
Antiretrovirals Promote Insulin Resistance in HepG2 Liver Cells through miRNA Regulation and Transcriptional Activation of the NLRP3 Inflammasome


Abstract
:1. Introduction
2. Results
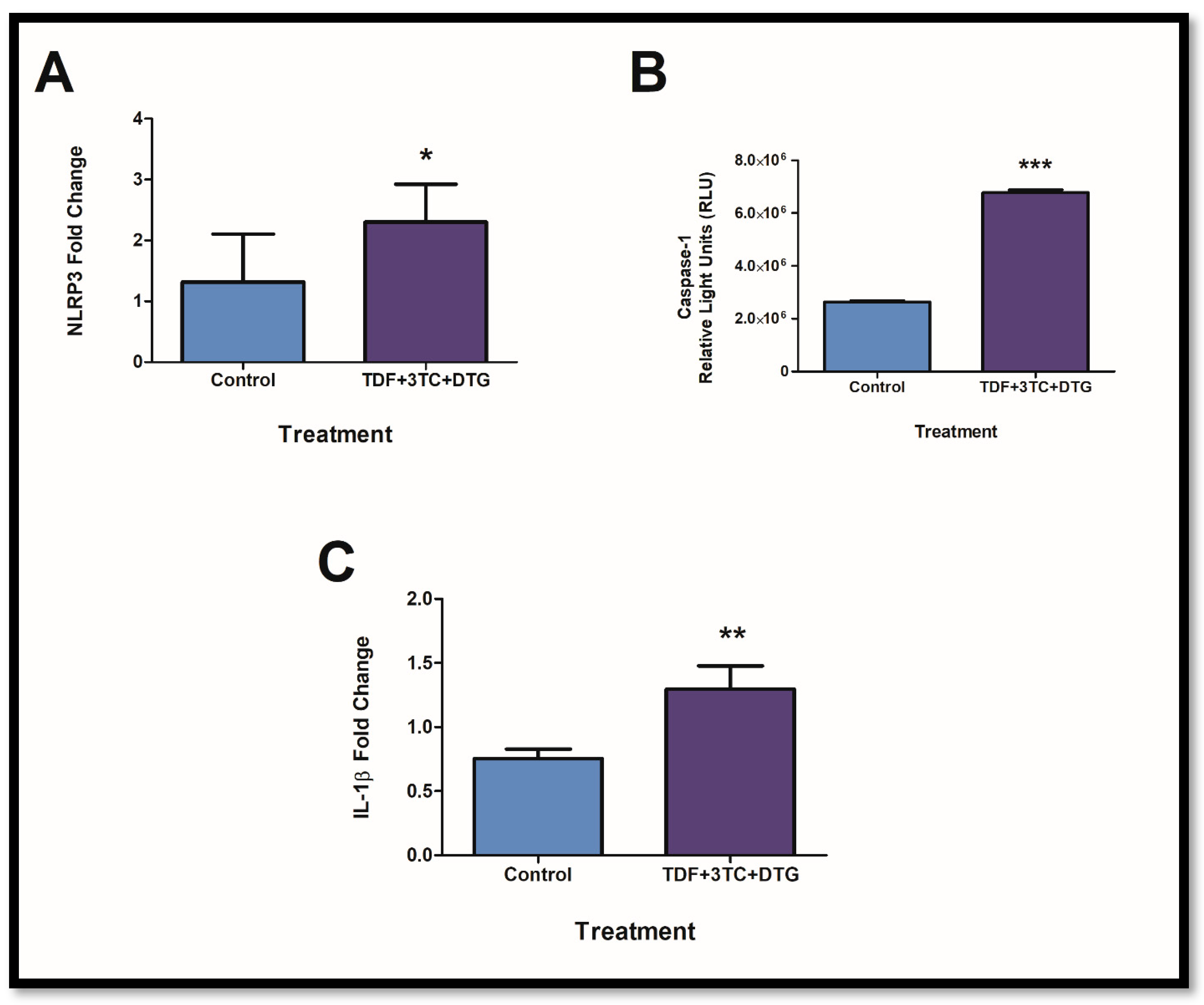
2.1. Combinational Usage of ARVs Results in the Upregulation of Key Components of the Inflammasome Pathway
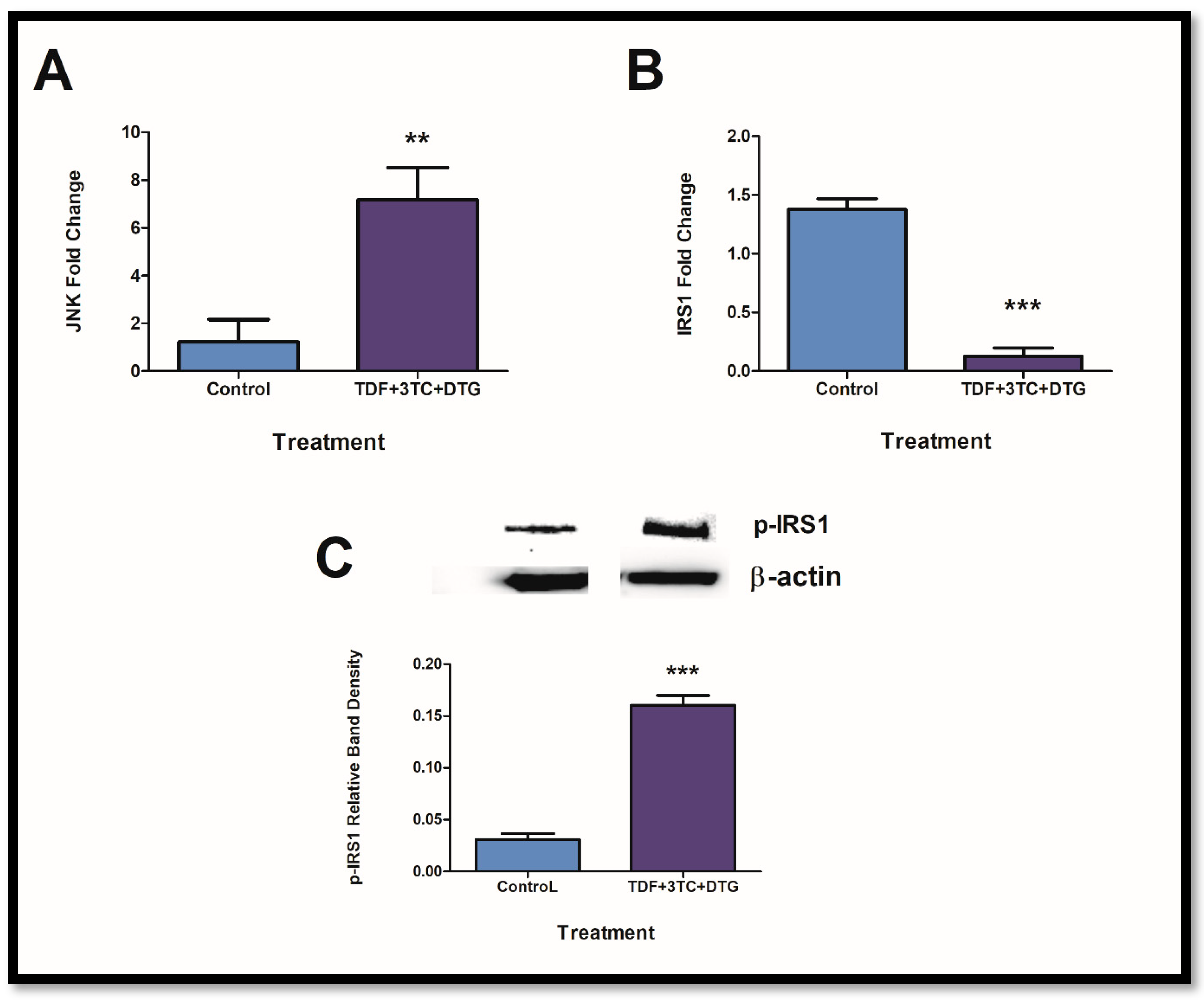
2.2. Exposure to ARVs Alters JNK and Insulin Receptor Expressions
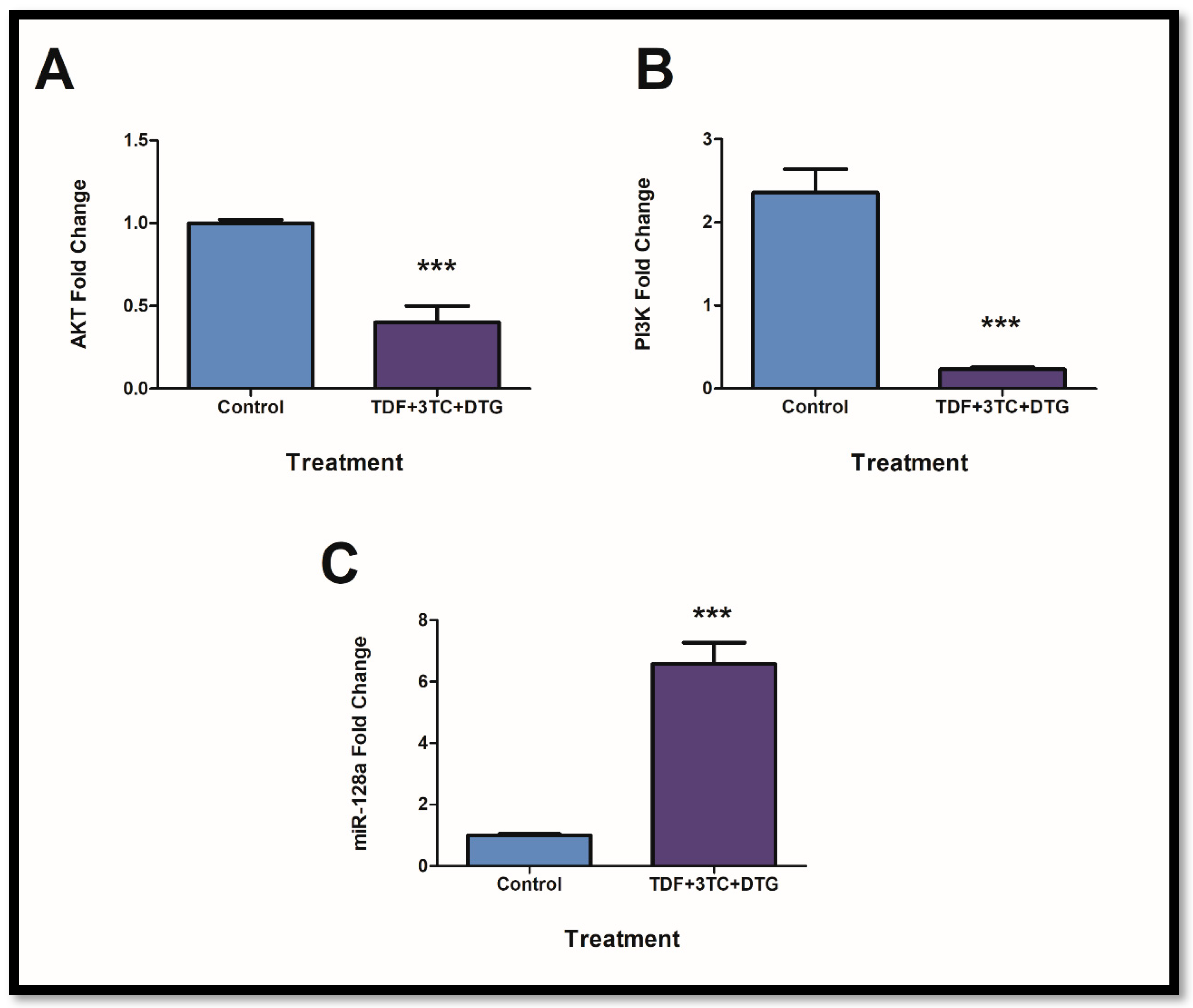
2.3. Prolonged Exposure to ARVs Resulted in Disruption of the PI3K/AKT Pathway via Inflammasome Activation and miR-128a Expression
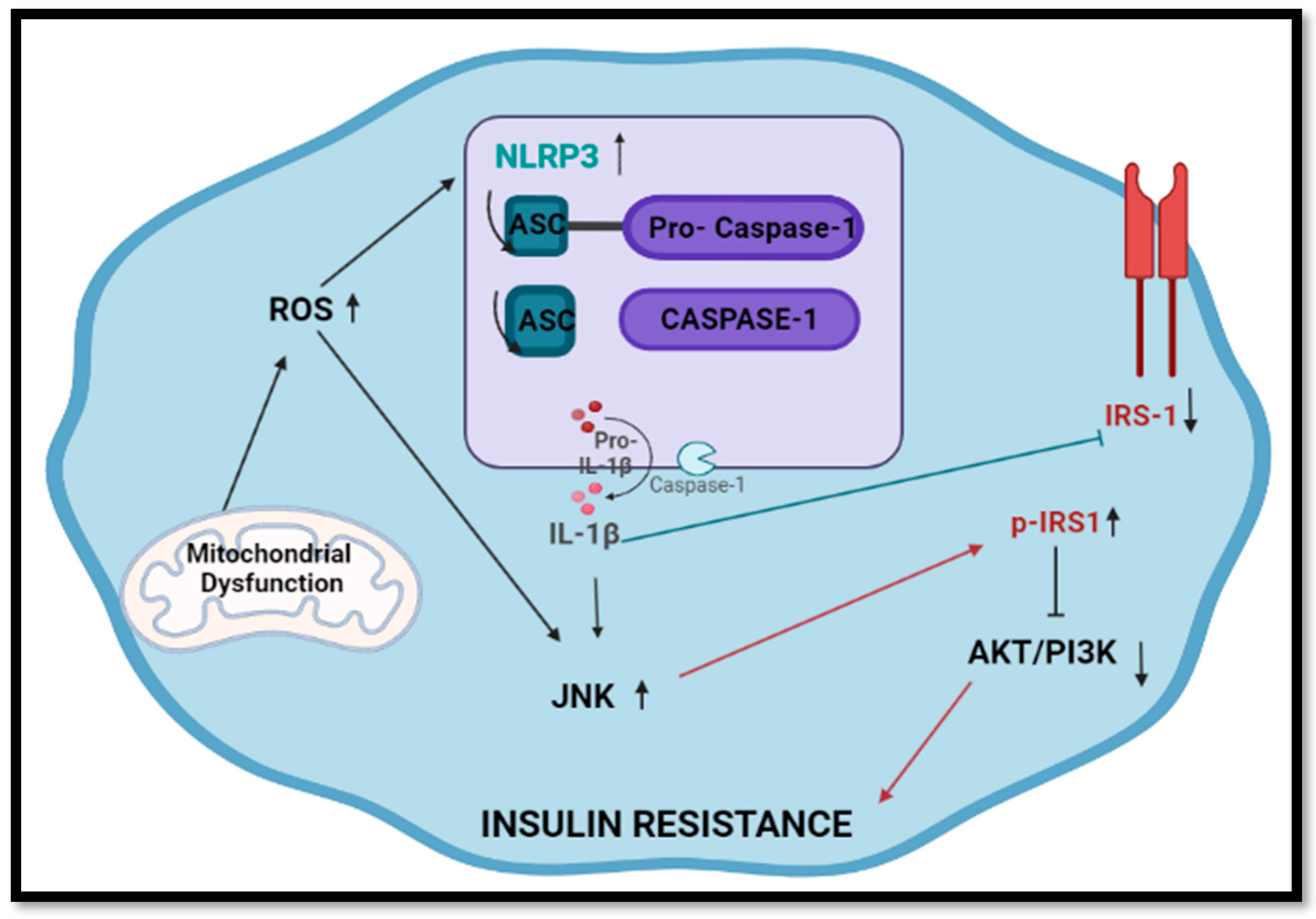
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Caspase 1 Detection
4.3. Western Blot
4.4. Quantitative PCR
4.4.1. RNA Isolation and Quantification
4.4.2. Quantification of mRNA Expression
4.4.3. Quantification of miR-128a Expression
4.5. Statistical Analysis
5. Future Recommendations and Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prasun, P. Mitochondrial dysfunction in metabolic syndrome. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2021, 1866, 165838. [Google Scholar] [CrossRef]
- Saklayen, M.G. The global epidemic of the metabolic syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Todowede, O.O.; Mianda, S.Z.; Sartorius, B. Prevalence of metabolic syndrome among HIV-positive and HIV-negative populations in sub-Saharan Africa—A systematic review and meta-analysis. Syst. Rev. 2019, 8, 4. [Google Scholar] [CrossRef]
- Chang, H.-H. Weight gain and metabolic syndrome in human immunodeficiency virus patients. Infect. Chemother. 2022, 54, 220. [Google Scholar] [CrossRef]
- Sapuła, M.; Suchacz, M.; Załęski, A.; Wiercińska-Drapało, A. Impact of combined antiretroviral therapy on metabolic syndrome components in adult people living with HIV: A literature review. Viruses 2022, 14, 122. [Google Scholar] [CrossRef]
- UNAIDS. Global HIV & AIDS Statistics—2020 Fact Sheet. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 11 February 2022).
- Stats-SA. 2020 Mid-Year Population Estimates. Available online: http://www.statssa.gov.za/?p=13453 (accessed on 11 April 2022).
- WHO. HIV/AIDS. Available online: https://www.who.int/news-room/fact-sheets/detail/hiv-aids (accessed on 20 January 2022).
- Cheney, L.; Barbaro, J.M.; Berman, J.W. Antiretroviral Drugs Impact Autophagy with Toxic Outcomes. Cells 2021, 10, 909. [Google Scholar] [CrossRef]
- Tagliari, C.; de Oliveira, C.N.; Vogel, G.M.; da Silva, P.B.; Linden, R.; Lazzaretti, R.K.; Notti, R.K.; Sprinz, E.; Mattevi, V.S. Investigation of SIRT1 gene variants in HIV-associated lipodystrophy and metabolic syndrome. Genet. Mol. Biol. 2020, 43, e20190142. [Google Scholar] [CrossRef]
- Chhoun, P.; Tuot, S.; Harries, A.D.; Kyaw, N.T.T.; Pal, K.; Mun, P.; Brody, C.; Mburu, G.; Yi, S. High prevalence of non-communicable diseases and associated risk factors amongst adults living with HIV in Cambodia. PLoS ONE 2017, 12, e0187591. [Google Scholar] [CrossRef] [Green Version]
- Hyle, E.P.; Naidoo, K.; Su, A.E.; El-Sadr, W.M.; Freedberg, K.A. HIV, tuberculosis, and non-communicable diseases: What is known about the costs, effects, and cost-effectiveness of integrated care? J. Acquir. Immune Defic. Syndr. (1999) 2014, 67, S87. [Google Scholar] [CrossRef] [Green Version]
- Araujo, S.; Bañó, S.; Machuca, I.; Moreno, A.; Pérez-Elıas, M.J.; Casado, J.L. Prevalence of insulin resistance and risk of diabetes mellitus in HIV-infected patients receiving current antiretroviral drugs. Eur. J. Endocrinol. 2014, 171, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, N.; Nakao, M.; Matsui, S.; Satoh, T. A Crucial Role for the Small GTPase Rac1 Downstream of the Protein Kinase Akt2 in Insulin Signaling that Regulates Glucose Uptake in Mouse Adipocytes. Int. J. Mol. Sci. 2019, 20, 5443. [Google Scholar] [CrossRef] [Green Version]
- Sokolowska, E.; Blachnio-Zabielska, A. The role of ceramides in insulin resistance. Front. Endocrinol. 2019, 10, 577. [Google Scholar] [CrossRef] [Green Version]
- Leavens, K.F.; Birnbaum, M.J. Insulin signaling to hepatic lipid metabolism in health and disease. Crit. Rev. Biochem. Mol. Biol. 2011, 46, 200–215. [Google Scholar] [CrossRef]
- Targher, G.; Corey, K.E.; Byrne, C.D.; Roden, M. The complex link between NAFLD and type 2 diabetes mellitus—Mechanisms and treatments. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 599–612. [Google Scholar] [CrossRef]
- Gnanasekaran, N. The missing link between HAART, mitochondrial damage and insulin resistance. Biomed. Pharmacol. J. 2020, 13, 965–972. [Google Scholar] [CrossRef]
- Jager, J.; Grémeaux, T.; Cormont, M.; Le Marchand-Brustel, Y.; Tanti, J.-F. Interleukin-1β-induced insulin resistance in adipocytes through down-regulation of insulin receptor substrate-1 expression. Endocrinology 2007, 148, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-A.; Wei, Y.; Sowers, J.R. Role of mitochondrial dysfunction in insulin resistance. Circ. Res. 2008, 102, 401–414. [Google Scholar] [CrossRef]
- Jorquera, G.; Russell, J.; Monsalves-Álvarez, M.; Cruz, G.; Valladares-Ide, D.; Basualto-Alarcón, C.; Barrientos, G.; Estrada, M.; Llanos, P. NLRP3 inflammasome: Potential role in obesity related low-grade inflammation and insulin resistance in skeletal muscle. Int. J. Mol. Sci. 2021, 22, 3254. [Google Scholar] [CrossRef]
- Mullis, C.; Swartz, T.H. NLRP3 Inflammasome signaling as a link between HIV-1 infection and atherosclerotic cardiovascular disease. Front. Cardiovasc. Med. 2020, 7, 95. [Google Scholar] [CrossRef]
- Choi, H.; Koh, H.W.; Zhou, L.; Cheng, H.; Loh, T.P.; Parvaresh Rizi, E.; Toh, S.A.; Ronnett, G.V.; Huang, B.E.; Khoo, C.M. Plasma protein and microRNA biomarkers of insulin resistance: A network-based integrative-omics analysis. Front. Physiol. 2019, 10, 379. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, N.; Alexander, M.S.; Shimizu-Motohashi, Y.; Myers, J.A.; Kawahara, G.; Kunkel, L.M. Regulation of IRS1/Akt insulin signaling by microRNA-128a during myogenesis. J. Cell Sci. 2013, 126, 2678–2691. [Google Scholar] [CrossRef] [Green Version]
- WHO. Update of Recommendations on First-and Second-Line Antiretroviral Regimens. Available online: https://apps.who.int/iris/bitstream/handle/10665/325892/WHO-CDS-HIV-19.15-eng.pdf (accessed on 12 December 2021).
- Mohan, J.; Ghazi, T.; Chuturgoon, A.A. A Critical Review of the Biochemical Mechanisms and Epigenetic Modifications in HIV- and Antiretroviral-Induced Metabolic Syndrome. Int. J. Mol. Sci. 2021, 22, 12020. [Google Scholar] [CrossRef]
- Ruoß, M.; Damm, G.; Vosough, M.; Ehret, L.; Grom-Baumgarten, C.; Petkov, M.; Naddalin, S.; Ladurner, R.; Seehofer, D.; Nussler, A. Epigenetic modifications of the liver tumor cell line HepG2 increase their drug metabolic capacity. Int. J. Mol. Sci. 2019, 20, 347. [Google Scholar] [CrossRef] [Green Version]
- Nagiah, S.; Phulukdaree, A.; Chuturgoon, A. Mitochondrial and oxidative stress response in HepG2 cells following acute and prolonged exposure to antiretroviral drugs. J. Cell. Biochem. 2015, 116, 1939–1946. [Google Scholar] [CrossRef] [Green Version]
- Nagiah, S.; Phulukdaree, A.; Chuturgoon, A. Inverse association between microRNA-124a and ABCC4 in HepG2 cells treated with antiretroviral drugs. Xenobiotica 2016, 46, 825–830. [Google Scholar] [CrossRef]
- Sibiya, T.; Ghazi, T.; Mohan, J.; Nagiah, S.; Chuturgoon, A.A. Spirulina platensis Ameliorates Oxidative Stress Associated with Antiretroviral Drugs in HepG2 Cells. Plants 2022, 11, 3143. [Google Scholar] [CrossRef]
- Otto, A.O.; Rivera, C.G.; Zeuli, J.D.; Temesgen, Z. Hepatotoxicity of Contemporary Antiretroviral Drugs: A Review and Evaluation of Published Clinical Data. Cells 2021, 10, 1263. [Google Scholar] [CrossRef]
- Leal, V.N.C.; Reis, E.C.; Pontillo, A. Inflammasome in HIV infection: Lights and shadows. Mol. Immunol. 2020, 118, 9–18. [Google Scholar] [CrossRef]
- Guo, H.; Callaway, J.B.; Ting, J.P. Inflammasomes: Mechanism of action, role in disease, and therapeutics. Nat. Med. 2015, 21, 677–687. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Xu, W.; Zhou, R. NLRP3 inflammasome activation and cell death. Cell. Mol. Immunol. 2021, 18, 2114–2127. [Google Scholar] [CrossRef]
- Lu, A.; Magupalli, V.G.; Ruan, J.; Yin, Q.; Atianand, M.K.; Vos, M.R.; Schröder, G.F.; Fitzgerald, K.A.; Wu, H.; Egelman, E.H. Unified polymerization mechanism for the assembly of ASC-dependent inflammasomes. Cell 2014, 156, 1193–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Chen, J.; Xu, H.; Liu, S.; Jiang, Q.-X.; Halfmann, R.; Chen, Z.J. Prion-like polymerization underlies signal transduction in antiviral immune defense and inflammasome activation. Cell 2014, 156, 1207–1222. [Google Scholar] [CrossRef] [Green Version]
- El-Sharkawy, L.Y.; Brough, D.; Freeman, S. Inhibiting the NLRP3 Inflammasome. Molecules 2020, 25, 5533. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-J.; Mao, L.-F.; Lai, H.-L.; Wang, Y.-W.; Jiang, Z.-B.; Li, W.; Huang, J.-M.; Xie, Y.-J.; Xu, C.; Liu, P. Dolutegravir derivative inhibits proliferation and induces apoptosis of non-small cell lung cancer cells via calcium signaling pathway. Pharmacol. Res. 2020, 161, 105129. [Google Scholar] [CrossRef] [PubMed]
- Hamed, M.; Aremu, G.; Akhigbe, R. Concomitant administration of HAART aggravates anti-Koch-induced oxidative hepatorenal damage via dysregulation of glutathione and elevation of uric acid production. Biomed. Pharmacother. 2021, 137, 111309. [Google Scholar] [CrossRef]
- George, J.W.; Mattingly, J.E.; Roland, N.J.; Small, C.M.; Lamberty, B.G.; Fox, H.S.; Stauch, K.L. Physiologically Relevant Concentrations of Dolutegravir, Emtricitabine, and Efavirenz Induce Distinct Metabolic Alterations in HeLa Epithelial and BV2 Microglial Cells. Front. Immunol. 2021, 12, 639378. [Google Scholar] [CrossRef]
- Ramamoorthy, H.; Abraham, P.; Isaac, B. Mitochondrial dysfunction and electron transport chain complex defect in a rat model of tenofovir disoproxil fumarate nephrotoxicity. J. Biochem. Mol. Toxicol. 2014, 28, 246–255. [Google Scholar] [CrossRef]
- Gorwood, J.; Bourgeois, C.; Pourcher, V.; Pourcher, G.; Charlotte, F.; Mantecon, M.; Rose, C.; Morichon, R.; Atlan, M.; Le Grand, R. The integrase inhibitors dolutegravir and raltegravir exert proadipogenic and profibrotic effects and induce insulin resistance in human/simian adipose tissue and human adipocytes. Clin. Infect. Dis. 2020, 71, e549–e560. [Google Scholar] [CrossRef]
- Blümer, R.M.; van Vonderen, M.G.; Sutinen, J.; Hassink, E.; Ackermans, M.; van Agtmael, M.A.; Yki-Jarvinen, H.; Danner, S.A.; Reiss, P.; Sauerwein, H.P. Zidovudine/lamivudine contributes to insulin resistance within 3 months of starting combination antiretroviral therapy. Aids 2008, 22, 227–236. [Google Scholar] [CrossRef]
- Jia, J.; Yao, P.; Arif, A.; Fox, P.L. Regulation and dysregulation of 3′ UTR-mediated translational control. Curr. Opin. Genet. Dev. 2013, 23, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Ge, Q.; Chen, X.; Zhao, Y.; Mu, H.; Zhang, J. Modulatory mechanisms of NLRP3: Potential roles in inflammasome activation. Life Sci. 2021, 267, 118918. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Tsai, L.T.; Rosen, E.D. Nuclear mechanisms of insulin resistance. Trends Cell Biol. 2016, 26, 341–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Dey, D.; Roy, S.S. Molecular mechanism of insulin resistance. J. Biosci. 2007, 32, 405–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottrell, M.L.; Hadzic, T.; Kashuba, A.D. Clinical pharmacokinetic, pharmacodynamic and drug-interaction profile of the integrase inhibitor dolutegravir. Clin. Pharmacokinet. 2013, 52, 981–994. [Google Scholar] [CrossRef] [Green Version]
- Erickson-Viitanen, S.; Wu, J.-T.; Shi, G.; Unger, S.; King, R.W.; Fish, B.; Klabe, R.; Geleziunas, R.; Gallagher, K.; Otto, M.J. Cellular pharmacology of D-d4FC, a nucleoside analogue active against drug-resistant HIV. Antivir. Chem. Chemother. 2003, 14, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Adefolaju, G.A.; Scholtz, K.E.; Hosie, M.J. Expresión de VEGF165b Antiangiogénico en Células MCF-7 y MCF-10A de Mama Humana Expuesto a Inhibidores de la Transcriptasa Inversa y la Proteasa. Int. J. Morphol. 2017, 35, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | Annealing Temperature (°C) | |
| NLRP3 | Forward Reverse | CAGGTGTTGGAATTAGACAAC TTCAGACAACCCCAGGTTCT | 60 |
| IL-1β | Forward Reverse | ACGAATCTCCGACCACCACTAC TCCATGGCCACAACAACTGACG | 60 |
| AKT | Forward Reverse | TGGACTACCTGCACTCGGAGAA GTGCCGCAAAAGGTCTTCATGG | 59 |
| PI3K | Forward Reverse | GAAGCACCTGAATAGGCAAGTCG GAGCATCCATGAAATCTGGTCGC | 59 |
| IRS1 | Forward Reverse | AGTCTGTCGTCCAGTAGCACCA ACTGGAGCCATACTCATCCGAG | 59 |
| JNK | Forward Reverse | GACGCCTTATGTAGTGACTCGC TCCTGGAAAGAGGATTTTGTGGC | 59 |
| GAPDH | Forward Reverse | TCCACCACCCTGTTGCTGTA ACCACAGTCCATGCCATCAC | --- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohan, J.; Ghazi, T.; Mazibuko, M.S.; Chuturgoon, A.A. Antiretrovirals Promote Insulin Resistance in HepG2 Liver Cells through miRNA Regulation and Transcriptional Activation of the NLRP3 Inflammasome. Int. J. Mol. Sci. 2023, 24, 6267. https://doi.org/10.3390/ijms24076267
Mohan J, Ghazi T, Mazibuko MS, Chuturgoon AA. Antiretrovirals Promote Insulin Resistance in HepG2 Liver Cells through miRNA Regulation and Transcriptional Activation of the NLRP3 Inflammasome. International Journal of Molecular Sciences. 2023; 24(7):6267. https://doi.org/10.3390/ijms24076267
Chicago/Turabian StyleMohan, Jivanka, Terisha Ghazi, Makabongwe S. Mazibuko, and Anil A. Chuturgoon. 2023. "Antiretrovirals Promote Insulin Resistance in HepG2 Liver Cells through miRNA Regulation and Transcriptional Activation of the NLRP3 Inflammasome" International Journal of Molecular Sciences 24, no. 7: 6267. https://doi.org/10.3390/ijms24076267
APA StyleMohan, J., Ghazi, T., Mazibuko, M. S., & Chuturgoon, A. A. (2023). Antiretrovirals Promote Insulin Resistance in HepG2 Liver Cells through miRNA Regulation and Transcriptional Activation of the NLRP3 Inflammasome. International Journal of Molecular Sciences, 24(7), 6267. https://doi.org/10.3390/ijms24076267