
Comparative Transcriptomic Analysis Reveals the Functionally Segmented Intestine in Tunicate Ascidian



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
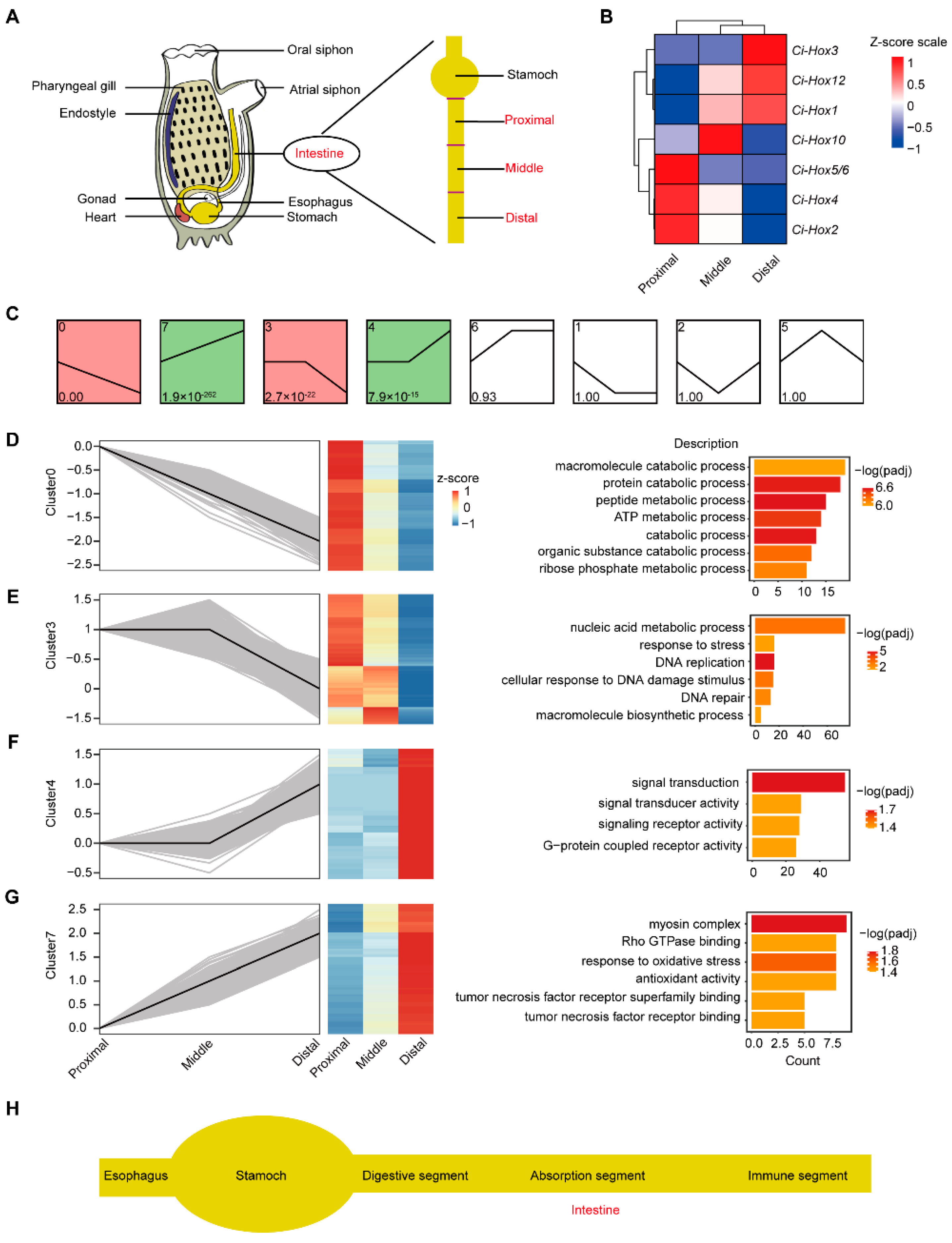
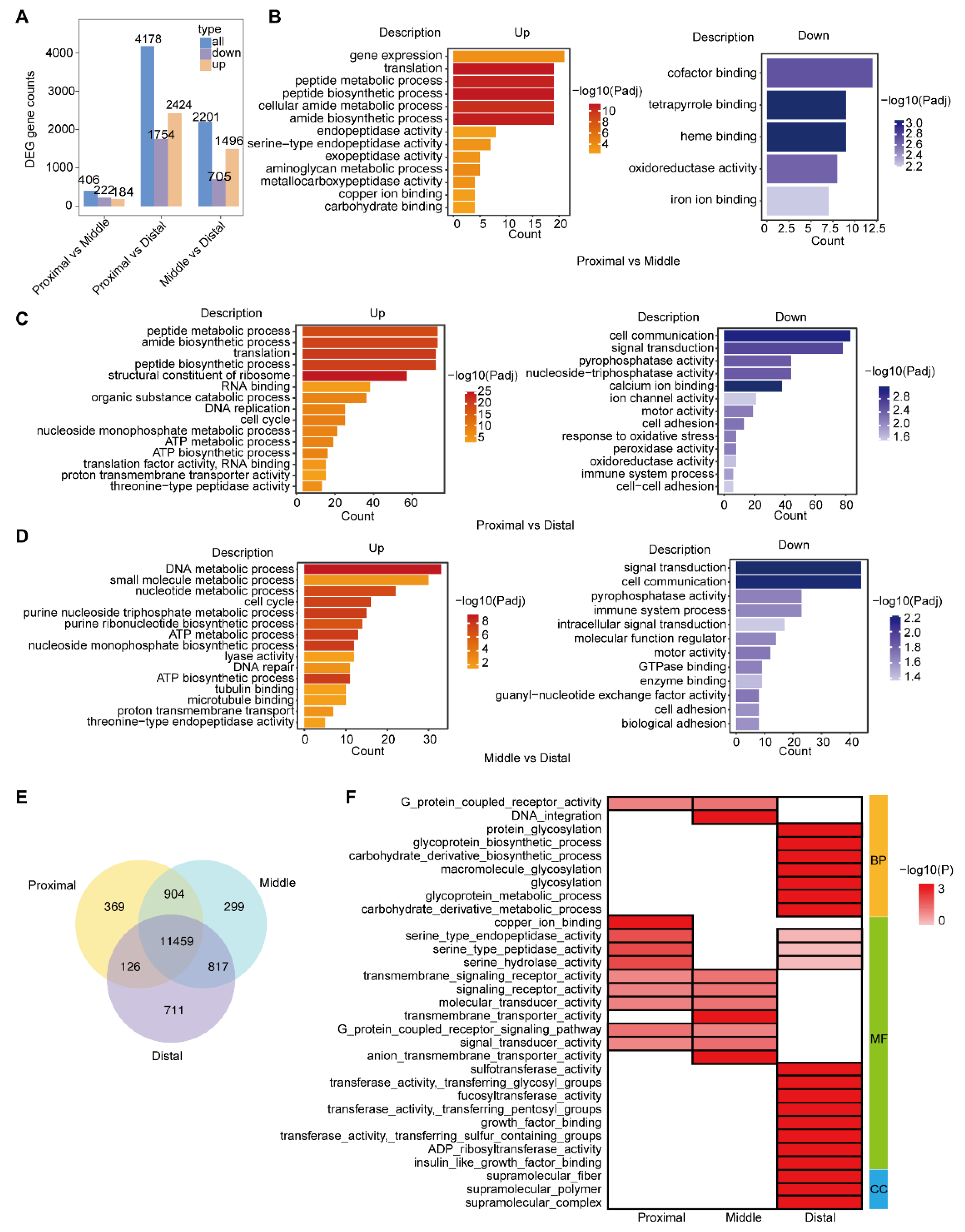
2.1. Gene Enrichment Analysis Reveals the Segmented Expression Patterns of C. savignyi Intestine
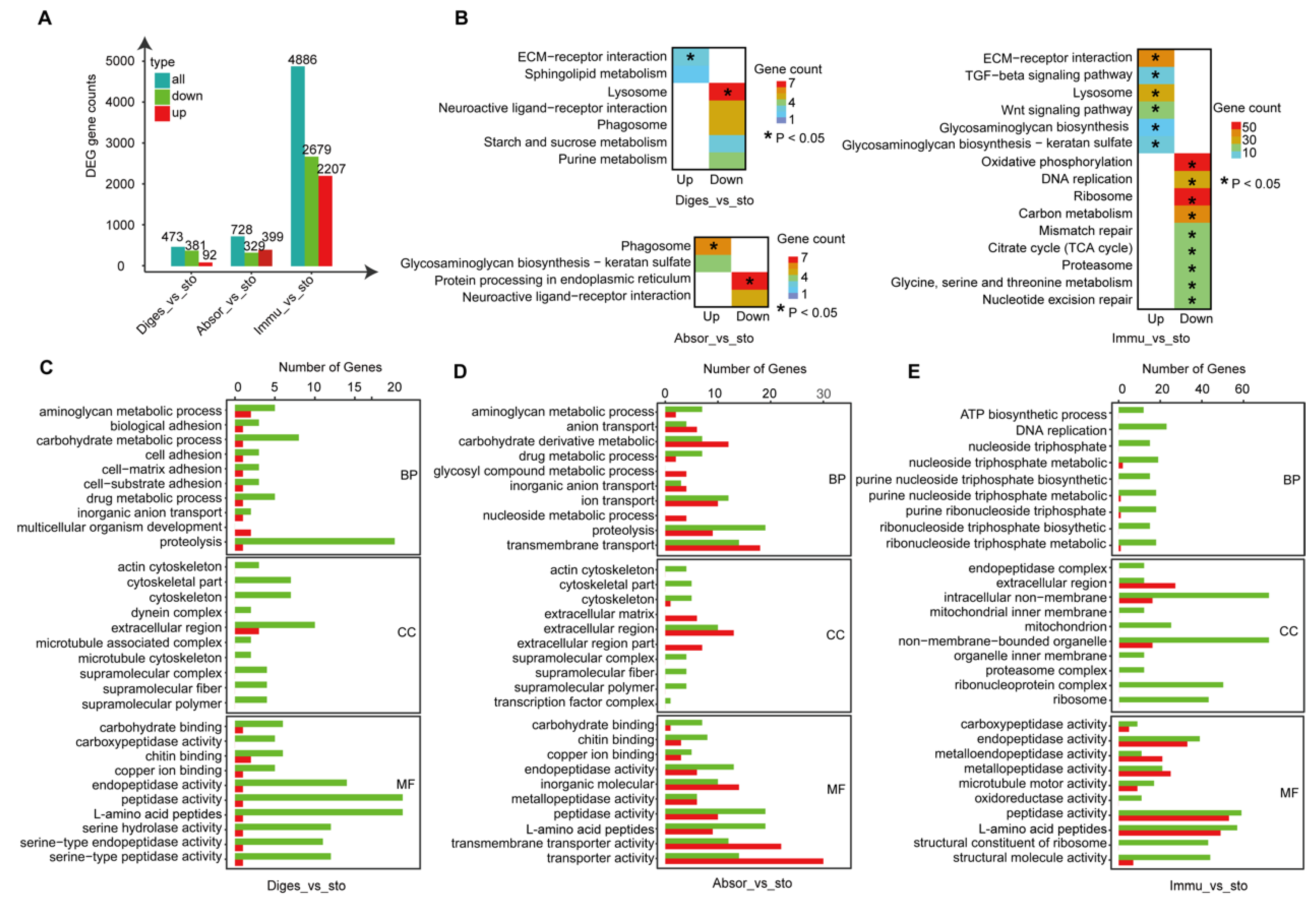
2.2. The Functional Relationship between Intestine and Stomach
2.3. Expression of the Orthologs of Vertebrate Digestive Organ−Specific Genes in the C. savignyi Intestine
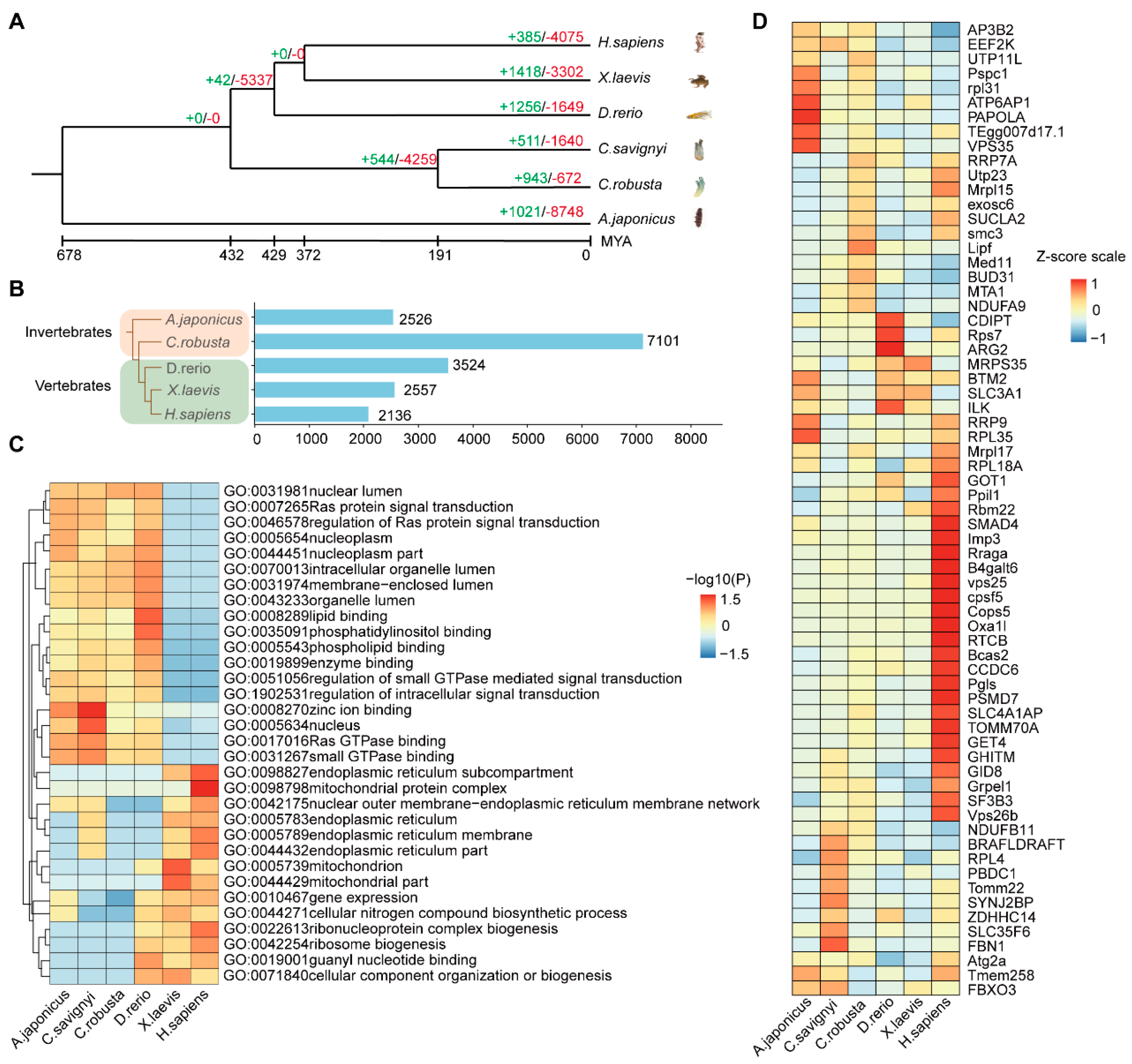
2.4. The Multi−Species Comparison Explains the Conservation and Divergence of Intestine Function during Evolution
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Preparation
4.2. RNA Extraction and Transcriptome Sequencing
4.3. Transcriptome Analysis
4.4. Combinatorial Analyses of Transcriptome Data
4.5. Expression Tendency Analysis of all Genes
4.6. Selection and Phylogenetic Analysis of Orthologous Genes
4.7. The Alignment Analysis of Homologous Genes of C. savignyi against Multiple Species
4.8. Quantitative Reverse−Transcription PCR (qRT−PCR)
4.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karasov, W.H.; Douglas, A.E. Comparative digestive physiology. Compr. Physiol. 2013, 3, 741–783. [Google Scholar] [PubMed] [Green Version]
- Spence, J.R.; Lauf, R.; Shroyer, N.F. Vertebrate intestinal endoderm development. Dev. Dyn. 2011, 240, 501–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marks, V.; Samols, E. Intestinal factors in the regulation of insulin secretion. Adv. Metab. Disord. 1970, 4, 1–38. [Google Scholar]
- Miguel-Aliaga, I.; Jasper, H.; Lemaitre, B. Anatomy and Physiology of the Digestive Tract of Drosophila melanogaster. Genetics 2018, 210, 357–396. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, J.; Ding, X.; Zhang, P.; Jia, K.; Fang, C.; Zhu, Y.; Spink, D.C.; Zhang, Q.Y. Potential biological functions of cytochrome P450 reductase-dependent enzymes in small intestine: Novel link to expression of major histocompatibility complex class II genes. J. Biol. Chem. 2012, 287, 17777–17788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, D.H.; Twumasi-Boateng, K.; Kang, H.S.; Carlisle, J.A.; Hanganu, A.; Lai, T.Y.; Shapira, M. The Developmental Intestinal Regulator ELT-2 Controls p38-Dependent Immune Responses in Adult C. elegans. PLoS Genet. 2015, 11, e1005265. [Google Scholar] [CrossRef] [Green Version]
- Grapin-Botton, A.; Majithia, A.R.; Melton, D.A. Key events of pancreas formation are triggered in gut endoderm by ectopic expression of pancreatic regulatory genes. Genes. Dev. 2001, 15, 444–454. [Google Scholar] [CrossRef] [Green Version]
- Naftalin, R. Comparative Physiology of the Digestive System of Vertebrates. Gut 1996, 39, 498. [Google Scholar] [CrossRef] [Green Version]
- Koelz, H.R. Gastric acid in vertebrates. Scand. J. Gastroenterol. Suppl. 1992, 27 (Suppl. S193), 2–6. [Google Scholar] [CrossRef]
- Richardson, M.K.; Wright, G.M. Developmental transformations in a normal series of embryos of the sea lamprey Petromyzon marinus (Linnaeus). J. Morphol. 2003, 257, 348–363. [Google Scholar] [CrossRef]
- Joseph, C.C.; Gregorio, A.D.; Levine, M. The Ascidian as a Model Organism in Developmental and Evolutionary Biology. Cell Press 2001, 106, 535–538. [Google Scholar]
- Delsuc, F.; Tsagkogeorga, G.; Lartillot, N.; Philippe, H. Additional molecular support for the new chordate phylogeny. Genesis 2008, 46, 592–604. [Google Scholar] [CrossRef] [Green Version]
- Ikuta, T.; Yoshida, N.; Satoh, N.; Saiga, H. Ciona intestinalis Hox gene cluster: Its dispersed structure and residual colinear expression in development. Proc. Natl. Acad. Sci. USA 2004, 101, 15118–15123. [Google Scholar] [CrossRef] [Green Version]
- Sasakura, Y.; Kanda, M.; Ikeda, T.; Horie, T.; Kawai, N.; Ogura, Y.; Yoshida, R.; Hozumi, A.; Satoh, N.; Fujiwara, S. Retinoic acid-driven Hox1 is required in the epidermis for forming the otic/atrial placodes during ascidian metamorphosis. Development 2012, 139, 2156–2160. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, S.; Ogasawara, M. Compartmentalized expression patterns of pancreatic- and gastric-related genes in the alimentary canal of the ascidian Ciona intestinalis: Evolutionary insights into the functional regionality of the gastrointestinal tract in Olfactores. Cell Tissue Res. 2017, 370, 113–128. [Google Scholar] [CrossRef]
- Matsubara, S.; Osugi, T.; Shiraishi, A.; Wada, A.; Satake, H. Comparative analysis of transcriptomic profiles among ascidians, zebrafish, and mice: Insights from tissue-specific gene expression. PLoS ONE 2021, 16, e0254308. [Google Scholar] [CrossRef]
- Zhao, L.; Song, W.; Chen, Y.G. Mesenchymal-epithelial interaction regulates gastrointestinal tract development in mouse embryos. Cell Rep. 2022, 40, 111053. [Google Scholar] [CrossRef]
- Byrnes, L.E.; Wong, D.M.; Subramaniam, M.; Meyer, N.P.; Gilchrist, C.L.; Knox, S.M.; Tward, A.D.; Ye, C.J.; Sneddon, J.B. Lineage dynamics of murine pancreatic development at single-cell resolution. Nat. Commun. 2018, 9, 3922. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, S.; Sekiguchi, T.; Ogasawara, M. Molecular and evolutionary aspects of the protochordate digestive system. Cell Tissue Res. 2019, 377, 309–320. [Google Scholar] [CrossRef]
- Beumer, J.; Clevers, H. Cell fate specification and differentiation in the adult mammalian intestine. Nat. Rev. Mol. Cell Biol. 2021, 22, 39–53. [Google Scholar] [CrossRef]
- Clevers, H. The intestinal crypt, a prototype stem cell compartment. Cell 2013, 154, 274–284. [Google Scholar] [CrossRef] [Green Version]
- Lehane, M.J.; Billingsley, P.F. Mechanisms controlling the synthesis and secretion of digestive enzyme in insects. Biol. Insect Midgut 1996, 7, 195–205. [Google Scholar]
- Hou, S.X. Intestinal stem cell asymmetric division in the Drosophila posterior midgut. J. Cell. Physiol. 2010, 224, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Leung, B.; Hermann, G.J.; Priess, J.R. Organogenesis of the Caenorhabditis elegans Intestine. Dev. Biol. 1999, 216, 114–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, B.M.; Avery, L. The pharynx of the nematode C. elegans: A model system for the study of motor control. Worm 2013, 2, e21833. [Google Scholar] [CrossRef] [Green Version]
- Dimov, I.; Maduro, M.F. The C. elegans intestine: Organogenesis, digestion, and physiology. Cell Tissue Res. 2019, 377, 383–396. [Google Scholar] [CrossRef]
- Li, J.; Prochaska, M.; Maney, L.; Wallace, K.N. Development and organization of the zebrafish intestinal epithelial stem cell niche. Dev. Dyn. 2020, 249, 76–87. [Google Scholar] [CrossRef]
- Wallace, K.N.; Pack, M. Unique and conserved aspects of gut development in zebrafish. Dev. Biol. 2003, 255, 12–29. [Google Scholar] [CrossRef] [Green Version]
- Ng, A.N.; de Jong-Curtain, T.A.; Mawdsley, D.J.; White, S.J.; Shin, J.; Appel, B.; Dong, P.D.; Stainier, D.Y.; Heath, J.K. Formation of the digestive system in zebrafish: III. Intestinal epithelium morphogenesis. Dev. Biol. 2005, 286, 114–135. [Google Scholar] [CrossRef] [Green Version]
- Wallace, K.N.; Akhter, S.; Smith, E.M.; Lorent, K.; Pack, M. Intestinal growth and differentiation in zebrafish. Mech. Dev. 2005, 122, 157–173. [Google Scholar] [CrossRef]
- Nakayama, S.; Satou, K.; Orito, W.; Ogasawara, M. Ordered expression pattern of Hox and ParaHox genes along the alimentary canal in the ascidian juvenile. Cell Tissue Res. 2016, 365, 65–75. [Google Scholar] [CrossRef]
- Ikuta, T.; Satoh, N.; Saiga, H. Limited functions of Hox genes in the larval development of the ascidian Ciona intestinalis. Development 2010, 137, 1505–1513. [Google Scholar] [CrossRef] [Green Version]
- Hartenstein, V.; Martinez, P. Structure, development and evolution of the digestive system. Cell Tissue Res. 2019, 377, 289–292. [Google Scholar] [CrossRef] [Green Version]
- Annunziata, R.; Andrikou, C.; Perillo, M.; Cuomo, C.; Arnone, M.I. Development and evolution of gut structures: From molecules to function. Cell Tissue Res. 2019, 377, 445–458. [Google Scholar] [CrossRef]
- Nakano, E.; Geisz, A.; Masamune, A.; Niihori, T.; Hamada, S.; Kume, K.; Kakuta, Y.; Aoki, Y.; Matsubara, Y.; Ebert, K.; et al. Variants in pancreatic carboxypeptidase genes CPA2 and CPB1 are not associated with chronic pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G688–G694. [Google Scholar] [CrossRef] [Green Version]
- Campbell, E.M.; Fares, H. Roles of CUP-5, the Caenorhabditis elegans orthologue of human TRPML1, in lysosome and gut granule biogenesis. BMC Cell Biol. 2010, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Jahan, S.; Awaja, N.; Hess, B.; Hajjar, S.; Sad, S.; Lohnes, D. The transcription factor Cdx2 regulates inflammasome activity through expression of the NLRP3 suppressor TRIM31 to maintain intestinal homeostasis. J. Biol. Chem. 2022, 298, 102386. [Google Scholar] [CrossRef]
- Jang, D.I.; Lee, A.H.; Shin, H.Y.; Song, H.R.; Park, J.H.; Kang, T.B.; Lee, S.R.; Yang, S.H. The role of tumor necrosis factor alpha (TNF-α) in autoimmune disease and current TNF-α inhibitors in therapeutics. Int. J. Mol. Sci. 2021, 22, 2719. [Google Scholar] [CrossRef]
- Willrich, M.A.; Murray, D.L.; Snyder, M.R. Tumor necrosis factor inhibitors: Clinical utility in autoimmune diseases. Transl. Res. 2015, 165, 270–282. [Google Scholar] [CrossRef]
- Dipankar, P.; Kumar, P.; Dash, S.P.; Sarangi, P.P. Functional and therapeutic relevance of Rho GTPases in innate immune cell migration and function during inflammation: An in silico perspective. Mediat. Inflamm. 2021, 2021, 6655412. [Google Scholar] [CrossRef]
- El Masri, R.; Delon, J. RHO GTPases: From new partners to complex immune syndromes. Nat. Rev. Immunol. 2021, 21, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef] [PubMed]
- Beumer, J.; Puschhof, J.; Yengej, F.Y.; Zhao, L.; Martinez-Silgado, A.; Blotenburg, M.; Begthel, H.; Boot, C.; van Oudenaarden, A.; Chen, Y.G.; et al. BMP gradient along the intestinal villus axis controls zonated enterocyte and goblet cell states. Cell Rep. 2022, 38, 110438. [Google Scholar] [CrossRef] [PubMed]
- Snoeck, V.; Goddeeris, B.; Cox, E. The role of enterocytes in the intestinal barrier function and antigen uptake. Microbes Infect. 2005, 7, 997–1004. [Google Scholar] [CrossRef]
- Haber, A.L.; Biton, M.; Rogel, N.; Herbst, R.H.; Shekhar, K.; Smillie, C.; Burgin, G.; Delorey, T.M.; Howitt, M.R.; Katz, Y.; et al. A single-cell survey of the small intestinal epithelium. Nature 2017, 551, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Grun, D.; Lyubimova, A.; Kester, L.; Wiebrands, K.; Basak, O.; Sasaki, N.; Clevers, H.; van Oudenaarden, A. Single-cell messenger RNA sequencing reveals rare intestinal cell types. Nature 2015, 525, 251–255. [Google Scholar] [CrossRef]
- Sklan, D. Development of the digestive tract of poultry. World’s Poult. Sci. J. 2019, 57, 415–428. [Google Scholar] [CrossRef]
- Orito, W.; Ohhira, F.; Ogasawara, M. Gene expression profiles of FABP genes in protochordates, Ciona intestinalis and Branchiostoma belcheri. Cell Tissue Res. 2015, 362, 331–345. [Google Scholar] [CrossRef]
- Hirano, T.; Nishida, H. Developmental fates of larval tissues after metamorphosis in the ascidian, Halocynthia roretzi. II. Origin of endodermal tissues of the juvenile. Dev. Genes. Evol. 2000, 210, 55–63. [Google Scholar] [CrossRef]
- Arenas, A.F.; Osorio-Méndez, J.F.; Gutierrez, A.J.; Gomez-Marin, J.E. Genome-Wide Survey and Evolutionary Analysis of Trypsin Proteases in Apicomplexan Parasites. Genom. Proteom. Bioinform. 2010, 8, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Parkhomchuk, D.; Borodina, T.; Amstislavskiy, V.; Banaru, M.; Hallen, L.; Krobitsch, S.; Lehrach, H.; Soldatov, A. Transcriptome analysis by strand-specific sequencing of complementary DNA. Nucleic Acids Res. 2009, 37, e123. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.W., Jr.; Lucero, R.J.; Foss, A.B. A situation where the Pearson correlation coefficient leads to erroneous assessment of reliability. J. Clin. Psychol. 1962, 18, 95–97. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bie, T.; Cristianini, N.; Demuth, J.P.; Hahn, M.W. CAFE: A computational tool for the study of gene family evolution. Bioinformatics 2006, 22, 1269–1271. [Google Scholar] [CrossRef] [Green Version]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Giron, C.G.; et al. Ensembl 2018. Nucleic Acids Res. 2018, 46, D754–D761. [Google Scholar] [CrossRef]
- Huang, D.; Zhang, B.; Han, T.; Liu, G.; Chen, X.; Zhao, Z.; Feng, J.; Yang, J.; Wang, T. Genome-wide prediction and comparative transcriptomic analysis reveals the G protein-coupled receptors involved in gonadal development of Apostichopus japonicus. Genomics 2021, 113, 967–978. [Google Scholar] [CrossRef]
- Fujihara, Y.; Herberg, S.; Blaha, A.; Panser, K.; Kobayashi, K.; Larasati, T.; Novatchkova, M.; Theussl, H.C.; Olszanska, O.; Ikawa, M.; et al. The conserved fertility factor SPACA4/Bouncer has divergent modes of action in vertebrate fertilization. Proc. Natl. Acad. Sci. USA 2021, 118, e210877718. [Google Scholar] [CrossRef]
- Session, A.M.; Uno, Y.; Kwon, T.; Chapman, J.A.; Toyoda, A.; Takahashi, S.; Fukui, A.; Hikosaka, A.; Suzuki, A.; Kondo, M.; et al. Genome evolution in the allotetraploid frog Xenopus laevis. Nature 2016, 538, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Fagerberg, L.; Hallstrom, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Liu, P.; Liu, F.; Jiang, A.; Qiao, J.; Pu, Z.; Wang, B.; Zhang, J.; Jia, D.; Li, Y.; et al. EDomics: A comprehensive and comparative multi-omics database for animal evo-devo. Nucleic Acids Res. 2023, 51, D913–D923. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Jiang, A.; Yu, H.; Dong, B. Comparative Transcriptomic Analysis Reveals the Functionally Segmented Intestine in Tunicate Ascidian. Int. J. Mol. Sci. 2023, 24, 6270. https://doi.org/10.3390/ijms24076270
Zhang W, Jiang A, Yu H, Dong B. Comparative Transcriptomic Analysis Reveals the Functionally Segmented Intestine in Tunicate Ascidian. International Journal of Molecular Sciences. 2023; 24(7):6270. https://doi.org/10.3390/ijms24076270
Chicago/Turabian StyleZhang, Wei, An Jiang, Haiyan Yu, and Bo Dong. 2023. "Comparative Transcriptomic Analysis Reveals the Functionally Segmented Intestine in Tunicate Ascidian" International Journal of Molecular Sciences 24, no. 7: 6270. https://doi.org/10.3390/ijms24076270
APA StyleZhang, W., Jiang, A., Yu, H., & Dong, B. (2023). Comparative Transcriptomic Analysis Reveals the Functionally Segmented Intestine in Tunicate Ascidian. International Journal of Molecular Sciences, 24(7), 6270. https://doi.org/10.3390/ijms24076270