
Notch-RBPJ Pathway for the Differentiation of Bone Marrow Mesenchymal Stem Cells in Femoral Head Necrosis

Abstract
:1. Introduction
2. Results
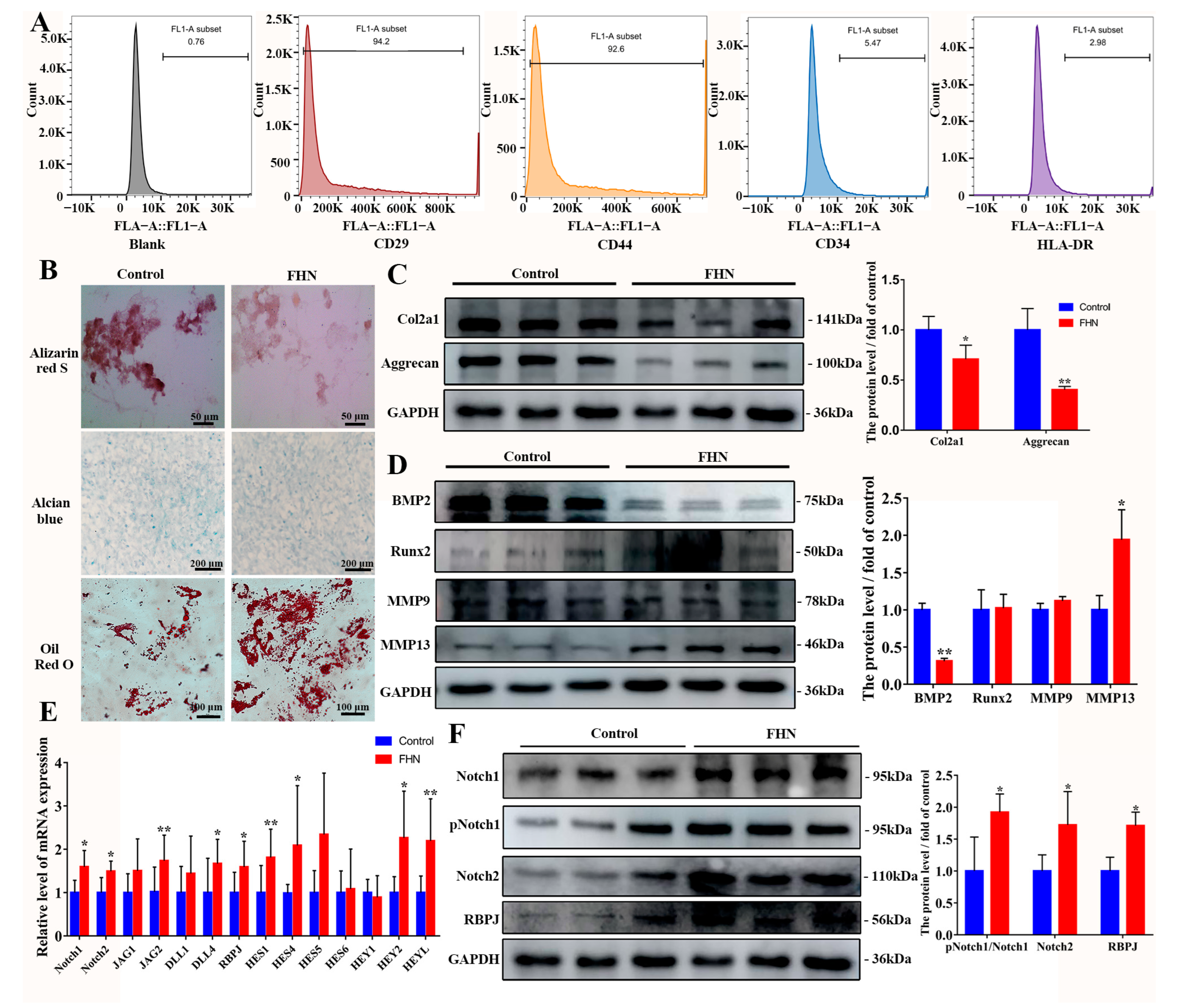
2.1. Decreased Osteogenic and Chondrogenic Differentiation and Increased Lipogenic Capacity of BMSCs in Spontaneous FHN Broiler Femur Tissue
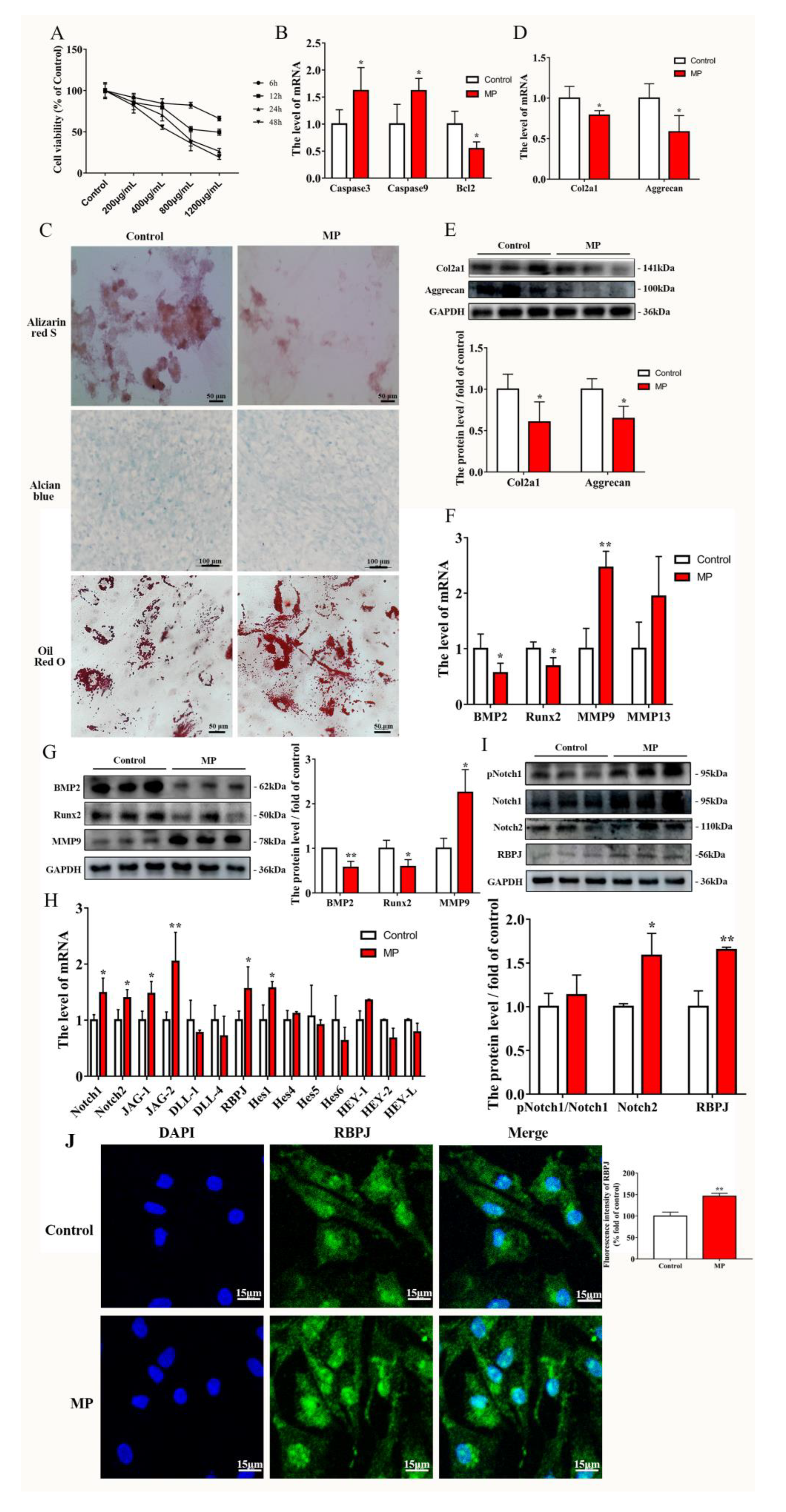
2.2. MP Causes the Homeostatic Imbalance of BMSCs and Activation of the Notch Pathway
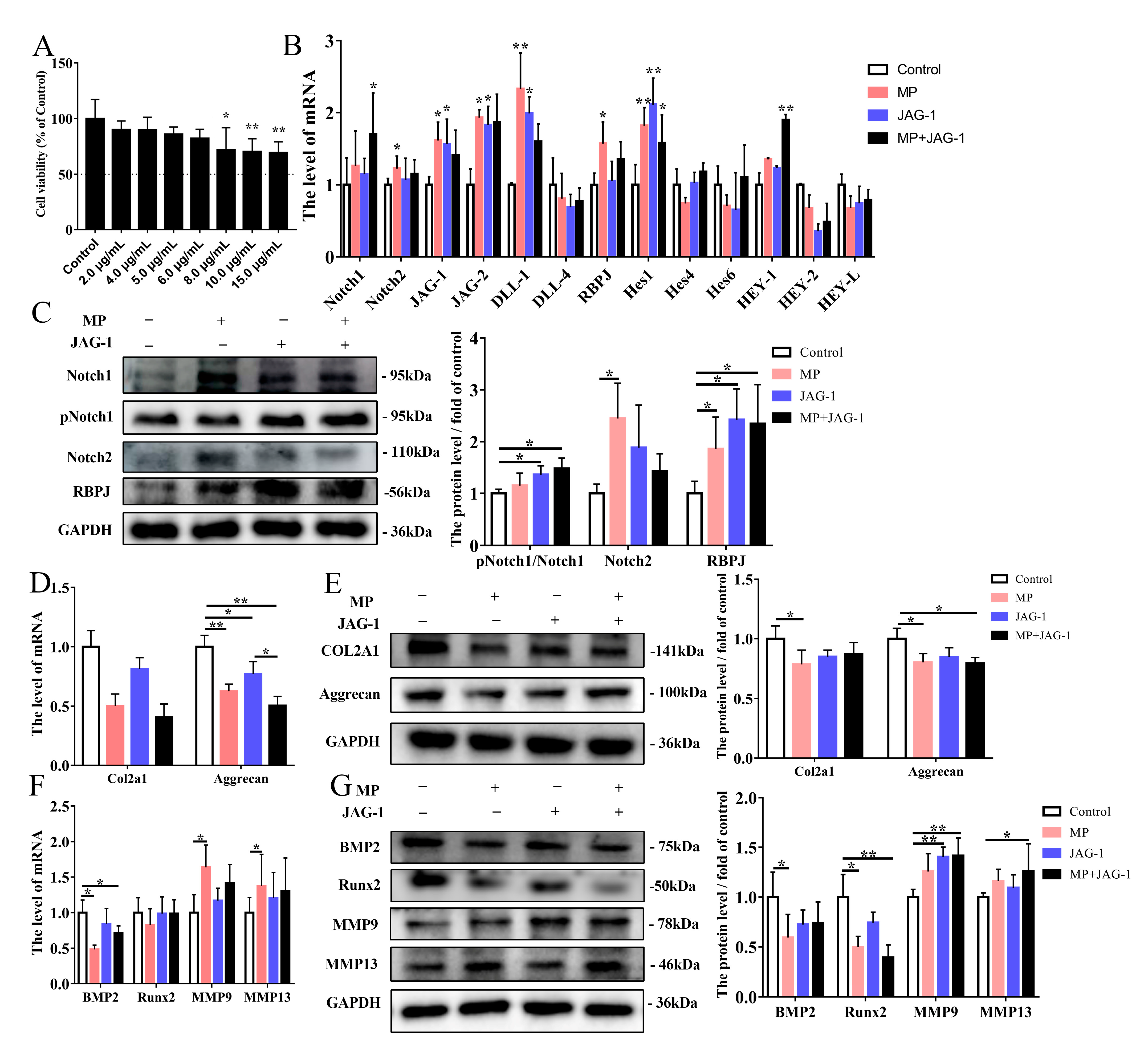
2.3. Regulation of MP-Induced Cellular Osteogenesis and Chondrogenesis Metabolism by the Notch Pathway
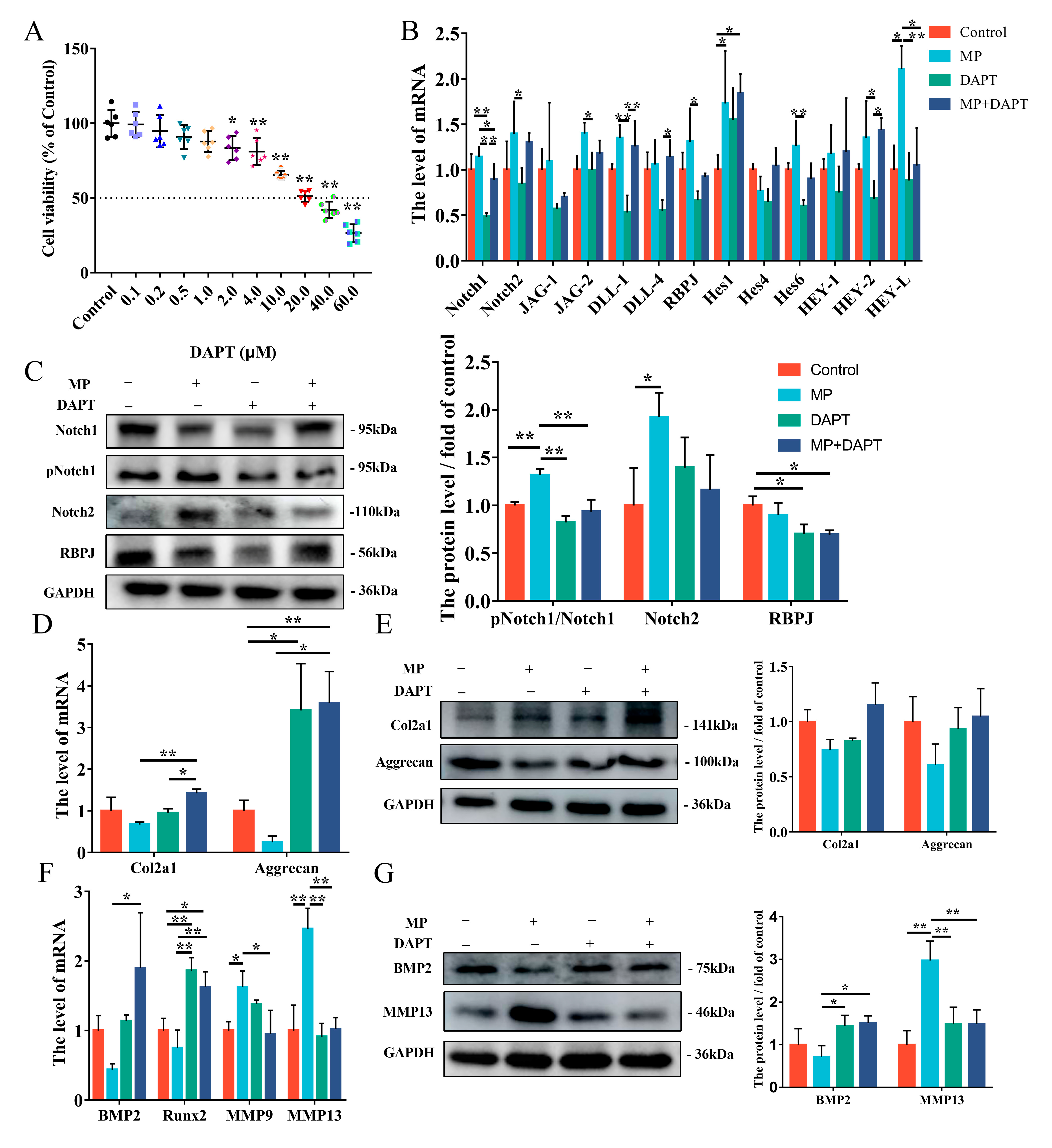
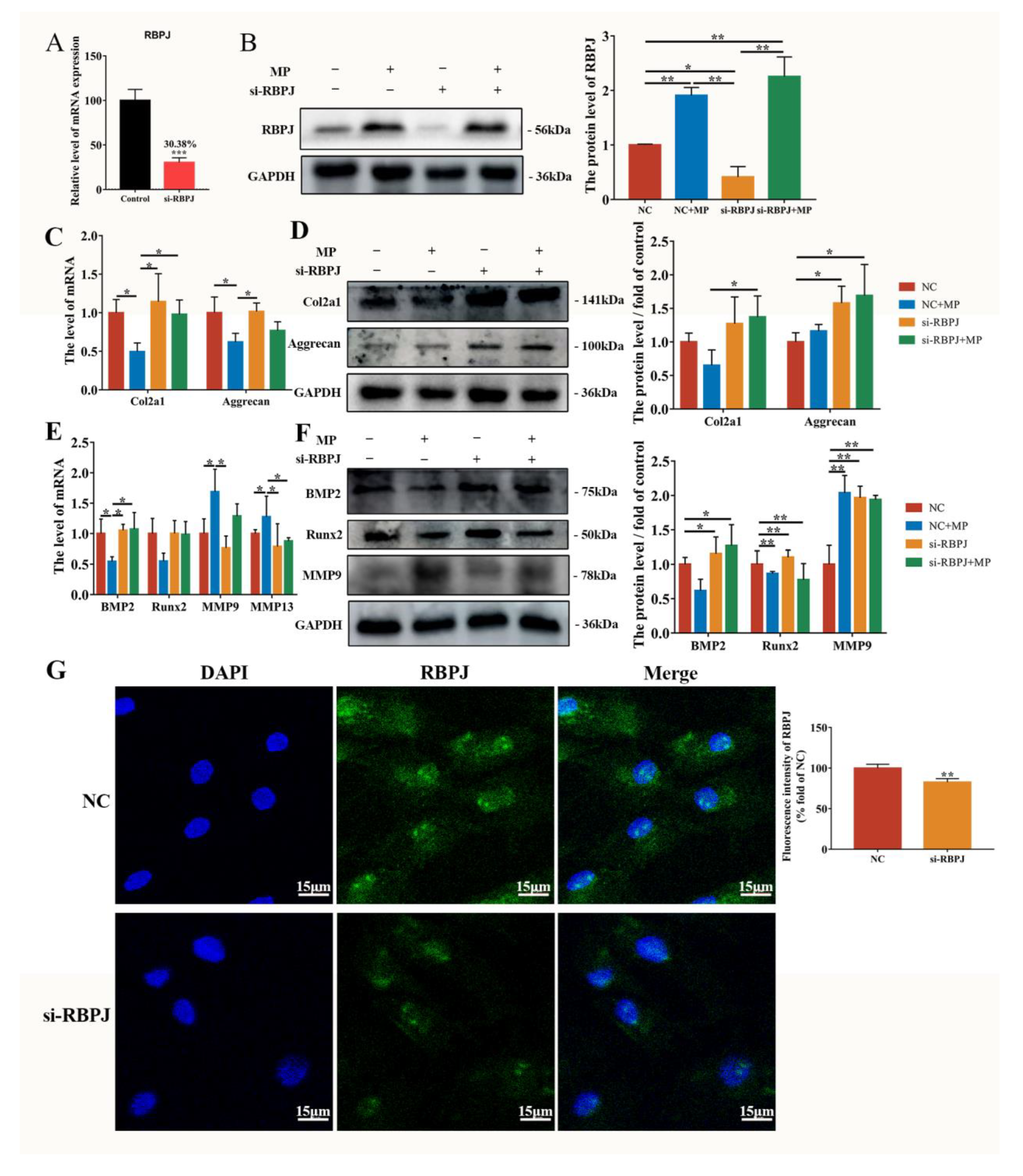
2.4. MP-Induced Alterations in BMSCs Differentiation Are Mitigated by si-RBPJ
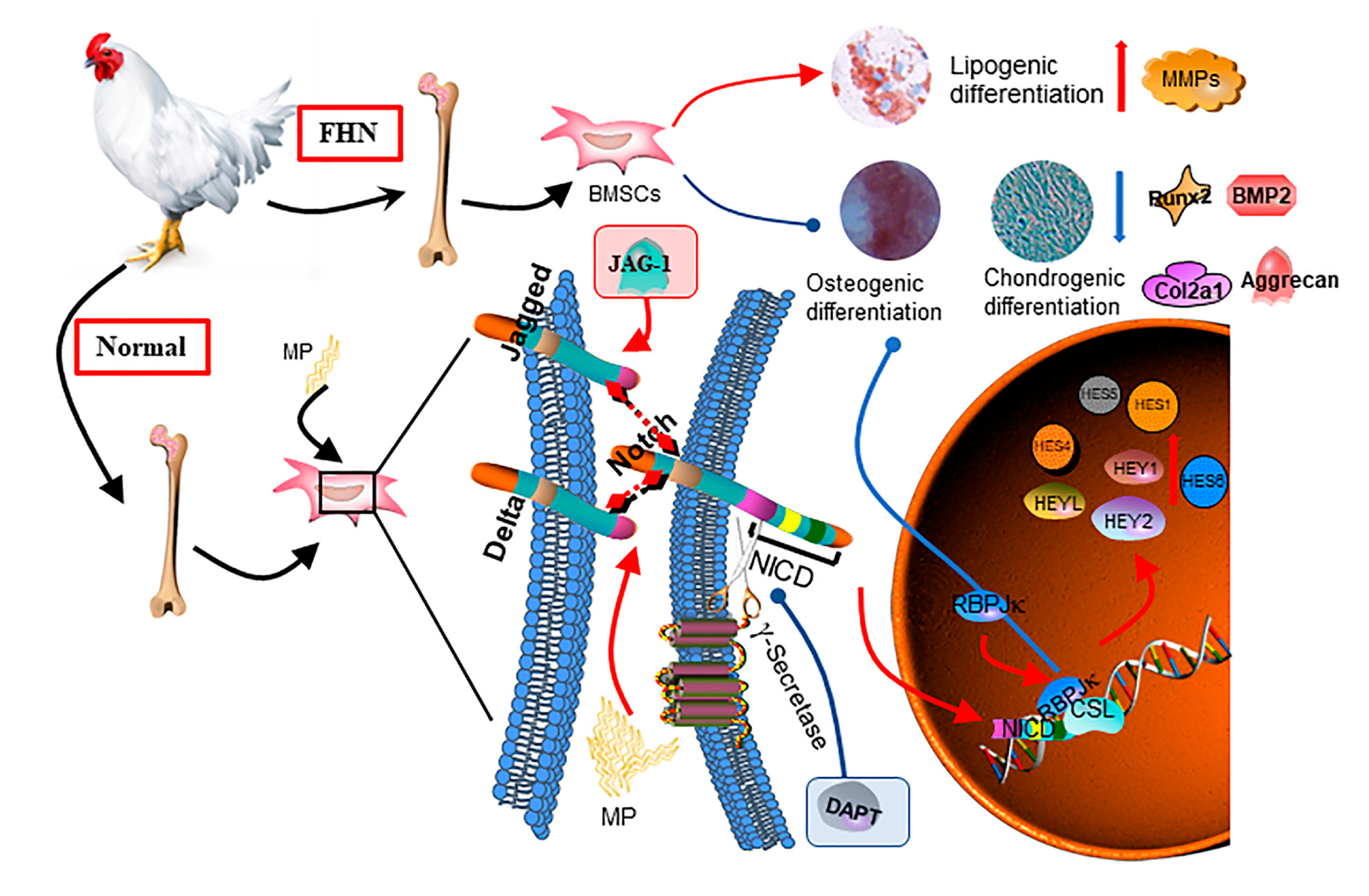
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Isolation, Culture, and Identification of Primary BMSCs
4.3. Cell Treatment
4.4. Cell Viability Assay
4.5. Osteogenesis Differentiation
4.6. Lipogenic Differentiation
4.7. Chondrogenic Differentiation
4.8. Alizarin Red S (ARS) Staining
4.9. Oil Red O Staining
4.10. Alcian Blue Staining
4.11. Flow Cytometry Analysis of the Surface Markers
4.12. RNA Isolation and qRT-RCR
4.13. Western Blot
4.14. Immunofluorescence
4.15. Cell Transfection
4.16. Statistics Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuang, M.J.; Huang, Y.; Zhao, X.G.; Zhang, R.; Ma, J.X.; Wang, D.C.; Ma, X.L. Exosomes derived from Wharton’s jelly of human umbilical cord mesenchymal stem cells reduce osteocyte apoptosis in glucocorticoid-induced osteonecrosis of the femoral head in rats via the miR-21-PTEN-AKT signalling pathway. Int. J. Biol. Sci. 2019, 15, 1861–1871. [Google Scholar] [CrossRef]
- Lee, K.S.; Lee, J.; Kim, H.K.; Yeom, S.H.; Woo, C.H.; Jung, Y.J.; Yun, Y.E.; Park, S.Y.; Han, J.; Kim, E.; et al. Extracellular vesicles from adipose tissue-derived stem cells alleviate osteoporosis through osteoprotegerin and miR-21-5p. J. Extracell. Vesicles 2021, 10, e12152. [Google Scholar] [CrossRef]
- Takam Kamga, P.; Bazzoni, R.; Dal Collo, G.; Cassaro, A.; Tanasi, I.; Russignan, A.; Tecchio, C.; Krampera, M. The Role of Notch and Wnt Signaling in MSC Communication in Normal and Leukemic Bone Marrow Niche. Front. Cell Dev. Biol. 2020, 8, 599276. [Google Scholar] [CrossRef]
- Malizos, K.N.; Quarles, L.D.; Seaber, A.V.; Rizk, W.S.; Urbaniak, J.R. An experimental canine model of osteonecrosis: Characterization of the repair process. J. Orthop. Res. 1993, 11, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, F.; Connolly, S.; Zurakowski, D.; Menezes, N.; Olear, E.; Jimenez, M.; Flynn, E.; Jaramillo, D. Femoral head deformation and repair following induction of ischemic necrosis: A histologic and magnetic resonance imaging study in the piglet. J. Bone Jt. Surgery. Am. Vol. 2009, 91, 2903–2914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernigou, P.; Beaujean, F.; Lambotte, J.C. Decrease in the mesenchymal stem-cell pool in the proximal femur in corticosteroid-induced osteonecrosis. J. Bone Jt. Surgery. Br. Vol. 1999, 81, 349–355. [Google Scholar] [CrossRef]
- Hernigou, P.; Beaujean, F. Abnormalities in the bone marrow of the iliac crest in patients who have osteonecrosis secondary to corticosteroid therapy or alcohol abuse. J. Bone Jt. Surgery. Am. Vol. 1997, 79, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Chen, Y.X.; Yin, J.H.; Tao, S.C.; Guo, S.C.; Wei, Z.Y.; Feng, Y.; Zhang, C.Q. Valproic acid prevents glucocorticoid-induced osteonecrosis of the femoral head of rats. Int. J. Mol. Med. 2018, 41, 3433–3447. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, J.S.; Roh, H.L.; Kim, C.H.; Jung, J.S.; Suh, K.T. Alterations in the differentiation ability of mesenchymal stem cells in patients with nontraumatic osteonecrosis of the femoral head: Comparative analysis according to the risk factor. J. Orthop. Res. 2006, 24, 604–609. [Google Scholar] [CrossRef] [Green Version]
- Houdek, M.T.; Wyles, C.C.; Packard, B.D.; Terzic, A.; Behfar, A.; Sierra, R.J. Decreased Osteogenic Activity of Mesenchymal Stem Cells in Patients With Corticosteroid-Induced Osteonecrosis of the Femoral Head. J. Arthroplast. 2016, 31, 893–898. [Google Scholar] [CrossRef]
- Wang, B.L.; Sun, W.; Shi, Z.C.; Lou, J.N.; Zhang, N.F.; Shi, S.H.; Guo, W.S.; Cheng, L.M.; Ye, L.Y.; Zhang, W.J.; et al. Decreased proliferation of mesenchymal stem cells in corticosteroid-induced osteonecrosis of femoral head. Orthopedics 2008, 31, 444. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.A.; Zheng, H.X.; Wang, C.W.; Shi, D.; Li, J.J. Influence of glucocorticoids on the osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells. BMC Musculoskelet. Disord. 2014, 15, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Yang, N.; Shi, X.M. Regulation of mesenchymal stem cell osteogenic differentiation by glucocorticoid-induced leucine zipper (GILZ). J. Biol. Chem. 2008, 283, 4723–4729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch signaling: Cell fate control and signal integration in development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S. Notch signaling in stem cell systems. Stem Cells 2006, 24, 2437–2447. [Google Scholar] [CrossRef]
- Lai, E.C. Notch signaling: Control of cell communication and cell fate. Development 2004, 131, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Shang, X.; Zhang, H.; Wang, G.; Massey, P.A.; Barton, S.R.; Kevil, C.G.; Dong, Y. Notch Signaling in Osteogenesis, Osteoclastogenesis, and Angiogenesis. Am. J. Pathol. 2019, 189, 1495–1500. [Google Scholar] [CrossRef] [Green Version]
- Sprinzak, D.; Blacklow, S.C. Biophysics of Notch Signaling. Annu. Rev. Biophys. 2021, 50, 157–189. [Google Scholar] [CrossRef]
- van Tetering, G.; Vooijs, M. Proteolytic cleavage of Notch: “HIT and RUN”. Curr. Mol. Med. 2011, 11, 255–269. [Google Scholar] [CrossRef]
- Dong, Y.; Long, T.; Wang, C.; Mirando, A.J.; Chen, J.; O’Keefe, R.J.; Hilton, M.J. NOTCH-Mediated Maintenance and Expansion of Human Bone Marrow Stromal/Stem Cells: A Technology Designed for Orthopedic Regenerative Medicine. Stem Cells Transl. Med. 2014, 3, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Chia, W.; Liu, J.; Huang, Y.G.; Zhang, C. A circular RNA derived from DAB1 promotes cell proliferation and osteogenic differentiation of BMSCs via RBPJ/DAB1 axis. Cell Death Dis. 2020, 11, 372. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, J.B.; Andrade, D.M.; Santiago, M.B. The use of bisphosphonate in the treatment of avascular necrosis: A systematic review. Clin. Rheumatol. 2008, 27, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Fan, R.; Zhou, Z. Endoplasmic reticulum stress, chondrocyte apoptosis and oxidative stress in cartilage of broilers affected by spontaneous femoral head necrosis. Poult. Sci. 2021, 100, 101258. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Wang, K.; Wang, L.; Zhou, Z. Changes of lipid and bone metabolism in broilers with spontaneous femoral head necrosis. Poult. Sci. 2021, 100, 100808. [Google Scholar] [CrossRef]
- Fan, R.; Liu, K.; Zhou, Z. Abnormal Lipid Profile in Fast-Growing Broilers With Spontaneous Femoral Head Necrosis. Front. Physiol. 2021, 12, 685968. [Google Scholar] [CrossRef]
- Plotkin, L.I.; Manolagas, S.C.; Bellido, T. Glucocorticoids induce osteocyte apoptosis by blocking focal adhesion kinase-mediated survival. Evidence for inside-out signaling leading to anoikis. J. Biol. Chem. 2007, 282, 24120–24130. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Molecular Mechanism of Runx2-Dependent Bone Development. Mol. Cells 2020, 43, 168–175. [Google Scholar] [CrossRef]
- Bruderer, M.; Richards, R.G.; Alini, M.; Stoddart, M.J. Role and regulation of RUNX2 in osteogenesis. Eur. Cells Mater. 2014, 28, 269–286. [Google Scholar] [CrossRef]
- Huang, Z.; Ren, P.G.; Ma, T.; Smith, R.L.; Goodman, S.B. Modulating osteogenesis of mesenchymal stem cells by modifying growth factor availability. Cytokine 2010, 51, 305–310. [Google Scholar] [CrossRef]
- Noël, D.; Gazit, D.; Bouquet, C.; Apparailly, F.; Bony, C.; Plence, P.; Millet, V.; Turgeman, G.; Perricaudet, M.; Sany, J.; et al. Short-term BMP-2 expression is sufficient for in vivo osteochondral differentiation of mesenchymal stem cells. Stem Cells 2004, 22, 74–85. [Google Scholar] [CrossRef]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Wei, B.; Li, J.; Huang, C.; Lin, R.; Tang, C.; Xu, Y.; Yao, Q.; Wang, L. Investigations of Cartilage Matrix Degeneration in Patients with Early-Stage Femoral Head Necrosis. Med. Sci. Monit. 2017, 23, 5783–5792. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.; Xing, R.; Liu, J.; Xing, F. Role of CSL-dependent and independent Notch signaling pathways in cell apoptosis. Apoptosis 2016, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Xi, J.; Cheng, Y.; Sun, D.; Shu, P.; Chi, S.; Tian, S.; Ye, S. Reprogrammed mesenchymal stem cells derived from iPSCs promote bone repair in steroid-associated osteonecrosis of the femoral head. Stem Cell Res. Ther. 2021, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Osathanon, T.; Manokawinchoke, J.; Sa-Ard-Iam, N.; Mahanonda, R.; Pavasant, P.; Suwanwela, J. Jagged1 promotes mineralization in human bone-derived cells. Arch. Oral Biol. 2019, 99, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Tocados, J.M.; Herencia, C.; Martínez-Moreno, J.M.; Montes de Oca, A.; Rodríguez-Ortiz, M.E.; Vergara, N.; Blanco, A.; Steppan, S.; Almadén, Y.; Rodríguez, M.; et al. Magnesium Chloride promotes Osteogenesis through Notch signaling activation and expansion of Mesenchymal Stem Cells. Sci. Rep. 2017, 7, 7839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilton, M.J.; Tu, X.; Wu, X.; Bai, S.; Zhao, H.; Kobayashi, T.; Kronenberg, H.M.; Teitelbaum, S.L.; Ross, F.P.; Kopan, R.; et al. Notch signaling maintains bone marrow mesenchymal progenitors by suppressing osteoblast differentiation. Nat. Med. 2008, 14, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Engin, F.; Yao, Z.; Yang, T.; Zhou, G.; Bertin, T.; Jiang, M.M.; Chen, Y.; Wang, L.; Zheng, H.; Sutton, R.E.; et al. Dimorphic effects of Notch signaling in bone homeostasis. Nat. Med. 2008, 14, 299–305. [Google Scholar] [CrossRef]
- Shang, X.; Luo, Z.; Wang, X.; Jaeblon, T.; Marymont, J.V.; Dong, Y. Deletion of RBPJK in Mesenchymal Stem Cells Enhances Osteogenic Activity by Up-Regulation of BMP Signaling. PLoS ONE 2015, 10, e0135971. [Google Scholar] [CrossRef]
- Wagley, Y.; Chesi, A.; Acevedo, P.K.; Lu, S.; Wells, A.D.; Johnson, M.E.; Grant, S.F.A.; Hankenson, K.D. Canonical Notch signaling is required for bone morphogenetic protein-mediated human osteoblast differentiation. Stem Cells 2020, 38, 1332–1347. [Google Scholar] [CrossRef]
- Novak, S.; Roeder, E.; Sinder, B.P.; Adams, D.J.; Siebel, C.W.; Grcevic, D.; Hankenson, K.D.; Matthews, B.G.; Kalajzic, I. Modulation of Notch1 signaling regulates bone fracture healing. J. Orthop. Res. 2020, 38, 2350–2361. [Google Scholar] [CrossRef]
- Oldershaw, R.A.; Tew, S.R.; Russell, A.M.; Meade, K.; Hawkins, R.; McKay, T.R.; Brennan, K.R.; Hardingham, T.E. Notch signaling through Jagged-1 is necessary to initiate chondrogenesis in human bone marrow stromal cells but must be switched off to complete chondrogenesis. Stem Cells 2008, 26, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, C.; Safarova, A.; Smith, M.; Chung, R.; Bruyneel, A.A.N.; Gomez-Galeno, J.; Oswald, F.; Larson, C.J.; Cashman, J.R.; Ruiz-Lozano, P.; et al. Disruption of NOTCH signaling by a small molecule inhibitor of the transcription factor RBPJ. Sci. Rep. 2019, 9, 10811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Wang, J.; Liu, Q.; Li, J.; Zhang, Q.; Zhuang, Z.; Yao, X.; Liu, C.; Li, Y.; Cao, L.; et al. DAPT, a γ-Secretase Inhibitor, Suppresses Tumorigenesis, and Progression of Growth Hormone-Producing Adenomas by Targeting Notch Signaling. Front. Oncol. 2019, 9, 809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giaimo, B.D.; Gagliani, E.K.; Kovall, R.A.; Borggrefe, T. Transcription Factor RBPJ as a Molecular Switch in Regulating the Notch Response. Adv. Exp. Med. Biol. 2021, 1287, 9–30. [Google Scholar] [CrossRef]
- Liu, K.; Chen, P.; Lu, J.; Zhu, Y.; Xu, Y.; Liu, Y.; Liu, J. Protective Effect of Purple Tomato Anthocyanidin on Chromium(VI)-Induced Autophagy in LMH Cells by Inhibiting Endoplasmic Reticulum Stress. Biol. Trace Elem. Res. 2020, 194, 570–580. [Google Scholar] [CrossRef]
- Liu, K.; Cui, Y.; Li, H.; Qi, C.; Cheng, G.; Gao, X.; Zhang, Z.; Liu, Y.; Liu, J. Hydrogen-Rich Medium Regulates Cr(VI)-Induced ER Stress and Autophagy Signaling in DF-1 Cells. Biol. Trace Elem. Res. 2022, 200, 2329–2337. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence (5’-3’) |
|---|---|
| Aggrecan | F: TGCAAGGCAAAGTCTTCTACG |
| R: GGCAGGGTTCAGGTAAACG | |
| Col2a1 | F: ACCTACAGCGTCTTGGAGGA |
| R: ATATCCACGCCAAACTCCTG | |
| BMP2 | F: CTCAGCTCAGGCCGTTGTTA |
| R: GTCATTCCACCCCACGTCAT | |
| Runx2 | F: TTCACAAGCATTTCATCCCTC |
| R: TTGCGGACATACCCAGTGACA | |
| MMP9 | F: GCCATCACTGAGATCAATGGAG |
| R: GATAGAGAAGGCGCCCTGAGT | |
| MMP13 | F: AGAGACCCTGGAGCACTGATGT |
| R: GGGATCTCTGTCTCCAGCACCA | |
| Notch1 | F: AGAGGAACTCAAGGGGAGCG |
| R: CGGCACTCGCATTTGTAGTC | |
| Notch2 | F: CACACCCGAGTGCTTGTTTG |
| R: ATTACAGCCCTGGTCACAGC | |
| JAG1 | F: AACTGGTACCGCTGTGAGTG |
| R: GCAAGGCCTCCCTGTAACTT | |
| JAG2 | F: CAAGTGGCTGGGAAGGAGAA |
| R: TGCATCGGCCACCATTATGA | |
| DLL1 | F: TGAACTACTGCACTCACCACAA |
| R: TCGTTGATTTCAATCTCGCAGC | |
| DLL4 | F: CAGCAGGTAACGGTCGGAG |
| R: TTGACGAACTCGTGCAGCTT | |
| RBPJ | F: GCAGATGATCCGGTATCGCAG |
| R: TTTGGGCATGGAGTGGCTTGA | |
| HES1 | F: CAGCGAGTGCATGAACGAAG |
| R: TGATGGCGTTGATCTGGGTC | |
| HES4 | F: CCCATCATGGAGAAGCGACG |
| R: GAGTGCCGAGAGCTGTCTTTT | |
| HES5 | F: TGAAATACAGCCGAGCTTTTGC |
| R: GCAGAAGGAGAACCGGAGTC | |
| HES6 | F: CGCATTCCACTTGGATCAGTCTA |
| R: AGGCCCACTTTGGAATCAGC | |
| HEY1 | F: GGCCGGAGGGAAAGGTTATTT |
| R: GTGATGTCCAAAGGCGTTGC | |
| HEY2 | F: TATTTCTCTTTGCCCCACGCC |
| R: TATGGCTTTGCCCGCAGTA | |
| HEYL | F: TCAGGATGAAGCGTCTGTGC R: GCCGCTTCTCAATGATCCCT |
| Caspase-3 | F: TGCTCCAGGCTACTACTCC R: CCACTCTGCGATTTACACGA |
| Caspase-9 | F: CGAAGGAGCAAGCACGAC R: CGCAGCCCTCATCTAGCAT |
| Bcl-2 | F: CGACTGGGATGACAGGAAAG R: GGAGCGCACAGGTGAGACA |
| GAPDH | F: GAACATCATCCCAGCGTCCA |
| R: CGGCAGGTCAGGTCAACAAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Ge, H.; Liu, C.; Jiang, Y.; Yu, Y.; Zhou, Z. Notch-RBPJ Pathway for the Differentiation of Bone Marrow Mesenchymal Stem Cells in Femoral Head Necrosis. Int. J. Mol. Sci. 2023, 24, 6295. https://doi.org/10.3390/ijms24076295
Liu K, Ge H, Liu C, Jiang Y, Yu Y, Zhou Z. Notch-RBPJ Pathway for the Differentiation of Bone Marrow Mesenchymal Stem Cells in Femoral Head Necrosis. International Journal of Molecular Sciences. 2023; 24(7):6295. https://doi.org/10.3390/ijms24076295
Chicago/Turabian StyleLiu, Kangping, Hongfan Ge, Chengyin Liu, Yixin Jiang, Yaling Yu, and Zhenlei Zhou. 2023. "Notch-RBPJ Pathway for the Differentiation of Bone Marrow Mesenchymal Stem Cells in Femoral Head Necrosis" International Journal of Molecular Sciences 24, no. 7: 6295. https://doi.org/10.3390/ijms24076295
APA StyleLiu, K., Ge, H., Liu, C., Jiang, Y., Yu, Y., & Zhou, Z. (2023). Notch-RBPJ Pathway for the Differentiation of Bone Marrow Mesenchymal Stem Cells in Femoral Head Necrosis. International Journal of Molecular Sciences, 24(7), 6295. https://doi.org/10.3390/ijms24076295