
Exposure to Insecticides Modifies Gene Expression and DNA Methylation in Hematopoietic Tissues In Vitro

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
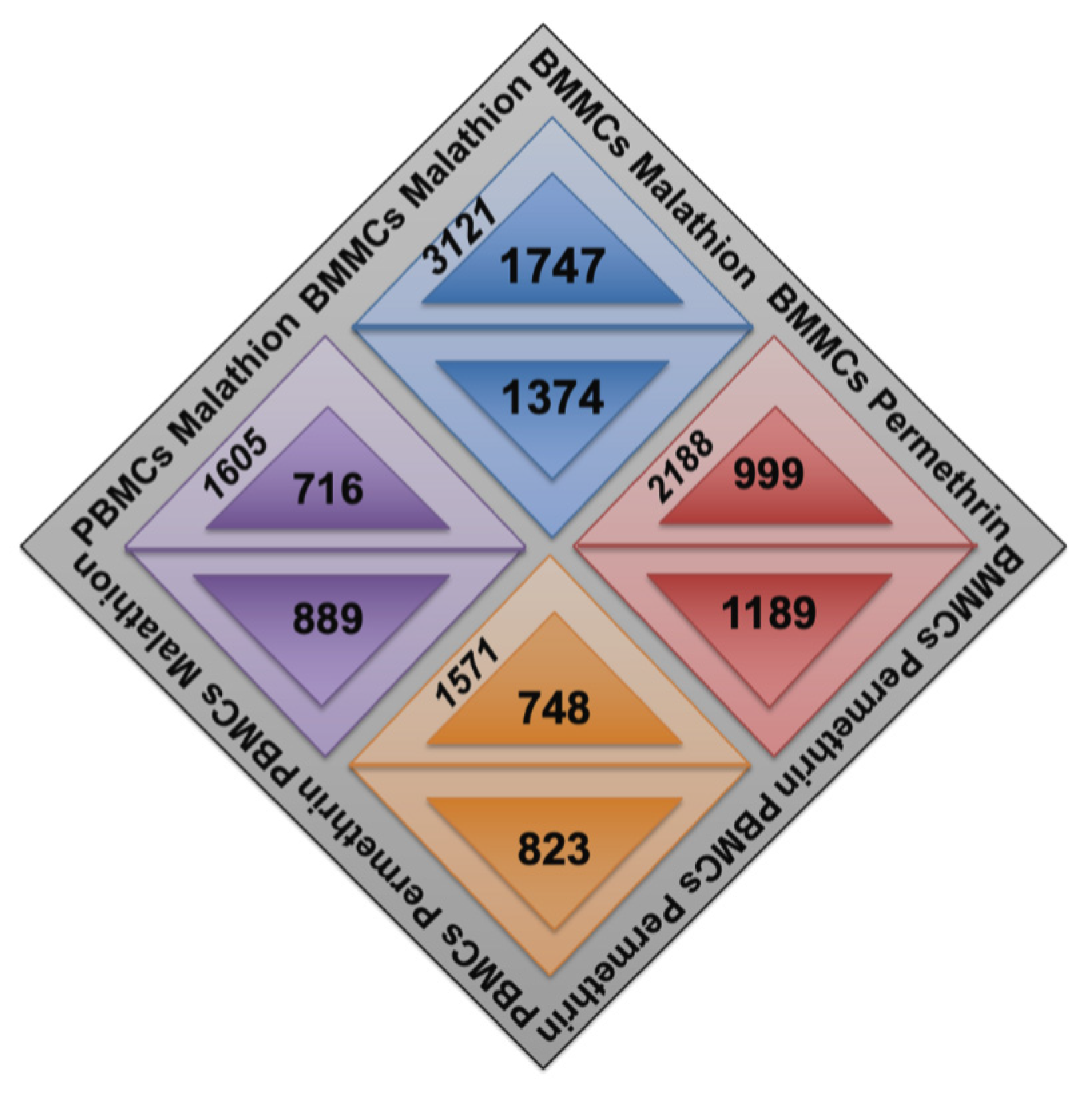
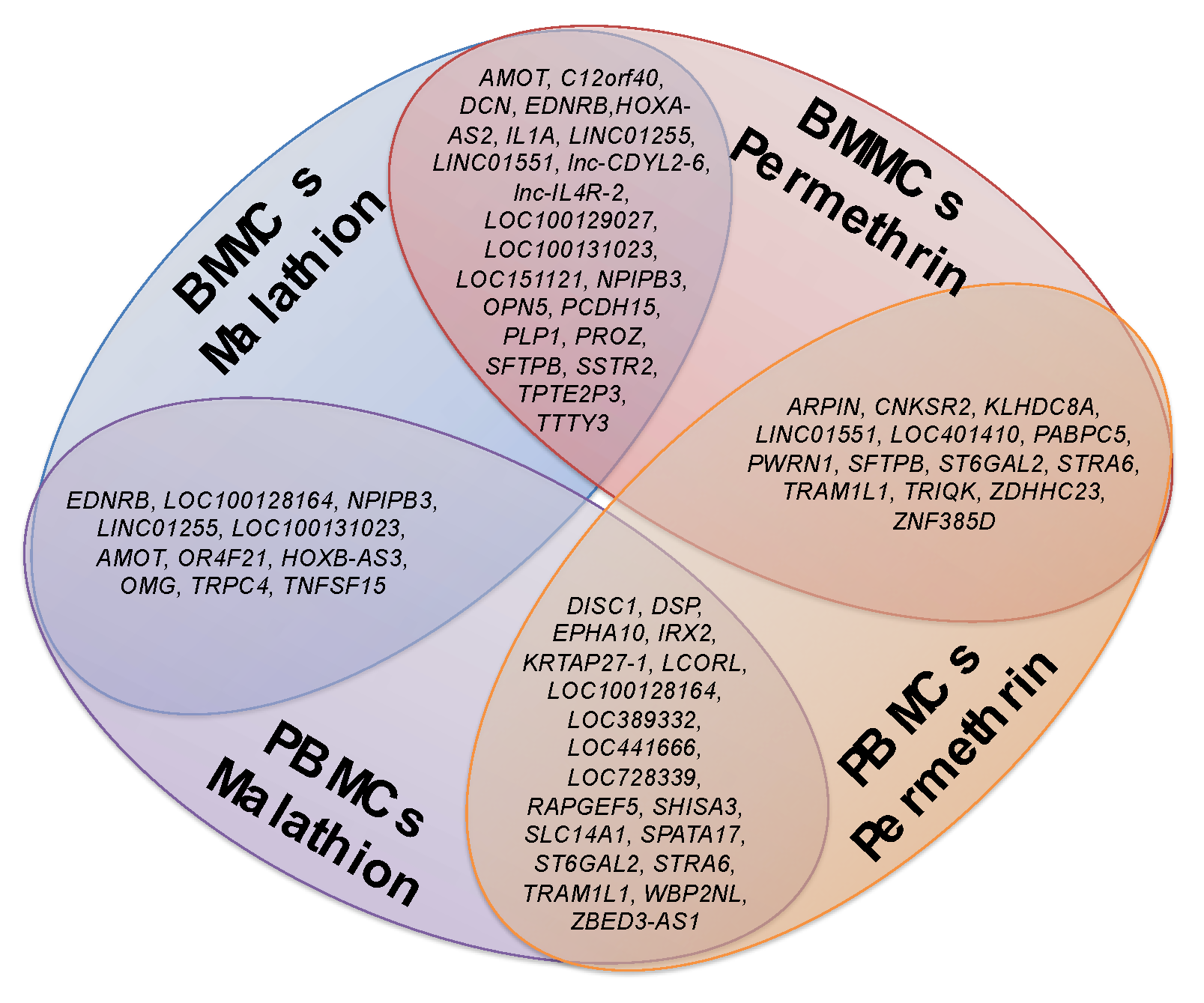
2.1. Changes in Gene Expression of BMMCs and PBMCs Exposed to Insecticides
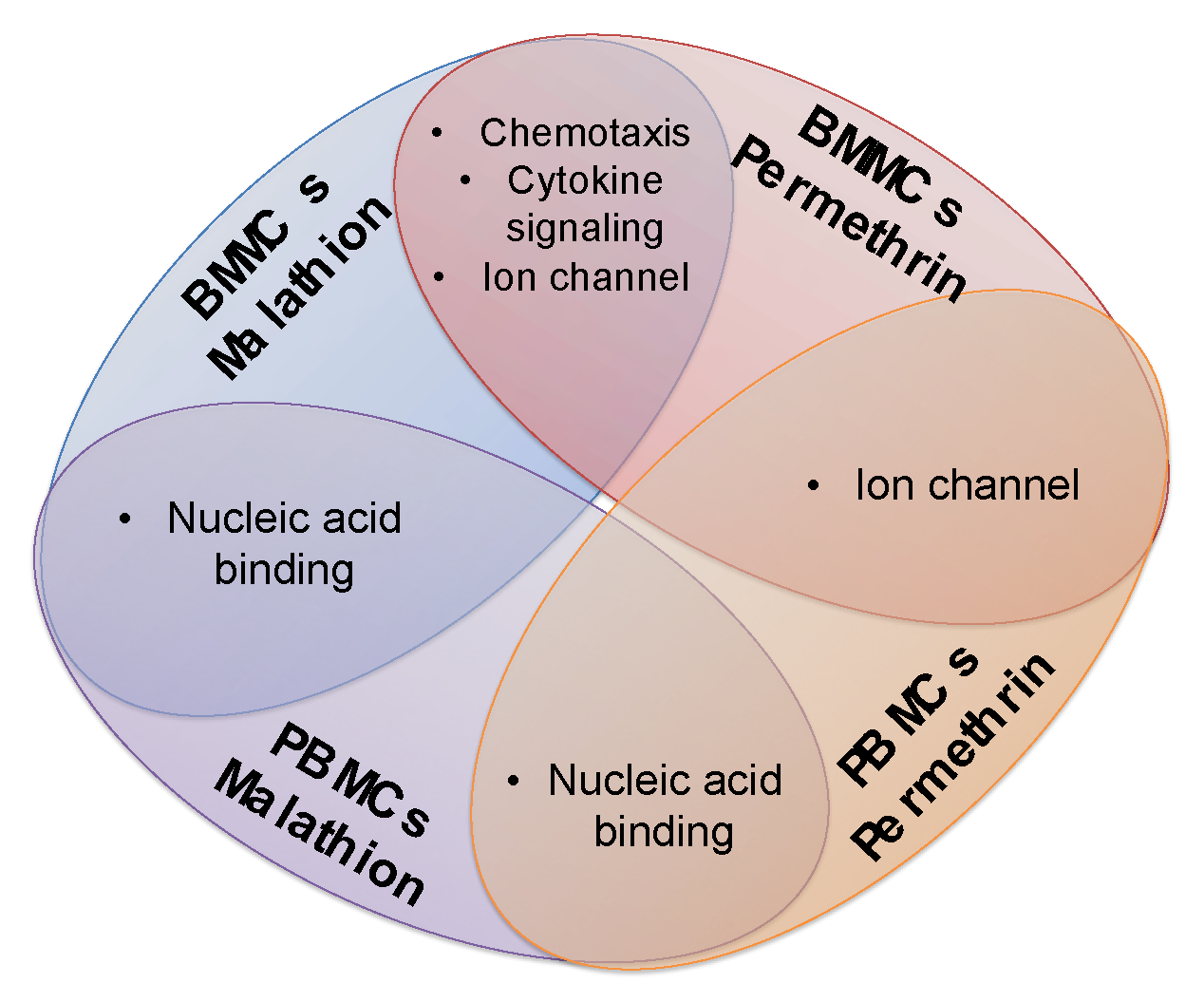
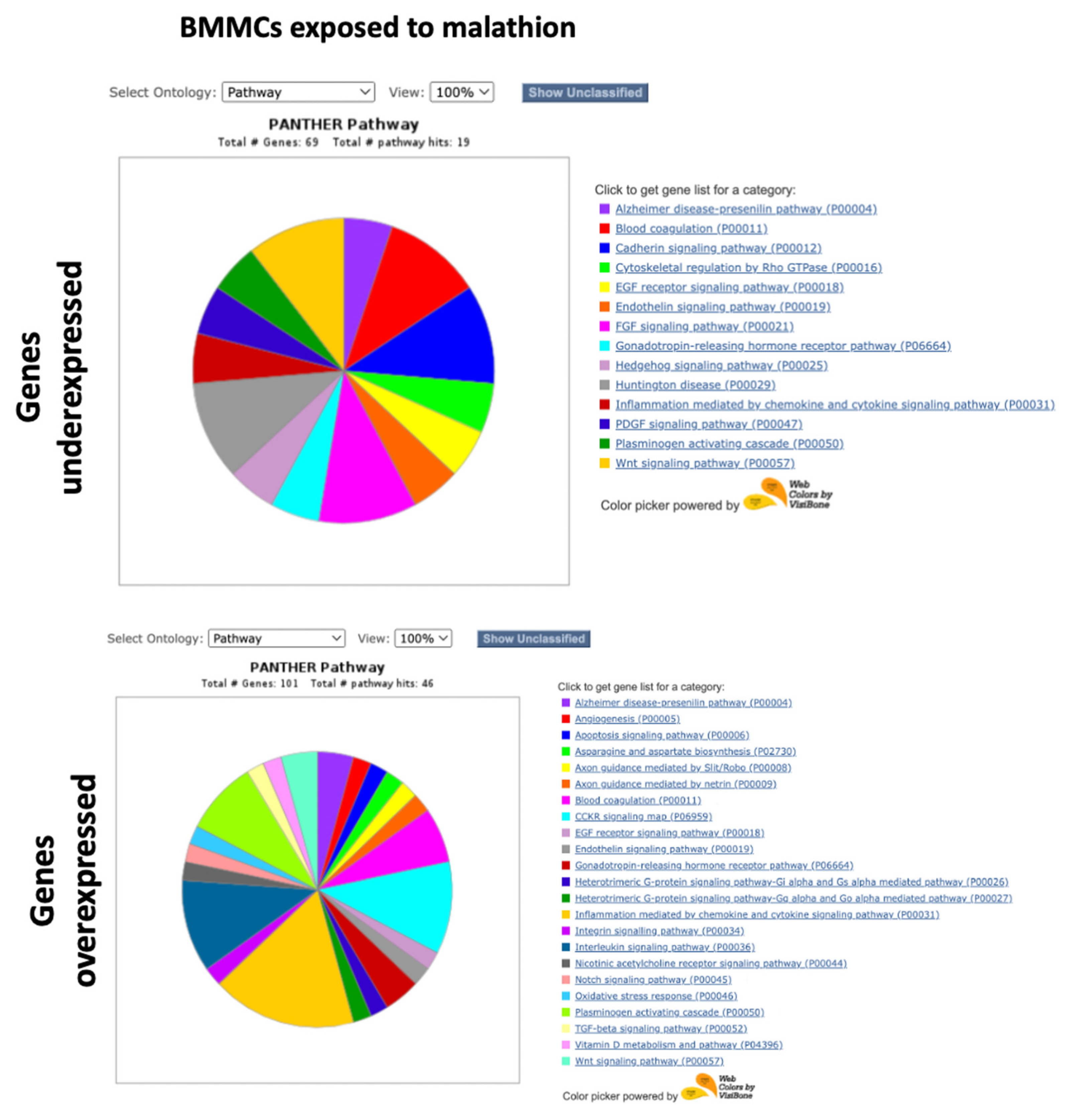
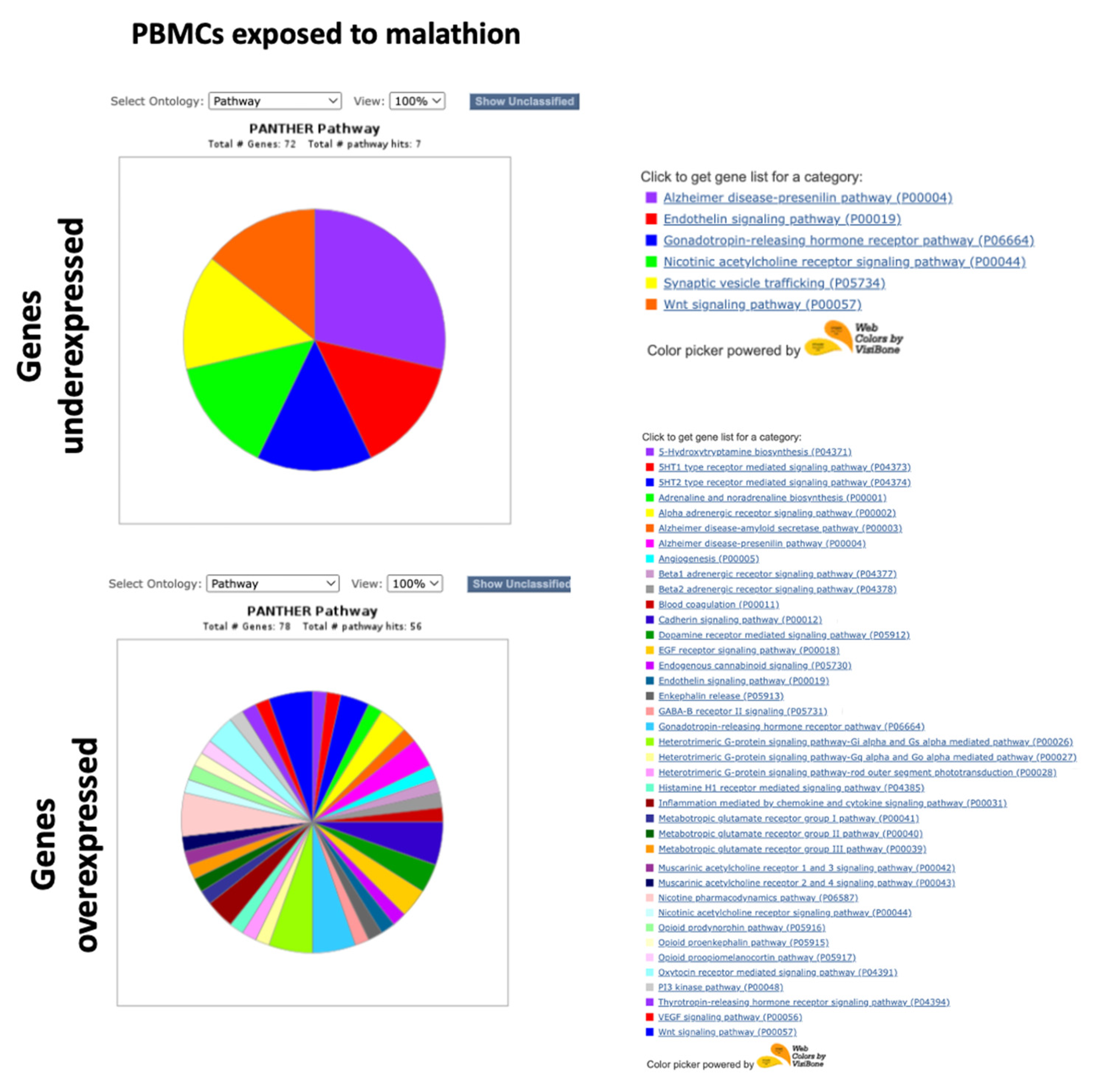
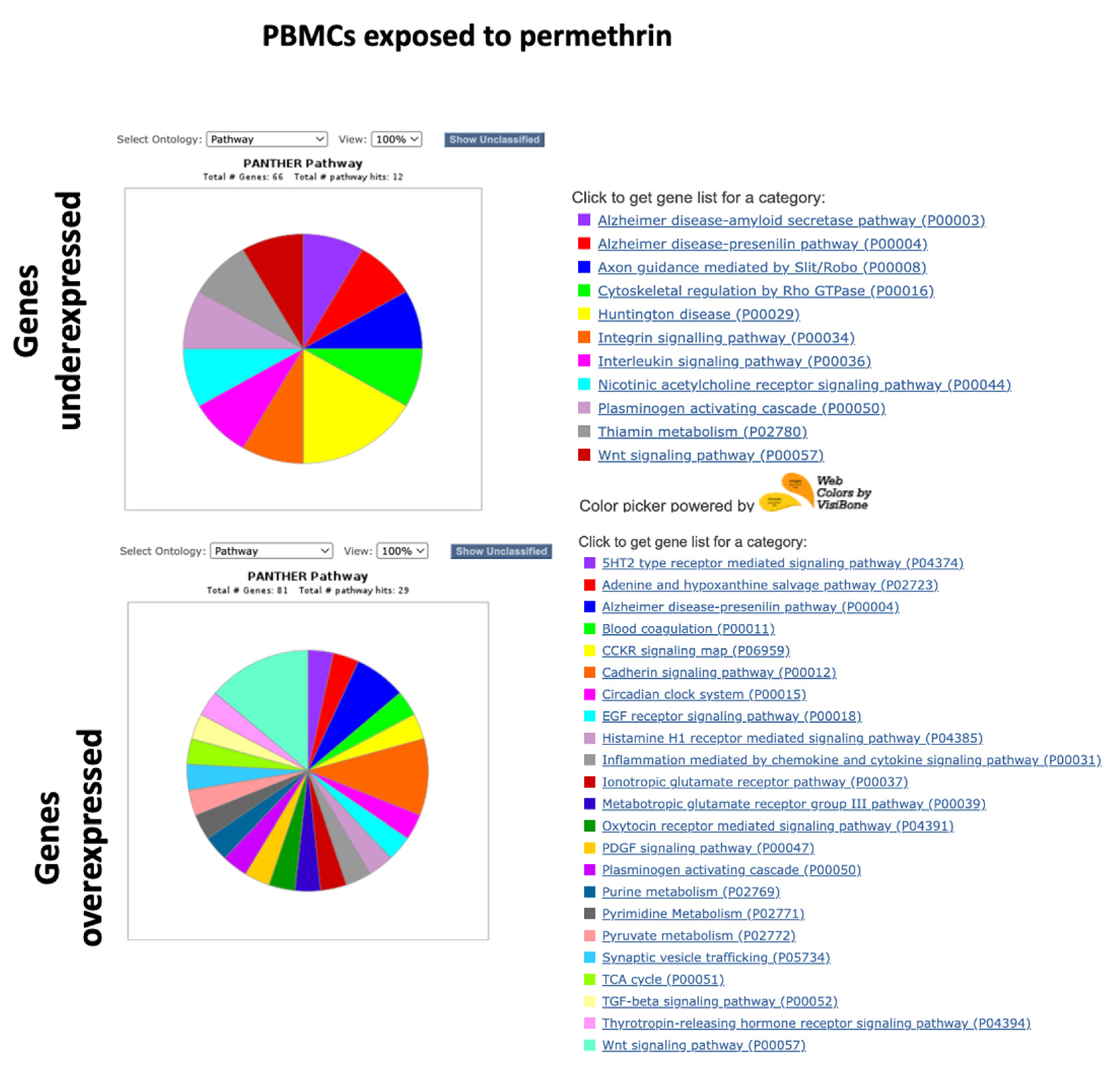
2.2. PANTHER GO Analysis with the Complete Lists of Altered Genes
2.3. PANTHER GO Analysis with the 100 Genes with the Highest Expression and 100 with the Lowest Expression
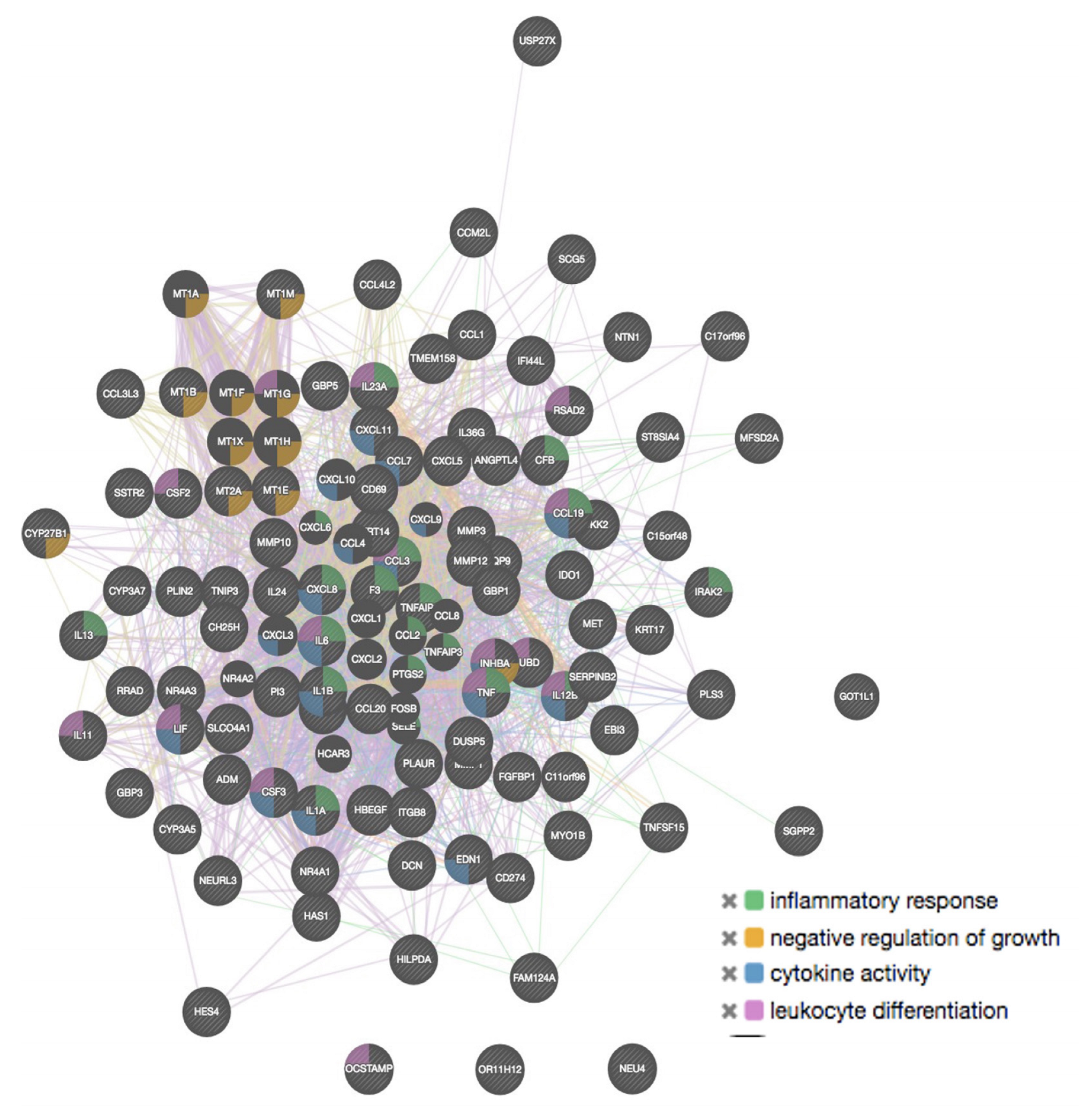
2.4. GeneMANIA Analysis
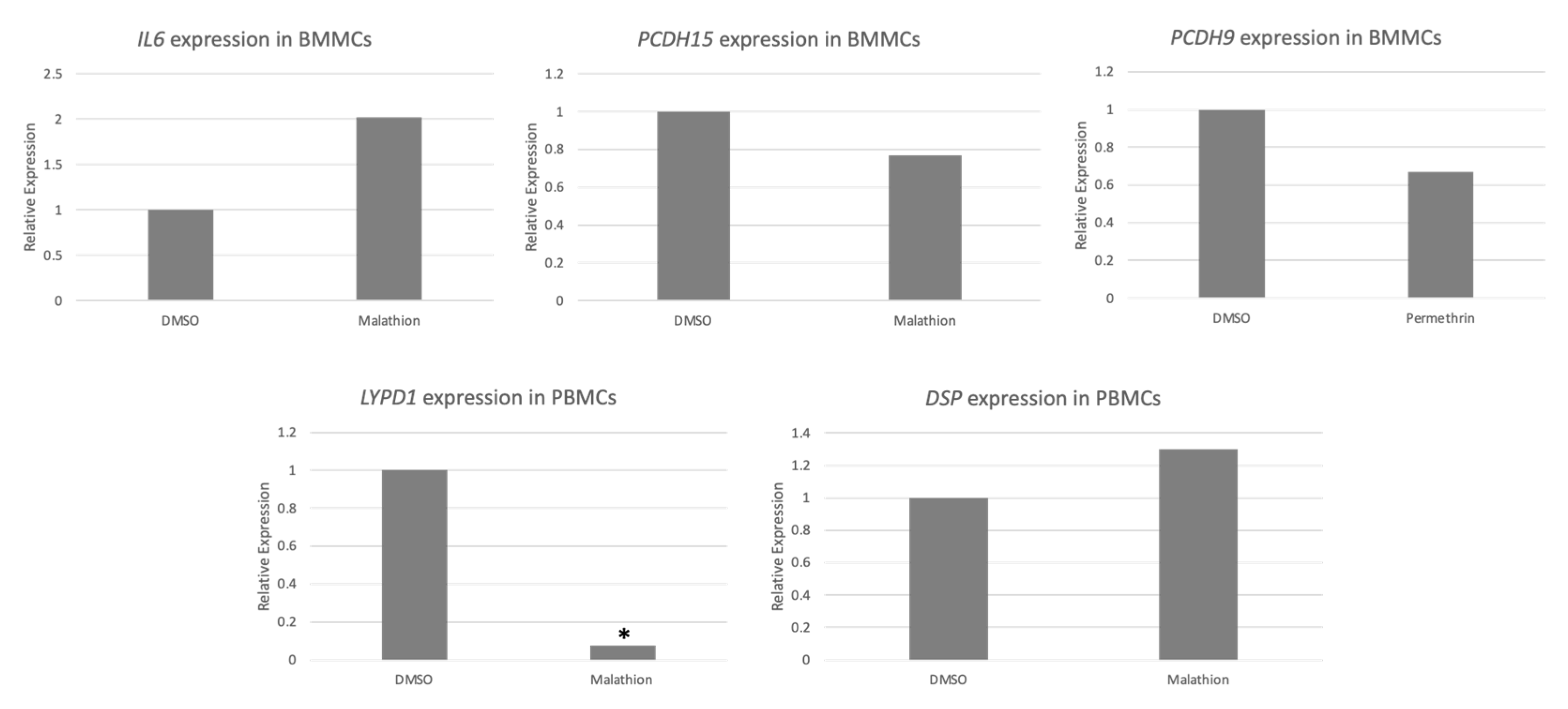
2.5. qPCR Validation
2.6. Changes in DNA Methylation of BMMCs Exposed to Insecticides
3. Discussion
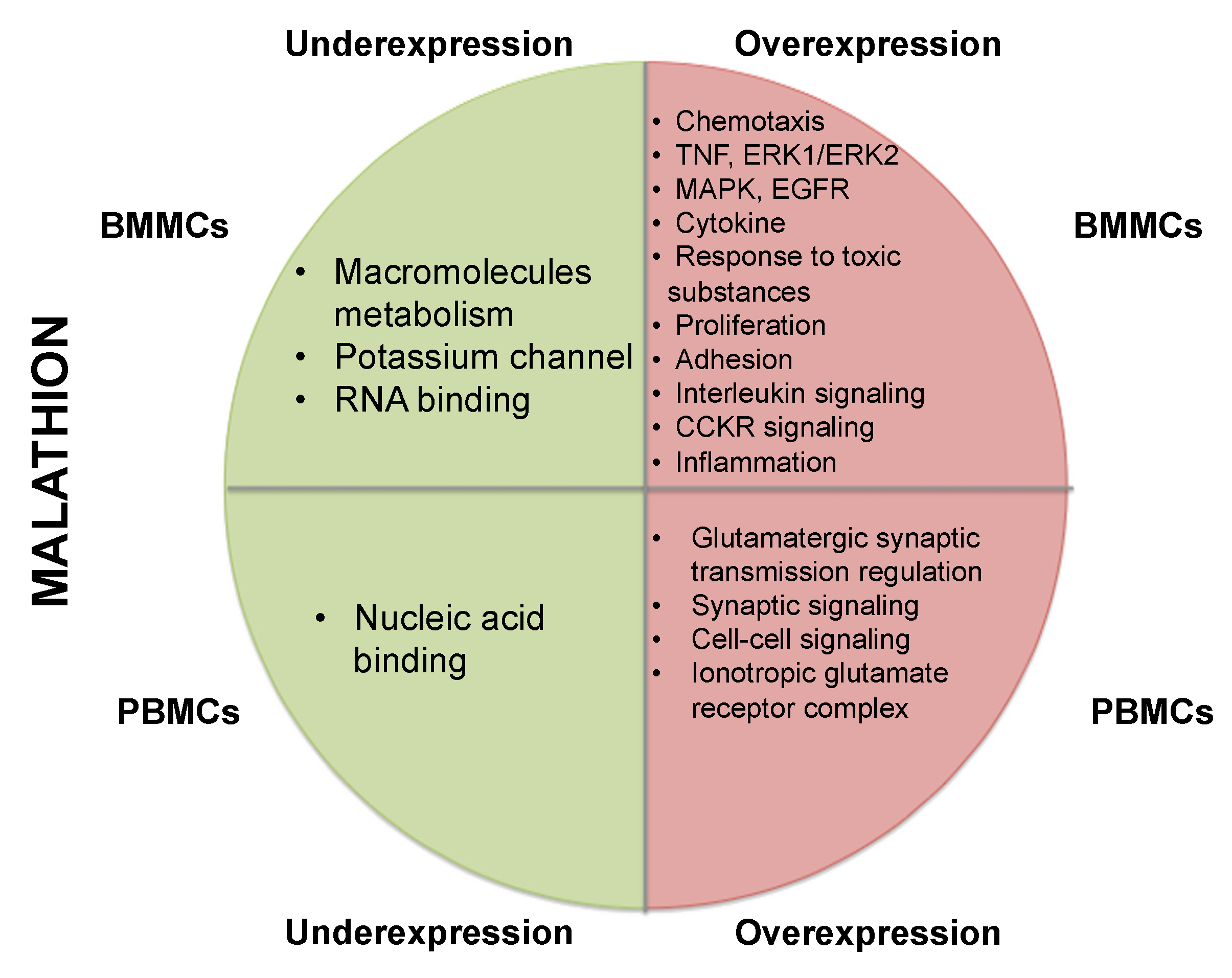
3.1. Underexpression of Genes in BMMCs Exposed to Malathion
3.2. Overexpression of Genes in BMMCs Exposed to Malathion
3.3. Gene Expression Deregulation in PBMCs Exposed to Malathion
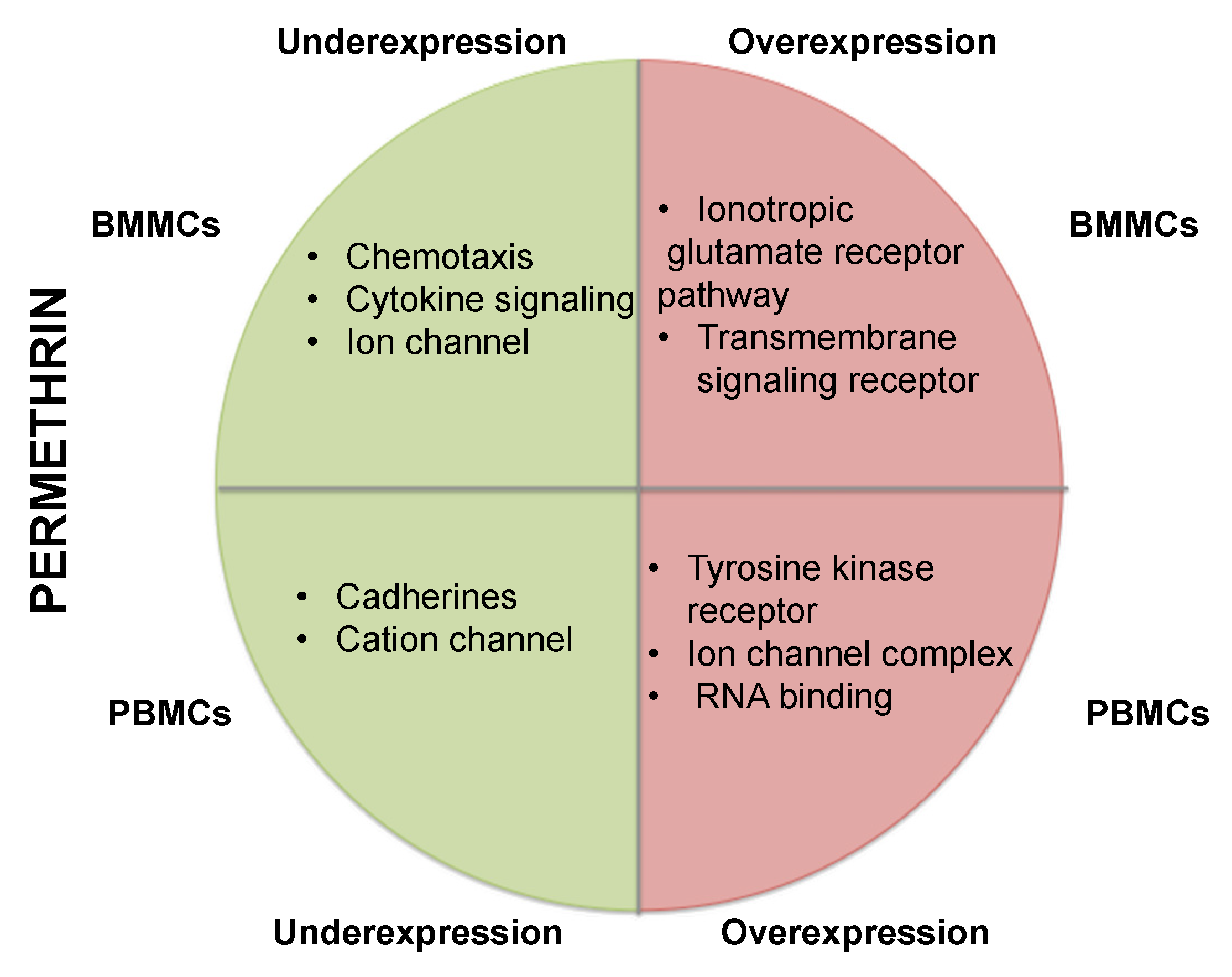
3.4. Gene Repression in BMMCs Exposed to Permethrin
3.5. Gene Overexpression in BMMCs Exposed to Permethrin
3.6. Gene Repression in PBMCs Exposed to Permethrin
3.7. Overexpression of Genes in PMBCs Exposed to Permethrin
3.8. Methylation Alteration in BMMCs Exposed to Insecticides
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Nucleic acid Extraction and Microarrays
4.3. Data Analysis
4.4. qPCR Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matthews, G. Pesticides: Health, Safety and the Environment, 2nd ed.; Wiley: Hoboken, NJ, USA, 2016; Available online: https://www.wiley.com/en-us/Pesticides%3A+Health%2C+Safety+and+the+Environment%2C+2nd+Edition-p-9781118976029 (accessed on 16 December 2022).
- Agopian, J.; Navarro, J.-M.; Gac, A.-C.; Lecluse, Y.; Briand, M.; Grenot, P.; Gauduchon, P.; Ruminy, P.; LeBailly, P.; Nadel, B.; et al. Agricultural pesticide exposure and the molecular connection to lymphomagenesis. J. Exp. Med. 2009, 206, 1473–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, G.; Shi, R.; Gao, Y.; Zhang, Y.; Kamijima, M.; Sakai, K.; Wang, G.; Feng, C.; Tian, Y. Pyrethroid pesticide exposure and risk of childhood acute lymphocytic leukemia in Shanghai. Environ. Sci. Technol. 2012, 46, 13480–13487. [Google Scholar] [CrossRef] [PubMed]
- Menegaux, F.; Baruchel, A.; Bertrand, Y.; Lescoeur, B.; Leverger, G.; Nelken, B.; Sommelet, D.; Hémon, D.; Clavel, J. Household exposure to pesticides and risk of childhood acute leukaemia. Occup. Environ. Med. 2006, 63, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Liu, X.; Liu, C.; Wang, J.; Li, C.; Liu, Q.; Li, Q.; Li, S.; Sun, S.; Yan, J.; et al. Chlorpyrifos induces MLL translocations through caspase 3-dependent genomic instability and topoisomerase II inhibition in human fetal liver hematopoietic stem cells. Toxicol. Sci. Off. J. Soc. Toxicol. 2015, 147, 588–606. [Google Scholar] [CrossRef] [Green Version]
- Alavanja, M.C.R.; Bonner, M.R. Occupational pesticide exposures and cancer risk: A review. J. Toxicol. Environ. Heal. Part B 2012, 15, 238–263. [Google Scholar] [CrossRef]
- Hernández, A.F.; Menéndez, P. Linking pesticide exposure with pediatric leukemia: Potential underlying mechanisms. Int. J. Mol. Sci. 2016, 17, 461. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, H.; Ghassemi-Barghi, N.; Malakshah, O.; Ashari, S. Pyrethroid exposure and neurotoxicity: A mechanistic approach. Arch. Ind. Hyg. Toxicol. 2019, 70, 74–89. [Google Scholar] [CrossRef] [Green Version]
- Naughton, S.X.; Terry, A.V. Neurotoxicity in acute and repeated organophosphate exposure. Toxicology 2018, 408, 101–112. [Google Scholar] [CrossRef]
- Navarrete-Meneses, M.D.P.; Pérez-Vera, P. Pyrethroid pesticide exposure and hematological cancer: Epidemiological, biological and molecular evidence. Rev. Environ. Health 2019, 34, 197–210. [Google Scholar] [CrossRef]
- Perry, J.; Cotton, J.; Rahman, M.A.; A Brumby, S. Organophosphate exposure and the chronic effects on farmers: A narrative review. Rural Remote Health 2020, 20, 4508. [Google Scholar] [CrossRef]
- Fghihi-Zarandi, A.; Dabaghzadeh, F.; Vaziri, A.; Karami-Mohajeri, S.; Ghorbaninejad, B.; Zamani, A.; Rahimi-Sadegh, K. Occupational risk assessment of organophosphates with an emphasis on psychological and oxidative stress factors. Toxicol. Ind. Health 2022, 38, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wallace, A.D.; Du, P.; Kibbe, W.A.; Jafari, N.; Xie, H.; Lin, S.; Baccarelli, A.; Soares, M.B.; Hou, L. DNA methylation alterations in response to pesticide exposure in vitro. Environ. Mol. Mutagen. 2012, 53, 542–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjitha, R.; Antony, A.; Shilpa, O.; Anupama, K.P.; Mallikarjunaiah, S.; Gurushankara, H.P. Malathion induced cancer-linked gene expression in human lymphocytes. Environ. Res. 2020, 182, 109131. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Ramos, E.; Ares, I.; Castellano, V.; Martínez, M.; Martínez-Larrañaga, M.-R.; Anadón, A.; Martínez, M.-A. Oxidative stress and gene expression profiling of cell death pathways in alpha-cypermethrin-treated SH-SY5Y cells. Arch. Toxicol. 2016, 91, 2151–2164. [Google Scholar] [CrossRef] [PubMed]
- Boffetta, P.; Desai, V. Exposure to permethrin and cancer risk: A systematic review. Crit. Rev. Toxicol. 2018, 48, 433–442. [Google Scholar] [CrossRef] [PubMed]
- McDuffie, H.H.; Pahwa, P.; McLaughlin, J.R.; Spinelli, J.J.; Fincham, S.; Dosman, J.A.; Robson, D.; Skinnider, L.F.; Choi, N.W. Non-Hodgkin’s lymphoma and specific pesticide exposures in men: Cross-Canada study of pesticides and health. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored. Am. Soc. Prev. Oncol. 2001, 10, 1155–1163. [Google Scholar]
- de Moura, L.T.R.; Bedor, C.N.G.; Lopez, R.V.M.; Santana, V.S.; de Silviera da Rocha, T.M.B.; Filho, V.W.; Curado, M.P. Occupational exposure to organophosphate pesticides and hematologic neoplasms: A systematic review. Rev. Bras. Epidemiol. Braz. J. Epidemiol. 2020, 23, e200022. [Google Scholar]
- Rodriguez-Cortez, V.C.; Navarrete-Meneses, M.P.; Molina, O.; Velasco-Hernandez, T.; Gonzalez, J.; Romecin, P.; Gutierrez-Agüera, F.; Roca-Ho, H.; Vinyoles, M.; Kowarz, E.; et al. The insecticides permethrin and chlorpyrifos show limited genotoxicity and no leukemogenic potential in human and murine hematopoietic stem progenitor cells. Haematologica 2022, 107, 544–559. [Google Scholar] [CrossRef]
- Navarrete-Meneses, M.P.; Pedraza-Meléndez, A.I.; Salas-Labadía, C.; Moreno-Lorenzana, D.; Pérez-Vera, P. Low concentrations of permethrin and malathion induce numerical and structural abnormalities in KMT2A and IGH genes in vitro. J. Appl. Toxicol. 2018, 38, 1262–1270. [Google Scholar] [CrossRef]
- Hoang, T.T.; Qi, C.; Paul, K.C.; Lee, M.; White, J.D.; Richards, M.; Auerbach, S.S.; Long, S.; Shrestha, S.; Wang, T.; et al. Epigenome-wide DNA methylation and pesticide use in the agricultural lung health study. Environ. Health Perspect. 2021, 129, 097008. [Google Scholar] [CrossRef]
- Paul, K.C.; Chuang, Y.-H.; Cockburn, M.; Bronstein, J.M.; Horvath, S.; Ritz, B. Organophosphate pesticide exposure and differential genome-wide DNA methylation. Sci. Total Environ. 2018, 645, 1135–1143. [Google Scholar] [CrossRef]
- Costa, M.B.; Farias, I.R.; da Silva, C.M.; Filho, L.I.P.F.; Borges, D.D.P.; de Oliveira, R.T.G.; Ribeiro-Junior, H.L.; Magalhães, S.M.M.; Pinheiro, R.F. Chromosomal abnormalities and dysregulated DNA repair gene expression in farmers exposed to pesticides. Environ. Toxicol. Pharmacol. 2020, 82, 103564. [Google Scholar] [CrossRef]
- Giambò, F.; Leone, G.M.; Gattuso, G.; Rizzo, R.; Cosentino, A.; Cinà, D.; Teodoro, M.; Costa, C.; Tsatsakis, A.; Fenga, C.; et al. Genetic and epigenetic alterations induced by pesticide exposure: Integrated analysis of gene expression, microRNA expression, and DNA methylation datasets. Int. J. Environ. Res. Public Health 2021, 18, 8697. [Google Scholar] [CrossRef]
- Wajed, S.A.; Laird, P.W.; Demeester, T.R. DNA methylation: An alternative pathway to cancer. Ann. Surg. 2001, 234, 10–20. [Google Scholar] [CrossRef]
- Mandarapu, R.; Prakhya, B.M. Exposure to cypermethrin and mancozeb alters the expression profile of THBS1, SPP1, FEZ1 and GPNMB in human peripheral blood mononuclear cells. J. Immunotoxicol. 2016, 13, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Collotta, M.; Bertazzi, P.; Bollati, V. Epigenetics and pesticides. Toxicology 2013, 307, 35–41. [Google Scholar] [CrossRef]
- Bordoni, L.; Nasuti, C.; Mirto, M.; CaraDonna, F.; Gabbianelli, R. Intergenerational effect of early life exposure to permethrin: Changes in global DNA methylation and in Nurr1 gene expression. Toxics 2015, 3, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-C.T.; Schmitt, M.; Yang, Z.; Que, L.G.; Stewart, J.C.; Frampton, M.W.; Devlin, R.B. Gene expression profile in circulating mononuclear cells after exposure to ultrafine carbon particles. Inhal. Toxicol. 2010, 22, 835–846. [Google Scholar] [CrossRef] [Green Version]
- Wens, B.; De Boever, P.; Verbeke, M.; Hollanders, K.; Schoeters, G. Cultured human peripheral blood mononuclear cells alter their gene expression when challenged with endocrine-disrupting chemicals. Toxicology 2013, 303, 17–24. [Google Scholar] [CrossRef]
- Gangemi, S.; Gofita, E.; Costa, C.; Teodoro, M.; Briguglio, G.; Nikitovic, D.; Tzanakakis, G.; Tsatsakis, A.M.; Wilks, M.F.; Spandidos, D.A.; et al. Occupational and environmental exposure to pesticides and cytokine pathways in chronic diseases. Int. J. Mol. Med. 2016, 38, 1012–1020. Available online: https://pubmed.ncbi.nlm.nih.gov/27600395/ (accessed on 16 December 2022). [CrossRef] [Green Version]
- Yang, J.S.; Symington, S.; Clark, J.M.; Park, Y. Permethrin, a pyrethroid insecticide, regulates ERK1/2 activation through membrane depolarization-mediated pathway in HepG2 hepatocytes. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 121, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-B.; Kim, S.-J.; Kang, S.-Y.; Yi, J.W.; Kim, S.-M. The large-conductance calcium-activated potassium channel holds the key to the conundrum of familial hypokalemic periodic paralysis. Korean J. Pediatr. 2014, 57, 445–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, D.R.M.M. Reduced levels of serum potassium and plasma cholinesterase in acute organophosphate poisoning: Possible predictive markers. Asia Pac. J. Med. Toxicol. 2014, 3, 68–72. [Google Scholar]
- Sieuwerts, A.M.; Gelder, M.E.M.-V.; Timmermans, M.; Trapman, A.M.; Garcia, R.R.; Arnold, M.; Goedheer, A.J.; Portengen, H.; Klijn, J.G.; Foekens, J.A. How ADAM-9 and ADAM-11 differentially from estrogen receptor predict response to tamoxifen treatment in patients with recurrent breast cancer: A retrospective study. Clin. Cancer Res. 2005, 11, 7311–7321. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, M.E.; Chen, S.-C.; Andersson, A.K.; Phillips, L.A.; Li, Y.; Sotzen, J.; Kundu, M.; Downing, J.R.; Melnick, A.; Mullighan, C.G. Integrated genetic and epigenetic analysis of childhood acute lymphoblastic leukemia. J. Clin. Investig. 2013, 123, 3099–3111. [Google Scholar] [CrossRef] [Green Version]
- Slotkin, T.A.; Seidler, F.J.; Fumagalli, F. Exposure to organophosphates reduces the expression of neurotrophic factors in neonatal rat brain regions: Similarities and differences in the effects of chlorpyrifos and diazinon on the fibroblast growth factor superfamily. Environ. Health Perspect. 2007, 115, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, P.-C.; Liu, M.-C.; Chen, L.-M.; Tsai, C.-Y.; Wang, Y.-T.; Chen, J.; Hsu, L.-S. Promoter methylation of p16 and EDNRB gene in leukemia patients in Taiwan. Chin. J. Physiol. 2008, 51, 27–31. [Google Scholar]
- Taghavian, F.; Vaezi, G.; Abdollahi, M.; Malekirad, A.A. Comparative toxicological study between exposed and non-exposed farmers to organophosphorus pesticides. Cell J. 2016, 18, 89–96. [Google Scholar]
- Banks, C.N.; Lein, P.J. A review of experimental evidence linking neurotoxic organophosphorus compounds and inflammation. Neurotoxicology 2012, 33, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Hemmati, S.; Haque, T.; Gritsman, K. Inflammatory signaling pathways in preleukemic and leukemic stem cells. Front. Oncol. 2017, 7, 265. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Li, Z.; Zhou, J. Tumor necrosis factor α in the onset and progression of leukemia. Exp. Hematol. 2017, 45, 17–26. [Google Scholar] [CrossRef]
- Navarrete-Meneses, M.D.P.; Pérez-Vera, P. Epigenetic alterations in acute lymphoblastic leukemia. Bol. Med. Hosp. Infant. Mex. 2017, 74, 243–264. [Google Scholar] [CrossRef]
- Knight, T.; Irving, J.A.E. Ras/Raf/MEK/ERK pathway activation in childhood acute lymphoblastic leukemia and its therapeutic targeting. Front. Oncol. 2014, 4, 160. [Google Scholar] [CrossRef]
- Nwabo Kamdje, A.H.; Krampera, M. Notch signaling in acute lymphoblastic leukemia: Any role for stromal microenvironment? Blood 2011, 118, 6506–6514. [Google Scholar] [CrossRef] [Green Version]
- Bubici, C.; Papa, S. JNK signalling in cancer: In need of new, smarter therapeutic targets. Br. J. Pharmacol. 2013, 171, 24–37. [Google Scholar] [CrossRef]
- Staal, F.J.T.; Famili, F.; Perez, L.G.; Pike-Overzet, K. Aberrant Wnt signaling in leukemia. Cancers 2016, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Kordes, U.; Krappmann, D.; Heissmeyer, V.; Ludwig, W.D.; Scheidereit, C. Transcription factor NF-kappaB is constitutively activated in acute lymphoblastic leukemia cells. Leukemia 2000, 14, 399–402. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, D.R.; Seo, K.-W.; Roh, K.-H.; Jung, J.-W.; Kang, S.-K.; Kang, K.-S. REX-1 expression and p38 MAPK activation status can determine proliferation/differentiation fates in human mesenchymal stem cells. PLoS ONE 2010, 5, e10493. [Google Scholar] [CrossRef] [Green Version]
- Kocabas, F.; Zheng, J.; Thet, S.; Copeland, N.G.; Jenkins, N.A.; DeBerardinis, R.J.; Zhang, C.; Sadek, H.A. Meis1 regulates the metabolic phenotype and oxidant defense of hematopoietic stem cells. Blood 2012, 120, 4963–4972. [Google Scholar] [CrossRef] [Green Version]
- Soltaninejad, K.; Abdollahi, M. Current opinion on the science of organophosphate pesticides and toxic stress: A systematic review. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2009, 15, RA75–RA90. [Google Scholar]
- Sturgill, J.L.; Mathews, J.; Scherle, P.; Conrad, D.H. Glutamate signaling through the kainate receptor enhances human immunoglobulin production. J. Neuroimmunol. 2011, 233, 80–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Takano, N.; Xiang, L.; Gilkes, D.M.; Luo, W.; Semenza, G.L. Hypoxia-inducible factors enhance glutamate signaling in cancer cells. Oncotarget 2014, 5, 8853–8868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grainger, S.; Traver, D.; Willert, K. Wnt signaling in hematological malignancies. Prog. Mol. Biol. Transl. Sci. 2018, 153, 321–341. [Google Scholar] [PubMed]
- Dwyer, M.A.; Joseph, J.D.; Wade, H.E.; Eaton, M.L.; Kunder, R.S.; Kazmin, D. WNT11 expression is induced by estrogen-related receptor alpha and beta-catenin and acts in an autocrine manner to increase cancer cell migration. Cancer Res. 2010, 70, 9298–9308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat. Rev. Immunol. 2017, 17, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Rose, T.M.; Bruce, A.G. Oncostatin M is a member of a cytokine family that includes leukemia-inhibitory factor, granulocyte colony-stimulating factor, and interleukin 6. Proc. Natl. Acad. Sci. USA 1991, 88, 8641–8645. [Google Scholar] [CrossRef] [Green Version]
- Dong, M.; Blobe, G.C. Role of transforming growth factor-beta in hematologic malignancies. Blood 2006, 107, 4589–4596. [Google Scholar] [CrossRef] [Green Version]
- Seth, B.; Yadav, A.; Agarwal, S.; Tiwari, S.K.; Chaturvedi, R.K. Inhibition of the transforming growth factor-β/SMAD cascade mitigates the anti-neurogenic effects of the carbamate pesticide carbofuran. J. Biol. Chem. 2017, 292, 19423–19440. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, Q.; Cai, J.; Yang, J.; Shen, Q.; Xu, S. Chlorpyrifos exposure in common carp (Cyprinus carpio L.) leads to oxidative stress and immune responses. Fish Shellfish. Immunol. 2017, 67, 604–611. [Google Scholar] [CrossRef]
- Tao, Y.-F.; Fang, F.; Hu, S.-Y.; Lu, J.; Cao, L.; Zhao, W.-L.; Xiao, P.-F.; Li, Z.-H.; Wang, N.-N.; Xu, L.-X.; et al. Hypermethylation of the GATA binding protein 4 (GATA4) promoter in Chinese pediatric acute myeloid leukemia. BMC Cancer 2015, 15, 756. [Google Scholar] [CrossRef] [Green Version]
- Soderlund, D.M. Molecular mechanisms of pyrethroid insecticide neurotoxicity: Recent advances. Arch. Toxicol. 2012, 86, 165–181. [Google Scholar] [CrossRef] [Green Version]
- Raymond, V.; Matsuda, K.; Sattelle, B.M.; Rauh, J.J.; Sattelle, D.B. Ion channels: Molecular targets of neuroactive insecticides. Invertebr. Neurosci. 2005, 5, 119–133. [Google Scholar] [CrossRef]
- Alvarado, S.; Wyglinski, J.; Suderman, M.; Andrews, S.A.; Szyf, M. Methylated DNA binding domain protein 2 (MBD2) coordinately silences gene expression through activation of the MicroRNA hsa-mir-496 promoter in breast cancer cell line. PLoS ONE 2013, 8, e74009. [Google Scholar] [CrossRef] [Green Version]
- Sarma, S.N.; Kim, Y.-J.; Ryu, J.-C. Gene expression profiles of human promyelocytic leukemia cell lines exposed to volatile organic compounds. Toxicology 2010, 271, 122–130. [Google Scholar] [CrossRef]
- Vainchenker, W.; Constantinescu, S.N. JAK/STAT signaling in hematological malignancies. Oncogene 2012, 32, 2601–2613. [Google Scholar] [CrossRef] [Green Version]
- Pidugu, V.K.; Pidugu, H.B.; Wu, M.-M.; Liu, C.-J.; Lee, T.-C. Emerging functions of human IFIT proteins in cancer. Front. Mol. Biosci. 2019, 6, 148. [Google Scholar] [CrossRef] [Green Version]
- Seth, C.; Mas, C.; Conod, A.; Mueller, J.; Siems, K.; Kuciak, M.; Borges, I.; Altaba, A.R. Long-lasting WNT-TCF response blocking and epigenetic modifying activities of withanolide F in human cancer cells. PLoS ONE 2016, 11, e0168170. [Google Scholar] [CrossRef] [Green Version]
- Kasat, K.; Go, V.; Pogo, B.G. Effects of pyrethroid insecticides and estrogen on WNT10B proto-oncogene expression. Environ. Int. 2002, 28, 429–432. [Google Scholar] [CrossRef]
- El Hajj, N.; Dittrich, M.; Haaf, T. Epigenetic dysregulation of protocadherins in human disease. Semin. Cell Dev. Biol. 2017, 69, 172–182. [Google Scholar] [CrossRef]
- Salameh, A.; Lee, A.K.; Cardó-Vila, M.; Nunes, D.N.; Efstathiou, E.; Staquicini, F.I.; Dobroff, A.S.; Marchiò, S.; Navone, N.M.; Hosoya, H.; et al. PRUNE2 is a human prostate cancer suppressor regulated by the intronic long noncoding RNA PCA3. Proc. Natl. Acad. Sci. USA 2015, 112, 8403–8408. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.X.; Tan, M.S.; Yu, J.T.; Tan, L. Matrix metalloproteinases and their multiple roles in Alzheimer’s Disease. BioMed Res. Int. 2014, 2014, 908636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijer, M.; Dingemans, M.M.; Berg, M.V.D.; Westerink, R.H. Inhibition of voltage-gated calcium channels as common mode of action for (Mixtures of) distinct classes of insecticides. Toxicol. Sci. 2014, 141, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.I.; Lin, D.T.; Chang, C.J.; Lee, C.Y.; Tang, J.L.; Tien, H.F. Marrow matrix metalloproteinases (MMPs) and tissue inhibitors of MMP in acute leukaemia: Potential role of MMP-9 as a surrogate marker to monitor leukaemic status in patients with acute myelogenous leukaemia. Br. J. Haematol. 2002, 117, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Nath, S.; Mandal, C.; Chatterjee, U.; Mandal, C. Association of cytosolic sialidase Neu2 with plasma membrane enhances Fas-mediated apoptosis by impairing PI3K-Akt/mTOR-mediated pathway in pancreatic cancer cells. Cell Death Dis. 2018, 9, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cilloni, D.; Carturan, S.; Bracco, E.; Campia, V.; Rosso, V.; Torti, D.; Calabrese, C.; Gaidano, V.; Niparuck, P.; Favole, A.; et al. Aberrant activation of ROS1 represents a new molecular defect in chronic myelomonocytic leukemia. Leuk. Res. 2013, 37, 520–530. [Google Scholar] [CrossRef]
- Abbaspour Babaei, M.; Kamalidehghan, B.; Saleem, M.; Huri, H.Z.; Ahmadipour, F. Receptor tyrosine kinase (c-Kit) inhibitors: A potential therapeutic target in cancer cells. Drug Des. Dev. Ther. 2016, 10, 2443–2459. [Google Scholar] [CrossRef] [Green Version]
- Holla, V.R.; Elamin, Y.Y.; Bailey, A.M.; Johnson, A.M.; Litzenburger, B.C.; Khotskaya, Y.B.; Sanchez, N.S.; Zeng, J.; Shufean, A.; Shaw, K.R.; et al. ALK: A tyrosine kinase target for cancer therapy. Mol. Case Stud. 2017, 3, a001115. [Google Scholar] [CrossRef] [Green Version]
- Rusiecki, J.A.; Freeman, L.E.B.; Bonner, M.R.; Alexander, M.; Chen, L.; Andreotti, G.; Barry, K.H.; Moore, L.E.; Byun, H.-M.; Kamel, F.; et al. High pesticide exposure events and DNA methylation among pesticide applicators in the agricultural health study. Environ. Mol. Mutagen. 2016, 58, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Agrahari, A.; Singh, R.; Yadav, S.; Srivastava, V.; Parmar, D. Imprinting of cerebral cytochrome P450s in offsprings prenatally exposed to cypermethrin augments toxicity on rechallenge. Sci. Rep. 2016, 6, 37426. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.-K. Active DNA demethylation mediated by DNA glycosylases. Annu. Rev. Genet. 2009, 43, 143–166. [Google Scholar] [CrossRef] [Green Version]
- Two-Color Microarray-Based Gene Expression Analysis Low Input Quick Amp Labeling Protocol. Available online: https://www.agilent.com/cs/library/usermanuals/public/G4140-90050_GeneExpression_TwoColor_6.9.pdf (accessed on 16 December 2022).
- Agilent Microarray Analysis of Methylated DNA Immunoprecipitation Protocol. Available online: https://www.agilent.com/cs/library/usermanuals/public/G4170-90012_Methylation_2.3.pdf (accessed on 16 December 2022).
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2018, 47, D419–D426. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BMMCs Exposed to Malathion | ||
|---|---|---|
| Underexpression | Overexpression | |
| Molecular Function | Structural, cytoskeleton: TUBB8 | Cytokines: CCL3, CCL7, CCL19, CCL1, CCL3L3, CCL20 |
| Cytokine receptor binding: IL23A | ||
| Hormonal receptor binding: ADM, SCG5, EDN1 | ||
| Biological process | Cell growth: LINGO1, LRIG3 | Negative regulation of apoptosis: MET |
| Cell differentiation: PLP1 | ||
| Cell death: ADAM11, ADAM32 | ||
| Mesoderm development: PITX1, FGF22 | ||
| Stimulus response: OPN5, LINGO1, LRIG3, DDR1, OR4D1, CEACAM7, OR2AT4, OR4F29, PROZ, OR4F21, AMOT, OR6C2, OR9G4 | ||
| Protein class | Defense/Immunity: CRISP3 | Adhesion: ITGB8, ANGPTL4 |
| Growth factor: INHBA, HBEGF | ||
| Signaling: PF4V1, FGG | Chemokines: CXCL8, CCL19, CXCL5, CXCL11 | |
| Transcription factor: PROX2, NFIB | Tumoral necrosis factor family: TNF, TNFSF15 | |
| Transcription factor: HES4, NR4A3, NR4A1 | ||
| PBMCs Exposed to Malathion | ||
|---|---|---|
| Underexpression | Overexpression | |
| Molecular Function | Methyltransferase: HNMT | Protein kinase: CDK15, GRK1 |
| Biological Process | Cell cycle: S100A1, CROCCP3 | Negative regulation of apoptosis: EPHA10, FLT4, ERBB4 |
| Development differentiation: GATA4, LEP1 | Epigenetic regulation: FAM172BP, MBD2 | |
| Immune response: CMA1 | Signals transduction: RAPGEF5 | |
| Stress response: APLF, PDIA2 | Differentiation during development: GP5 | |
| Embryonic development: DLG2 | ||
| DNA replication: GMNC | ||
| Cell component | Intracellular junctions: AMOT | Nucleus: TCF7L1 |
| Microtubules: KIF6 | ||
| Protein class | Cytokines: TNFSF15, SST | Adhesion: RS1 |
| Nuclease: TDRD6, TDRD1 | ||
| Transcription factor: IRX2, ZFPM2 | ||
| Transcription Factor: MEIS1, ARNT2, ZFP42, WBP2NL, EN1 | ||
| Pathway | Endothelin: EDNRB | Inflammation: PLCB4, GNAI1 |
| Wnt: PCDHGB1 | ||
| BMMCs Exposed to Permethrin | ||
|---|---|---|
| Underexpression | Overexpression | |
| Molecular Function | Antioxidant: GPX8 | Nucleic acid binding: IFIT1 |
| Hormonal activity: CGB2 | ||
| Biological process | Negative regulation of apoptosis: FGFR2 | Cell adhesion: ZNF645, PARD3, SSX21P |
| Negative regulation of apoptosis: BDNF | ||
| Epigenetic regulation: FAM172BP, MBD2 | ||
| Chromatin organization: MBD2 | ||
| Cell cycle: MAPRE3, TPTE2 | Signal transduction: MAPK: GDF1 | |
| Cell cycle: CDKL4, ACTL7B | ||
| Differentiation during development: FGFR2, HMCN2, BSPH1, PLP1, GATA4, LMOD1 | Differentiation during development: DCN, GDF1, BDNF, BRSK1 | |
| Cell death: ADAM30 | Cell Death: IFI27 | |
| Stress response: PROZ | Stress response: FIGF | |
| Cell component | Nucleus: CSN3, RBMXL2 | Nucleus: CDKL4 |
| Protein class | Adhesion: ITGB6 | Adhesion: FBLN2 |
| Nuclease: TDRD6 | ||
| Transcription factor: TBX15 | Growth factors: BDNF | |
| Voltage dependent sodium channel: SCN9A | ||
| Pathways | Angiogenesis: PDGFD | Interleukins: IL15RA |
| Cadherins: DCHS2, PCDH15, PCDH9 | ||
| Endothelin: EDNRB | ||
| PI3K pathway: TPTE2 | ||
| P53: TPTE2 | ||
| PBMCs Exposed to Permethrin | ||
|---|---|---|
| Underexpression | Overexpression | |
| Molecular Function | Nucleic acid binding: PRX, IFIT1B | |
| Transcription factor: ZFP42, WBP2NL | ||
| Cytokines: A2ML1, IL13RA2 | ||
| Biological Process | Negative regulation of apoptosis: LHX3 | Chromatin ensemble: HIST2H3BF |
| Cytoskeleton organization: TUBB8 | MAPK cascade: EPHA10, RAPGEF5, EPHA6, GAB1 | |
| Cell cycle: SAMD7 | Differentiation during development: SPRR1A | |
| Cell Growth: MXRA5, SEMA5B | ||
| Differentiation during development: PSAPL1 | Cell death: TNFSF11, PPEF1 | |
| DNA repair: HMGA2 | ||
| Cell death: PRUNE2 | Splicing: RBMX2 | |
| Mesoderm development: COL8A1 | Cytokine production: BTNL3 | |
| Stress response: ATG10 | ||
| Cell component | Nucleus: PUS7L, VGLL3 | |
| Protein Class | Kinases: MAPK4 | Transcription factor: HOXB5, FOXI1, PER3, MYT1, KDM5A |
| Pathways | Wnt: ANKYRIN | Inflammation: PLCH11 |
| Wnt: PCDHA8, PCDHB18P, MMTV, WNT11, MMP7 | ||
| Underexpression | Overexpression | ||
|---|---|---|---|
| Malathion | BMMCs | Channel activity | Inflammatory response |
| Negative regulation of cell development | Growth negative regulation | ||
| Ion channel activity | Cytokine activity | ||
| Cell differentiation negative regulation | Leukocyte differentiation | ||
| PBMCs | - | Synaptic regulation activity | |
| Permethrin | BMMCs | - | Type 1 Interferon signaling pathway |
| Type 1 Interferon cellular response | |||
| PBMCs | Wide pore cannel activity | Reactive oxygen species regulation | |
| Ephrins signaling | |||
| Protein-protein interaction, bridging |
| Repressed Genes in BMMCs Exposed to Malathion | |||||
| Gene | Locus | ΣZ DMSO | ΣZ Malathion | Methylation Status | ΣZ Malathion- ΣZ DMSO |
| KCNMA1 | 10q22.3 | 39.65 | 62.97 | Hypermethylated | 23.32 |
| ADAM11 | 17q21.31 | −44.58 | 55.14 | Hypermethylated | 99.72 |
| Overexpressed genes in BMMCs exposed to malathion | |||||
| Gene | Locus | ΣZ DMSO | ΣZ Malathion | Methylation status | ΣZ DMSO- ΣZ Malathion |
| INHBA | 7p14.1 | 9.74 | −11.36 | Hypomethylated | 21.10 |
| NR4A3 | 9q22 | 167.30 | 65.15 | Hypomethylated | 102.15 |
| ANGPTL4 | 19p13.2 | 31.23 | −62.18 | Hypomethylated | 93.41 |
| ITGB8 | 7p21.1 | 66.01 | 0.25 | Hypomethylated | 65.77 |
| Overexpressed genes in BMMCs exposed to permethrin | |||||
| Gene | Locus | ΣZ DMSO | ΣZ Permethrin | Methylation status | ΣZ DMSO- ΣZ Permethrin |
| ST6GAL2 | 2q12.3 | 86.51 | −3.67 | Hypomethylated | 90.17 |
| FIGF | Xp22.2 | 28.40 | 0.92 | Hypomethylated | 27.48 |
| BDNF | 11p14.1 | 128.06 | −1.27 | Hypomethylated | 129.32 |
| PARD3 | 10p11.22-p11.21 | 27.95 | −5.93 | Hypomethylated | 33.88 |
| MAPK | 22q11.22 | 18.07 | −13.69 | Hypomethylated | 31.76 |
| TDRD6 | 6p12.3 | 62.78 | 0.03 | Hypomethylated | 62.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarrete-Meneses, M.d.P.; Salas-Labadía, C.; Juárez-Velázquez, M.d.R.; Moreno-Lorenzana, D.; Gómez-Chávez, F.; Olaya-Vargas, A.; Pérez-Vera, P. Exposure to Insecticides Modifies Gene Expression and DNA Methylation in Hematopoietic Tissues In Vitro. Int. J. Mol. Sci. 2023, 24, 6259. https://doi.org/10.3390/ijms24076259
Navarrete-Meneses MdP, Salas-Labadía C, Juárez-Velázquez MdR, Moreno-Lorenzana D, Gómez-Chávez F, Olaya-Vargas A, Pérez-Vera P. Exposure to Insecticides Modifies Gene Expression and DNA Methylation in Hematopoietic Tissues In Vitro. International Journal of Molecular Sciences. 2023; 24(7):6259. https://doi.org/10.3390/ijms24076259
Chicago/Turabian StyleNavarrete-Meneses, María del Pilar, Consuelo Salas-Labadía, María del Rocío Juárez-Velázquez, Dafné Moreno-Lorenzana, Fernando Gómez-Chávez, Alberto Olaya-Vargas, and Patricia Pérez-Vera. 2023. "Exposure to Insecticides Modifies Gene Expression and DNA Methylation in Hematopoietic Tissues In Vitro" International Journal of Molecular Sciences 24, no. 7: 6259. https://doi.org/10.3390/ijms24076259
APA StyleNavarrete-Meneses, M. d. P., Salas-Labadía, C., Juárez-Velázquez, M. d. R., Moreno-Lorenzana, D., Gómez-Chávez, F., Olaya-Vargas, A., & Pérez-Vera, P. (2023). Exposure to Insecticides Modifies Gene Expression and DNA Methylation in Hematopoietic Tissues In Vitro. International Journal of Molecular Sciences, 24(7), 6259. https://doi.org/10.3390/ijms24076259