
Mutation at the Site of Hydroxylation in the Ribosomal Protein uL15 (RPL27a) Causes Specific Changes in the Repertoire of mRNAs Translated in Mammalian Cells
 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
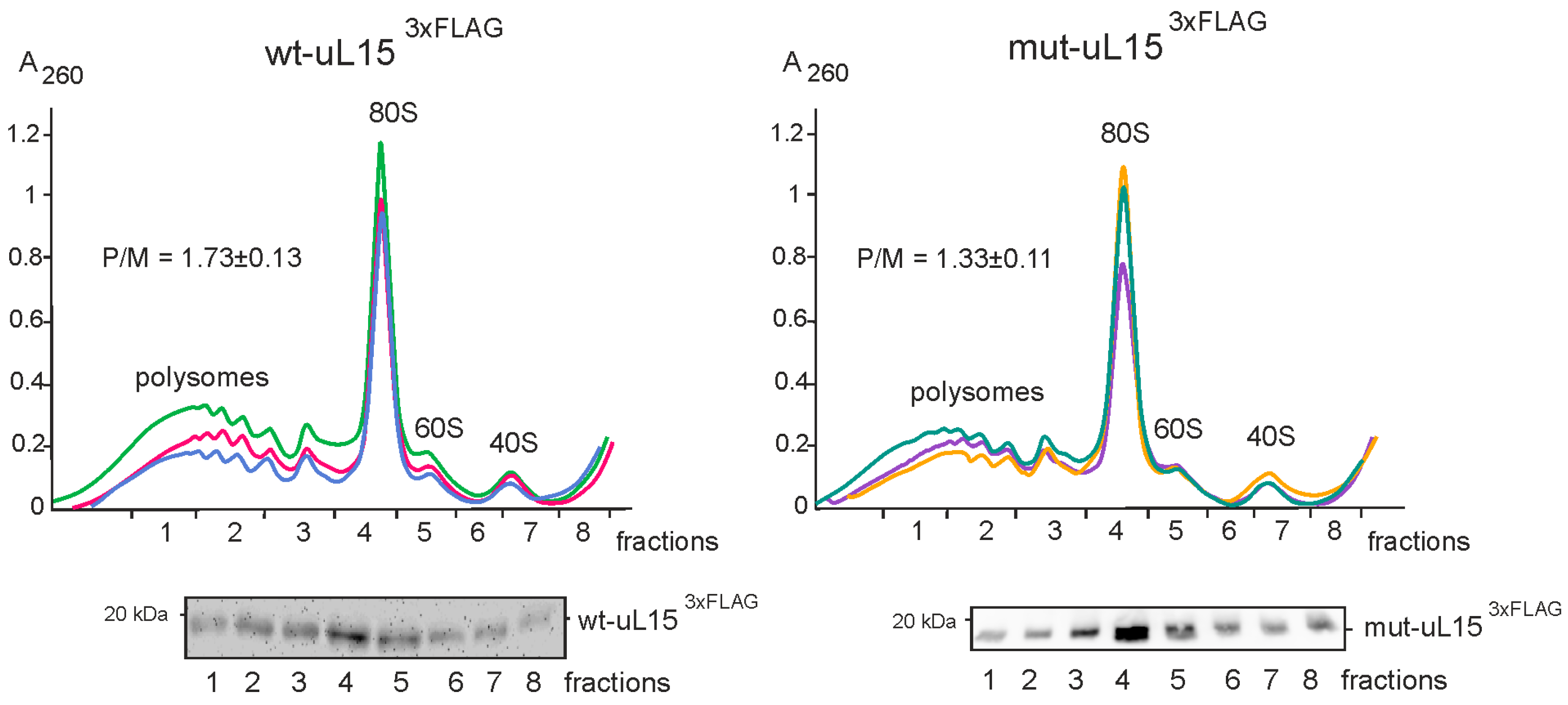
2.1. Incorporation of the Mutant uL15 Incapable of Hydroxylation into Ribosomes Reduces Translational Activity in HEK293T Cells
2.2. RNA-Seq of Total and Polysome-Associated mRNAs from Cells Producing Mut-uL153×FLAG or Its Wild-Type Form
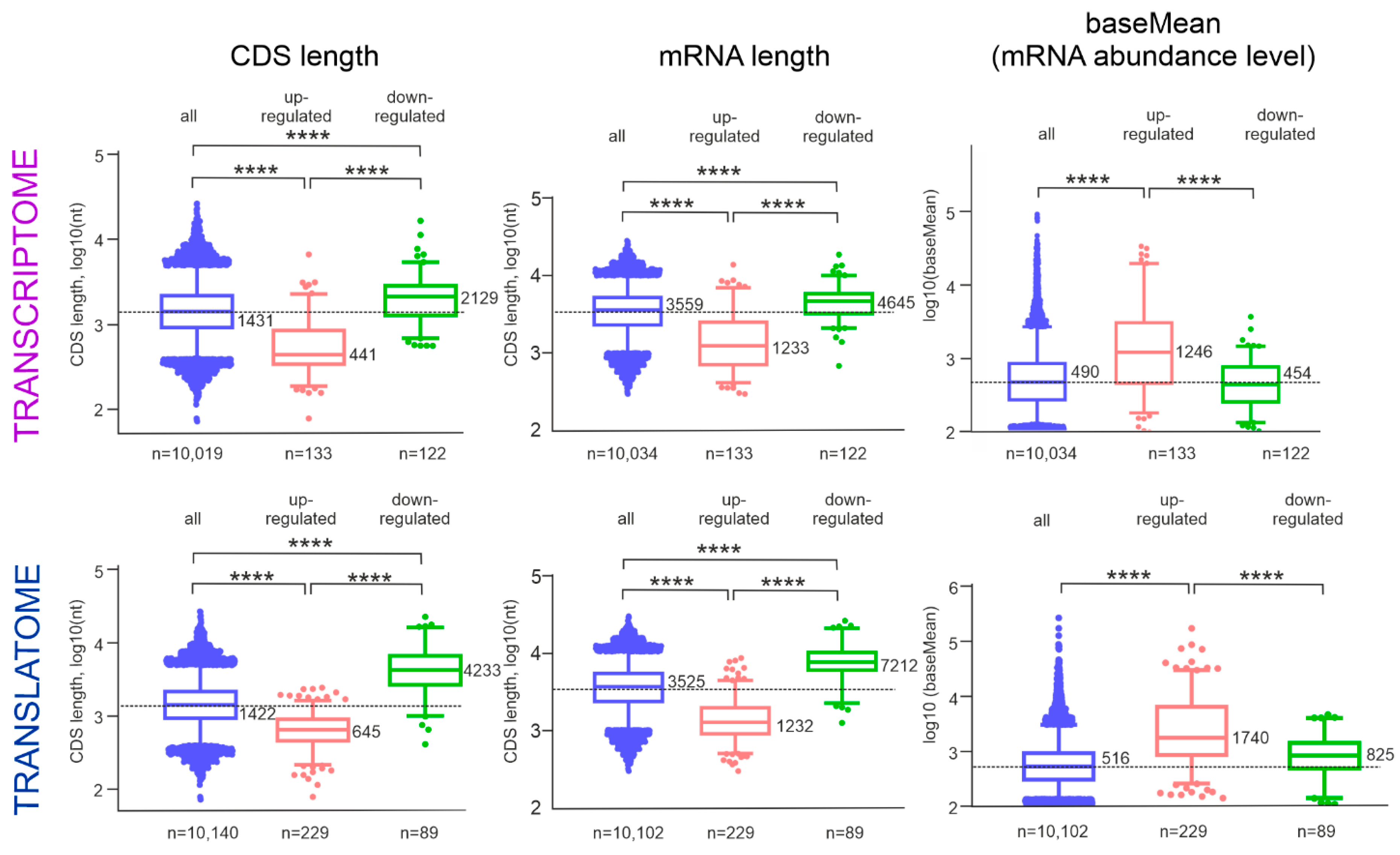
2.3. Analysis of the tDEG and pDEG mRNAs
3. Discussion
4. Materials and Methods
4.1. Plasmids, Cell Culturing, and Target Protein Production Analysis
4.2. DNA Library Preparation and Next-Generation Sequencing
4.3. Raw RNA-Seq Data Processing
4.4. Downstream Analysis of Processed RNA-Seq Data
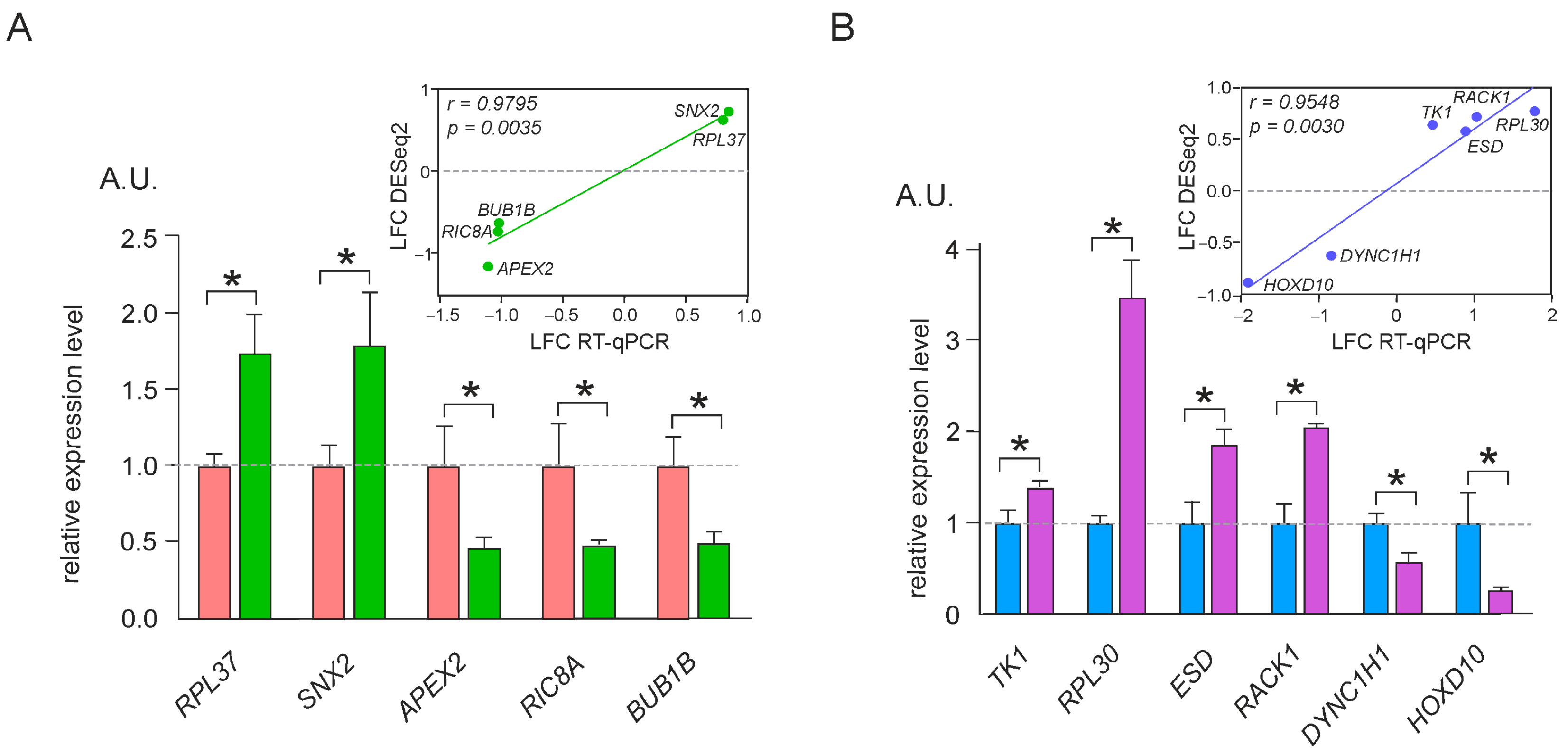
4.5. Validation of RNA-Seq Results Using RT-qPCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, D.N.; Cate, J.H.D. The structure and function of the eukaryotic ribosome. Cold Spring Harb. Perspect. Biol. 2012, 4, a011536. [Google Scholar] [CrossRef] [PubMed]
- Melnikov, S.; Ben-Shem, A.; Garreau de Loubresse, N.; Jenner, L.; Yusupova, G.; Yusupov, M. One core, two shells: Bacterial and eukaryotic ribosomes. Nat. Struct. Mol. Biol. 2012, 19, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Myasnikov, A.G.; Kundhavai Natchiar, S.; Nebout, M.; Hazemann, I.; Imbert, V.; Khatter, H.; Peyron, J.-F.; Klaholz, B.P. Structure–function insights reveal the human ribosome as a cancer target for antibiotics. Nat. Commun. 2016, 7, 12856. [Google Scholar] [CrossRef] [Green Version]
- Khatter, H.; Myasnikov, A.G.; Natchiar, S.K.; Klaholz, B.P. Structure of the human 80S ribosome. Nature 2015, 520, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, A.V.; Razin, S.V.; Spirin, A.S. Intersubunit mobility of the ribosome. Mol. Biol. 2018, 52, 799–811. [Google Scholar] [CrossRef]
- Thomas, G.; Siegmann, M.; Kubler, A.-M.; Gordon, J.; de Asua, L.J. Regulation of 40S ribosomal protein S6 phosphorylation in swiss mouse 3T3 cells. Cell 1980, 19, 1015–1023. [Google Scholar] [CrossRef]
- Shirai, A.; Sadaie, M.; Shinmyozu, K.; Nakayama, J. Methylation of ribosomal protein L42 regulates ribosomal function and stress-adapted cell growth. J. Biol. Chem. 2010, 285, 22448–22460. [Google Scholar] [CrossRef] [Green Version]
- Mazumder, B.; Sampath, P.; Seshadri, V.; Maitra, R.K.; DiCorleto, P.E.; Fox, P.L. Regulated release of L13a from the 60S ribosomal subunit as a mechanism of transcript-specific translational control. Cell 2003, 115, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Ge, W.; Wolf, A.; Feng, T.; Ho, C.; Sekirnik, R.; Zayer, A.; Granatino, N.; Cockman, M.E.; Loenarz, C.; Loik, N.D. Oxygenase-catalyzed ribosome hydroxylation occurs in prokaryotes and humans. Nat. Chem. Biol. 2012, 8, 960–962. [Google Scholar] [CrossRef]
- Singleton, R.S.; Liu-Yi, P.; Formenti, F.; Ge, W.; Sekirnik, R.; Fischer, R.; Adam, J.; Pollard, P.J.; Wolf, A.; Thalhammer, A. OGFOD1 catalyzes prolyl hydroxylation of RPS23 and is involved in translation control and stress granule formation. Proc. Natl. Acad. Sci. USA 2014, 111, 4031–4036. [Google Scholar] [CrossRef] [Green Version]
- Graifer, D.; Malygin, A.; Karpova, G. Hydroxylation of protein constituents of the human translation system: Structural aspects and functional assignments. Future Med. Chem. 2019, 11, 357–369. [Google Scholar] [CrossRef]
- Bräuer, K.E.; Brockers, K.; Moneer, J.; Feuchtinger, A.; Wollscheid-Lengeling, E.; Lengeling, A.; Wolf, A. Phylogenetic and genomic analyses of the ribosomal oxygenases Riox1 (No66) and Riox2 (Mina53) provide new insights into their evolution. BMC Evol. Biol. 2018, 18, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bundred, J.R.; Hendrix, E.; Coleman, M.L. The emerging roles of ribosomal histidyl hydroxylases in cell biology, physiology and disease. Cell. Mol. Life Sci. 2018, 75, 4093–4105. [Google Scholar] [CrossRef] [Green Version]
- Anger, A.M.; Armache, J.-P.; Berninghausen, O.; Habeck, M.; Subklewe, M.; Wilson, D.N.; Beckmann, R. Structures of the human and drosophila 80S ribosome. Nature 2013, 497, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Anthony, R.A.; Liebman, S.W. Alterations in ribosomal protein RPS28 can diversely affect translational accuracy in Saccharomyces Cerevisiae. Genetics 1995, 140, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Cukras, A.R.; Rogers, E.J.; Southworth, D.R.; Green, R. Mutational analysis of S12 protein and implications for the accuracy of decoding by the ribosome. J. Mol. Biol. 2007, 374, 1065–1076. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Su, Z.; Yang, K.; Wu, C.; Becker, T.; Bell-Pedersen, D.; Zhang, J.; Sachs, M.S. Structure of the translating neurospora ribosome arrested by cycloheximide. Proc. Natl. Acad. Sci. 2021, 118, e2111862118. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Li, X.; Nian, W.; Wang, J.; Wang, X.; Sun, L.; Zhu, Y.; Tong, Z. Ribosome proteins represented by RPL27A mark the development and metastasis of triple-negative breast cancer in mouse and human. Front. Cell Dev. Biol. 2021, 9, 716730. [Google Scholar] [CrossRef]
- Alkhatabi, H.A.; McLornan, D.P.; Kulasekararaj, A.G.; Malik, F.; Seidl, T.; Darling, D.; Gaken, J.; Mufti, G.J. RPL27A is a target of MiR-595 and may contribute to the myelodysplastic phenotype through ribosomal dysgenesis. Oncotarget 2016, 7, 47875. [Google Scholar] [CrossRef] [Green Version]
- Joseph, J.P.; Harishankar, M.K.; Pillai, A.A.; Devi, A. Hypoxia induced EMT: A review on the mechanism of tumor progression and metastasis in OSCC. Oral Oncol. 2018, 80, 23–32. [Google Scholar] [CrossRef]
- Mennerich, D.; Kubaichuk, K.; Kietzmann, T. DUBs, hypoxia, and cancer. Trends Cancer 2019, 5, 632–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanshina, D.D.; Gopanenko, A.V.; Karpova, G.G.; Malygin, A.A. Replacement of hydroxylated His39 in ribosomal protein uL15 with Ala or Thr impairs the translational activity of human ribosomes. Mol. Biol. 2020, 54, 449–457. [Google Scholar] [CrossRef]
- Babaylova, E.S.; Gopanenko, A.V.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A.; Karpova, G.G. Deficiency of the ribosomal protein uL5 Leads to significant rearrangements of the transcriptional and translational landscapes in mammalian cells. Int. J. Mol. Sci. 2021, 22, 13485. [Google Scholar] [CrossRef]
- Gopanenko, A.V.; Kolobova, A.V.; Meschaninova, M.I.; Venyaminova, A.G.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A.; Karpova, G.G. Knockdown of the ribosomal protein eL29 in mammalian cells leads to significant changes in gene expression at the transcription level. Cells 2020, 9, 1228. [Google Scholar] [CrossRef] [PubMed]
- Gopanenko, A.V.; Kolobova, A.V.; Meschaninova, M.I.; Venyaminova, A.G.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A.; Karpova, G.G. Knockdown of the mRNA encoding the ribosomal protein eL38 in mammalian cells causes a substantial reorganization of genomic transcription. Biochimie 2021, 184, 132–142. [Google Scholar] [CrossRef]
- Gopanenko, A.V.; Kolobova, A.V.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A.; Karpova, G.G. Knockdown of the ribosomal protein eL38 in HEK293 cells changes the translational efficiency of specific genes. Int. J. Mol. Sci. 2021, 22, 4531. [Google Scholar] [CrossRef]
- Luan, Y.; Tang, N.; Yang, J.; Liu, S.; Cheng, C.; Wang, Y.; Chen, C.; Guo, Y.; Wang, H.; Zhao, W. Deficiency of ribosomal proteins reshapes the transcriptional and translational landscape in human cells. Nucleic Acids Res. 2022, 50, 6601–6617. [Google Scholar] [CrossRef]
- Tian, Y.; Babaylova, E.S.; Gopanenko, A.V.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A.; Karpova, G.G. Changes in the transcriptome caused by mutations in the ribosomal protein uS10 associated with a predisposition to colorectal cancer. Int. J. Mol. Sci. 2022, 23, 6174. [Google Scholar] [CrossRef]
- Natchiar, S.K.; Myasnikov, A.G.; Kratzat, H.; Hazemann, I.; Klaholz, B.P. Visualization of chemical modifications in the human 80S ribosome structure. Nature 2017, 551, 472–477. [Google Scholar] [CrossRef]
- DeLano, W.L. PyMol Molecular Graphics System User’s Manual; DeLano Scientific, Palo Alto: Santa Clara, CA, USA, 2002. [Google Scholar]
- Baliga, B.S.; Pronczuk, A.W.; Munro, H.N. Mechanism of cycloheximide inhibition of protein synthesis in a cell-free system prepared from rat liver. J. Biol. Chem. 1969, 244, 4480–4489. [Google Scholar] [CrossRef]
- Schneider-Poetsch, T.; Ju, J.; Eyler, D.E.; Dang, Y.; Bhat, S.; Merrick, W.C.; Green, R.; Shen, B.; Liu, J.O. Inhibition of eukaryotic translation elongation by cycloheximide and lactimidomycin. Nat. Chem. Biol. 2010, 6, 209–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zolotenkova, E.A.; Gopanenko, A.V.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A. Mutation at the Site of Hydroxylation in the Ribosomal Protein uL15 (RPL27a) Causes Specific Changes in the Repertoire of mRNAs Translated in Mammalian Cells. Int. J. Mol. Sci. 2023, 24, 6173. https://doi.org/10.3390/ijms24076173
Zolotenkova EA, Gopanenko AV, Tupikin AE, Kabilov MR, Malygin AA. Mutation at the Site of Hydroxylation in the Ribosomal Protein uL15 (RPL27a) Causes Specific Changes in the Repertoire of mRNAs Translated in Mammalian Cells. International Journal of Molecular Sciences. 2023; 24(7):6173. https://doi.org/10.3390/ijms24076173
Chicago/Turabian StyleZolotenkova, Elizaveta A., Alexander V. Gopanenko, Alexey E. Tupikin, Marsel R. Kabilov, and Alexey A. Malygin. 2023. "Mutation at the Site of Hydroxylation in the Ribosomal Protein uL15 (RPL27a) Causes Specific Changes in the Repertoire of mRNAs Translated in Mammalian Cells" International Journal of Molecular Sciences 24, no. 7: 6173. https://doi.org/10.3390/ijms24076173
APA StyleZolotenkova, E. A., Gopanenko, A. V., Tupikin, A. E., Kabilov, M. R., & Malygin, A. A. (2023). Mutation at the Site of Hydroxylation in the Ribosomal Protein uL15 (RPL27a) Causes Specific Changes in the Repertoire of mRNAs Translated in Mammalian Cells. International Journal of Molecular Sciences, 24(7), 6173. https://doi.org/10.3390/ijms24076173