C.; Riaz, A.; Farooq, M.; Nawaz, I.; Wilkes, A.; Li, Y. Potential Mechanisms of Abiotic Stress Tolerance in Crop Plants Induced by Thiourea. Front. Plant Sci. 2019, 10, 1336. [Google Scholar] [CrossRef]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Daur, I.; Nawaz, M.A.; Ahmad, M.Q.; Rana, I.A.; Atif, R.M.; Yang, S.H.; Chung, G. Redox and Ionic Homeostasis Regulations against Oxidative, Salinity and Drought Stress in Wheat (a Systems Biology Approach). Front. Genet. 2017, 8, 141. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Mech. Salin. Toler. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Borges, C.V.; Minatel, I.O.; Gomez-Gomez, H.A.; Lima, G.P.P. Medicinal Plants: Influence of Environmental Factors on the Content of Secondary Metabolites. In Medicinal Plants and Environmental Challenges; Springer: Berlin/Heidelberg, Germany, 2017; pp. 259–277. [Google Scholar]
- Dresselhaus, T.; Hückelhoven, R. Biotic and Abiotic Stress Responses in Crop Plants. Agronomy 2018, 8, 267. [Google Scholar] [CrossRef] [Green Version]
- Isah, T. Stress and Defense Responses in Plant Secondary Metabolites Production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Environmental Stress and Secondary Metabolites in Plants: An Overview. Plant Metab. Regul. Environ. Stress 2018, 153–167. [Google Scholar] [CrossRef]
- Stanković, M.; Jakovljević, D.; Stojadinov, M.; Stevanović, Z.D. Halophyte Species as a Source of Secondary Metabolites with Antioxidant Activity. In Ecophysiology, Abiotic Stress Responses and Utilization of Halophytes; Springer: Berlin/Heidelberg, Germany, 2019; pp. 289–312. [Google Scholar]
- Lopes, M.; Sanches-Silva, A.; Castilho, M.; Cavaleiro, C.; Ramos, F. Halophytes as Source of Bioactive Phenolic Compounds and Their Potential Applications. Crit. Rev. Food Sci. Nutr. 2021, 63, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Mackay, A. Ion Transport in Halophytes. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2011; Volume 57, pp. 151–199. ISBN 0065-2296. [Google Scholar]
- Flowers, T.J.; Troke, P.F.; Yeo, A.R. The Mechanism of Salt Tolerance in Halophytes. Annu. Rev. Plant Physiol. 1977, 28, 89–121. [Google Scholar] [CrossRef]
- Manousaki, E.; Kalogerakis, N. Halophytes—An Emerging Trend in Phytoremediation. Int. J. Phytoremediation 2011, 13, 959–969. [Google Scholar] [CrossRef]
- O’Leary, J.W.; Glenn, E.P. Global Distribution and Potential for Halophytes. In Halophytes as a Resource for Livestock and for Rehabilitation of Degraded Lands; Springer: Berlin/Heidelberg, Germany, 1994; pp. 7–17. [Google Scholar]
- Al-Azzawi, M.J.; Flowers, T.J. Distribution and Potential Uses of Halophytes within the Gulf Cooperation Council States. Agronomy 2022, 12, 1030. [Google Scholar] [CrossRef]
- Batanouny, K.H. Halophytes and Halophytic Plant Communities in the Arab Region. In Halophytes as a Resource for Livestock and for Rehabilitation of Degraded Lands; Springer: Berlin/Heidelberg, Germany, 1994; pp. 139–163. [Google Scholar]
- Khan, M.A.; Qaiser, M. Halophytes of Pakistan: Characteristics, Distribution and Potential Economic Usages. In Sabkha Ecosystems; Springer: Berlin/Heidelberg, Germany, 2006; pp. 129–153. [Google Scholar]
- Waisel, Y. Biology of Halophytes; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 0323151582. [Google Scholar]
- Flowers, T.J.; Muscolo, A. Introduction to the Special Issue: Halophytes in a Changing World. AoB Plants 2015, 7, plv020. [Google Scholar] [CrossRef]
- Faustino, M.V.; Faustino, M.A.F.; Pinto, D.C.G.A. Halophytic Grasses, a New Source of Nutraceuticals? A Review on Their Secondary Metabolites and Biological Activities. Int. J. Mol. Sci. 2019, 20, 1067. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Tang, X.; Shao, H.; Wang, H. Salinity Tolerance Mechanism of Economic Halophytes from Physiological to Molecular Hierarchy for Improving Food Quality. Curr. Genom. 2016, 17, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hozzein, W.N.; Hassan, H.M. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants 2020, 9, 869. [Google Scholar] [CrossRef]
- Jithesh, M.N.; Prashanth, S.R.; Sivaprakash, K.R.; Parida, A.K. Antioxidative Response Mechanisms in Halophytes: Their Role in Stress Defence. J. Genet. 2006, 85, 237. [Google Scholar] [CrossRef] [PubMed]
- Duarte, B.; Santos, D.; Marques, J.C.; Caçador, I. Ecophysiological Adaptations of Two Halophytes to Salt Stress: Photosynthesis, PS II Photochemistry and Anti-Oxidant Feedback–Implications for Resilience in Climate Change. Plant Physiol. Biochem. 2013, 67, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.-H.; Wang, B.-S. Oxidative Stress and Salt Tolerance in Plants. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2008; pp. 231–245. [Google Scholar]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity Effects on Polyphenol Content and Antioxidant Activities in Leaves of the Halophyte Cakile Maritima. Plant Physiol. Biochem. 2007, 45, 244–249. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Falleh, H.; Trabelsi, N.; Boulaaba, M.; Smaoui, A.; Abdelly, C. Influence of Biological, Environmental and Technical Factors on Phenolic Content and Antioxidant Activities of Tunisian Halophytes. Comptes Rendus Biol. 2008, 331, 865–873. [Google Scholar] [CrossRef]
- Ozgur, R.; Uzilday, B.; Sekmen, A.H.; Turkan, I. Reactive Oxygen Species Regulation and Antioxidant Defence in Halophytes. Funct. Plant Biol. 2013, 40, 832–847. [Google Scholar] [CrossRef] [PubMed]
- Abideen, Z.; Qasim, M.; Rasheed, A.; Adnan, M.Y.; Gul, B.; Khan, M.A. Antioxidant Activity and Polyphenolic Content of Phragmites Karka under Saline Conditions. Pak. J. Bot. 2015, 47, 813–818. [Google Scholar]
- Boughalleb, F.; Denden, M. Physiological and Biochemical Changes of Two Halophytes, Nitraria retusa (Forssk.) and Atriplex halimus (L.) under Increasing Salinity. Agric. J. 2011, 6, 327–339. [Google Scholar] [CrossRef]
- Ben Taârit, M.; Msaada, K.; Hosni, K.; Marzouk, B. Physiological Changes, Phenolic Content and Antioxidant Activity of Salvia officinalis L. Grown under Saline Conditions. J. Sci. Food Agric. 2012, 92, 1614–1619. [Google Scholar] [CrossRef]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal Halophytes: Potent Source of Health Promoting Biomolecules with Medical, Nutraceutical and Food Applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef]
- Bayoumi, M.T.; El Shaer, H.M. Impact of Halophytes on Animal Health and Nutrition. In Halophytes as a Resource for Livestock and for Rehabilitation of Degraded Lands; Springer: Berlin/Heidelberg, Germany, 1994; pp. 267–272. [Google Scholar]
- Cybulska, I.; Brudecki, G.; Alassali, A.; Thomsen, M.; Brown, J.J. Phytochemical Composition of Some Common Coastal Halophytes of the United Arab Emirates. Emir. J. Food Agric. 2014, 26, 1046–1057. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Elansary, H.O.; Mattar, M.A.; M Elhindi, K.; A Alotaibi, M.; Mishra, A. Differential Accumulation of Metabolites in Suaeda Species Provides New Insights into Abiotic Stress Tolerance in C4-Halophytic Species in Elevated CO2 Conditions. Agronomy 2021, 11, 131. [Google Scholar] [CrossRef]
- Boutaoui, N.; Zaiter, L.; Benayache, F.; Benayache, S.; Cacciagrano, F.; Cesa, S.; Secci, D.; Carradori, S.; Giusti, A.M.; Campestre, C. Atriplex Mollis Desf. Aerial Parts: Extraction Procedures, Secondary Metabolites and Color Analysis. Molecules 2018, 23, 1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qasim, M.; Gulzar, S.; Shinwari, Z.K.; Aziz, I.; Khan, M.A. Traditional Ethnobotanical Uses of Halophytes from Hub, Balochistan. Pak. J. Bot 2010, 42, 1543–1551. [Google Scholar]
- Qasim, M.; Gulzar, S.; Khan, M.A. Halophytes as Medicinal Plants. In Urbanisation, Land Use, Land Degradation and Environment; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Al-Omar, M.S.; Mohammed, H.A.; Mohammed, S.A.A.; Abd-Elmoniem, E.; Kandil, Y.I.; Eldeeb, H.M.; Chigurupati, S.; Sulaiman, G.M.; Al-Khurayyif, H.K.; Almansour, B.S. Anti-Microbial, Anti-Oxidant, and α-Amylase Inhibitory Activity of Traditionally-Used Medicinal Herbs: A Comparative Analyses of Pharmacology, and Phytoconstituents of Regional Halophytic Plants’ Diaspora. Molecules 2020, 25, 5457. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Ali, H.M.; Qureshi, K.A.; Alsharidah, M.; Kandil, Y.I.; Said, R.; Mohammed, S.A.A.; Al-Omar, M.S.; Al Rugaie, O.; Abdellatif, A.A.H. Comparative Phytochemical Profile and Biological Activity of Four Major Medicinal Halophytes from Qassim Flora. Plants 2021, 10, 2208. [Google Scholar] [CrossRef] [PubMed]
- Giordano, R.; Saii, Z.; Fredsgaard, M.; Hulkko, L.S.S.; Poulsen, T.B.G.; Thomsen, M.E.; Henneberg, N.; Zucolotto, S.M.; Arendt-Nielsen, L.; Papenbrock, J. Pharmacological Insights into Halophyte Bioactive Extract Action on Anti-Inflammatory, Pain Relief and Antibiotics-Type Mechanisms. Molecules 2021, 26, 3140. [Google Scholar] [CrossRef]
- Qasim, M.; Abideen, Z.; Adnan, M.Y.; Ansari, R.; Gul, B.; Khan, M.A. Traditional Ethnobotanical Uses of Medicinal Plants from Coastal Areas. J. Coast Life Med. 2014, 2, 22–30. [Google Scholar]
- Medini, F.; Ksouri, R. Antimicrobial Capacities of the Medicinal Halophyte Plants. In Natural Antimicrobial Agents; Springer: Berlin/Heidelberg, Germany, 2018; pp. 271–288. [Google Scholar]
- Medini, F.; Legault, J.; Pichette, A.; Abdelly, C.; Ksouri, R. Antiviral Efficacy of Limonium Densiflorum against HSV-1 and Influenza Viruses. South Afr. J. Bot. 2014, 92, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, Antioxidants, and the Degenerative Diseases of Aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, H. Natural and Synthetic Flavonoid Derivatives with Potential Antioxidant and Anticancer Activities; University of Saarland: Saarbrücken, Germany, 2009. [Google Scholar]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive Oxygen Species and Neutrophil Function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Day, B.J. The Science of Licking Your Wounds: Function of Oxidants in the Innate Immune System. Biochem. Pharmacol. 2019, 163, 451–457. [Google Scholar] [CrossRef]
- Irshad, M.; Chaudhuri, P.S. Oxidant-Antioxidant System: Role and Significance in Human Body. Indian J. Exp. Biol. 2002, 40, 1233–1239. [Google Scholar]
- Castellani, R.; Hirai, K.; Aliev, G.; Drew, K.L.; Nunomura, A.; Takeda, A.; Cash, A.D.; Obrenovich, M.E.; Perry, G.; Smith, M.A. Role of Mitochondrial Dysfunction in Alzheimer’s Disease. J. Neurosci. Res. 2002, 70, 357–360. [Google Scholar] [CrossRef]
- Hsu, C.-C.; Tseng, L.-M.; Lee, H.-C. Role of Mitochondrial Dysfunction in Cancer Progression. Exp. Biol. Med. 2016, 241, 1281–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elejalde, J.I.G. Oxidative Stress, Diseases and Antioxidant Treatment. An. Med. Interna 2001, 18, 326–335. [Google Scholar]
- Jezierska-Drutel, A.; Rosenzweig, S.A.; Neumann, C.A. Role of Oxidative Stress and the Microenvironment in Breast Cancer Development and Progression. In Advances in Cancer Research; Elsevier: Amsterdam, The Netherlands, 2013; Volume 119, pp. 107–125. ISBN 0065-230X. [Google Scholar]
- Visconti, R.; Grieco, D. New Insights on Oxidative Stress in Cancer. Curr. Opin. Drug Discov. Dev. 2009, 12, 240–245. [Google Scholar]
- Kaur, C.; Kapoor, H.C. Antioxidants in Fruits and Vegetables—The Millennium’s Health. Int. J. Food Sci. Technol. 2001, 36, 703–725. [Google Scholar] [CrossRef]
- Asadi-Samani, M.; Farkhad, N.K.; Mahmoudian-Sani, M.R.; Shirzad, H. Antioxidants as a Double-Edged Sword in the Treatment of Cancer. In Antioxidants; IntechOpen: London, UK, 2019. [Google Scholar]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Ren, Z.; Zhang, J.; Chuang, C.-C.; Kandaswamy, E.; Zhou, T.; Zuo, L. Role of ROS and Nutritional Antioxidants in Human Diseases. Front. Physiol. 2018, 9, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; Leonart, M.E. Oxidative Stress and Cancer: An Overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef]
- Sytar, O.; Mbarki, S.; Zivcak, M.; Brestic, M. The Involvement of Different Secondary Metabolites in Salinity Tolerance of Crops. In Salinity Responses and Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2018; Volume 2, pp. 21–48. [Google Scholar]
- Huang, W.-Y.; Cai, Y.-Z.; Zhang, Y. Natural Phenolic Compounds from Medicinal Herbs and Dietary Plants: Potential Use for Cancer Prevention. Nutr. Cancer 2009, 62, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, T.; Akpinar-Bayizit, A.; Yilmaz-Ersan, L.; Delikanli, B. Phenolics in Human Health. Int. J. Chem. Eng. Appl. 2014, 5, 393. [Google Scholar] [CrossRef] [Green Version]
- Guengerich, F.P. Metabolic Activation of Carcinogens. Pharmacol. Ther. 1992, 54, 17–61. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, S.A.; Wahab, S.; Abullais, S.S.; Das, G.; Hani, U.; Ahmad, W.; Amir, M.; Ahmad, A.; Kandasamy, G.; Vasudevan, R. Pharmacological Efficacy of Tamarix Aphylla: A Comprehensive Review. Plants 2021, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Halophytic Herbs of the Mediterranean Basin: An Alternative Approach to Health. Food Chem. Toxicol. 2018, 114, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Mohammed, H.A. The Valuable Impacts of Halophytic Genus Suaeda Mathsemicolon Nutritional, Chemical and Biological Values. Med. Chem. 2020, 16, 1044–1057. [Google Scholar] [CrossRef]
- Davoodvandi, A.; Shabani Varkani, M.; Clark, C.C.T.; Jafarnejad, S. Quercetin as an Anticancer Agent: Focus on Esophageal Cancer. J. Food Biochem. 2020, 44, e13374. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Khan, I.A.; ur-Rehman, M.; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer Potential of Quercetin: A Comprehensive Review. Phyther. Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef] [PubMed]
- Teğin, İ.; Canpolat, G.; Fidan, M. The Antioxidant Capacity, Total Phenolic Content and Phenolic Compounds of Plantago coronopus L. Subsp. Coronopus in Naturally Distributed in Akdoğmuş-Siirt. In Proceedings of the 2018 2nd International Symposium on Multidisciplinary Studies and Innovative Technologies (ISMSIT), Ankara, Turkey, 19–21 October 2018; IEEE: Piscataway, NJ, USA, 2018; pp. 1–4. [Google Scholar]
- Baysal, I.; Ekizoglu, M.; Ertas, A.; Temiz, B.; Agalar, H.G.; Yabanoglu-Ciftci, S.; Temel, H.; Ucar, G.; Turkmenoglu, F.P. Identification of Phenolic Compounds by LC-MS/MS and Evaluation of Bioactive Properties of Two Edible Halophytes: Limonium effusum and L. sinuatum. Molecules 2021, 26, 4040. [Google Scholar] [CrossRef] [PubMed]
- Murshid, S.S.A.; Atoum, D.; Abou-Hussein, D.R.; Abdallah, H.M.; Hareeri, R.H.; Almukadi, H.; Edrada-Ebel, R. Genus Salsola: Chemistry, Biological Activities and Future Prospective—A Review. Plants 2022, 11, 714. [Google Scholar] [CrossRef] [PubMed]
- Manthey, J.A.; Guthrie, N. Antiproliferative Activities of Citrus Flavonoids against Six Human Cancer Cell Lines. J. Agric. Food Chem. 2002, 50, 5837–5843. [Google Scholar] [CrossRef] [PubMed]
- Manthey, J.A.; Guthrie, N.; Grohmann, K. Biological Properties of Citrus Flavonoids Pertaining to Cancer and Inflammation. Curr. Med. Chem. 2001, 8, 135–153. [Google Scholar] [CrossRef]
- Cazarolli, L.H.; Zanatta, L.; Alberton, E.H.; Bonorino Figueiredo, M.S.R.; Folador, P.; Damazio, R.G.; Pizzolatti, M.G.; Barreto Silva, F.R.M. Flavonoids: Prospective Drug Candidates. Mini Rev. Med. Chem. 2008, 8, 1429–1440. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Ba, L.A.; Burkholz, T.; Schumann, E.; Diesel, B.; Zapp, J.; Kiemer, A.K.; Ries, C.; Hartmann, R.W.; Hosny, M. Facile Synthesis of Chrysin-Derivatives with Promising Activities as Aromatase Inhibitors. Nat. Prod. Commun. 2011, 6, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Khoo, B.Y.; Chua, S.L.; Balaram, P. Apoptotic Effects of Chrysin in Human Cancer Cell Lines. Int. J. Mol. Sci. 2010, 11, 2188–2199. [Google Scholar] [CrossRef] [Green Version]
- Mani, R.; Natesan, V. Chrysin: Sources, Beneficial Pharmacological Activities, and Molecular Mechanism of Action. Phytochemistry 2018, 145, 187–196. [Google Scholar] [CrossRef]
- Lan, L.; Wang, Y.; Pan, Z.; Wang, B.; Yue, Z.; Jiang, Z.; Li, L.; Wang, C.; Tang, H. Rhamnetin Induces Apoptosis in Human Breast Cancer Cells via the MiR-34a/Notch-1 Signaling Pathway. Oncol. Lett. 2019, 17, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, D.; Jung, K.-H.; Zhang, H.; Nannapaneni, S.; Wang, X.; Amin, A.R.M.R.; Chen, Z.; Shin, D.M. Luteolin Nanoparticle in Chemoprevention: In Vitro and in Vivo Anticancer Activity. Cancer Prev. Res. 2014, 7, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M. Luteolin, a Flavonoid, as an Anticancer Agent: A Review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-Y.; Wang, C.-J.; Chen, N.-F.; Ho, W.-H.; Lu, F.-J.; Tseng, T.-H. Luteolin Enhances Paclitaxel-Induced Apoptosis in Human Breast Cancer MDA-MB-231 Cells by Blocking STAT3. Chem. Biol. Interact. 2014, 213, 60–68. [Google Scholar] [CrossRef]
- Rauf, A.; Shariati, M.A.; Imran, M.; Bashir, K.; Khan, S.A.; Mitra, S.; Bin Emran, T.; Badalova, K.; Uddin, M.; Mubarak, M.S. Comprehensive Review on Naringenin and Naringin Polyphenols as a Potent Anticancer Agent. Environ. Sci. Pollut. Res. 2022, 29, 31025–31041. [Google Scholar] [CrossRef]
- Pecoraro, C.; Faggion, B.; Balboni, B.; Carbone, D.; Peters, G.J.; Diana, P.; Assaraf, Y.G.; Giovannetti, E. GSK3β as a Novel Promising Target to Overcome Chemoresistance in Pancreatic Cancer. Drug Resist. Updat. 2021, 58, 100779. [Google Scholar] [CrossRef]
- Devi, K.P.; Rajavel, T.; Habtemariam, S.; Nabavi, S.F.; Nabavi, S.M. Molecular Mechanisms Underlying Anticancer Effects of Myricetin. Life Sci. 2015, 142, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, S.; Gholami, M.H.; Zabolian, A.; Saleki, H.; Farahani, M.V.; Hamzehlou, S.; Far, F.B.; Sharifzadeh, S.O.; Samarghandian, S.; Khan, H. Caffeic Acid and Its Derivatives as Potential Modulators of Oncogenic Molecular Pathways: New Hope in the Fight against Cancer. Pharmacol. Res. 2021, 171, 105759. [Google Scholar] [CrossRef]
- Kuo, Y.-Y.; Jim, W.-T.; Su, L.-C.; Chung, C.-J.; Lin, C.-Y.; Huo, C.; Tseng, J.-C.; Huang, S.-H.; Lai, C.-J.; Chen, B.-C. Caffeic Acid Phenethyl Ester Is a Potential Therapeutic Agent for Oral Cancer. Int. J. Mol. Sci. 2015, 16, 10748–10766. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef]
- Rosendahl, A.H.; Perks, C.M.; Zeng, L.; Markkula, A.; Simonsson, M.; Rose, C.; Ingvar, C.; Holly, J.M.P.; Jernström, H. Caffeine and Caffeic Acid Inhibit Growth and Modify Estrogen Receptor and Insulin-like Growth Factor I Receptor Levels in Human Breast Cancer. Clin. Cancer Res. 2015, 21, 1877–1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Jana, J.; Sengupta, P.; Jana, S.; Chatterjee, S. Myricetin Arrests Human Telomeric G-Quadruplex Structure: A New Mechanistic Approach as an Anticancer Agent. Mol. Biosyst. 2016, 12, 2506–2518. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhu, M.; Wang, L.; Yu, S. Anti-Tumor Effects and Associated Molecular Mechanisms of Myricetin. Biomed. Pharmacother. 2019, 120, 109506. [Google Scholar] [CrossRef]
- Yang, Y.; Wolfram, J.; Boom, K.; Fang, X.; Shen, H.; Ferrari, M. Hesperetin Impairs Glucose Uptake and Inhibits Proliferation of Breast Cancer Cells. Cell Biochem. Funct. 2013, 31, 374–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roohbakhsh, A.; Parhiz, H.; Soltani, F.; Rezaee, R.; Iranshahi, M. Molecular Mechanisms behind the Biological Effects of Hesperidin and Hesperetin for the Prevention of Cancer and Cardiovascular Diseases. Life Sci. 2015, 124, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in Cancer Therapy: Anti-Cancer Effects and Mechanisms of Action. Cell Biosci. 2017, 7, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, P.; Leung, Y.-K.; Tang, W.-Y.; Harwood, C.; Ho, S.-M. Apigenin Suppresses Cancer Cell Growth through ERβ. Neoplasia 2006, 8, 896–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Konishi, M.; Horinaka, M.; Yasuda, T.; Goda, A.E.; Taniguchi, H.; Yano, K.; Wakada, M.; Sakai, T. Kaempferol Sensitizes Colon Cancer Cells to TRAIL-Induced Apoptosis. Biochem. Biophys. Res. Commun. 2008, 375, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, Y.; An, Y.; Fang, G. The Mechanism of Anticancer Action and Potential Clinical Use of Kaempferol in the Treatment of Breast Cancer. Biomed. Pharmacother. 2019, 117, 109086. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Choi, K.-C. Anti-Cancer Effect and Underlying Mechanism(s) of Kaempferol, a Phytoestrogen, on the Regulation of Apoptosis in Diverse Cancer Cell Models. Toxicol. Res. 2013, 29, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.H.; Rajamanickam, V.; Nagarajan, S. Supplementation of P-Coumaric Acid Exhibits Chemopreventive Effect via Induction of Nrf2 in a Short-Term Preclinical Model of Colon Cancer. Eur. J. Cancer Prev. 2019, 28, 472–482. [Google Scholar] [CrossRef]
- Sharma, S.H.; Rajamanickam, V.; Nagarajan, S. Antiproliferative Effect of P-Coumaric Acid Targets UPR Activation by Downregulating Grp78 in Colon Cancer. Chem. Biol. Interact. 2018, 291, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.; Narayanankutty, A.; Nazeem, P.A.; Valsalan, R.; Babu, T.D.; Mathew, D. Plant Phenolics Ferulic Acid and P-Coumaric Acid Inhibit Colorectal Cancer Cell Proliferation through EGFR down-Regulation. Asian Pac. J. Cancer Prev. 2016, 17, 4019–4023. [Google Scholar] [PubMed]
- Jang, M.G.; Ko, H.C.; Kim, S.-J. Effects of P-Coumaric Acid on MicroRNA Expression Profiles in SNU-16 Human Gastric Cancer Cells. Genes Genom. 2020, 42, 817–825. [Google Scholar] [CrossRef]
- Eroğlu, C.; Seçme, M.; Bağcı, G.; Dodurga, Y. Assessment of the Anticancer Mechanism of Ferulic Acid via Cell Cycle and Apoptotic Pathways in Human Prostate Cancer Cell Lines. Tumor Biol. 2015, 36, 9437–9446. [Google Scholar] [CrossRef]
- Gao, J.; Yu, H.; Guo, W.; Kong, Y.; Li, Q.; Yang, S.; Zhang, Y.; Wang, Y. The Anticancer Effects of Ferulic Acid Is Associated with Induction of Cell Cycle Arrest and Autophagy in Cervical Cancer Cells. Cancer Cell Int. 2018, 18, 102. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.-F.; Shi, S.-L.; Liu, Q.-R.; Tang, J.; Song, J.; Liang, Y. Anticancer Effects of Ginsenoside Rg1, Cinnamic Acid and Tanshinone IIA in Osteosarcoma MG-63 Cells: Nuclear Matrix Downregulation and Cytoplasmic Trafficking of Nucleophosmin. Int. J. Biochem. Cell Biol. 2008, 40, 1918–1929. [Google Scholar] [CrossRef]
- Mir, B.A.; Rasool, S.; Rehman, M.U.; Amin, I.; Ali, R. Anticancer Mechanistic Insights of Epigallocatechin-3-Gallate, an Active Ingredient of Green Tea (Camellia sinensis). Plant Hum. Health 2019, 2, 139–157. [Google Scholar]
- Hayakawa, S.; Ohishi, T.; Miyoshi, N.; Oishi, Y.; Nakamura, Y.; Isemura, M. Anti-Cancer Effects of Green Tea Epigallocatchin-3-Gallate and Coffee Chlorogenic Acid. Molecules 2020, 25, 4553. [Google Scholar] [CrossRef]
- Anwar, S.; Shamsi, A.; Shahbaaz, M.; Queen, A.; Khan, P.; Hasan, G.M.; Islam, A.; Alajmi, M.F.; Hussain, A.; Ahmad, F. Rosmarinic Acid Exhibits Anticancer Effects via MARK4 Inhibition. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Swamy, M.K.; Sinniah, U.R.; Ghasemzadeh, A. Anticancer Potential of Rosmarinic Acid and Its Improved Production through Biotechnological Interventions and Functional Genomics. Appl. Microbiol. Biotechnol. 2018, 102, 7775–7793. [Google Scholar] [CrossRef] [PubMed]
- Jacobo-Herrera, N.J.; Jacobo-Herrera, F.E.; Zentella-Dehesa, A.; Andrade-Cetto, A.; Heinrich, M.; Pérez-Plasencia, C. Medicinal Plants Used in Mexican Traditional Medicine for the Treatment of Colorectal Cancer. J. Ethnopharmacol. 2016, 179, 391–402. [Google Scholar] [CrossRef]
- Graham, J.G.; Quinn, M.L.; Fabricant, D.S.; Farnsworth, N.R. Plants Used against Cancer–an Extension of the Work of Jonathan Hartwell. J. Ethnopharmacol. 2000, 73, 347–377. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Castro, A.J.; Villarreal, M.L.; Salazar-Olivo, L.A.; Gomez-Sanchez, M.; Dominguez, F.; Garcia-Carranca, A. Mexican Medicinal Plants Used for Cancer Treatment: Pharmacological, Phytochemical and Ethnobotanical Studies. J. Ethnopharmacol. 2011, 133, 945–972. [Google Scholar] [CrossRef] [PubMed]
- Arya, S.S.; Devi, S.; Ram, K.; Kumar, S.; Kumar, N.; Mann, A.; Kumar, A.; Chand, G. Halophytes: The Plants of Therapeutic Medicine. In Ecophysiology, Abiotic Stress Responses and Utilization of Halophytes; Springer: Berlin/Heidelberg, Germany, 2019; pp. 271–287. [Google Scholar]
- Hammiche, V.; Maiza, K. Traditional Medicine in Central Sahara: Pharmacopoeia of Tassili N’ajjer. J. Ethnopharmacol. 2006, 105, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Bozyel, M.E.; Bozyel, E.M.; Canli, K.; Altuner, E.M. Anticancer Uses of Medicinal Plants in Turkish Traditional Medicine. J. Agric. Nat. 2019, 22, 465–484. [Google Scholar]
- Benarba, B. Use of Medicinal Plants by Breast Cancer Patients in Algeria. EXCLI J. 2015, 14, 1164. [Google Scholar]
- Stevanovic, Z.D.; Stankovic, M.S.; Stankovic, J.; Janackovic, P.; Stankovic, M. Use of Halophytes as Medicinal Plants: Phytochemical Diversity and Biological Activity. In Halophytes and Climate Change: Adaptive Mechanisms and Potential Uses; CABI: Wallingford, UK, 2019; p. 343. [Google Scholar]
- Seo, J.H.; Jin, M.H.; Chang, Y.H. Anti-Inflammatory Effect of Salsola Komarovii Extract with Dissociated Glucocorticoid Activity. BMC Complement. Med. Ther. 2020, 20, 176. [Google Scholar] [CrossRef] [PubMed]
- Saleh, K.A.; Albinhassan, T.H.; Al-Ghazzawi, A.M.; Mohaya, A.; Shati, A.A.; Ayoub, H.J.; Abdallah, Q.M. Anticancer Property of Hexane Extract of Suaeda Fruticose Plant Leaves against Different Cancer Cell Lines. Trop. J. Pharm. Res. 2020, 19, 129–136. [Google Scholar] [CrossRef]
- Albinhassan, T.H.; Saleh, K.A.; Barhoumi, Z.; Alshehri, M.A.; Al-Ghazzawi, A.M. Anticancer, Anti-Proliferative Activity of Avicennia Marina Plant Extracts. J. Cancer Res. Ther. 2021, 17, 879. [Google Scholar]
- Mohammed, H.A.; Al-Omar, M.S.; Mohammed, S.A.A.; Alhowail, A.H.; Eldeeb, H.M.; Sajid, M.S.M.; Abd-Elmoniem, E.M.; Alghulayqeh, O.A.; Kandil, Y.I.; Khan, R.A. Phytochemical Analysis, Pharmacological and Safety Evaluations of Halophytic Plant, Salsola cyclophylla. Molecules 2021, 26, 2384. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Khan, R.A.; Abdel-Hafez, A.A.; Abdel-Aziz, M.; Ahmed, E.; Enany, S.; Mahgoub, S.; Al-Rugaie, O.; Alsharidah, M.; Aly, M.S.A. Phytochemical Profiling, In Vitro and In Silico Anti-Microbial and Anti-Cancer Activity Evaluations and Staph GyraseB and h-TOP-IIβ Receptor-Docking Studies of Major Constituents of Zygophyllum coccineum L. Aqueous-Ethanolic Extract and Its Subsequent Fra. Molecules 2021, 26, 577. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, H.A.; Al-Omar, M.S.; Khan, R.A.; Mohammed, S.A.A.; Qureshi, K.A.; Abbas, M.M.; Al Rugaie, O.; Abd-Elmoniem, E.; Ahmad, A.M.; Kandil, Y.I. Chemical Profile, Antioxidant, Antimicrobial, and Anticancer Activities of the Water-Ethanol Extract of Pulicaria undulata Growing in the Oasis of Central Saudi Arabian Desert. Plants 2021, 10, 1811. [Google Scholar] [CrossRef] [PubMed]
- Ksouri, W.M.; Medini, F.; Mkadmini, K.; Legault, J.; Magné, C.; Abdelly, C.; Ksouri, R. LC–ESI-TOF–MS Identification of Bioactive Secondary Metabolites Involved in the Antioxidant, Anti-Inflammatory and Anticancer Activities of the Edible Halophyte Zygophyllum album Desf. Food Chem. 2013, 139, 1073–1080. [Google Scholar] [CrossRef]
- Dhahri, M.; Sioud, S.; Dridi, R.; Hassine, M.; Boughattas, N.A.; Almulhim, F.; Al Talla, Z.; Jaremko, M.; Emwas, A.-H.M. Extraction, Characterization, and Anticoagulant Activity of a Sulfated Polysaccharide from Bursatella Leachii Viscera. ACS Omega 2020, 5, 14786–14795. [Google Scholar] [CrossRef] [PubMed]
- Oueslati, S.; Ksouri, R.; Falleh, H.; Pichette, A.; Abdelly, C.; Legault, J. Phenolic Content, Antioxidant, Anti-Inflammatory and Anticancer Activities of the Edible Halophyte Suaeda Fruticosa Forssk. Food Chem. 2012, 132, 943–947. [Google Scholar] [CrossRef]
- Karker, M.; Falleh, H.; Msaada, K.; Smaoui, A.; Abdelly, C.; Legault, J.; Ksouri, R. Antioxidant, Anti-Inflammatory and Anticancer Activities of the Medicinal Halophyte Reaumuria Vermiculata. EXCLI J. 2016, 15, 297. [Google Scholar] [PubMed]
- Sun, Z.; Wang, H.; Wang, J.; Zhou, L.; Yang, P. Chemical Composition and Anti-Inflammatory, Cytotoxic and Antioxidant Activities of Essential Oil from Leaves of Mentha Piperita Grown in China. PLoS ONE 2014, 9, e114767. [Google Scholar] [CrossRef] [Green Version]
- Hamed, A.R.; El-Hawary, S.S.; Ibrahim, R.M.; Abdelmohsen, U.R.; El-Halawany, A.M. Identification of Chemopreventive Components from Halophytes Belonging to Aizoaceae and Cactaceae Through LC/MS—Bioassay Guided Approach. J. Chromatogr. Sci. 2021, 59, 618–626. [Google Scholar] [CrossRef]
- Sehrawat, A.; Sultana, S. Evaluation of Possible Mechanisms of Protective Role of Tamarix gallica against DEN Initiated and 2-AAF Promoted Hepatocarcinogenesis in Male Wistar Rats. Life Sci. 2006, 79, 1456–1465. [Google Scholar] [CrossRef]
- Gamal, G.; Abo-El-Seoud, K.A.; Attia, G. Triterpenoids from the Aerial Parts of Anabasis articulata (Forssk) Moq: Gastroprotective Effect in Vivo with in Silico Studies, Cytotoxic and Antimicrobial Activities. Nat. Prod. Res. 2021, 36, 4076–4084. [Google Scholar] [CrossRef]
- Boulaaba, M.; Mkadmini, K.; Tsolmon, S.; Han, J.; Smaoui, A.; Kawada, K.; Ksouri, R.; Isoda, H.; Abdelly, C. In Vitro Antiproliferative Effect of Arthrocnemum indicum Extracts on Caco-2 Cancer Cells through Cell Cycle Control and Related Phenol LC-TOF-MS Identification. Evid.-Based Complement. Altern. Med. 2013, 2013, 529375. [Google Scholar] [CrossRef] [Green Version]
- Sharawi, Z.W. Therapeutic Effect of Arthrocnemum machrostachyum Methanolic Extract on Ehrlich Solid Tumor in Mice. BMC Complement. Med. Ther. 2020, 20, 153. [Google Scholar] [CrossRef]
- Petkov, V.; Batsalova, T.; Stoyanov, P.; Mladenova, T.; Kolchakova, D.; Argirova, M.; Raycheva, T.; Dzhambazov, B. Selective Anticancer Properties, Proapoptotic and Antibacterial Potential of Three Asplenium Species. Plants 2021, 10, 1053. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.; Vasas, A.; Schelz, Z.S.; Viveiros, M.; Molnar, J.; Hohmann, J.; Amaral, L. Constituents of Carpobrotus edulis Inhibit P-Glycoprotein of MDR1-Transfected Mouse Lymphoma Cells. Anticancer Res. 2010, 30, 829–835. [Google Scholar] [PubMed]
- Lee, C.-W.; Chen, H.-J.; Xie, G.-R.; Shih, C.-K. Djulis (Chenopodium formosanum) Prevents Colon Carcinogenesis via Regulating Antioxidative and Apoptotic Pathways in Rats. Nutrients 2019, 11, 2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-W.; Chen, H.-J.; Chien, Y.-H.; Hsia, S.-M.; Chen, J.-H.; Shih, C.-K. Synbiotic Combination of Djulis (Chenopodium formosanum) and Lactobacillus acidophilus Inhibits Colon Carcinogenesis in Rats. Nutrients 2020, 12, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.A.; Ju, J. Antioxidant and Growth Inhibitory Activities of Mesembryanthemum crystallinum L. in HCT116 Human Colon Cancer Cells. J. Nutr. Health 2019, 52, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Fraternale, D.; Ricci, D.; Calcabrini, C.; Guescini, M.; Martinelli, C.; Sestili, P. Cytotoxic Activity of Essential Oils of Aerial Parts and Ripe Fruits of Echinophora spinosa (Apiaceae). Nat. Prod. Commun. 2013, 8, 1645–1649. [Google Scholar] [CrossRef] [Green Version]
- Boulaaba, M.; Kalai, F.Z.; Dakhlaoui, S.; Ezzine, Y.; Selmi, S.; Bourgou, S.; Smaoui, A.; Isoda, H.; Ksouri, R. Antioxidant, Antiproliferative and Anti-Inflammatory Effects of Glaucium flavum Fractions Enriched in Phenolic Compounds. Med. Chem. Res. 2019, 28, 1995–2001. [Google Scholar] [CrossRef]
- Bournine, L.; Bensalem, S.; Peixoto, P.; Gonzalez, A.; Maiza-Benabdesselam, F.; Bedjou, F.; Wauters, J.-N.; Tits, M.; Frederich, M.; Castronovo, V. Revealing the Anti-Tumoral Effect of Algerian Glaucium flavum Roots against Human Cancer Cells. Phytomedicine 2013, 20, 1211–1218. [Google Scholar] [CrossRef]
- Haji, A.F.; Ostad, S.N.; Khanavi, M.; Haji, A.A.; Farahanikia, B.; Salarytabar, A. Cytotoxicity of Two Species of Glaucium from Iran. J. Med. Plants 2013, 12, 85–92. [Google Scholar]
- Um, Y.R.; Kong, C.-S.; Im Lee, J.; Kim, Y.A.; Nam, T.J.; Seo, Y. Evaluation of Chemical Constituents from Glehnia littoralis for Antiproliferative Activity against HT-29 Human Colon Cancer Cells. Process Biochem. 2010, 45, 114–119. [Google Scholar] [CrossRef]
- Medini, F.; Bourgou, S.; Lalancette, K.; Snoussi, M.; Mkadmini, K.; Coté, I.; Abdelly, C.; Legault, J.; Ksouri, R. Phytochemical Analysis, Antioxidant, Anti-Inflammatory, and Anticancer Activities of the Halophyte Limonium densiflorum Extracts on Human Cell Lines and Murine Macrophages. South Afr. J. Bot. 2015, 99, 158–164. [Google Scholar] [CrossRef]
- Amrani, A.; Lahneche, A.M.; Benaissa, O.; Boubekri, N.; Demirtaş, I.; Benayache, F.; Benayache, S.; Zama, D. In Vitro Antiproliferative and Inhibition of Oxidative DNA Damage Activities of N-Butanol Extract of Limonium bonduelli from Algeria. Braz. Arch. Biol. Technol. 2019, 62. [Google Scholar] [CrossRef]
- Placines, C.; Castaneda-Loaiza, V.; Rodrigues, M.J.; Pereira, C.G.; da Silva, J.P.; Zengin, G.; Custódio, L. In Vitro Enzyme Inhibitory and Antioxidant Properties, Cytotoxicity, and LC-DAD-ESI-MS/MS Profile of Extracts from the Halophyte Lotus creticus L. Jundishapur J. Nat. Pharm. Prod. 2021, 16, e101125. [Google Scholar] [CrossRef]
- Castañeda-Loaiza, V.; Placines, C.; Rodrigues, M.J.; Pereira, C.G.; Zengin, G.; Neng, N.R.; Nogueira, J.M.F.; Custódio, L. In Vitro Enzyme Inhibitory and Anti-Oxidant Properties, Cytotoxicity and Chemical Composition of the Halophyte Malcolmia littorea (L.) R. Br. (Brassicaceae). Nat. Prod. Res. 2020, 35, 4753–4756. [Google Scholar] [CrossRef] [PubMed]
- Barnawi, I.O.; Ali, I. Anticancer Potential of Pulicaria Crispa Extract on Human Breast Cancer MDA-MB-231 Cells. Lett. Drug Des. Discov. 2019, 16, 1354–1359. [Google Scholar] [CrossRef]
- Nawwar, M.A.; Ayoub, N.A.; El-Rai, M.A.; Bassyouny, F.; Mostafa, E.S.; Al-Abd, A.M.; Harms, M.; Wende, K.; Lindequist, U.; Linscheid, M.W. Cytotoxic Ellagitannins from Reaumuria vermiculata. Fitoterapia 2012, 83, 1256–1266. [Google Scholar] [CrossRef]
- Ryu, D.-S.; Kim, S.-H.; Lee, D.-S. Anti-Proliferative Effect of Polysaccharides from Salicornia Herbacea on Induction of G2/M Arrest and Apoptosis in Human Colon Cancer Cells. J. Microbiol. Biotechnol. 2009, 19, 1482–1489. [Google Scholar] [CrossRef]
- Al Bratty, M.; Makeen, H.A.; Alhazmi, H.A.; Syame, S.M.; Abdalla, A.N.; Homeida, H.E.; Sultana, S.; Ahsan, W.; Khalid, A. Phytochemical, Cytotoxic, and Antimicrobial Evaluation of the Fruits of Miswak Plant, Salvadora persica L. J. Chem. 2020, 2020, 4521951. [Google Scholar] [CrossRef]
- Ibrahim, A.Y.; El-Gengaihi, S.E.; Motawe, H.M. Phytochemical and Cytotoxicity Investigations of Salvadora persica Bark Extracts. JASMR 2011, 6, 127–133. [Google Scholar]
- Saleem, H.; Khurshid, U.; Sarfraz, M.; Tousif, M.I.; Alamri, A.; Anwar, S.; Alamri, A.; Ahmad, I.; Abdallah, H.H.; Mahomoodally, F.M. A Comprehensive Phytochemical, Biological, Toxicological and Molecular Docking Evaluation of Suaeda fruticosa (L.) Forssk.: An Edible Halophyte Medicinal Plant. Food Chem. Toxicol. 2021, 154, 112348. [Google Scholar] [CrossRef]
- Boulaaba, M.; Tsolmon, S.; Ksouri, R.; Han, J.; Kawada, K.; Smaoui, A.; Abdelly, C.; Isoda, H. Anticancer Effect of Tamarix gallica Extracts on Human Colon Cancer Cells Involves Erk1/2 and P38 Action on G 2/M Cell Cycle Arrest. Cytotechnology 2013, 65, 927–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Attar, M.M.; Abdel-Tawab, F.M.; Awad, A.A.; Ahmad, E.S.; Kamel, H.A.; Hassan, A.I. Anti-Cancer Effects of Zygophyllum album and Suaeda palaestina Extracts on Human Liver Cancer Cell Lines. Egypt. J. Genet. Cytol. 2019, 48, 77–90. [Google Scholar]
- Nipun, T.S.; Khatib, A.; Ibrahim, Z.; Ahmed, Q.U.; Redzwan, I.E.; Saiman, M.Z.; Supandi, F.; Primaharinastiti, R.; El-Seedi, H.R. Characterization of α-Glucosidase Inhibitors from Psychotria malayana Jack Leaves Extract Using LC-MS-Based Multivariate Data Analysis and In-Silico Molecular Docking. Molecules 2020, 25, 5885. [Google Scholar] [CrossRef]
- Noumi, E.; Snoussi, M.; Anouar, E.H.; Alreshidi, M.; Veettil, V.N.; Elkahoui, S.; Adnan, M.; Patel, M.; Kadri, A.; Aouadi, K. HR-LCMS-Based Metabolite Profiling, Antioxidant, and Anticancer Properties of Teucrium polium L. Methanolic Extract: Computational and In Vitro Study. Antioxidants 2020, 9, 1089. [Google Scholar] [CrossRef]
- Safitri, A.; Putri, A.S.; Octavianty, T.D.; Sari, D.R.T. Metabolomic Profiles of Curcuma Longa L and Cosmos Caudatus Extracts and Their In-Silico Anti-Cancer Activity. J. Phys. Conf. Ser. 2020, 1665, 12022. [Google Scholar] [CrossRef]
- Sahu, R.; Kar, R.K.; Sunita, P.; Bose, P.; Kumari, P.; Bharti, S.; Srivastava, S.; Pattanayak, S.P. LC-MS Characterized Methanolic Extract of Zanthoxylum armatum Possess Anti-Breast Cancer Activity through Nrf2-Keap1 Pathway: An In-Silico, In-Vitro and In-Vivo Evaluation. J. Ethnopharmacol. 2021, 269, 113758. [Google Scholar]
- Zengin, G.; Aumeeruddy-Elalfi, Z.; Mollica, A.; Yilmaz, M.A.; Mahomoodally, M.F. In Vitro and In Silico Perspectives on Biological and Phytochemical Profile of Three Halophyte Species—A Source of Innovative Phytopharmaceuticals from Nature. Phytomedicine 2018, 38, 35–44. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Almahmoud, S.A.; Arfeen, M.; Srivastava, A.; El-Readi, M.Z.; Ragab, E.A.; Shehata, S.M.; Mohammed, S.A.A.; Mostafa, E.M.; El-khawaga, H.A.; et al. Phytochemical Profiling, Molecular Docking, and In Vitro Anti-Hepatocellular Carcinoid Bioactivity of Suaeda Vermiculata Extracts. Arab. J. Chem. 2022, 15, 103950. [Google Scholar] [CrossRef]
- Farooq Khan, M.; Nasr, F.A.; Baabbad, A.A.; Alqahtani, A.S.; Wadaan, M.A.M. Investigating the Anticancer Activity and Characterization of Bioactive Constituents of Moricandia sinaica (Boiss.) Boiss through In Vitro and In Silico Approaches in Triple-Negative Breast Cancer Cell Line. Appl. Sci. 2021, 11, 1244. [Google Scholar] [CrossRef]
- Mohammed, H.; Al-Mahmoud, S.; Arfeen, M.; Srivastava, A.; Al-Raidy, M.Z.; Ragab, E.A.; Shehata, S.M.; Mohammed, S.A.A.; Mostafa, E.M.; El-Khawaga, H.A. LC-MS Profiled Chemical Constituents, Molecular Modeling, and In Vitro Bioactivity Evaluations of Suaeda vermiculata Extracts as Anti-Hepatocellular Carcinoma Preparation: Assessment of the Constituents’ Role, and Receptor Docking Feasibility Based Activity Projections. Arab. J. Chem. 2022, 15, 103950. [Google Scholar]
- Kim, Y.A.; Kim, H.; Seo, Y. Antiproliferative Effect of Flavonoids from the Halophyte Vitex rotundifolia on Human Cancer Cells. Nat. Prod. Commun. 2013, 8, 1405–1408. [Google Scholar] [CrossRef] [Green Version]
- Orabi, M.A.A.; Zidan, S.A.H.; Attia, G.H.; Alyami, H.S.; Matsunami, K.; Hatano, T. Ellagitannins and Simple Phenolics from the Halophytic Plant Tamarix nilotica. Nat. Prod. Res. 2020, 36, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Bournine, L.; Bensalem, S.; Wauters, J.-N.; Iguer-Ouada, M.; Maiza-Benabdesselam, F.; Bedjou, F.; Castronovo, V.; Bellahcène, A.; Tits, M.; Frédérich, M. Identification and Quantification of the Main Active Anticancer Alkaloids from the Root of Glaucium flavum. Int. J. Mol. Sci. 2013, 14, 23533–23544. [Google Scholar] [CrossRef]
- Rodrigues, M.J.; Gangadhar, K.N.; Vizetto-Duarte, C.; Wubshet, S.G.; Nyberg, N.T.; Barreira, L.; Varela, J.; Custódio, L. Maritime Halophyte Species from Southern Portugal as Sources of Bioactive Molecules. Mar. Drugs 2014, 12, 2228–2244. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.-S.; Kim, Y.A.; Kim, H.; Seo, Y. Evaluation of a Furochromone from the Halophyte Corydalis Heterocarpa for Cytotoxic Activity against Human Gastric Cancer (AGS) Cells. Food Funct. 2016, 7, 4823–4829. [Google Scholar] [CrossRef] [PubMed]
- Stefkó, D.; Kúsz, N.; Barta, A.; Kele, Z.; Bakacsy, L.; Szepesi, Á.; Fazakas, C.; Wilhelm, I.; Krizbai, I.A.; Hohmann, J. Gerardiins A–L and Structurally Related Phenanthrenes from the Halophyte Plant Juncus Gerardii and Their Cytotoxicity against Triple-Negative Breast Cancer Cells. J. Nat. Prod. 2020, 83, 3058–3068. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, M.; Zhao, Y.; Wang, H.; Liu, T.; Xin, Z. Pentadecyl Ferulate, a Potent Antioxidant and Antiproliferative Agent from the Halophyte Salicornia herbacea. Food Chem. 2013, 141, 2066–2074. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, X.; Shan, Y.; Guan, F.; Chen, Y.; Wang, X.; Wang, M.; Feng, X. Two New Nortriterpenoid Saponins from Salicornia bigelovii Torr. and Their Cytotoxic Activity. Fitoterapia 2012, 83, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Shan, Y.; Wang, Q.; Wang, M.; Chen, Y.; Yin, M.; Liu, F.; Zhao, Y.; Zhang, J.; Feng, X. Induction of Apoptosis by Bigelovii A through Inhibition of NF-κB Activity. Mol. Med. Rep. 2018, 18, 1600–1608. [Google Scholar] [CrossRef] [Green Version]
- Parvez, M.K.; Al-Dosari, M.S.; Rehman, M.T.; Alajmi, M.F.; Alqahtani, A.S.; AlSaid, M.S. New Terpenic and Phenolic Compounds from Suaeda Monoica Reverse Oxidative and Apoptotic Damages in Human Endothelial Cells. Saudi Pharm. J. 2021, 29, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Gupta, C.; Prakash, D. Phytochemicals as the Source of Natural Immunomodulator and Their Role in Cancer Chemoprevention. In Immunomodulators and Human Health; Springer: Berlin/Heidelberg, Germany, 2022; pp. 209–241. [Google Scholar]
- Masad, R.J.; Haneefa, S.M.; Mohamed, Y.A.; Al-Sbiei, A.; Bashir, G.; Fernandez-Cabezudo, M.J.; Al-Ramadi, B.K. The Immunomodulatory Effects of Honey and Associated Flavonoids in Cancer. Nutrients 2021, 13, 1269. [Google Scholar] [CrossRef] [PubMed]
- Oo, A.M.; Mohd Adnan, L.H.; Nor, N.M.; Simbak, N.; Ahmad, N.Z.; Lwin, O.M. Immunomodulatory Effects of Flavonoids: An Experimental Study on Natural-Killer-Cell-Mediated Cytotoxicity against Lung Cancer and Cytotoxic Granule Secretion Profile. Proc. Singapore Healthc. 2021, 30, 279–285. [Google Scholar] [CrossRef]
- Yilma, A.N.; Singh, S.R.; Morici, L.; Dennis, V.A. Flavonoid Naringenin: A Potential Immunomodulator for Chlamydia Trachomatis Inflammation. Mediat. Inflamm. 2013, 2013, 102457. [Google Scholar]
- Esmaeil, N.; Anaraki, S.B.; Gharagozloo, M.; Moayedi, B. Silymarin Impacts on Immune System as an Immunomodulator: One Key for Many Locks. Int. Immunopharmacol. 2017, 50, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Ramiro-Puig, E.; Castell, M. Cocoa: Antioxidant and Immunomodulator. Br. J. Nutr. 2009, 101, 931–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rachmawati, I.; Rifa’i, M. In Vitro Immunomodulatory Activity of Aqueous Extract of Moringa Oleifera Lam. Leaf to the CD4+, CD8+ and B220+ Cells in Mus Musculus. J. Exp. Life Sci. 2014, 4, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Zalizar, L. Flavonoids of Phylanthus Niruri as Immunomodulators a Prospect to Animal Disease Control. ARPN J. Sci. Technol. 2013, 3, 529–532. [Google Scholar]
- Uivarosi, V.; Munteanu, A. Flavonoid Complexes as Promising Anticancer Metallodrugs. In Flavonoids-From Biosynthesis to Human Health; IntechOpen: London, UK, 2017; pp. 305–337. [Google Scholar]
- Montbriand, M.J. Herbs or Natural Products That Decrease Cancer Growth: Part One of a Four-Part Series. Oncol. Nurs. Forum 1969, 31, E75–E90. [Google Scholar]
- Durga, M.; Nathiya, S.; Devasena, T. Immunomodulatory and Antioxidant Actions of Dietary Flavonoids. Int. J. Pharm. Pharm. Sci. 2014, 6, 50–56. [Google Scholar]
- González-Gallego, J.; García-Mediavilla, M.V.; Sánchez-Campos, S.; Tuñón, M.J. Anti-Inflammatory and Immunomodulatory Properties of Dietary Flavonoids. In Polyphenols in Human Health and Disease; Elsevier: Amsterdam, The Netherlands, 2014; pp. 435–452. [Google Scholar]
- Hosseinzade, A.; Sadeghi, O.; Naghdipour Biregani, A.; Soukhtehzari, S.; Brandt, G.S.; Esmaillzadeh, A. Immunomodulatory Effects of Flavonoids: Possible Induction of T CD4+ Regulatory Cells through Suppression of MTOR Pathway Signaling Activity. Front. Immunol. 2019, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Araki, K.; Ellebedy, A.H.; Ahmed, R. TOR in the Immune System. Curr. Opin. Cell Biol. 2011, 23, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Baranowski, M.; Enns, J.; Blewett, H.; Yakandawala, U.; Zahradka, P.; Taylor, C.G. Dietary Flaxseed Oil Reduces Adipocyte Size, Adipose Monocyte Chemoattractant Protein-1 Levels and T-Cell Infiltration in Obese, Insulin-Resistant Rats. Cytokine 2012, 59, 382–391. [Google Scholar] [CrossRef]
- Odegaard, J.I.; Chawla, A. The Immune System as a Sensor of the Metabolic State. Immunity 2013, 38, 644–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of Effector CD4 T Cell Populations. Annu. Rev. Immunol. 2009, 28, 445–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez-Agell, M.; Urpi-Sarda, M.; Sacanella, E.; Camino-López, S.; Chiva-Blanch, G.; Llorente-Cortés, V.; Tobias, E.; Roura, E.; Andres-Lacueva, C.; Lamuela-Raventos, R.M. Cocoa Consumption Reduces NF-ΚB Activation in Peripheral Blood Mononuclear Cells in Humans. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 257–263. [Google Scholar] [CrossRef]
- Mao, T.K. Van de, WJ, Keen, CL, Schmitz, HH & Gershwin, ME Modulation of TNF-Alpha Secretion in Peripheral Blood Mononuclear Cells by Cocoa Flavanols and Procyanidins. Dev. Immunol. 2002, 9, 135–141. [Google Scholar]
- Ramos-Romero, S.; Pérez-Cano, F.J.; Pérez-Berezo, T.; Castellote, C.; Franch, A.; Castell, M. Effect of a Cocoa Flavonoid-Enriched Diet on Experimental Autoimmune Arthritis. Br. J. Nutr. 2012, 107, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Seo, B.R.; Min, K.; Cho, I.J.; Kim, S.C.; Kwon, T.K. Curcumin Significantly Enhances Dual PI3K/Akt and MTOR Inhibitor NVP-BEZ235-Induced Apoptosis in Human Renal Carcinoma Caki Cells through down-Regulation of P53-Dependent Bcl-2 Expression and Inhibition of Mcl-1 Protein Stability. PLoS ONE 2014, 9, e95588. [Google Scholar] [CrossRef]
- Zhao, G.; Han, X.; Zheng, S.; Li, Z.; Sha, Y.; Ni, J.; Sun, Z.; Qiao, S.; Song, Z. Curcumin Induces Autophagy, Inhibits Proliferation and Invasion by Downregulating AKT/MTOR Signaling Pathway in Human Melanoma Cells. Oncol. Rep. 2016, 35, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Alanazi, I.O.; Khan, Z. Understanding EGFR Signaling in Breast Cancer and Breast Cancer Stem Cells: Overexpression and Therapeutic Implications. Asian Pac. J. Cancer Prev. 2016, 17, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Dienstmann, R.; Salazar, R.; Tabernero, J. Overcoming Resistance to Anti-EGFR Therapy in Colorectal Cancer. Am. Soc. Clin. Oncol. Educ. B. 2015, 35, e149–e156. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Afaq, F.; Syed, D.N.; Mukhtar, H. Fisetin, a Novel Dietary Flavonoid, Causes Apoptosis and Cell Cycle Arrest in Human Prostate Cancer LNCaP Cells. Carcinogenesis 2008, 29, 1049–1056. [Google Scholar] [CrossRef] [Green Version]
- Suh, Y.; Afaq, F.; Khan, N.; Johnson, J.J.; Khusro, F.H.; Mukhtar, H. Fisetin Induces Autophagic Cell Death through Suppression of MTOR Signaling Pathway in Prostate Cancer Cells. Carcinogenesis 2010, 31, 1424–1433. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.T.; Lee, S.H.; Kim, Y.M. Quercetin Regulates Sestrin 2-AMPK-MTOR Signaling Pathway and Induces Apoptosis via Increased Intracellular ROS in HCT116 Colon Cancer Cells. J. Cancer Prev. 2013, 18, 264. [Google Scholar] [CrossRef] [Green Version]
- Granato, M.; Rizzello, C.; Montani, M.S.G.; Cuomo, L.; Vitillo, M.; Santarelli, R.; Gonnella, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. Quercetin Induces Apoptosis and Autophagy in Primary Effusion Lymphoma Cells by Inhibiting PI3K/AKT/MTOR and STAT3 Signaling Pathways. J. Nutr. Biochem. 2017, 41, 124–136. [Google Scholar] [CrossRef]
- Granato, M.; Rizzello, C.; Romeo, M.A.; Yadav, S.; Santarelli, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. Concomitant Reduction of C-Myc Expression and PI3K/AKT/MTOR Signaling by Quercetin Induces a Strong Cytotoxic Effect against Burkitt’s Lymphoma. Int. J. Biochem. Cell Biol. 2016, 79, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, K.; Ghosh, S.; Mukherjee, A.; Sadhukhan, R.; Mondal, J.; Khuda-Bukhsh, A.R. Quercetin Induces Cytochrome-c Release and ROS Accumulation to Promote Apoptosis and Arrest the Cell Cycle in G2/M, in Cervical Carcinoma: Signal Cascade and Drug-DNA Interaction. Cell Prolif. 2013, 46, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, N.; Kim, H.; Talcott, S.; Mertens-Talcott, S. Pomegranate Polyphenolics Suppressed Azoxymethane-Induced Colorectal Aberrant Crypt Foci and Inflammation: Possible Role of MiR-126/VCAM-1 and MiR-126/PI3K/AKT/MTOR. Carcinogenesis 2013, 34, 2814–2822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezrich, J.D.; Fechner, J.H.; Zhang, X.; Johnson, B.P.; Burlingham, W.J.; Bradfield, C.A. An Interaction between Kynurenine and the Aryl Hydrocarbon Receptor Can Generate Regulatory T Cells. J. Immunol. 2010, 185, 3190–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Zhang, L.; Hoagland, M.S.; Swanson, H.I. Lack of the Aryl Hydrocarbon Receptor Leads to Impaired Activation of AKT/Protein Kinase B and Enhanced Sensitivity to Apoptosis Induced via the Intrinsic Pathway. J. Pharmacol. Exp. Ther. 2007, 320, 448–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. | Compound | Structure | Mechanism(s) of Action | References |
|---|---|---|---|---|
| 1. | Luteolin |  | Regulates cell proliferation mediated by EGF, leading to the suppression of p-STAT3, p-EGFR, p-PKB, and p-Erk1/2 expression in MCF-7; induces cytoskeleton shrinkage and the expression of E-cadherin, while reducing the expressions of mesenchymal biomarkers N-cadherin, snail, and vimentin; inhibits EGFR and activation of Bax in pancreatic cancer; inhibits FASN in prostate cancer; activates FAS in oral cancer, and caspase-3, -9, TP53, and Bax in lung, gastric, and liver cancers; inhibits PI3K-Akt and DR5 in cervical cancer. | [112,113] |
| 2. | Quercetin |  | Activates TP53 gene, inhibits Bcl-2 expression; enhances efficacy and sensitization of doxorubicin against MCF-7 by reducing multidrug-resistant protein 1 gene expression level; induces G1-phase arrest and reduces the expression of Twist, Cyclin-D1, p21, and phospho p38 mitogen-activated protein kinases in MCF-7 and MDA-MB-231; inhibits PI3K, MAPK, AKT, COX-2, and Bcl-2 signaling and activates TP53, Bax, caspase-3, -8, and -9. | [99,100] |
| 3. | Chrysin |  | Inhibits aromatase enzyme in hormone-dependent breast cancer; induces apoptosis by caspase activation, Akt suppression, and stabilization of TP53 expression through ERK1/2 activation. | [107,108,109] |
| 4. | Rhamnetin |  | Upregulates caspase-3 and -9 and enhances TP53 protein and microRNA (miR-)34a. | [110] |
| 5. | Myricetin |  | Arrests G-quadruplex structure, represses hTERT expression in MCF-7; attenuates neoplastic transformation of cancer cells; suppresses cyclin-dependent kinase 1 (CDK1), PAK1, MEK, phosphorylated (p)-ERK1/2, β-catenin, cyclin D1, PCNA, and survivin. | [121,122] |
| 6. | Hesperetin |  | Reduces cellular proliferation by reducing cellular glucose uptake in MDA-MB-231; suppresses insulin receptor-beta subunit (IR-beta) and Akt; inhibits angiogenesis and metastasis by suppressing COX-2, MMP-2, and MMP-9; activates caspases and suppresses Bcl-2 and Bax to induce apoptosis. | [123,124] |
| 7. | Apigenin |  | Induces cell cycle arrest, apoptosis, and autophagy and suppresses cancer migration and invasion by inhibiting NEDD9/Src/AKT, PI3K/AKT, ERK1/2, JAK/STAT, Wnt/β-catenin, SAPK/JNK, and FAK molecular pathways and upregulating ATM/ATR and AMPK expression; suppresses overexpression of ERβ in breast and prostate cancerous cells. | [125,126] |
| 8. | Kaempferol |  | Upregulates TRAIL receptors and induces apoptosis in SW480 (human colon cancer line); induces apoptotic genes TP53, PUMA, NOXA, Bax, BCL-2, Apaf-1, caspase-9, and cytochrome c; downregulates Bcl-2 and Bcl-xL expression; reduces iNOS, COX2, CRP, and NF-κB protein levels; inhibits cancer invasion of breast cancer by blocking the PKCδ/MAPK/AP-1 cascade and subsequent MMP-9 expression. | [127,128,129] |
| 9. | Coumaric acid |  | Reduces the expression of COX-2, IL-6, TNF-α and PGE2; downregulates EGFR and GRP78 and activates the unfolded protein response (UPR) leading to apoptosis in cancer cells; modulates the expression of microRNAs in gastric cancer cells; induces Nrf2 transcription factor in colon cancer. | [130,131,132,133] |
| 10. | Ellagic acid |  | Reduces P-VEGFR2, MAPK, and PI3K/Akt expression in MDA-MB-231, descreases Bcl-2 levels, increases Bax levels, and inhibits SphK1 and integrin-linked kinase (ILK); reduces eicosanoid synthesis and downregulates the heme oxygenase (HO) system in prostate cancer. | [130,131,132,133] |
| 11. | Caffeic acid |  | Decreases the expression of TRIF, TLR4, and IRAK4 proteins in breast cancer cells; enhances HO-1, GCLC, and GCLM expression by the Nrf2/ERK pathway in liver cancer cells; enhances the activity of caspases and p53 enzymes and blocks Bcl-2 activity in cervical cancer. | [119] |
| 12. | Ferulic acid |  | Stops cell division by upregulating the expressions of ATR, ATM, CDKN1A, CDKN1B, E2F4, RB1, and TP53 and downregulating the expressions of CCND1, CCND2, CCND3, CDK2, CDK4, and CDK6 in PC-3 cells; upregulates the expressions of CASP1, CASP2, CASP8, CYCS, FAS, FASLG, and TRADD and downregulates the expressions of BCL2 and XIAP in LNCaP cells; inhibits autophagy-related proteins such as LC3-II, Beclin1, and Atg12-Atg5. | [134,135] |
| 13. | Cinnamic acid |  | Regulates oncogenes c-myc, c-fos, and TP53. | [136] |
| 14. | Epicatechin (R=R1= H)/epigallocatechin (R=OH, R1=H)/epigallocatechin-3-gallate (R=OH, R1=gallic acid) |  | Enhances the gene expression of GST, TP53, PTEN, CYP450, and Bax; suppresses VEGF, COX-2, and NF-KB activity. | [137] |
| 15. | Chlorogenic acid |  | Reduces the inflammatory markers IL-1β, IL-6, IL-8, COX-2, and TNF-α by down-regulating NF-KB expression; reduces ERK1/2, Akt/PI3K, EGFR, and Bcl-2 expressions. | [138] |
| 16. | Rosmarinic acid |  | Inhibits microtubule affinity regulating kinase (MARK4); suppresses inflammatory markers COX-2 and TNF-α and the expression of NF-KB; reduces MAPK/ERK and PI3K/Akt signaling pathways; activates caspases and PARP. | [139,140] |
| 17. | Caffeic acidphenethyl ester |  | Arrests cell cycle in G1 and G2/M; decreases the phosphorylation of EGFR in different cancer cells; reduces the expression of Akt, Akt1, Akt2, Akt3, phospho-Akt Ser473, phospho-Akt Thr308, GSK3β, FOXO1, FOXO3a, phospho-FOXO1 Thr24, and phospho-FoxO3a Thr32 in Tw2.6 cells. | [118] |
| Plant | Location | Active Extract | Main Constituents | Cell Lines/In Vivo Testing, IC50 Values | Proposed Mechanism | References |
|---|---|---|---|---|---|---|
| Anabasis articulata | Saudi Arabia | Aq. ethanolic extract | Kaempferol 3-neohesperidosid, 6-gingerol, triterpenes, steroidal saponins, and alkaloids | Panc1 (human pancreatic cancer cell line, derived from ductal cell pancreatic carcinoma), IC50 998.5 | [70] | |
| Egypt | Methylene chloride | HePG-2 (human, hepatic carcinoma cell), IC50 6.9; HCT-116, IC50 5.5 | [162] | |||
| Arthrocnemum indicum | Tunisia | Shoot aqueous methanol extract | Gallic acid, 3-hydroxy-4′-methoxyflavone, cyanidin, chrysoeriol, quercetin, catechol, syringic acid, luteolin | Shoot extracts inhibited Caco-2 (human, colorectal adenocarcinoma cells) colon cancer cell growth in a dose-dependent manner | Cell cycle blocking at the G2/M phase | [163] |
| Arthrocnemum macrostachyum | Egypt | Methanol extract | Phenolic acids and flavonoids | In vivo anticancer effect against Ehrlich solid tumor in mice | Increased tissue necrosis and apoptosis, enhanced DNA fragmentation, upregulated cell cycle regulatory genes (Cdc2 and connexin26), and decreased TNFa levels in tumor tissues | [164] |
| Asplenium ceterach | Bulgaria | Aqueous methanol | Phenolic acids and flavonoids | A549 (human, adenocarcinoma, hypotriploid alveolar basal epithelial cells), FL, HeLa (IC50 40.48) | Strong proapoptotic potential against HeLa (human, cervical cancer cell line) | [165] |
| Avicennia marina | Saudi Arabia | Hexane fraction | Betaine and hymecromone | HCT-116, IC50 23.7; HepG2, IC50 44.9; MCF-7, IC50 79.55 | Inhibition of cell cycle in G0/G1 and S phases in HepG2 and MCF-7 | [151] |
| Carpobrotus edulis | Portugal | Leaf methanol extract and different fractions. | β-amyrin, uvaol, oleanolic acid, monogalactosyl diacylglycerol, catechin, epicatechin, and procyanidin B5 | L5178 (mouse, T-cell lymphoma cells), and L5178 (mouse, T-cell lymphoma cells) transfected with pHa MDR1/A retrovirus | Inhibition of P-glycoprotein in MDR1-transfected mouse lymphoma cells | [166] |
| Chenopodium formosanum | Taiwan | Grain extract | Polyphenols and prebiotic dietary fiber | In vivo colon carcinogenesis induced by 1,2-dimethylhydrazine and dextran sulfate sodium in rats | Increase Bax and caspase-9 expressions; reduced TP53 and Bcl-2 expression; decreased expressions of proliferating cell nuclear antigen and cyclooxygenase-2; regulation of apoptosis-related proteins | [167,168] |
| Mesembryanthemum crystallinum | Korea | Ethanol extracts and its fractions | Phenolics and flavonoids | Inhibition of HCT116 cell growth in dose-dependent manner | Increased G2/M cell population and increased ROS levels in cells | [169] |
| Echinophora spinosa | Italy | Essential oils | p-Cymene, β-Phellandrene, β-Phellandrene, myristicin | U937, IC50 14.5–43.4 | Induced apoptosis in U937 cell line (human monocytic cell based) | [170] |
| Glaucium flavum | Tunis | Ethyl acetate extract | Isoquinoline alkaloids, kaempferol, caffeic acid, catechin hydrate, syringic acid, chlorogenic acid, isoquercitrin, and trans-hydroxycinnamic acid | MCF-7, IC50 135 | [171] | |
| Algeria | CH2Cl2 extract | MDA-MB-435, MDA-MB-231, and Hs578T (IC50 7.9–13.6) as well as in vivo tumor chorioallantoic membrane (CAM) model | Hinders angiogenesis, induction of apoptotic processes, and/or limited neovessel formation inside the tumor | [172] | ||
| Iran | Methanol extract and rich alkaloid fraction | HT-29, IC50 22.32 L; Caco-2, IC50 52.38 | [173] | |||
| Glehnia littoralis | Korea | Hexane fraction Aqueous methanol fraction | Furanocoumarin bitter principle and polyacetylene alcohols | HT-29 (77% inhibition at 50 mg/mL extract) | Induced chromatin condensation and nuclear fragmentation, suggesting the presence of apoptotic cells; reduced mRNA expression of Bcl-2, cyclooxygenase (COX-2), and inducible nitric oxide synthase (iNOS) | [174] |
| Limonium densiflorum | Tunisia | CHCl3 extract | Gallic acid, epigallocatechin, quercitrin, myricetin, dihydrokaempferol, isorhamnetin | A-549, IC50 29 µg/mL; DLD-1, IC50 85) | Isorhamnetin induced apoptosis through activation of peroxisome proliferator-activated receptor γ pathway in gastric cancer | [74,175] |
| Limonium bonduelli | Algeria | n-Butanol extract | Flavonoids (eriodictyol, luteolin, apigenin) and 4-hydroxy-3-methoxy benzoic acid; ethyl acetate extract of L. bonduelli and pure flavonoids, eriodictyol and luteolin | Dose-dependent growth inhibition of HT-29 and HeLa cell-lines | [176] | |
| Lotus creticus L | Portugal | Acetone extract (aerial part) Ethanol extract (fruits) | Steroids, coumarins, tannins, and flavonoids, e.g., catechin, epicatechin, isorhamnetin, quercetin, isorhamnetin-O-hexoside, quercetin-O-hexoside, myricetin-O-hexoside | Extracts had low toxicity RAW 264.7 | [177] | |
| Lycium shawii | Saudi Arabia | Aqueous ethanol extract | Flavonoids, 3-gluco-7-rhamnosyl quercetin, luteolin 7-O-glucoside, kaempferol-3-O-glucoside | MCF7, 194.5 µg/mL; K562, 464.9 µg/mL | Induced apoptosis and cell membrane damage due to necrosis and late apoptosis | [70] |
| Malcolmia littorea | Portugal | Polar extracts of flower and roots | Phenolic acids and flavonoids including salicylic acid and luteolin-7-O-glucoside. | HepG2 (viability 38.3%) HEK 293 cells (viability 93.1%) | [178] | |
| Mentha piperita | China | Essential oils | Menthyl acetate, cineol, menthol, pulegone, and caryophyllene oxide | SPC-A1, IC50 10.89; K562, IC50 16.16; and SGC-7901, IC50 38.76. | [159] | |
| Pulicaria undulata | Saudi Arabia | Aqueous ethanolic extract | Flavonoids of kaempferol-, luteolin-, and quercetin-based glycosides | MCF-7, 519.2 µg/mL; K562, 1212 µg/mL; PANC-1, 1535 µg/mL | Cell cycle arrest at the Q1 and Q2 quadrants, and necrosis in late apoptosis | [154] |
| Pulicaria crispa | Saudi Arabia | Aqueous ethanolic extract | Sterols, triterpenoids, essential oils, phenolics, and flavonoids | MDA-MB-231, IC50 180 µg/mL | Loss of cancer cell integrity, shrinkage of cytoplasm, and cell detachment | [179] |
| Reaumuria vermiculata | Tunisia | Hexane and CH2Cl2 | Myricetin, phenolics, and flavonoids | A-549, IC50 17, (hexane extract), and 23 (dichloromethane extract) | [158] | |
| Egypt | Aqueous methanol extract | Huh-7, IC50 2.4; HCT-116, IC50 1.8; MCF-7, IC50 1.3; PC-3, IC50 1.5 | [180] | |||
| Salicornia herbacea | Korea | Crude and fine polysaccharide | Polysaccharides and phenolic compounds | HT-29 | Inhibition of cyclin B1 and Cdc2 mRNAG2/M arrest | [181] |
| Salvadora persica | Saudi Arabia | Ethanol extracts of fruits | Essential oils, alkaloids, steroids, cetyl dasycarpidan-1-methanol, tetracosamethyl-cyclododecasiloxane, eicosamethyl-cyclodecasiloxane, and 1-monolinoleoylglycerol | MCF7, IC50 17.50; A2780, IC50 8.35; HT29, IC50 5.12 | [182] | |
| Salvadora persica L | Egypt | Bark petroleum ether | HepG, IC50 43.6l; MCF-7, IC50 44.3; A549, IC50 19.87 L | [183] | ||
| Suaeda fruticosa | Pakistan | Methanol and CHCl3 extracts | Phenolics, flavonoids, saponins, fatty acids | MCF-7 (63.44% and 45.01% cell viability in methanol and CH2Cl2 at 200 μg/mL), MDA-MB-231 (77.75% and 67.22% cell viability in methanol and dichloromethane at 200 μg/mL), and DU-145 (62.83% and 25.88% cells viability in methanol and dichloromethane at 200 μg/mL) | [184] | |
| Tunisia | CH2Cl2 extract | A-549, IC50 49 ± 7; DLD-1, IC50 10 ± 1; Caco-2, IC50 140 ± 13 µg/mL; HT-29, (IC50 12 ± 14 | [157] | |||
| Saudi Arabia | Hexane extract | HCT-116, IC50 17.15; MCF-7, IC50 28.1; HepG2, IC50 33.2 | Arrest the cell cycle at the G0-G1 phase | [150] | ||
| Tamarix gallica | Tunisia | Methanolic extracts | Phenolic acids and flavonoids | Caco-2, 38% inhibition in cell growth at 100 µg/mL | Decreased DNA synthesis, arrested cell mitosis at G2/M phase; changes in the cell-cycle-associated proteins (cyclin B1, p38, Erk1/2, Chk1, and Chk2) correlated with changes in the cell cycle distribution | [185] |
| India | Protects against liver carcinogenesis initiated by diethylnitrosamine and 2-acetylaminofluorene | Restoration of cellular antioxidant enzymes, detoxifying enzymes, ODC activity, and DNA synthesis. | [161] | |||
| Zygophyllum album | Tunisia | CH2Cl2 extract | Isorhamnetin-3-O-rutinoside, quinovic acid derivatives, malvidin 3-rhamnoside, quercetin 3-sulfate | A-549, IC50 37; DLD-1, IC50 48 | Downregulation of cyclin B1 and cyclin dependent kinase; upregulation of TP53 and caspase 3 | [155] |
| Egypt | HepG2 IC50 27.74 | [186] | ||||
| Zygophyllum coccineum | Saudi Arabia | Aqueous ethanolic extract | Phenolics, flavonoids, alkaloids, quinovic acid derivatives. | MCF-7, IC50 3.47; HCT-116, IC50 3.19; HepG2, IC50 2.27 | Inhibition of human topoisomerase-IIβ | [153] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed, H.A.; Emwas, A.-H.; Khan, R.A. Salt-Tolerant Plants, Halophytes, as Renewable Natural Resources for Cancer Prevention and Treatment: Roles of Phenolics and Flavonoids in Immunomodulation and Suppression of Oxidative Stress towards Cancer Management. Int. J. Mol. Sci. 2023, 24, 5171. https://doi.org/10.3390/ijms24065171
Mohammed HA, Emwas A-H, Khan RA. Salt-Tolerant Plants, Halophytes, as Renewable Natural Resources for Cancer Prevention and Treatment: Roles of Phenolics and Flavonoids in Immunomodulation and Suppression of Oxidative Stress towards Cancer Management. International Journal of Molecular Sciences. 2023; 24(6):5171. https://doi.org/10.3390/ijms24065171
Chicago/Turabian StyleMohammed, Hamdoon A., Abdul-Hamid Emwas, and Riaz A. Khan. 2023. "Salt-Tolerant Plants, Halophytes, as Renewable Natural Resources for Cancer Prevention and Treatment: Roles of Phenolics and Flavonoids in Immunomodulation and Suppression of Oxidative Stress towards Cancer Management" International Journal of Molecular Sciences 24, no. 6: 5171. https://doi.org/10.3390/ijms24065171
APA StyleMohammed, H. A., Emwas, A. -H., & Khan, R. A. (2023). Salt-Tolerant Plants, Halophytes, as Renewable Natural Resources for Cancer Prevention and Treatment: Roles of Phenolics and Flavonoids in Immunomodulation and Suppression of Oxidative Stress towards Cancer Management. International Journal of Molecular Sciences, 24(6), 5171. https://doi.org/10.3390/ijms24065171