

Fatty Acid Induced Hypermethylation in the Slc2a4 Gene in Visceral Adipose Tissue Is Associated to Insulin-Resistance and Obesity

 , , ,
, , ,
Abstract
:1. Introduction
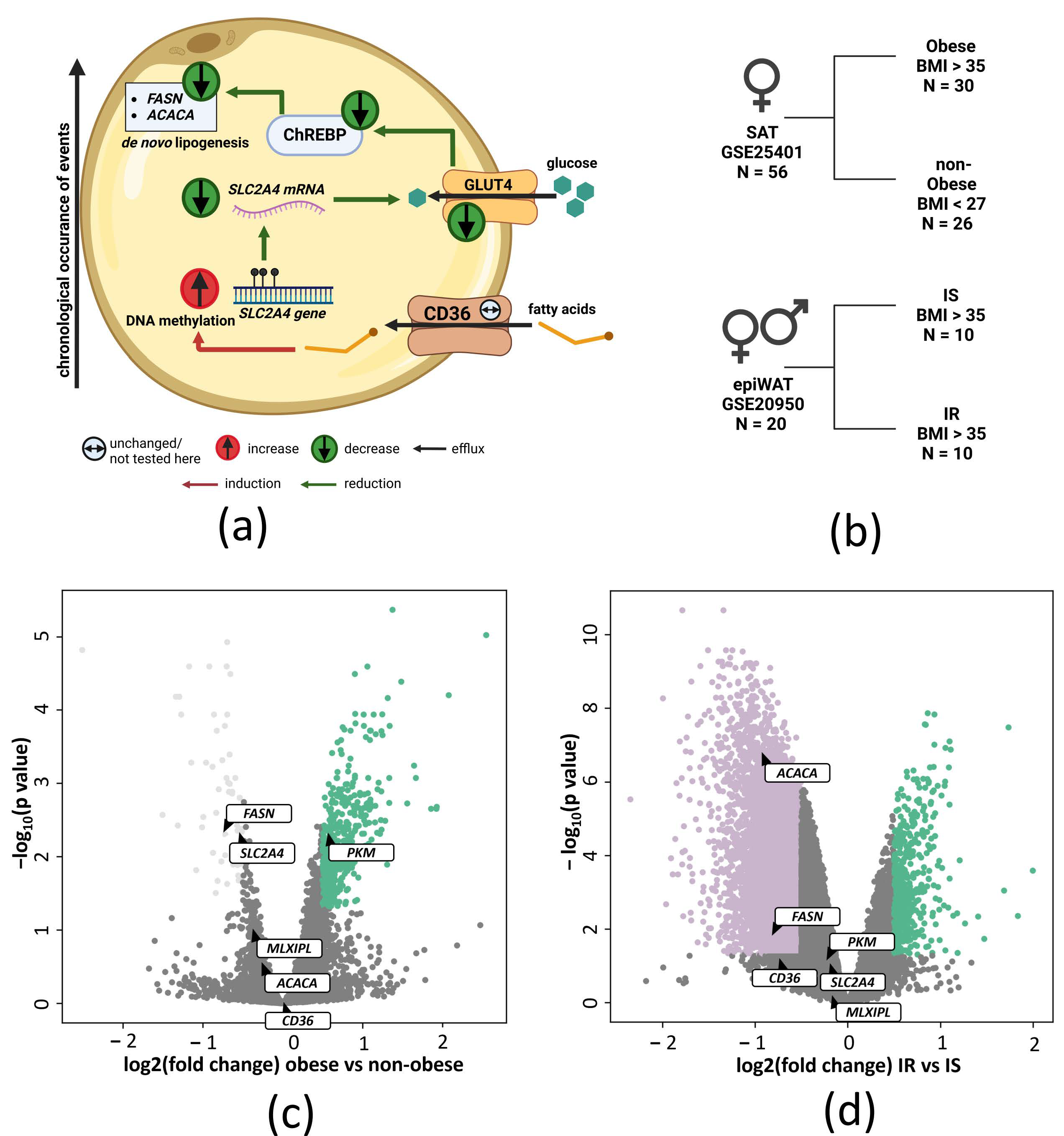
2. Results
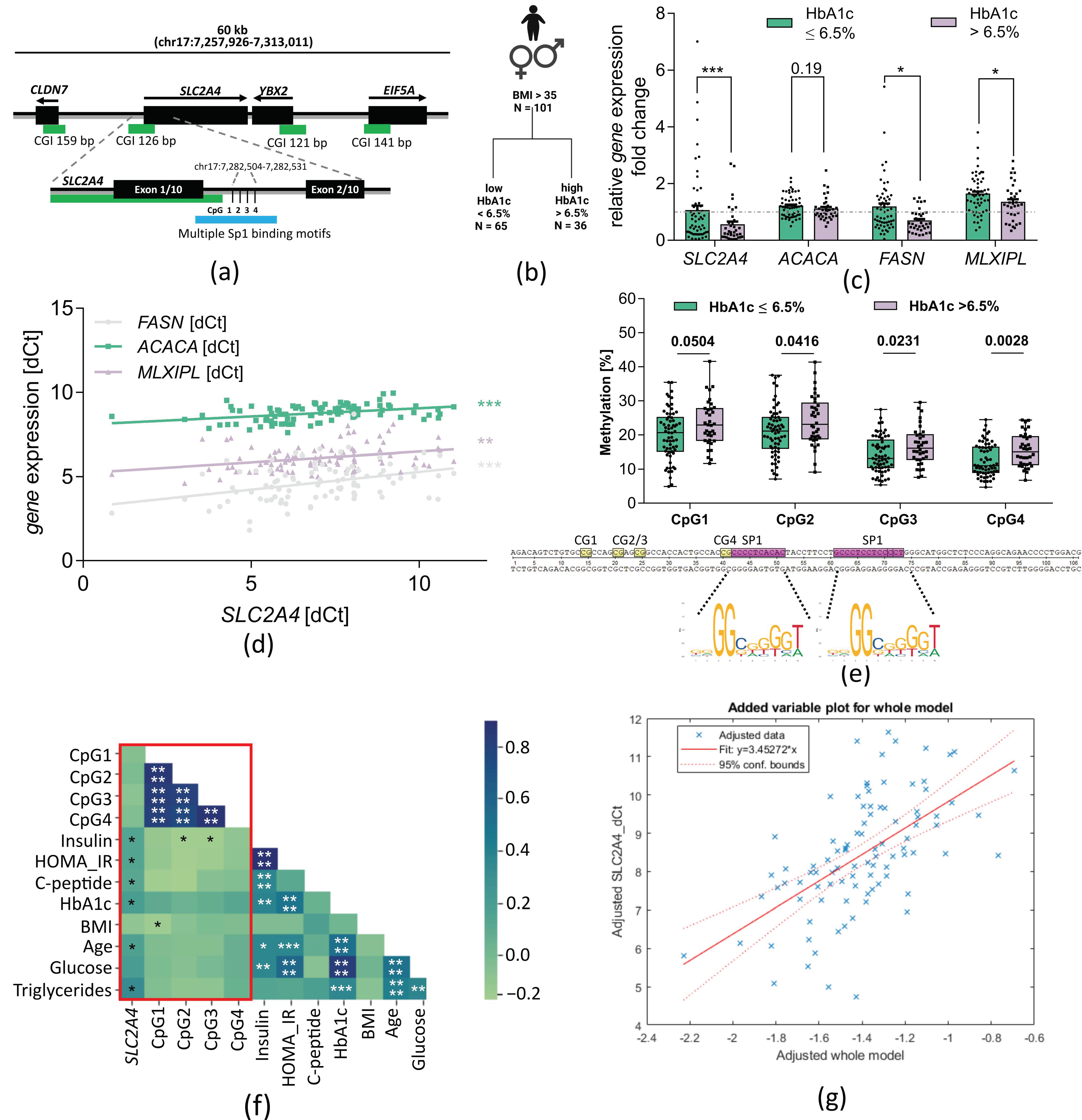
2.1. Enhancer-Associated SLC2A4 DNA Methylation Is Increased in Visceral Adipose Tissue of Morbidly Obese Subjects with High HbA1c and Is Associated with Decreased Gene Expression
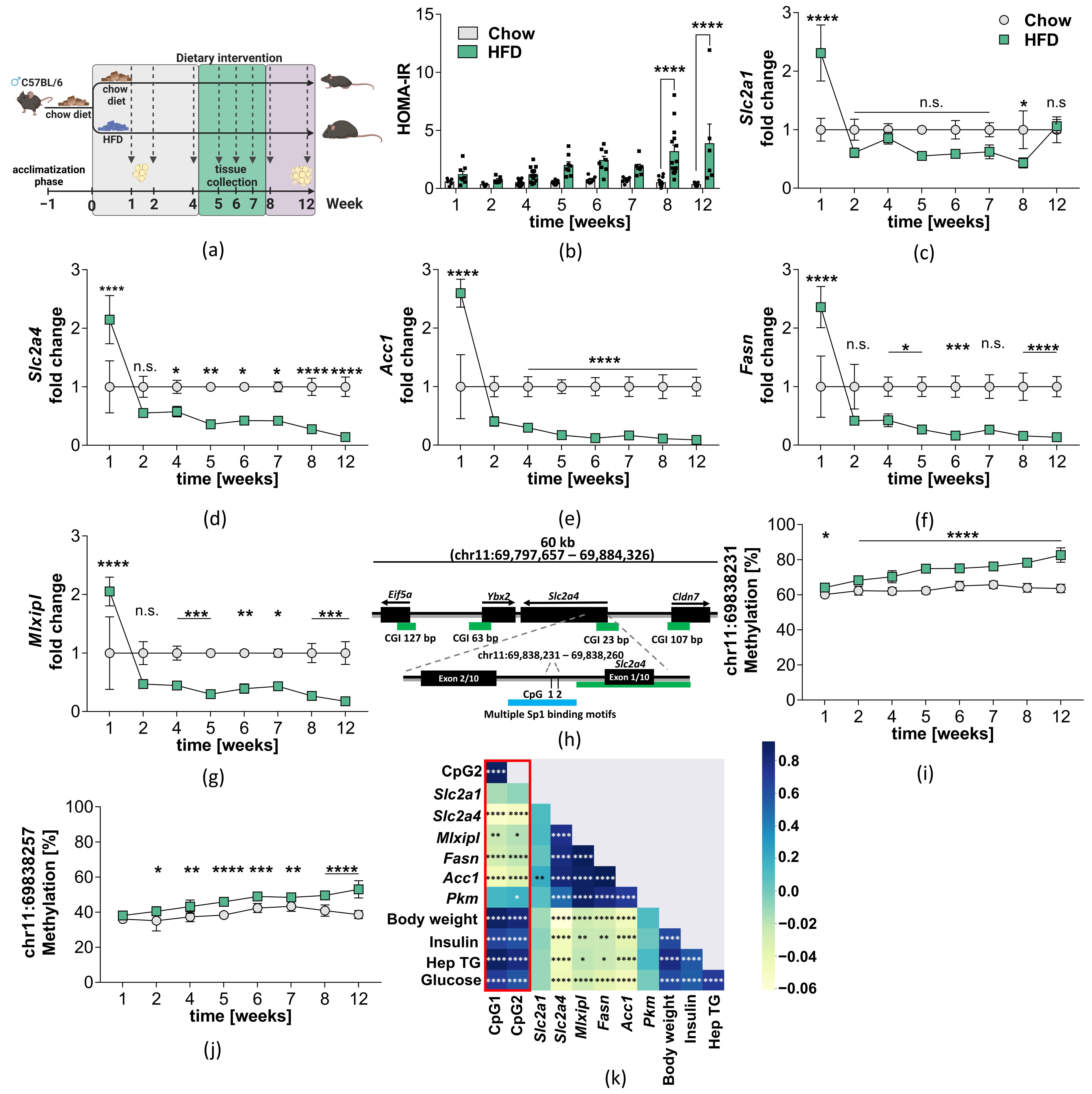
2.2. Hypermethylation of the Slc2a4 Gene in VAT of Diet-Induced Obese Mice
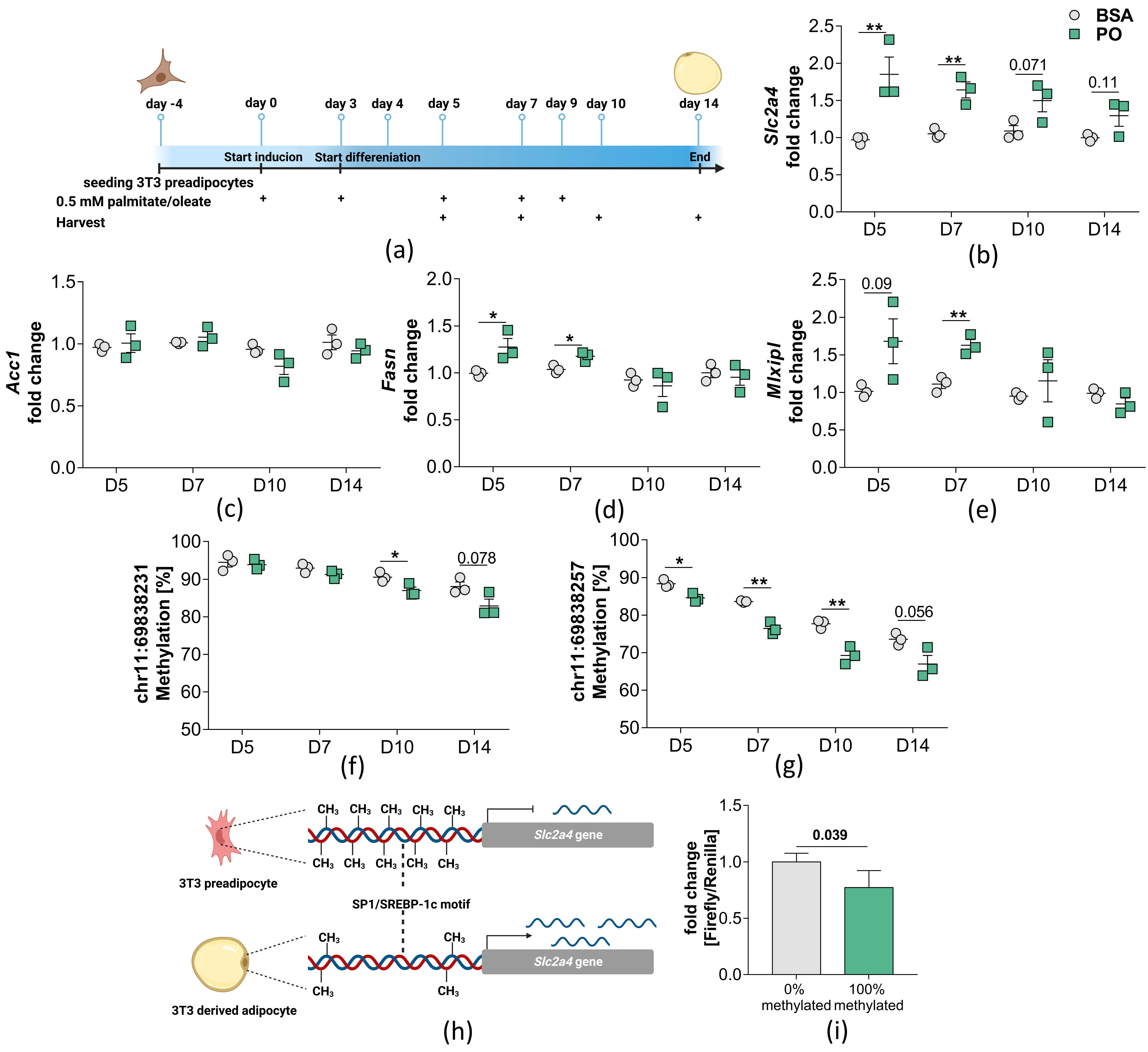
2.3. Palmitate/Oleate Treatment of Differentiating Preadipocytes Alters DNA Methylation in the Slc2a4 Gene
3. Discussion
4. Materials and Methods
4.1. Analysis of Publicly Available Transcriptomic Data
4.2. Study Subjects Used for qPCR and Pyrosequencing Analysis
4.3. Mouse Model of Diet-Induced Obesity
4.4. RNA Extraction, cDNA Synthesis and RT-qPCR
4.5. DNA Extraction from VAT, Bisulfite Conversion, and Pyrosequencing
4.6. Western Blot
4.7. Methylation-Sensitive Reporter Gene Assay
4.8. Treatment of 3T3 Preadipocytes with Palmitate/Oleate Mixture
4.9. Statistical Analysis
4.10. Stepwise Regression Model
(OPBMI × SLC2A4_CpG2)β3 + (OPBMI × SLC2A4_CpG4)β4 + (Age × sex_2_w_1_m)β5 +
(Age × SLC2A4_CpG1)β6 + (SLC2A4_CpG1 × SLC2A4_CpG2)β7 +
(SLC2A4_CpG2 × SLC2A4_CpG3)β8 + (SLC2A4_CpG3 × SLC2A4_CpG4)β9
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kalra, S.; Choudhary, N.; Unnikrishnan, A.G.; Ajish, T. Preventive pharmacotherapy in type 2 diabetes mellitus. Indian J. Endocrinol. Metab. 2012, 16, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M.; Newgard, C.B. Mechanisms of disease:Molecular and metabolic mechanisms of insulin resistance and β-cell failure in type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Aruleba, R.T.; Adekiya, T.A.; Oyinloye, B.E.; Kappo, A.P. Structural Studies of Predicted Ligand Binding Sites and Molecular Docking Analysis of Slc2a4 as a Therapeutic Target for the Treatment of Cancer. Int. J. Mol. Sci. 2018, 19, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrêa-Giannella, M.L.; Machado, U.F. SLC2A4 gene: A promising target for pharmacogenomics of insulin resistance. Pharmacogenomics 2013, 14, 847–850. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Czech, M.P. The GLUT4 Glucose Transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Machado, U.F.; Shimizu, I.; Saito, M. Reduced content and preserved translocation of glucose transporter (GLUT 4) in white adipose tissue of obese mice. Physiol. Behav. 1994, 55, 621–625. [Google Scholar] [CrossRef]
- Song, Z.; Xiaoli, A.M.; Yang, F. Regulation and Metabolic Significance of De Novo Lipogenesis in Adipose Tissues. Nutrients 2018, 10, 1383. [Google Scholar] [CrossRef] [Green Version]
- Eissing, L.; Scherer, T.; Tödter, K.; Knippschild, U.; Greve, J.W.; Buurman, W.A.; Pinnschmidt, H.O.; Rensen, S.S.; Wolf, A.M.; Bartelt, A.; et al. De novo lipogenesis in human fat and liver is linked to ChREBP-β and metabolic health. Nat. Commun. 2013, 4, 1528. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Gerhold, K.; Mayers, J.R.; Wiest, M.M.; Watkins, S.M.; Hotamisligil, G.S. Identification of a Lipokine, a Lipid Hormone Linking Adipose Tissue to Systemic Metabolism. Cell 2008, 134, 933–944. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, E.; Kotani, K.; Peroni, O.D.; Kahn, B.B. Adipose-specific overexpression of GLUT4 reverses insulin resistance and diabetes in mice lacking GLUT4 selectively in muscle. Am. J. Physiol. Metab. 2005, 289, E551–E561. [Google Scholar] [CrossRef] [Green Version]
- Barreto-Andrade, J.N.; de Fátima, L.A.; Campello, R.S.; Guedes, J.A.C.; de Freitas, H.S.; Machado, M.M.F. Estrogen Receptor 1 (ESR1) Enhances Slc2a4/GLUT4 Expression by a SP1 Cooperative Mechanism. Int. J. Med. Sci. 2018, 15, 1320–1328. [Google Scholar] [CrossRef] [Green Version]
- Im, S.-S.; Kwon, S.-K.; Kang, S.-Y.; Kim, T.-H.; Kim, H.-I.; Hur, M.-W.; Kim, K.-S.; Ahn, Y.-H. Regulation of GLUT4 gene expression by SREBP-1c in adipocytes. Biochem. J. 2006, 399, 131–139. [Google Scholar] [CrossRef]
- Herrera, B.M.; Keildson, S.; Lindgren, C.M. Genetics and epigenetics of obesity. Maturitas 2011, 69, 41–49. [Google Scholar] [CrossRef]
- Ardekani, A.M. Role of Epigenetics in Biology and Human Diseases. Iran. Biomed. J. 2016, 20, 246–258. [Google Scholar] [CrossRef]
- Cierzniak, A.; Pawelka, D.; Kaliszewski, K.; Rudnicki, J.; Dobosz, T.; Malodobra-Mazur, M. DNA methylation in adipocytes from visceral and subcutaneous adipose tissue influences insulin-signaling gene expression in obese individuals. Int. J. Obes. 2021, 45, 650–658. [Google Scholar] [CrossRef]
- Esteves, J.V.; Yonamine, C.Y.; Machado, U.F. SLC2A4 expression and its epigenetic regulation as biomarkers for insulin resistance treatment in diabetes mellitus. Biomarkers Med. 2020, 14, 413–416. [Google Scholar] [CrossRef]
- Hardy, O.T.; Perugini, R.A.; Nicoloro, S.M.; Gallagher-Dorval, K.; Puri, V.; Straubhaar, J.; Czech, M.P. Body mass index-independent inflammation in omental adipose tissue associated with insulin resistance in morbid obesity. Surg. Obes. Relat. Dis. Off. J. Am. Soc. Bariatr. Surg. 2011, 7, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Arner, E.; Mejhert, N.; Kulyté, A.; Balwierz, P.J.; Pachkov, M.; Cormont, M.; Lorente-Cebrián, S.; Ehrlund, A.; Laurencikiene, J.; Hedén, P.; et al. Adipose Tissue MicroRNAs as Regulators of CCL2 Production in Human Obesity. Diabetes 2012, 61, 1986–1993. [Google Scholar] [CrossRef] [Green Version]
- Douet, V.; Heller, M.B.; Le Saux, O. DNA methylation and Sp1 binding determine the tissue-specific transcriptional activity of the mouse Abcc6 promoter. Biochem. Biophys. Res. Commun. 2007, 354, 66–71. [Google Scholar] [CrossRef] [Green Version]
- The ENCODE Project Consortium. An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [Green Version]
- Sievert, H.; Krause, C.; Geißler, C.; Grohs, M.; El-Gammal, A.T.; Wolter, S.; Mann, O.; Lehnert, H.; Kirchner, H. Epigenetic Downregulation of FASN in Visceral Adipose Tissue of Insulin Resistant Subjects. Exp. Clin. Endocrinol. Diabetes 2021, 129, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Katsogiannos, P.; Kamble, P.G.; Boersma, G.J.; Karlsson, F.A.; Lundkvist, P.; Sundbom, M.; Pereira, M.J.; Eriksson, J.W. Early Changes in Adipose Tissue Morphology, Gene Expression, and Metabolism After RYGB in Patients with Obesity and T2D. J. Clin. Endocrinol. Metab. 2019, 104, 2601–2613. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.C.D.; Zhou, T.; Rao, S.; Goel, P.; Rastogi, C.; Lazarovici, A.; Bussemaker, H.J.; Rohs, R. Evolving insights on how cytosine methylation affects protein-DNA binding. Briefings Funct. Genom. 2014, 14, 61–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuyàs, E.; Fernández-Arroyo, S.; Verdura, S.; García, R.Á.-F.; Stursa, J.; Werner, L.; Blanco-González, E.; Montes-Bayón, M.; Joven, J.; Viollet, B.; et al. Metformin regulates global DNA methylation via mitochondrial one-carbon metabolism. Oncogene 2018, 37, 963–970. [Google Scholar] [CrossRef]
- Moraes, P.A.; Yonamine, C.Y.; Junior, D.C.P.; Esteves, J.V.D.; Machado, U.F.; Mori, R.C. Insulin acutely triggers transcription of Slc2a4 gene: Participation of the AT-rich, E-box and NFKB-binding sites. Life Sci. 2014, 114, 36–44. [Google Scholar] [CrossRef]
- Bereal-Williams, C.; Machado, R.F.; McGowan, V.; Chi, A.; Hunter, C.J.; Kato, G.J. Atorvastatin reduces serum cholesterol and triglycerides with limited improvement in vascular function in adults with sickle cell anemia. Haematologica 2012, 97, 1768–1770. [Google Scholar] [CrossRef] [Green Version]
- Poletto, A.C.; David-Silva, A.; Yamamoto, A.P.D.M.; Machado, U.F.; Furuya, D.T. Reduced Slc2a4/GLUT4 expression in subcutaneous adipose tissue of monosodium glutamate obese mice is recovered after atorvastatin treatment. Diabetol. Metab. Syndr. 2015, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride Metabolism in the Liver. Compr. Physiol. 2018, 8, 1–22. [Google Scholar] [CrossRef]
- Ley, S.H.; Hamdy, O.; Mohan, V.; Hu, F.B. Prevention and management of type 2 diabetes: Dietary components and nutritional strategies. Lancet 2014, 383, 1999–2007. [Google Scholar] [CrossRef] [Green Version]
- Fraulob, J.C.; Ogg-Diamantino, R.; Fernandes-Santos, C.; Aguila, M.B.; Mandarim-De-Lacerda, C.A. A Mouse Model of Metabolic Syndrome: Insulin Resistance, Fatty Liver and Non-Alcoholic Fatty Pancreas Disease (NAFPD) in C57BL/6 Mice Fed a High Fat Diet. J. Clin. Biochem. Nutr. 2010, 46, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Geißler, C.; Krause, C.; Neumann, A.-M.; Britsemmer, J.H.; Taege, N.; Grohs, M.; Kaehler, M.; Cascorbi, I.; Lewis, A.G.; Seeley, R.J.; et al. Dietary induction of obesity and insulin resistance is associated with changes in Fgf21 DNA methylation in liver of mice. J. Nutr. Biochem. 2022, 100, 108907. [Google Scholar] [CrossRef]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0 Contributors. SciPy 1.0 Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Obese (n = 30) | Non-Obese (n = 26) | p Value |
|---|---|---|---|
| Age [years] | 43 ± 2 | 43 ± 3 | 0.8 |
| BMI 1 [kg/m2] | 40.9 ± 1.3 | 24 ± 1.8 | <0.0001 |
| Log10 HOMAIR 2 | 0.458 ± 0.06 | −0.01 ± 0.049 | <0.0001 |
| Characteristic | Insulin Sensitive (HOMA < 2.4; n = 10) | Insulin Resistant (HOMA > 2.4; n = 10) | p Value |
|---|---|---|---|
| Age [years] | 39 ± 6 | 43 ± 9 | 0.2 (n.s.) |
| Woman [number] | 8 | 6 | 0.4 (n.s.) |
| BMI 1 | 48 ± 3 | 49 ± 7 | 0.7 (n.s.) |
| Total cholesterol [mg/dL] | 176 ± 27 | 179 ± 50 | 0.9 (n.s.) |
| LDL cholesterol 2 [mg/dL] | 109 ± 15 | 101 ± 45 | 0.6 (n.s.) |
| HDL cholesterol 3 [mg/dL] | 44 ± 5 | 42 ± 9 | 0.5 (n.s.) |
| Triglycerides [mg/dL] | 129 ± 23 | 166 ± 74 | 0.2 (n.s.) |
| Fasting glucose [mg/dL] | 88 ± 9 | 101 ± 12 | 0.02 |
| Fasting insulin [mIU/mL] | 9 ± 4 | 24 ± 7 | <0.001 |
| HOMA 4 | 1.3 ± 0.5 | 3.6 ± 5.3 | <0.001 |
| Characteristic | Low-HbA1c (n = 65) | High-HbA1c (n = 36) | p Value |
|---|---|---|---|
| Age [years] | 39 ± 12 | 53 ± 10 | <0.0001 |
| Woman [number] | 52 | 21 | 0.01 |
| BMI 1 | 52.51 ± 11.18 | 50.54 ± 9.07 | 0.36 (n.s.) |
| Total cholesterol [mg/dL] | 187.02 ± 32.32 | 188.31 ± 48.50 | 0.88 (n.s.) |
| LDL cholesterol 2 [mg/dL] | 106.36 ± 30.12 | 90.67 ± 35.48 | 0.03 |
| HDL cholesterol 3 [mg/dL] | 47.02 ± 11.66 | 41.19 ± 13.45 | 0.03 |
| Triglycerides [mg/dL] | 170.69 ± 89.89 | 280.81 ± 155.17 | <0.0001 |
| Fasting glucose [mg/dL] | 94.58 ± 15.99 | 179.81 ± 71.67 | <0.0001 |
| Fasting insulin [mIU/mL] | 13.25 ± 11.53 | 12.61 ± 29.75 | 0.003 |
| HOMA 4 | 2.9 ± 2.82 | 12.61 ± 18.82 | 0.0003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Britsemmer, J.H.; Krause, C.; Taege, N.; Geißler, C.; Lopez-Alcantara, N.; Schmidtke, L.; Naujack, A.-M.; Wagner, J.; Wolter, S.; Mann, O.; et al. Fatty Acid Induced Hypermethylation in the Slc2a4 Gene in Visceral Adipose Tissue Is Associated to Insulin-Resistance and Obesity. Int. J. Mol. Sci. 2023, 24, 6417. https://doi.org/10.3390/ijms24076417
Britsemmer JH, Krause C, Taege N, Geißler C, Lopez-Alcantara N, Schmidtke L, Naujack A-M, Wagner J, Wolter S, Mann O, et al. Fatty Acid Induced Hypermethylation in the Slc2a4 Gene in Visceral Adipose Tissue Is Associated to Insulin-Resistance and Obesity. International Journal of Molecular Sciences. 2023; 24(7):6417. https://doi.org/10.3390/ijms24076417
Chicago/Turabian StyleBritsemmer, Jan H., Christin Krause, Natalie Taege, Cathleen Geißler, Nuria Lopez-Alcantara, Linda Schmidtke, Alison-Michelle Naujack, Jonas Wagner, Stefan Wolter, Oliver Mann, and et al. 2023. "Fatty Acid Induced Hypermethylation in the Slc2a4 Gene in Visceral Adipose Tissue Is Associated to Insulin-Resistance and Obesity" International Journal of Molecular Sciences 24, no. 7: 6417. https://doi.org/10.3390/ijms24076417
APA StyleBritsemmer, J. H., Krause, C., Taege, N., Geißler, C., Lopez-Alcantara, N., Schmidtke, L., Naujack, A. -M., Wagner, J., Wolter, S., Mann, O., & Kirchner, H. (2023). Fatty Acid Induced Hypermethylation in the Slc2a4 Gene in Visceral Adipose Tissue Is Associated to Insulin-Resistance and Obesity. International Journal of Molecular Sciences, 24(7), 6417. https://doi.org/10.3390/ijms24076417