
Transcriptomic Profile of Canine Mammary Ductal Carcinoma
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
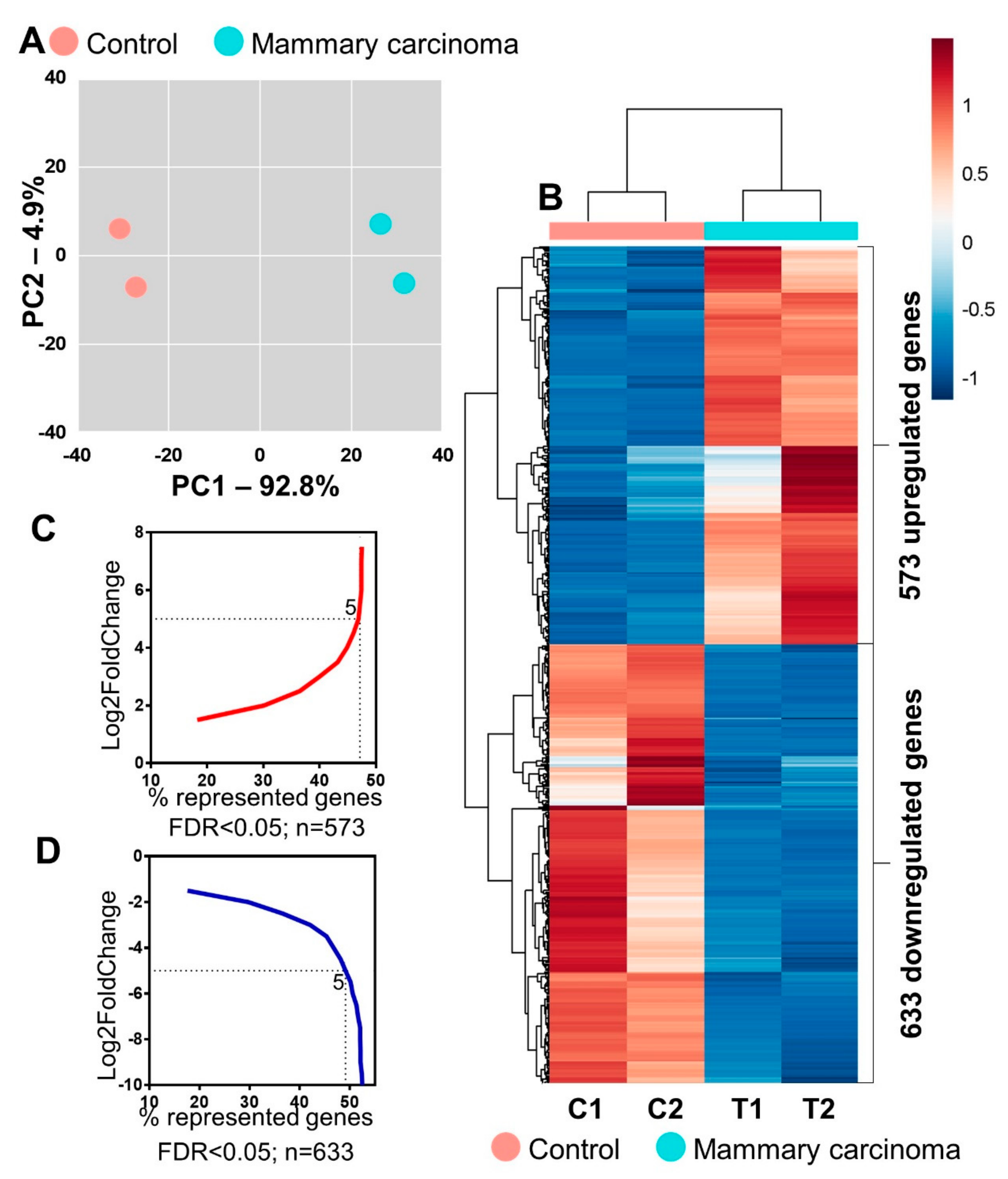
2. Results
3. Discussion
4. Materials and Methods
4.1. Casuistry
4.2. Ethical Premises
4.3. Methodology
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Biller, B.; Berg, J.; Garret, L.; Ruslander, D.; Wearing, R.; Abbott, B.; Patel, M.; Smith, D.; Bryan, C. 2016 AAHA oncology guidelines for dogs and cats. J. Am. Anim. Hosp. Assoc. 2016, 52, 181–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Sorenmo, K.U.; Rasotto, R.; Zappulli, V.; Goldschmidt, M.H. Development, anatomy, histology, lymphatic drainage, clinical features, and cell differentiation markers of canine mammary gland neoplasms. Vet. Pathol. 2011, 48, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilhena, H.; Figueira, A.C.; Schmitt, F.; Canadas, A.; Chaves, R.; Gama, A.; Dias-Pereira, P. Canine and feline spontaneous mammary tumours as models of human breast cancer. In Pets as Sentinels of Human Health in Pets as Sentinels, Forecasters and Promoters of Human Health; Pastorinho, M., Ed.; Springer: Cham, Switzerland, 2019; pp. 137–207. [Google Scholar]
- Rivera, P.; von Euler, H. Molecular biological aspects on canine and human mammary tumors. Vet. Pathol. 2011, 48, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Strandberg, J.D.; Goodman, D.G. Animal model: Canine mammary neoplasia. Am. J. Pathol. 1974, 75, 225–228. [Google Scholar]
- Liu, D.; Xing, H.; Ellis, A.E.; Northrup, N.C.; Rodriguez, C.O.; O’Regan, R.M.; Dalton, S.; Zhao, S. Molecular homology and difference between spontaneous canine mammary cancer and human breast cancer. Cancer Res. 2014, 74, 5045–5056. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, Y.; Morimatsu, M.; Ochiai, K.; Ishaiguro-Oonuma, T.; Wada, S.; Orino, K.; Watanabe, K. Reduced canine BRCA2 expression levels in mammary gland tumors. BMC Vet. Res. 2015, 11, 159. [Google Scholar] [CrossRef] [Green Version]
- Nieto, A.; Pérez-Alenza, M.D.; Del Castillo, N.; Tabanera, E.; Castaño, M.; Peña, L. BRCA1 expression in canine mammary dysplasias and tumours: Relationship with prognostic variables. J. Comp. Pathol. 2003, 128, 260–268. [Google Scholar] [CrossRef]
- Gray, M.; Meehan, J.; Martínez-Pérez, C.; Kay, C.; Turnbull, A.K.; Morrison, L.R.; Pang, L.Y.; Argyle, D. Naturally-Occurring Canine Mammary Tumors as a Translational Model for Human Breast Cancer. Front. Oncol. 2020, 10, 617. [Google Scholar] [CrossRef]
- Goldschmidt, M.H.; Peña, L.; Rasotto, R.; Zappulli, V. Classification and grading of canine mammary tumors. Vet. Pathol. 2011, 48, 117–131. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’Ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Clarke, D.J.B.; Kuleshov, M.V.; Shilder, B.M.; Torre, D.; Duffy, M.E.; Keenan, A.B.; Lachmann, A.; Feldmann, A.S.; Gundersen, G.W.; Ma’Ayan, A.; et al. eXpression2Kinases (X2K) Web: Linking expression signatures to upstream cell signaling networks. Nucleic Acids Res. 2018, 46, W171–W179. [Google Scholar] [CrossRef] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Taylor, G.N.; Shabestari, L.; Williams, J.; Mays, C.W.; Angus, W.; McFarlande, S. Mammary neoplasia in a closed beagle colony. Cancer Res. 1976, 36, 2740–2743. [Google Scholar]
- National Center for Biotechnology Information. NCBI Canis lupus Familiaris Annotation Release 105, National Library of Medicine. Available online: https://www.ncbi.nlm.nih.gov/genome/annotation_euk/Canis_lupus_familiaris/105/ (accessed on 11 January 2021).
- Lis, C.G.; Grutsch, J.F.; Vashi, P.G.; Lammersfeld, C.A. Is serum albumin an independent predictor of survival in patients with breast cancer? J. Parenter. Enter. Nutr. 2003, 27, 10–15. [Google Scholar] [CrossRef]
- Xie, X.W.; Jiang, S.S.; Li, X. CLEC3B as a potential prognostic biomarker in hepatocellular carcinoma. Front. Mol. Biosci. 2021, 7, 614034. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, D.; Zhu, Y.; Guo, H.; Zhao, X.; Sun, B. Clusterin expression and univariate analysis of overall survival in human breast cancer. Technol. Cancer Res. Treat. 2006, 5, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Sayers, K.T.; Brooks, A.D.; Sayers, T.J.; Chertov, O. Increased Secretory Leukocyte Protease Inhibitor (SLPI) production by highly metastatic mouse breast cancer cells. PLoS ONE 2014, 9, e104223. [Google Scholar] [CrossRef]
- Wagenblast, E.; Soto, M.; Gutiérrez-Ángel, S.; Hartl, C.A.; Gable, A.L.; Maceli, A.R.; Erard, N.; Williams, A.M.; Kim, S.Y.; Canott, S.R.; et al. A model of breast cancer heterogeneity reveals vascular mimicry as a driver of metastasis. Nature 2015, 520, 358–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koltai, T. Clusterin: A key player in cancer chemoresistance and its inhibition. Onco. Targets. Ther. 2014, 7, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Nugteren, S.; Samsom, J.N. Secretory Leukocyte Protease Inhibitor (SLPI) in mucosal tissues: Protects against inflammation, but promotes cancer. Cytokine Growth Factor Rev. 2021, 59, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Shen, Y.; Jiang, X.; Peng, M.; Liu, Y.; Mo, Y.; Ren, D.; Hua, Y.; Yu, B.; Zeng, Z.; et al. Mechanisms of vasculogenic mimicry in hypoxic tumor microenvironments. Mol. Cancer 2021, 20, 7. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Liu, Y.E.; Gentz, R.; Sang, Q.A.; Ni, J.; Goldberg, I.D.; Chi, Y.E. Suppression of breast cancer growth and metastasis by a serpin myoepithelium-derived serine proteinase inhibitor expressed in the mammary myoepithelial cells. Proc. Natl. Acad. Sci. USA 1999, 96, 3700–3705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scimone, C. Possible related functions of the non-homologous co-regulated gene pair PDCD10 and serpini1. EuroMediterranean Biomed. J. 2017, 12, 41–46. [Google Scholar] [CrossRef]
- Neer, E.J.; Schmidt, C.J.; Nambudripad, R.; Smith, T.F. The ancient regulatory-protein family of WD-repeat proteins. Nature 1994, 371, 297–300. [Google Scholar] [CrossRef]
- Li, D.; Roberts, R. WD-repeat proteins: Structure characteristics, biological function, and their involvement in human diseases. Cell. Mol. Life Sci. 2001, 58, 2085–2097. [Google Scholar] [CrossRef]
- Han, C.; Yu, T.; Qin, W.; Liao, X.; Huang, J.; Liu, Z.; Yu, L.; Liu, X.; Chen, Z.; Yang, C.; et al. Genome-wide association study of the TP53 R249S mutation in hepatocellular carcinoma with aflatoxin B1 exposure and infection with hepatitis B virus. J. Gastrointest. Oncol. 2020, 11, 1333–1349. [Google Scholar] [CrossRef]
- Kauppila, S.; Stenbäck, F.; Risteli, J.; Jukkola, A.; Risteli, L. Aberrant type I and type III collagen gene expression in human breast cancer in vivo. J. Pathol. 1998, 186, 262–268. [Google Scholar] [CrossRef]
- Case, A.; Brisson, B.K.; Durhan, A.C.; Rosen, S.; Monslow, J.; Buza, E.; Salah, P.; Gillem, J.; Ruthel, G.; Volk, S.W.; et al. Identification of prognostic collagen signatures and potential therapeutic stromal targets in canine mammary gland carcinoma. PLoS ONE 2017, 12, e0180448. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Saxena, S.; Shrivastava, S.; Mohanty, A.K.; Kumar, S.; Singh, R.J.; Kumar, A.; Wani, S.A.; Gandham, R.K.; Singh, R.K.; et al. Gene expression profiling of spontaneously occurring canine mammary tumours: Insight into gene networks and pathways linked to cancer pathogenesis. PLoS ONE 2018, 13, e0208656. [Google Scholar] [CrossRef] [Green Version]
- Amini, P.; Nassiri, S.; Ettlin, J.; Malbon, A.; Markkanen, E. Next-generation RNA sequencing of FFPE subsections reveals highly conserved stromal reprogramming between canine and human mammary carcinoma. DMM Dis. Model. Mech. 2019, 12, dmm040444. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, S.I.; Utturkar, S.; Lee, M.; Yang, H.H.; Kui, Z.; Lanman, N.A.; Zheng, G.; Cardona, X.E.R.; Mittal, S.K.; Miller, M.A. Ductal carcinoma in situ progression in dog model of breast cancer. Cancers 2020, 12, 418. [Google Scholar] [CrossRef] [Green Version]
- Al Saleh, S.; Al Mulla, F.; Luqmani, Y.A. Estrogen receptor silencing induces epithelial to mesenchymal transition in human breast cancer cells. PLoS ONE 2011, 6, e20610. [Google Scholar] [CrossRef]
- Ma, J.; Zhao, J.; Wang, P.; Feng, H.; Zong, Y.; Ou, B.; Zheng, M.; Lu, A. Cadherin-12 enhances proliferation in colorectal cancer cells and increases progression by promoting EMT. Tumor Biol. 2016, 37, 9077–9088. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.Y.; Fu, Z.D.; Dempsey, J. The Role of Histone Methylation and Methyltransferases in Gene Regulation. In Toxicoepigenetics; McCullough, S.D., Dolinoy, D.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Chapter 1–2; pp. 31–84. [Google Scholar]
- Ye, M.; Li, L.; Liu, D.; Wang, Q.; Zhang, Y.; Zhang, J. Identification and validation of a novel zinc finger protein-related gene-based prognostic model for breast cancer. PeerJ 2021, 9, e12276. [Google Scholar] [CrossRef]
- Kusafuka, K.; Muramatsu, K.; Kasami, M.; Kuriki, K.; Hirobe, K.; Hayashi, I.; Watanabe, H.; Hiraki, Y.; Shukunami, C.; Mochizuki, T.; et al. Cartilaginous features in matrix-producing carcinoma of the breast: Four cases report with histochemical and immunohistochemical analysis of matrix molecules. Mod. Pathol. 2008, 21, 1282–1292. [Google Scholar] [CrossRef] [Green Version]
- Shawky, M.S.; Ricciardelli, C.; Lord, M.; Whitelock, J.; Ferro, V.; Britt, K.; Thompson, E.W. Proteoglycans: Potential agents in mammographic density and the associated breast cancer risk. J. Mammary Gland Biol. Neoplasia 2015, 20, 121–131. [Google Scholar] [CrossRef]
- Eshchenko, T.Y.; Rykova, V.I.; Chernakov, A.E.; Sidorov, S.V.; Grigorieva, E.V. Expression of different proteoglycans in human breast tumors. Biochemistry 2007, 72, 1016–1020. [Google Scholar] [CrossRef]
- Cadenas, C.; Franckenstein, D.; Schmidt, M.; Gehrmann, M.; Hermes, M.; Geppert, B.; Schormann, W.; Maccoux, L.J.; Schug, M.; Schumann, A.; et al. Role of thioredoxin reductase 1 and thioredoxin interacting protein in prognosis of breast cancer. Breast Cancer Res. 2010, 12, R44. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ning, J.; Cao, W.; Wang, S.; Du, T.; Jiang, J.; Feng, X.; Zhang, B. Research progress of TXNIP as a tumor suppressor gene participating in the metabolic reprogramming and oxidative stress of cancer cells in various cancers. Front. Oncol. 2020, 10, 568574. [Google Scholar] [CrossRef]
- Park, J.W.; Lee, S.H.; Woo, G.H.; Kwon, H.J.; Kim, D.Y. Downregulation of TXNIP leads to high proliferative activity and estrogen-dependent cell growth in breast cancer. Biochem. Biophys. Res. Commun. 2018, 498, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Park, H.M.; Son, K.H.; Shin, T.J.; Cho, J.Y. Transcriptome signatures of canine mammary gland tumors and its comparison to human breast cancers. Cancers 2018, 10, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pöschel, A.; Beebe, E.; Kunz, L.; Amini, P.; Guscetti, F.; Malbon, A.; Markkanen, E. Identification of disease-promoting stromal components by comparative proteomic and transcriptomic profiling of canine mammary tumors using laser-capture microdissected FFPE tissue. Neoplasia 2021, 23, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Bhat, F.A.; Mohan, S.V.; Patil, A.; Advani, J.; Bhat, M.Y.; Patel, K.; Mangalaparthi, K.K.; Datta, K.K.; Routray, A.; Mohanty, N.; et al. Proteomic alterations associated with oral cancer patients with tobacco using habits. OMICS J. Integr. Biol. 2021, 25, 255–268. [Google Scholar] [CrossRef]
- Kengkarn, S.; Petmitr, S.; Boonyuen, U.; Reamtong, O.; Poomsawat, S.; Sanguansin, S. Identification of novel candidate biomarkers for oral squamous cell carcinoma based on whole gene expression profiling. Pathol. Oncol. Res. 2020, 26, 2315–2325. [Google Scholar] [CrossRef]
- Zhang, H.X.; Liu, O.S.; Deng, C.; He, Y.; Feng, Y.Q.; Ma, J.A.; Hu, C.H.; Tang, Z.G. Genome-wide gene expression profiling of tongue squamous cell carcinoma by RNA-seq. Clin. Oral Investig. 2018, 22, 209–216. [Google Scholar] [CrossRef]
- Shaikh, I.; Ansari, A.; Ayachit, G.; Gandhi, M.; Sharma, P.; Bhairappanavar, S.; Joshi, C.G.; Das, J. Differential gene expression analysis of HNSCC tumors deciphered tobacco dependent and independent molecular signatures. Oncotarget 2019, 10, 6168–6183. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Zhang, S.; Zhang, D.; Tao, Q.; Zhang, T.; Liu, G.; Liu, X.; Zhao, T. Identification of SERPINE1, PLAU and ACTA1 as biomarkers of head and neck squamous cell carcinoma based on integrated bioinformatics analysis. Int. J. Clin. Oncol. 2019, 24, 1030–1041. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Xie, Q.; Zhang, G.; Li, S.; Wu, Z.; Ma, Z.; He, X.; Gao, Y.; Wang, Y.; Kang, X.; et al. Slow skeletal muscle troponin T, titin and myosin light chain 3 are candidate prognostic biomarkers for Ewing’s sarcoma. Oncol. Lett. 2019, 18, 6431–6442. [Google Scholar] [CrossRef] [Green Version]
- Bizama, C.; Benavente, F.; Salvatierra, E.; Gutiérrez-Moraga, A.; Espinoza, J.A.; Fernández, E.A.; Roa, I.; Mazzolini, G.; Sagredo, E.A.; Gidekel, M.; et al. The low-abundance transcriptome reveals novel biomarkers, specific intracellular pathways and targetable genes associated with advanced gastric cancer. Int. J. Cancer 2014, 134, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Midha, M.K.; Huang, Y.; Yang, H.; Fan, T.; Chang, N.; Chen, T.; Wang, Y.; Kuo, W.; Chang, K.; Shen, C.; et al. Comprehensive cohort analysis of mutational spectrum in early onset breast cancer patients. Cancers 2020, 12, 2089. [Google Scholar] [CrossRef]
- Lian, J.; Li, K. A review of breast density implications and breast cancer screening. Clin. Breast Cancer 2020, 20, 283–290. [Google Scholar] [CrossRef]
- Kristiansen, G.; Hu, J.; Wichmann, D.; Stiehl, D.P.; Rose, M.; Gerhardt, J.; Bohnert, A.; ten Haaf, A.; Moch, H.; Raleigh, J.; et al. Endogenous myoglobin in breast cancer is hypoxia-inducible by alternative transcription and functions to impair mitochondrial activity: A role in tumor suppression? J. Biol. Chem. 2011, 286, 43417–43428. [Google Scholar] [CrossRef] [Green Version]
- Bicker, A.; Brahmer, A.M.; Meller, S.; Kristiansen, G.; Gorr, T.A.; Hankein, T. The distinct gene regulatory network of myoglobin in prostate and breast cancer. PLoS ONE 2015, 10, e0142662. [Google Scholar] [CrossRef] [Green Version]
- Kristiansen, G.; Rose, M.; Geisler, C.; Fritzsche, F.R.; Gerhardt, J.; Lüke, C.; Ladhoff, A.-M.; Knüchel, R.; Diete, M.; Moch, H.; et al. Endogenous myoglobin in human breast cancer is a hallmark of luminal cancer phenotype. Br. J. Cancer 2010, 102, 1736–1745. [Google Scholar] [CrossRef] [Green Version]
- Braganza, A.; Quesnelle, K.; Bickta, J.; Reyes, C.; Wang, Y.; Jessup, M.; Croix, C.S.; Arlotti, J.; Singh, S.V.; Shiva, S. Myoglobin induces mitochondrial fusion, thereby inhibiting breast cancer cell proliferation. J. Biol. Chem. 2019, 294, 7269–7282. [Google Scholar] [CrossRef]
- Laursen, I.; Briand, P.; Lykkesfeldt, A.E. Serum albumin as a modulator on growth of the human breast cancer cell line, MCF-7. Anticancer Res. 1990, 10, 343–351. [Google Scholar]
- Hwang, K.; Chung, J.K.; Roh, E.Y.; Kim, J.; Oh, S.; Kim, Y.A.; Rhu, J.; Kim, S. Prognostic influence of preoperative fibrinogen to albumin ratio for breast cancer. J. Breast Cancer 2017, 20, 254–263. [Google Scholar] [CrossRef]
- Liu, X.; Meng, Q.H.; Ye, Y.; Hildebrandt, M.A.T.; Gu, J.; Wu, X. Prognostic significance of pretreatment serum levels of albumin, LDH and total bilirubin in patients with nonmetastatic breast cancer. Carcinogenesis 2015, 36, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Röcken, C.; Kronsbein, H.; Sletten, K.; Roessner, A.; Bässler, R. Amyloidosis of the breast. Virchows Arch. 2002, 440, 527–535. [Google Scholar] [CrossRef]
- Stein, T.; Salomonis, N.; Nuyten, D.S.A.; Vijver, M.J.; Gusterson, B.A. A mouse mammary gland involution mRNA signature identifies biological pathways potentially associated with breast cancer metastasis. J. Mammary Gland Biol. Neoplasia 2009, 14, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Noguchi, K.; Hachiya, N.; Kametani, F.; Tasaki, M.; Nakaba, S.; Sassa, Y.; Yamashita, T.; Obayashi, K.; Ando, Y.; et al. Needle-shaped amyloid deposition in rat mammary gland: Evidence of a novel amyloid fibril protein. Amyloid 2020, 27, 25–35. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, D.B.; Fernandez, G.J.; Pardini, L.M.C.; Pardini, M.I.M.C.; Ferrasi, A.C. Transcriptomic Profile of Canine Mammary Ductal Carcinoma. Int. J. Mol. Sci. 2023, 24, 5212. https://doi.org/10.3390/ijms24065212
Santos DB, Fernandez GJ, Pardini LMC, Pardini MIMC, Ferrasi AC. Transcriptomic Profile of Canine Mammary Ductal Carcinoma. International Journal of Molecular Sciences. 2023; 24(6):5212. https://doi.org/10.3390/ijms24065212
Chicago/Turabian StyleSantos, Driéle B., Geysson J. Fernandez, Luciana M. C. Pardini, Maria Inês M. C. Pardini, and Adriana C. Ferrasi. 2023. "Transcriptomic Profile of Canine Mammary Ductal Carcinoma" International Journal of Molecular Sciences 24, no. 6: 5212. https://doi.org/10.3390/ijms24065212
APA StyleSantos, D. B., Fernandez, G. J., Pardini, L. M. C., Pardini, M. I. M. C., & Ferrasi, A. C. (2023). Transcriptomic Profile of Canine Mammary Ductal Carcinoma. International Journal of Molecular Sciences, 24(6), 5212. https://doi.org/10.3390/ijms24065212