
Long Non-Coding RNAs (lncRNAs) as Regulators of the PI3K/AKT/mTOR Pathway in Gastric Carcinoma

 ,
,  ,
, 
Abstract
:1. Introduction
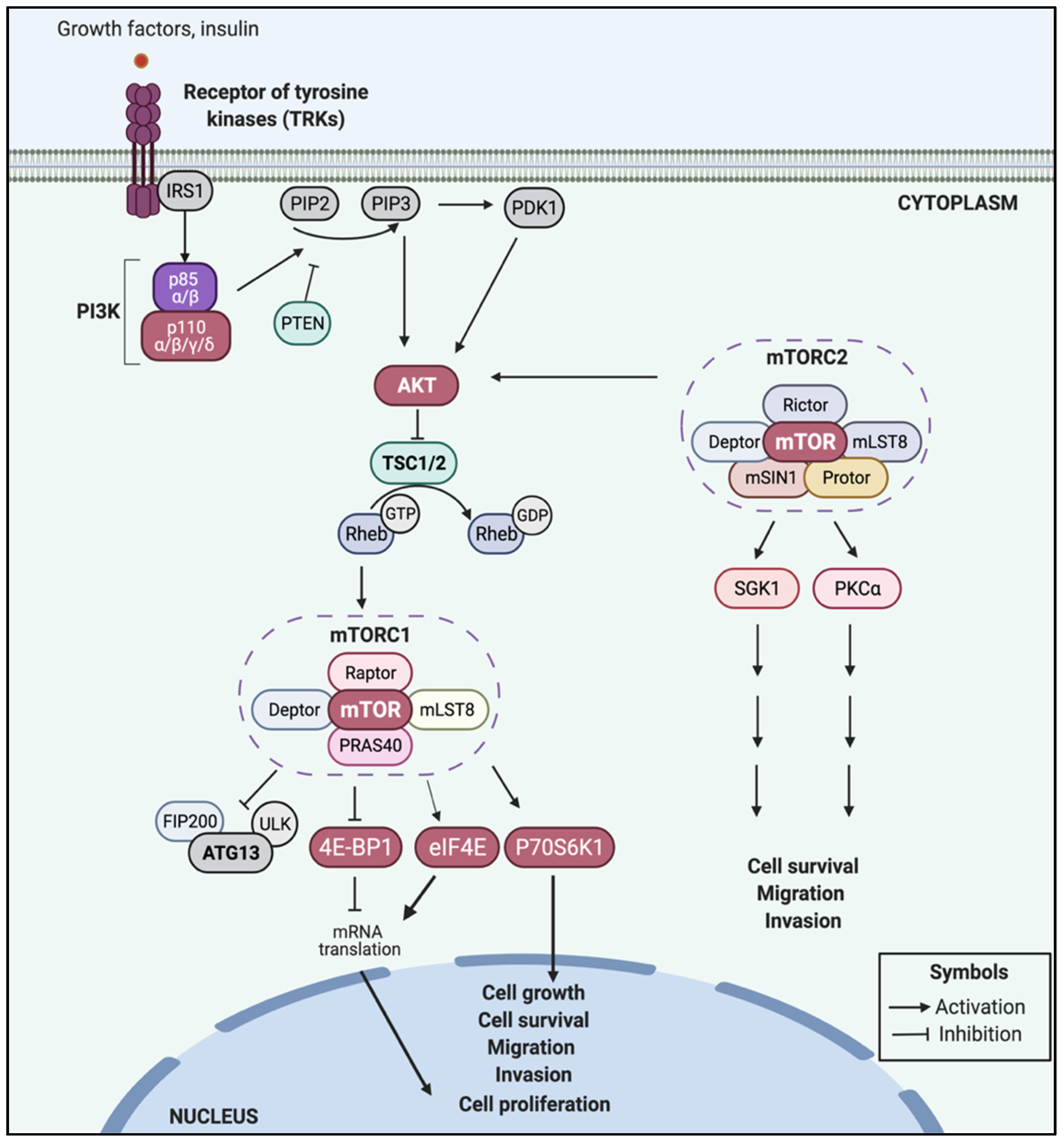
2. The PI3K/AKT/mTOR Signaling Pathway
3. The Role of the PI3K/AKT/mTOR Signaling Pathway in Cancer
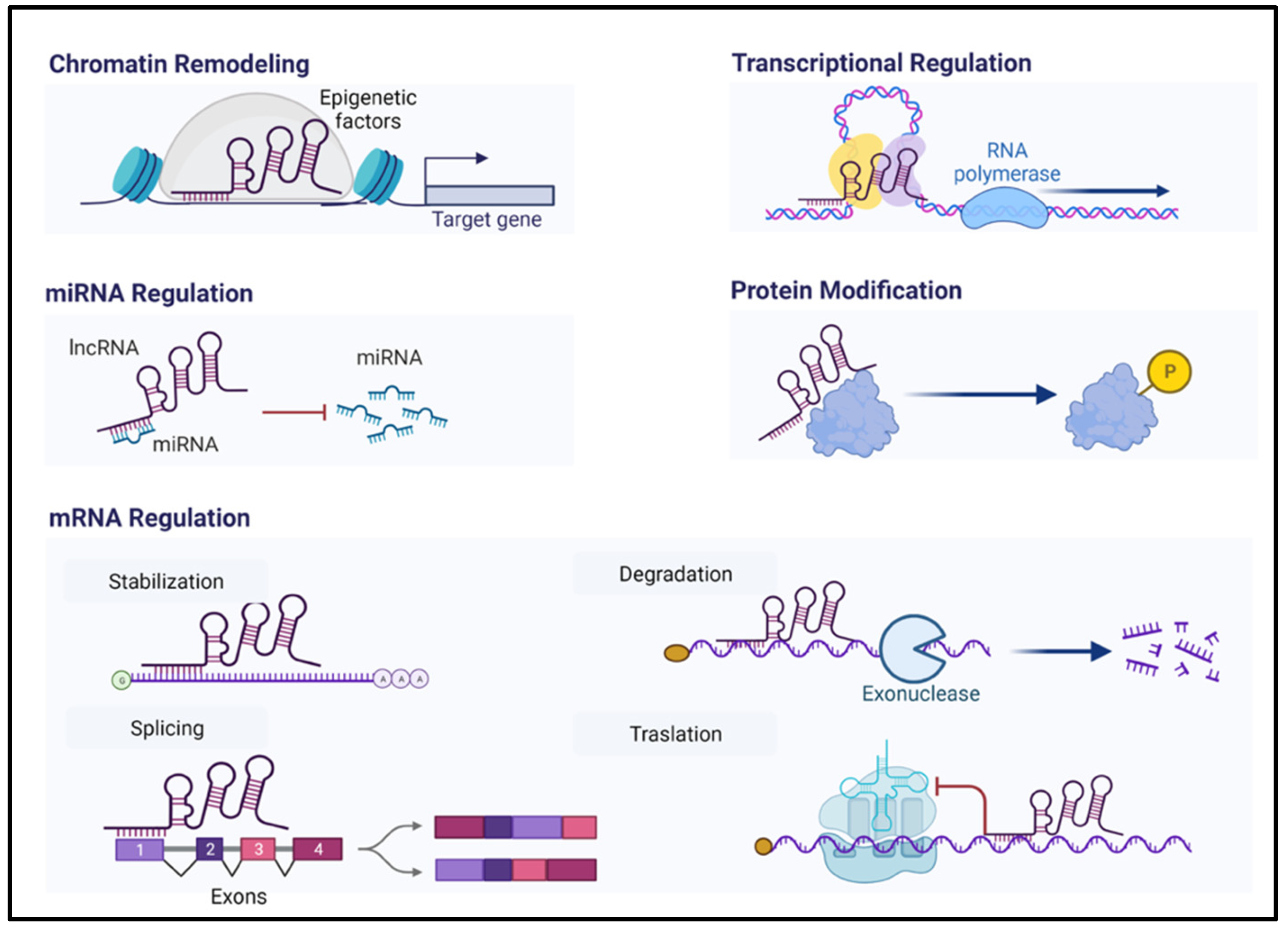
4. Long Non-Coding RNAs (lncRNAs)
5. The Clinical Significance of the PI3K/AKT/mTOR Pathway-Related lncRNAs in GC
6. LncRNAs Involved in the PI3K/AKT/mTOR Pathway Activation in GC
7. LncRNAs That Sponge miRNAs to Activate the PI3K/AKT/mTOR Pathway in GC
8. LncRNAs Involved in Drug Resistance via the PI3K/AKT/mTOR Pathway in GC
9. LncRNAs as Potential Diagnostic, Prognostic, and Therapeutic Markers in GC
10. Experimental and Technical Considerations for lncRNA-Involving Studies
11. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The Ever-Increasing Importance of Cancer as a Leading Cause of Premature Death Worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.P.; Weiderpass, E.; Stewart, B.W. World Cancer Report 2020: Cancer Research for Cancer Precention; International Agency for Research on Cancer: Lyon, France, 2020; Volume 199. [Google Scholar]
- Ministerio de Salud de Chile. Guía Clínica Cáncer Gástrico; MINSAL: Santiago, Chile, 2009.
- Pasechnikov, V.; Chukov, S.; Fedorov, E.; Kikuste, I.; Leja, M. Gastric Cancer: Prevention, Screening and Early Diagnosis. World J. Gastroenterol. 2014, 20, 13842–13862. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.W.; Gill, R.S.; Schiller, D.; Sawyer, M.B. Potential Role of Metabolomics in Diagnosis and Surveillance of Gastric Cancer. World J. Gastroenterol. 2014, 20, 12874–12882. [Google Scholar] [CrossRef]
- Van Cutsem, E.; Sagaert, X.; Topal, B.; Haustermans, K.; Prenen, H. Gastric Cancer. Lancet 2016, 388, 2654–2664. [Google Scholar] [CrossRef]
- Catalano, V.; Labianca, R.; Beretta, G.D.; Gatta, G.; de Braud, F.; van Cutsem, E. Gastric Cancer. Crit. Rev. Oncol. Hematol. 2009, 71, 127–164. [Google Scholar] [CrossRef]
- Sexton, R.E.; al Hallak, M.N.; Diab, M.; Azmi, A.S. Gastric Cancer: A Comprehensive Review of Current and Future Treatment Strategies. Cancer Metastasis Rev. 2020, 39, 1179–1203. [Google Scholar] [CrossRef]
- Mora-Lagos, B.; Cartas-Espinel, I.; Riquelme, I.; Parker, A.C.A.C.; Piccolo, S.R.S.R.; Viscarra, T.; Reyes, M.E.M.E.; Zanella, L.; Buchegger, K.; Ili, C.; et al. Functional and Transcriptomic Characterization of Cisplatin-Resistant AGS and MKN-28 Gastric Cancer Cell Lines. PLoS ONE 2020, 15, e0228331. [Google Scholar] [CrossRef]
- Riquelme, I.; Letelier, P.; Riffo-Campos, A.A.L.; Brebi, P.; Roa, J.C.J. Emerging Role of MiRNAs in the Drug Resistance of Gastric Cancer. Int. J. Mol. Sci. 2016, 17, 424. [Google Scholar] [CrossRef] [Green Version]
- Rivera, F.; Grávalos, C.; Garcia-Carbonero, R. SEOM Clinical Guidelines for the Diagnosis and Treatment of Gastric Adenocarcinoma. Clin. Transl. Oncol. 2012, 14, 528–535. [Google Scholar] [CrossRef]
- Shi, W.-J.; Gao, J.-B. Molecular Mechanisms of Chemoresistance in Gastric Cancer. World J. Gastrointest. Oncol. 2016, 8, 673. [Google Scholar] [CrossRef]
- Holohan, C.; van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer Drug Resistance: An Evolving Paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug Resistance in Cancer: An Overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [Green Version]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Lee, H.S.; Cho, S.B.; Lee, H.E.; Kim, M.A.; Kim, J.H.; Park, J.D.; Kim, J.H.; Yang, H.K.; Lee, B.L.; Kim, W.H. Protein Expression Profiling and Molecular Classification of Gastric Cancer by the Tissue Array Method. Clin. Cancer Res. 2007, 13, 4154–4163. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, I.; Tapia, O.; Espinoza, J.A.; Leal, P.; Buchegger, K.; Sandoval, A.; Bizama, C.; Araya, J.C.; Peek, R.M.; Roa, J.C. The Gene Expression Status of the PI3K/AKT/MTOR Pathway in Gastric Cancer Tissues and Cell Lines. Pathol. Oncol. Res. 2016, 22, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Murugan, A.K. MTOR: Role in Cancer, Metastasis and Drug Resistance. Semin. Cancer Biol. 2019, 59, 92–111. [Google Scholar] [CrossRef]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-Coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef]
- Foster, K.G.; Fingar, D.C. Mammalian Target of Rapamycin (MTOR): Conducting the Cellular Signaling Symphony. J. Biol. Chem. 2010, 285, 14071–14077. [Google Scholar] [CrossRef] [Green Version]
- Caron, E.; Ghosh, S.; Matsuoka, Y.; Ashton-Beaucage, D.; Therrien, M.; Lemieux, S.; Perreault, C.; Roux, P.P.; Kitano, H. A Comprehensive Map of the MTOR Signaling Network. Mol. Syst. Biol. 2010, 6, 453. [Google Scholar] [CrossRef]
- Yoo, Y.A.; Kang, M.H.; Lee, H.J.; Kim, B.; Park, J.K.; Kim, H.K.; Kim, J.S.; Oh, S.C. Sonic Hedgehog Pathway Promotes Metastasis and Lymphangiogenesis via Activation of Akt, EMT, and MMP-9 Pathway in Gastric Cancer. Cancer Res. 2011, 71, 7061–7070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelman, J.A.; Luo, J.; Cantley, L.C. The Evolution of Phosphatidylinositol 3-Kinases as Regulators of Growth and Metabolism. Nat. Rev. Genet. 2006, 7, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Fresno Vara, J.A.; Casado, E.; de Castro, J.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt Signalling Pathway and Cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Arcaro, A.; Guerreiro, A.S. The Phosphoinositide 3-Kinase Pathway in Human Cancer: Genetic Alterations and Therapeutic Implications. Curr. Genom. 2007, 8, 271–306. [Google Scholar] [CrossRef]
- Dibble, C.C.; Cantley, L.C. Regulation of MTORC1 by PI3K Signaling. Trends Cell Biol. 2015, 25, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.-G.; Wang, Q.; Zhou, C.-Z.; Qiu, G.-Q.; Peng, Z.-H.; Tang, H.-M. Mutation Analysis of Tumor Suppressor Gene PTEN in Patients with Gastric Carcinomas and Its Impact on PI3K/AKT Pathway. Oncol. Rep. 2010, 24, 89–95. [Google Scholar]
- Tapia, O.; Riquelme, I.; Leal, P.; Sandoval, A.; Aedo, S.; Weber, H.; Letelier, P.; Bellolio, E.; Villaseca, M.; Garcia, P.; et al. The PI3K/AKT/MTOR Pathway Is Activated in Gastric Cancer with Potential Prognostic and Predictive Significance. Virchows Arch. 2014, 465, 25–33. [Google Scholar] [CrossRef]
- Yuan, H.-X.; Guan, K.-L. Structural Insights of MTOR Complex 1. Cell Res. 2016, 26, 267–268. [Google Scholar] [CrossRef] [Green Version]
- Aylett, C.H.S.; Sauer, E.; Imseng, S.; Boehringer, D.; Hall, M.N.; Ban, N.; Maier, T. Architecture of Human MTOR Complex 1. Science 2015, 351, 48–52. [Google Scholar] [CrossRef]
- Xiao, L.; Wang, Y.C.; Li, W.S.; Du, Y. The Role of MTOR and Phospho-P70S6K in Pathogenesis and Progression of Gastric Carcinomas: An Immunohistochemical Study on Tissue Microarray. J. Exp. Clin. Cancer Res. 2009, 28, 152. [Google Scholar] [CrossRef] [Green Version]
- Berven, L.A.; Willard, F.S.; Crouch, M.F. Role of the P70(S6K) Pathway in Regulating the Actin Cytoskeleton and Cell Migration. Exp. Cell Res. 2004, 296, 183–195. [Google Scholar] [CrossRef]
- Yang, H.Y.; Xue, L.Y.; Xing, L.X.; Wang, J.L.J.; Wang, J.L.J.; Yan, X.; Zhang, X.H. Putative Role of the MTOR/4E-BP1 Signaling Pathway in the Carcinogenesis and Progression of Gastric Cardiac Adenocarcinoma. Mol. Med. Rep. 2013, 7, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Martini, M.; de Santis, M.C.; Braccini, L.; Gulluni, F.; Hirsch, E. PI3K/AKT Signaling Pathway and Cancer: An Updated Review. Ann. Med. 2014, 46, 372–383. [Google Scholar] [CrossRef]
- Dennis, M.D.; Jefferson, L.S.; Kimball, S.R. Role of P70S6K1-Mediated Phosphorylation of EIF4B and PDCD4 Proteins in the Regulation of Protein Synthesis. J. Biol. Chem. 2012, 287, 42890–42899. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Jiang, B.; Zhang, Y. 4E-BP1, a Multifactor Regulated Multifunctional Protein. Cell Cycle 2016, 15, 781–786. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Huang, S. Role of MTOR Signaling in Tumor Cell Motility, Invasion and Metastasis. Curr. Protein Pept. Sci. 2011, 12, 30–42. [Google Scholar]
- Weber, H.; Leal, P.; Stein, S.; Kunkel, H.; García, P.; Bizama, C.; Espinoza, J.A.; Riquelme, I.; Nervi, B.; Araya, J.C.; et al. Rapamycin and WYE-354 Suppress Human Gallbladder Cancer Xenografts in Mice. Oncotarget 2015, 6, 31877–31888. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Graham, P.H.; Hao, J.; Ni, J.; Bucci, J.; Cozzi, P.J.; Kearsley, J.H.; Li, Y. PI3K/Akt/MTOR Pathway Inhibitors Enhance Radiosensitivity in Radioresistant Prostate Cancer Cells through Inducing Apoptosis, Reducing Autophagy, Suppressing NHEJ and HR Repair Pathways. Cell Death Dis. 2014, 5, e1437. [Google Scholar] [CrossRef] [Green Version]
- Fattahi, S.; Amjadi-Moheb, F.; Tabaripour, R.; Ashrafi, G.H.; Akhavan-Niaki, H. PI3K/AKT/MTOR Signaling in Gastric Cancer: Epigenetics and Beyond. Life Sci. 2020, 262, 118513. [Google Scholar] [CrossRef]
- Riquelme, I.; Saavedra, K.; Espinoza, J.A.J.A.; Weber, H.; García, P.; Nervi, B.; Garrido, M.; Corvalán, A.H.A.H.; Roa, J.C.J.C.; Bizama, C.; et al. Molecular Classification of Gastric Cancer: Towards a Pathway-Driven Targeted Therapy. Oncotarget 2015, 5, 24750–24779. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Davies, B.R.; Han, S.; Zhou, M.; Bai, Y.; Zhang, J.; Xu, Y.; Tang, L.; Wang, H.; Liu, Y.J.; et al. The AKT Inhibitor AZD5363 Is Selectively Active in PI3KCA Mutant Gastric Cancer, and Sensitizes a Patient-Derived Gastric Cancer Xenograft Model with PTEN Loss to Taxotere. J. Transl. Med. 2013, 11, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellacosa, A.; Kumar, C.C.; di Cristofano, A.; Testa, J.R. Activation of AKT Kinases in Cancer: Implications for Therapeutic Targeting. Adv. Cancer Res. 2005, 94, 29–86. [Google Scholar] [CrossRef] [PubMed]
- Altomare, D.A.; Testa, J.R. Perturbations of the AKT Signaling Pathway in Human Cancer. Oncogene 2005, 24, 7455–7464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riquelme, I.; Tapia, O.; Leal, P.; Sandoval, A.; Varga, M.G.M.G.; Letelier, P.; Buchegger, K.; Bizama, C.; Espinoza, J.A.J.A.; Peek, R.M.R.M.; et al. MiR-101-2, MiR-125b-2 and MiR-451a Act as Potential Tumor Suppressors in Gastric Cancer through Regulation of the PI3K/AKT/MTOR Pathway. Cell Oncol. 2015, 39, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Fruman, D.A.; Rommel, C. PI3K and Cancer: Lessons, Challenges and Opportunities. Nat. Rev. Drug Discov. 2014, 13, 140–156. [Google Scholar] [CrossRef] [Green Version]
- Polivka, J.; Janku, F. Molecular Targets for Cancer Therapy in the PI3K/AKT/MTOR Pathway. Pharmacol. Ther. 2014, 142, 164–175. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin Signature Reveals over a Thousand Highly Conserved Large Non-Coding RNAs in Mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef]
- Vance, K.W.; Ponting, C.P. Transcriptional Regulatory Functions of Nuclear Long Noncoding RNAs. Trends Genet. 2014, 30, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Dykes, I.M.; Emanueli, C. Transcriptional and Post-Transcriptional Gene Regulation by Long Non-Coding RNA. Genom. Proteom. Bioinform. 2017, 15, 177–186. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long Non-Coding RNAs: Definitions, Functions, Challenges and Recommendations. Nat. Rev. Mol. Cell Biol. 2023, 17, 1–17. [Google Scholar] [CrossRef]
- Huarte, M. The Emerging Role of LncRNAs in Cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2020, 22, 96–118. [Google Scholar] [CrossRef]
- Xu, Q.; Deng, F.; Qin, Y.; Zhao, Z.; Wu, Z.; Xing, Z.; Ji, A.; Wang, Q.J. Long Non-Coding RNA Regulation of Epithelial–Mesenchymal Transition in Cancer Metastasis. Cell Death Dis. 2016, 7, e2254. [Google Scholar] [CrossRef]
- Liu, S.J.; Dang, H.X.; Lim, D.A.; Feng, F.Y.; Maher, C.A. Long Noncoding RNAs in Cancer Metastasis. Nat. Rev. Cancer 2021, 21, 446–460. [Google Scholar] [CrossRef]
- Liu, K.; Gao, L.; Ma, X.; Huang, J.J.; Chen, J.; Zeng, L.; Ashby, C.R.; Zou, C.; Chen, Z.S. Long Non-Coding RNAs Regulate Drug Resistance in Cancer. Mol. Cancer 2020, 19, 54. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Tan, H.Y.; Chan, Y.T.; Jiang, H.; Wang, N.; Wang, D. The Functional Role of Long Noncoding RNA in Resistance to Anticancer Treatment. Ther. Adv. Med. Oncol. 2020, 12, 1–18. [Google Scholar] [CrossRef]
- Zhang, E.; Kong, R.; Yin, D.; You, L.; Sun, M.; Han, L.; Xu, T.; Xia, R.; Yang, J.; De, W.; et al. Long Noncoding RNA ANRIL Indicates a Poor Prognosis of Gastric Cancer and Promotes Tumor Growth by Epigenetically Silencing of MiR-99a/MiR-449a. Oncotarget 2014, 5, 2276–2292. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Bu, P.; Liu, L.; Zhang, X.Z.; Li, J. Overexpression of Long Non-Coding RNA PVT1 in Gastric Cancer Cells Promotes the Development of Multidrug Resistance. Biochem. Biophys. Res. Commun. 2015, 462, 227–232. [Google Scholar] [CrossRef]
- Du, D.-X.; Lian, D.-B.; Amin, B.-H.; Yan, W. Long Non-Coding RNA CRNDE Is a Novel Tumor Promoter by Modulating PI3K/AKT Signal Pathways in Human Gastric Cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5392–5398. [Google Scholar]
- Li, C.; Liang, G.; Yang, S.; Sui, J.; Yao, W.; Shen, X.; Zhang, Y.; Peng, H.; Hong, W.; Xu, S.; et al. Dysregulated LncRNA-UCA1 Contributes to the Progression of Gastric Cancer through Regulation of the PI3K-Akt-MTOR Signaling Pathway. Oncotarget 2017, 8, 93476–93491. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Gu, J.; Lu, H. The GAS5/MiR-222 Axis Regulates Proliferation of Gastric Cancer Cells Through the PTEN/Akt/MTOR Pathway. Dig. Dis. Sci. 2017, 62, 3426–3437. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Liao, Q.; Zou, Z.; Wen, Y.; Wang, J.; Liu, C.; He, Q.; Weng, N.; Zeng, J.; Tang, H.; et al. Long Non-Coding RNA XLOC_006753 Promotes the Development of Multidrug Resistance in Gastric Cancer Cells Through the PI3K/AKT/MTOR Signaling Pathway. Cell. Physiol. Biochem. 2018, 51, 1221–1236. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Wu, P.; Xia, R.; Yang, J.; Huo, X.Y.; Gu, D.Y.; Tang, C.J.; De, W.; Yang, F. STAT3-Induced LncRNA HAGLROS Overexpression Contributes to the Malignant Progression of Gastric Cancer Cells via MTOR Signal-Mediated Inhibition of Autophagy. Mol. Cancer 2018, 17, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, W.; Li, Y.; Zhu, M.; Li, C.; Li, P. LncRNA NORAD Promotes Proliferation And Inhibits Apoptosis Of Gastric Cancer By Regulating MiR-214/Akt/MTOR Axis. OncoTargets Ther. 2019, 12, 8841. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Liang, G.; Yang, S.; Sui, J.; Wu, W.; Xu, S.; Ye, Y.; Shen, B.; Zhang, X.; Zhang, Y. LncRNA-LOC101928316 Contributes to Gastric Cancer Progression through Regulating PI3K-Akt-MTOR Signaling Pathway. Cancer Med. 2019, 8, 4428–4440. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Zhao, X.; Wang, H.; Zheng, M.; Wang, Q.; Chang, W. The Down-Regulation of LncRNA PCAT18 Promotes the Progression of Gastric Cancer via MiR-107/PTEN/PI3K/AKT Signaling Pathway. OncoTargets Ther. 2019, 12, 11017–11031. [Google Scholar] [CrossRef] [Green Version]
- Ba, M.-C.; Ba, Z.; Long, H.; Cui, S.Z.; Gong, Y.F.; Yan, Z.F.; Lin, K.P.; Wu, Y.B.; Tu, Y.N. LncRNA AC093818.1 Accelerates Gastric Cancer Metastasis by Epigenetically Promoting PDK1 Expression. Cell Death Dis. 2020, 11, 64. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhang, Y.; Ding, M.; Xu, R. Long Noncoding RNA TMPO-AS1/MiR-126-5p/BRCC3 Axis Accelerates Gastric Cancer Progression and Angiogenesis via Activating PI3K/Akt/MTOR Pathway. J. Gastroenterol. Hepatol. 2021, 36, 1877–1888. [Google Scholar] [CrossRef]
- Lin, S.; Wang, H.; Yang, W.; Wang, A.; Geng, C. Silencing of Long Non-Coding RNA Colon Cancer-Associated Transcript 2 Inhibits the Growth and Metastasis of Gastric Cancer Through Blocking MTOR Signaling. OncoTargets Ther. 2020, 13, 337. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Tan, X.; Tang, H.; Liu, J. CCL21 Activation of the MALAT1/SRSF1/MTOR Axis Underpins the Development of Gastric Carcinoma. J. Transl. Med. 2021, 19, 210. [Google Scholar] [CrossRef]
- Bai, L.; Dong, K.; Tong, D.; Shi, X.; Wei, S.; Cai, Y. LncRNA HIT000218960 Enhances Resistance to 5-fluorouracil by Promoting HMGA2 and Activating the AKT/MTOR/P70S6K Pathway in Gastric Cancer Cells. Exp. Ther. Med. 2022, 24, 527. [Google Scholar] [CrossRef]
- Chen, W.; Li, Z.; Duan, J.; Liu, Y.; Fang, C.; Bai, Z. LncRNA SNHG6 Inhibits Autophagy of Gastric Carcinoma Cells via PI3K/AKT/MTOR Signaling Pathway. Am. J. Transl. Res. 2022, 14, 2861. [Google Scholar]
- Dong, S.; Zhang, X.; Liu, D. Overexpression of Long Noncoding RNA GAS5 Suppresses Tumorigenesis and Development of Gastric Cancer by Sponging MiR-106a-5p through the Akt/MTOR Pathway. Biol. Open 2019, 8, bio041343. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Tang, L. HDAC3-Mediated LncRNA-LOC101928316 Contributes to Cisplatin Resistance in Gastric Cancer via Activating the PI3K-Akt-MTOR Pathway. Neoplasma 2021, 68, 1043–1051. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, X.; Yin, J.; Tang, J.; Hu, N.; Hong, Y.; Song, Z.; Bian, B.; Wu, F. LncRNA OGFRP1 Promotes Tumor Progression by Activating the AKT/MTOR Pathway in Human Gastric Cancer. Aging 2021, 13, 9766. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, X.; Zhang, T.; Su, L.; Liu, B.; Zhu, Z.; Li, C. LncRNA MALAT1 Promotes Gastric Cancer Progression via Inhibiting Autophagic Flux and Inducing Fibroblast Activation. Cell Death Dis. 2021, 12, 368. [Google Scholar] [CrossRef]
- Wang, B.; Deng, Y.; Jin, J.; Wu, Y.; Shen, L. Long Noncoding RNA LIT3527 Knockdown Induces Apoptosis and Autophagy through Inhibiting MTOR Pathway in Gastric Cancer Cells. J. Cancer 2021, 12, 4901. [Google Scholar] [CrossRef]
- Wu, Q.; Ma, J.; Wei, J.; Meng, W.; Wang, Y.; Shi, M. FOXD1-AS1 Regulates FOXD1 Translation and Promotes Gastric Cancer Progression and Chemoresistance by Activating the PI3K/AKT/MTOR Pathway. Mol. Oncol. 2021, 15, 299–316. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, J.; Wang, Y.; Li, J.; Zhao, L.; Zhang, T.; Liao, X. Long Non-Coding LEF1-AS1 Sponge MiR-5100 Regulates Apoptosis and Autophagy in Gastric Cancer Cells via the MiR-5100/DEK/AMPK-MTOR Axis. Int. J. Mol. Sci. 2022, 23, 4787. [Google Scholar] [CrossRef]
- Dong, C.; Wu, J.; Chen, Y.; Nie, J.; Chen, C. Activation of PI3K/AKT/MTOR Pathway Causes Drug Resistance in Breast Cancer. Front. Pharmacol. 2021, 12, 143. [Google Scholar] [CrossRef]
- Dong, Y.Y.; Zhou, Q.; Li, H.; Lv, Z.; Yuan, Y.; Sun, L.P. Abnormally Expressed LncRNAs as Potential Biomarkers for Gastric Cancer Risk: A Diagnostic Meta-Bioinformatics Analysis. BioMed Res. Int 2022, 2022, 6712625. [Google Scholar] [CrossRef]
- Keller, S.; Ridinger, J.; Rupp, A.-K.; Janssen, J.W.G.; Altevogt, P. Body Fluid Derived Exosomes as a Novel Template for Clinical Diagnostics. J. Transl. Med. 2011, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Sharples, R.A.; Scicluna, B.J.; Hill, A.F. Exosomes Provide a Protective and Enriched Source of MiRNA for Biomarker Profiling Compared to Intracellular and Cell-Free Blood. J. Extracell. Vesicles 2014, 3, 23743. [Google Scholar] [CrossRef] [PubMed]
- Quek, C.; Bellingham, S.A.; Jung, C.-H.; Scicluna, B.J.; Shambrook, M.C.; Sharples, R.A.; Cheng, L.; Hill, A.F. Defining the Purity of Exosomes Required for Diagnostic Profiling of Small RNA Suitable for Biomarker Discovery. RNA Biol. 2017, 14, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Dowdy, S.F. Overcoming Cellular Barriers for RNA Therapeutics. Nat. Biotechnol. 2017, 35, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA Therapeutics—Challenges and Potential Solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Ning, B.; Yu, D.; Yu, A.-M. Advances and Challenges in Studying Noncoding RNA Regulation of Drug Metabolism and Development of RNA Therapeutics. Biochem. Pharmacol. 2019, 169, 113638. [Google Scholar] [CrossRef]
- Shah, S.G.; Rashid, M.; Verma, T.; Ludbe, M.; Khade, B.; Gera, P.B.; Gupta, S. Establishing a Correlation between RIN and A260/280 along with the Multivariate Evaluation of Factors Affecting the Quality of RNA in Cryopreserved Cancer Bio-Specimen. Cell Tissue Bank 2019, 20, 489–499. [Google Scholar] [CrossRef]
- Charles Richard, J.L.; Eichhorn, P.J.A. Platforms for Investigating LncRNA Functions. SLAS Technol. 2018, 23, 493–506. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, I.; Pérez-Moreno, P.; Letelier, P.; Brebi, P.; Roa, J.C. The Emerging Role of PIWI-Interacting RNAs (PiRNAs) in Gastrointestinal Cancers: An Updated Perspective. Cancers 2022, 14, 202. [Google Scholar] [CrossRef]
- Pinkney, H.R.; Wright, B.M.; Diermeier, S.D. The LncRNA Toolkit: Databases and In Silico Tools for LncRNA Analysis. Non-Coding RNA 2020, 6, 49. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| lncRNA/Refs | Expression in GC | Clinicopathological Features |
|---|---|---|
| ANRIL [59] | Up | ANRIL overexpression was associated with greater tumor size, advanced TNM stage, and poor prognosis (OS and DFS) in GC patients. |
| PVT1 [60] | Up | PVT1 overexpression was frequent in GC patients with CDDP-resistant tumors. |
| CRNDE [61] | Up | CRNDE overexpression was correlated with invasion depth, advanced TNM stage, greater lymph node metastasis, and shorter survival (OS) of GC patients. |
| UCA1 [62] | Up | UCA1 overexpression was associated with advanced TNM stage and presence of metastasis in GC patients. |
| GAS5 [63] | Down | GAS5 expression was inversely correlated with TNM stage of tumors in GC patients. |
| XLOC_006753 [64] | Up | XLOC_006753 overexpression was associated with metastasis, advanced TNM stage, tumor size, and poor prognosis (OS and PFS) in GC patients. |
| HAGLROS [65] | Up | HAGLROS overexpression was correlated with increased invasion depth, advanced TNM stage, and poor prognosis (OS) in GC patients. |
| NORAD [66] | Up | NORAD overexpression was correlated with larger tumor size (>5 cm), lymph node metastasis, higher tumor grade, and advanced TNM stage in GC patients. |
| LOC101928316 [67] | Down | LOC101928316 expression was inversely correlated with the differentiation degree and TNM stage of gastric tumors. |
| PCAT18 [68] | Down | PCAT18 expression was inversely associated with tumor size. |
| AC093818.1 [69] | Up | AC093818.1 overexpression was correlated with invasion, lymphatic metastasis, distal metastasis, and advance TNM stage. Additionally, AC093818.1 seems to be useful to differentiate metastatic from non-metastatic GC. |
| TMPO-AS1 [70] | Up | TMPO-AS1 overexpression was correlated with advanced TNM stage, lymph node metastasis, and poorer survival (OS) in GC patients. |
| CCAT2 [71] | Up | CCAT2 overexpression was associated with greater tumor size, presence of lymph node metastasis, advanced TNM staging, and lower survival (OS) in GC patients. |
| MALAT1 [72] | Up | MALAT1 was significantly associated with worse pathological stage, differentiation degree, and presence of lymph node metastasis. |
| HIT000218960 [73] | Up | HIT000218960 expression was inversely correlated with the response to 5-FU in GC patients. |
| SNHG6 [74] | Up | SNHG6 overexpression was associated with lower survival (PPS) of patients, and showed a significant predictive value for the development of GC and the death of patients. Serum SNHG6 levels could be a promising prognostic marker because it would allow monitoring of GC patients before and after therapy. |
| lncRNA/Refs | Expression in GC | Phenotypic Features | Molecular Mechanism on the PI3K/AKT/mTOR Pathway |
|---|---|---|---|
| ANRIL [59] | Up | P, T | ANRIL modulates the expression of miR-99a/miR-449a by binding to PRC2, thus regulating mTOR and CDK6 pathways, thereby controlling GC cell proliferation. |
| PVT1 [60] | Up | A, DR | PVT1 induces the upregulation of MDR1, MRP, mTOR, and HIF-1A genes in GC cells. * |
| CRNDE [61] | Up | P, M, I | CRNDE induces higher expression levels of p-PI3K and p-AKT in GC cells. * |
| UCA1 [62] | Up | P, A, M, I, T | UCA1 overexpression increased the expression of AKT3, p-AKT3, p-mTOR, and P70S6K1, and inhibited the expression of EIF4E. * |
| GAS5 [63,75] | Down | P, A, M, I, T | GAS5 downregulation increases miR-222 levels, thereby inducing PTEN repression and the subsequent overexpression of p-AKT and p-mTOR. Similarly, GAS5 downregulation increases miR-106a-5p levels, which also induces the overexpression of p-Akt and p-mTOR. * |
| XLOC_006753 [64] | Up | P, V, A, CCP, DR, M, EMT | XLOC_006753 overexpression induces the higher expression levels of PI3K, p-AKT, p-mTOR, P70S6K1, p-P70S6K1, and p-4E-BP1. * |
| HAGLROS [65] | Up | P, M, T | HAGLROS sponges miR-100-5p to produce an increase of mTOR mRNA expression. HAGLROS also interacts with mTORC1 components to induce an inhibition of autophagy, thereby promoting proliferation and malignant phenotype of GC cells. |
| NORAD [66] | Up | P, A, T | NORAD sponges miR-214 to finally produce an increase of p-AKT and p-mTOR levels. |
| LOC101928316 [67,76] | Down | P, M, I, DR, T | LOC101928316 downregulation induces repression of PTEN protein and, therefore, higher expression of PI3K, p-AKT, mTOR, and p-mTOR in vitro, and higher expression of p-AKT and p-mTOR in vivo. * |
| PCAT18 [68] | Down | V, A, CCP, T | PCAT18 downregulation increases miR-107 levels, which represses PTEN to activate the Akt/mTOR pathway via producing overexpression of p-PI3K and p-AKT. |
| AC093818.1 [69] | Up | M, I | In DNA, the AC093818.1 sequence overlaps on the promotor sequence of PDK1 gene; thus, this lncRNA could induce a higher expression of PDK1. In addition, AC093818.1 binds transcription factors STAT3 and SP1 to also induce a higher expression of PDK1. Therefore, AC093818.1 overexpression finally increases the levels of PDK1 and its downstream targets p-AKT1 and p-mTOR in GC cells. |
| TMPO-AS1 [70] | Up | P, M, ANG | TMPO-AS1 overexpression decreases miR-126-5p levels, which increases BRCC3 expression that subsequently would induce activation of the PI3K/Akt/mTOR pathway via producing overexpression of p-PI3K, p-AKT, and p-mTOR. * |
| CCAT2 [71] | Up | P, A, CCP, ATG | CCAT2 overexpression would induce higher levels of p-AKT, p-mTOR, and p-P70S6K1. * |
| NEAT1 [77] | Up | P, A, M, I | NEAT1 sponges miR-1294 to finally produce an increase in the AKT1 mRNA levels. Additionally, NEAT1 induces lower PTEN expression and higher levels of AKT, p-AKT, and P70S6K1. * |
| MALAT1 [72,78] | Up | M, I, EMT, A, ATG, T | The CCL21 gene expression increases MALAT1 levels, which then reduces miR-202-3p levels, inciting the SRSF1 upregulation. This last event triggers p-mTOR overexpression. * MALAT1 inhibits PTEN mRNA expression and this event could induce a greater downstream activation of the PI3K/Akt/mTOR pathway. |
| LIT3527 [79] | Up | P, V, A, CD, ATG, M, MET | LIT3527 upregulation could induce higher levels of AKT, ERK, p-mTOR, and p-4E-BP1. * |
| FOXD1-AS1 [80] | Up | P, V, A, M, I, DR, T, MET | FOXD1-AS1 increases FOXD1 expression through strengthening eIF4G- eIF4E interaction via phosphorylation of 4E-BP1. FOXD1-AS1 also sponges miR-466 to thereby produce an increase of PIK3CA expression. |
| OGFRP1 [77] | Up | P, A, CCP, M, EMT, T | OGFRP1 overexpression would induce higher levels of p-AKT and p-mTOR. * |
| HIT000218960 [73] | Up | A, DR | HIT000218960 overexpression induces increase of HMGA2 levels, which also triggers higher expression of p-AKT, p-mTOR, and p-P70S6K1. * |
| SNHG6 [74] | Up | V, A, ATG, M | SNHG6 overexpression could induce higher protein levels of PI3K, p-PI3K, AKT, p-AKT, mTOR, and p-mTOR, and lower protein levels of Beclin1 and LC3. * |
| LEF1-AS1 [81] | --- | A, ATG, T, MET | LEF1-AS1 sponges miR-5100, inducing a higher DEK expression that subsequently triggers increased levels of p-mTOR. * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riquelme, I.; Pérez-Moreno, P.; Mora-Lagos, B.; Ili, C.; Brebi, P.; Roa, J.C. Long Non-Coding RNAs (lncRNAs) as Regulators of the PI3K/AKT/mTOR Pathway in Gastric Carcinoma. Int. J. Mol. Sci. 2023, 24, 6294. https://doi.org/10.3390/ijms24076294
Riquelme I, Pérez-Moreno P, Mora-Lagos B, Ili C, Brebi P, Roa JC. Long Non-Coding RNAs (lncRNAs) as Regulators of the PI3K/AKT/mTOR Pathway in Gastric Carcinoma. International Journal of Molecular Sciences. 2023; 24(7):6294. https://doi.org/10.3390/ijms24076294
Chicago/Turabian StyleRiquelme, Ismael, Pablo Pérez-Moreno, Bárbara Mora-Lagos, Carmen Ili, Priscilla Brebi, and Juan Carlos Roa. 2023. "Long Non-Coding RNAs (lncRNAs) as Regulators of the PI3K/AKT/mTOR Pathway in Gastric Carcinoma" International Journal of Molecular Sciences 24, no. 7: 6294. https://doi.org/10.3390/ijms24076294
APA StyleRiquelme, I., Pérez-Moreno, P., Mora-Lagos, B., Ili, C., Brebi, P., & Roa, J. C. (2023). Long Non-Coding RNAs (lncRNAs) as Regulators of the PI3K/AKT/mTOR Pathway in Gastric Carcinoma. International Journal of Molecular Sciences, 24(7), 6294. https://doi.org/10.3390/ijms24076294