
Bilateral Shifts in Deuterium Supply Similarly Change Physiology of the Pituitary–Thyroid Axis, but Differentially Influence Na+/I− Symporter Production

 ,
,  , and
, and 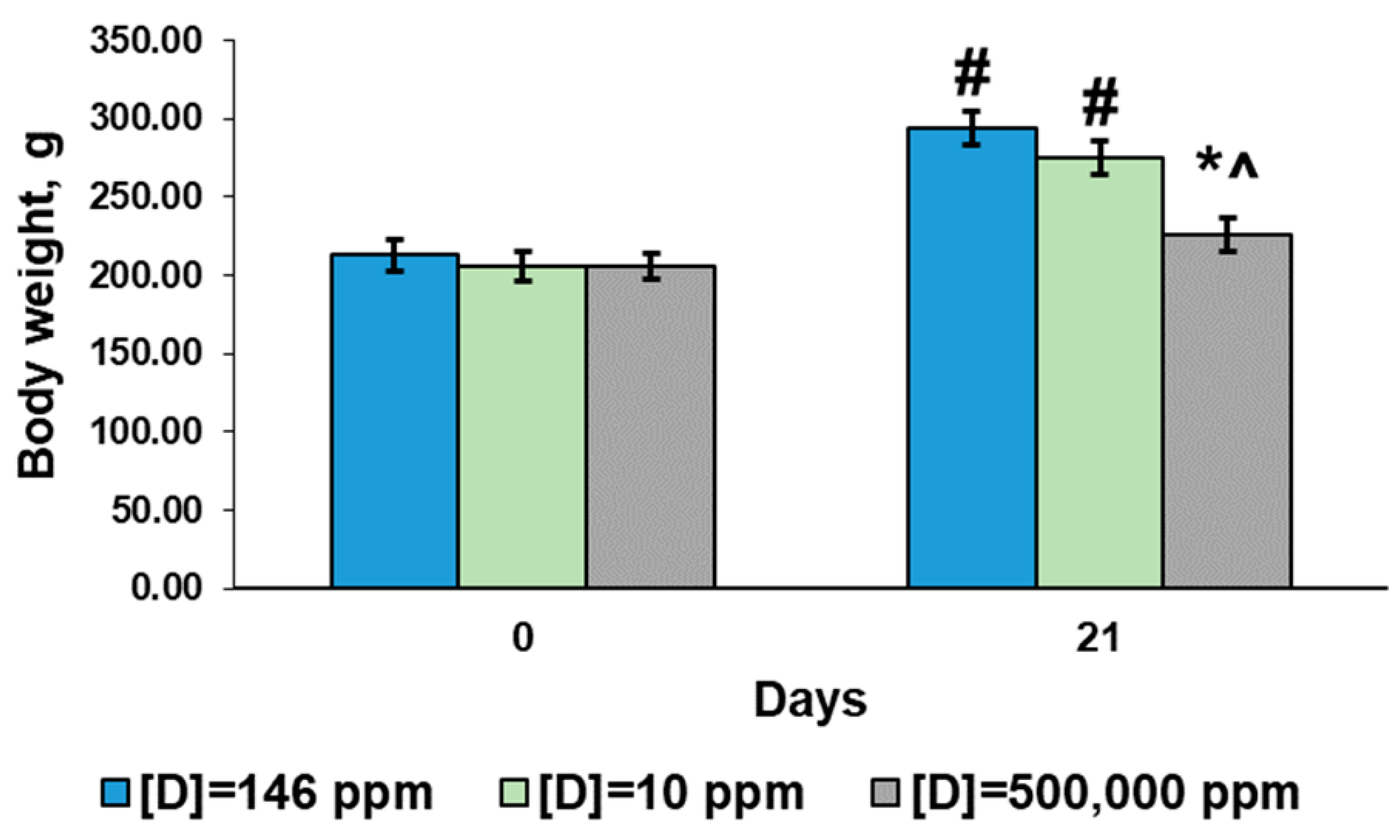{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Body Weight Parameters
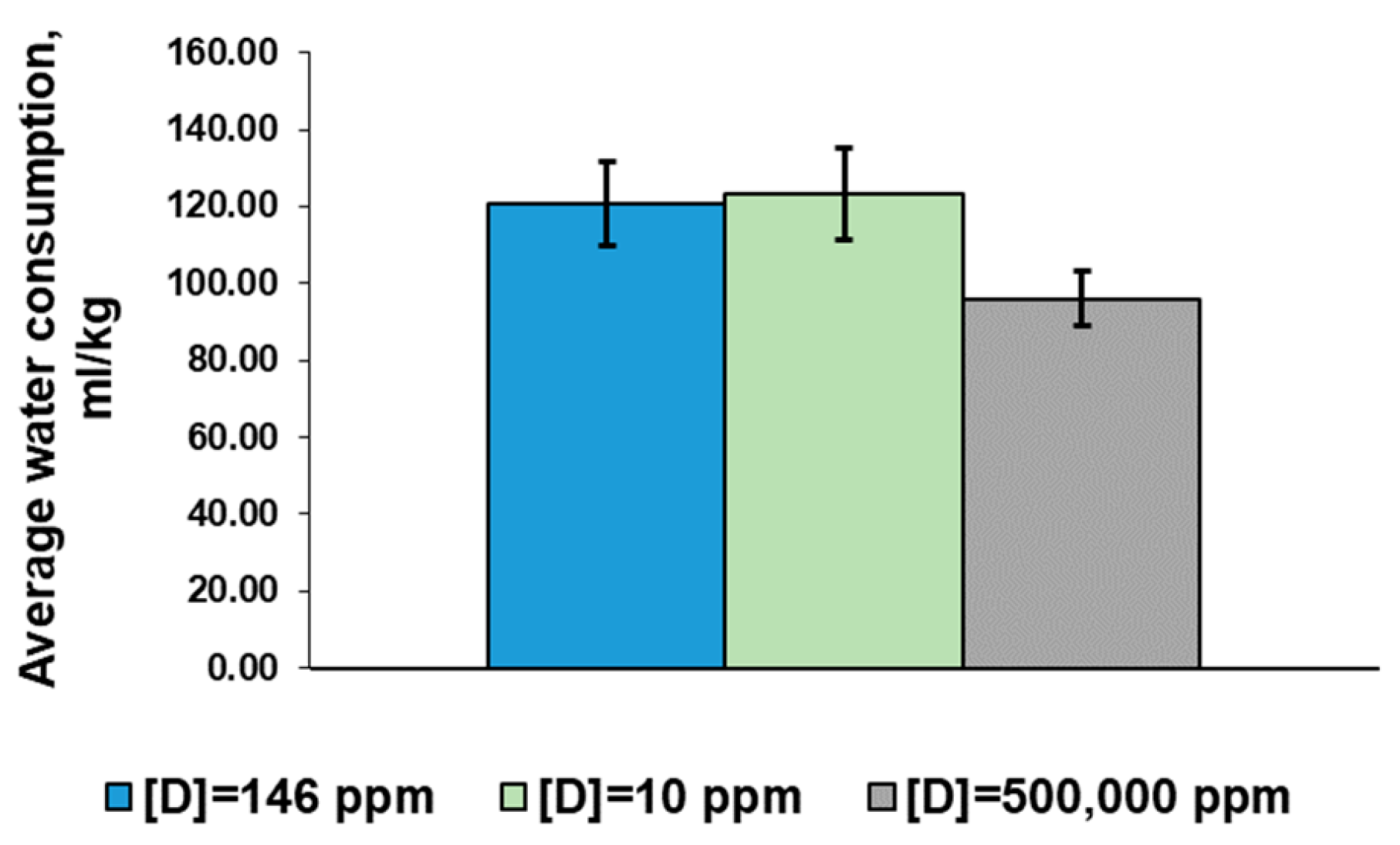
2.2. Water Consumption Parameters
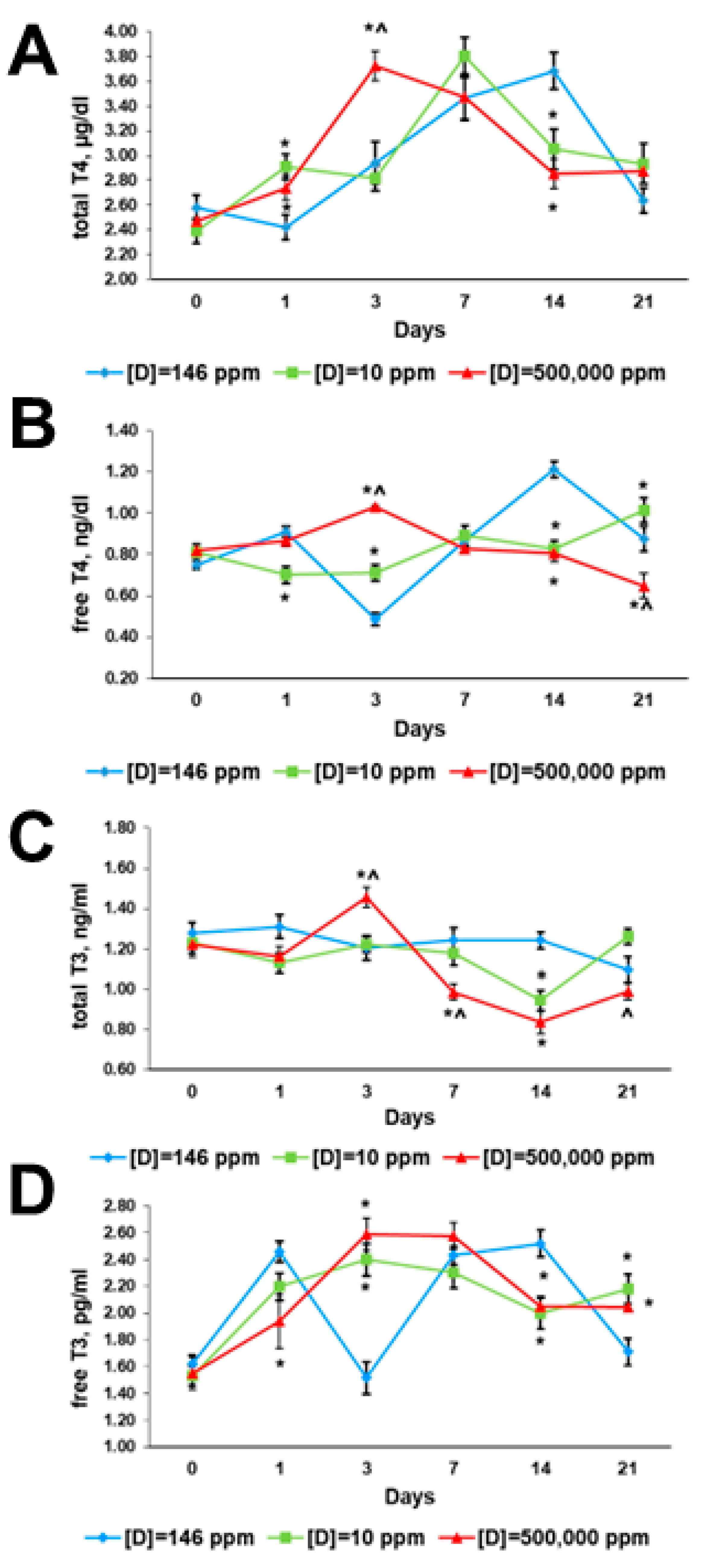
2.3. Thyroid Hormone Profiles
2.3.1. Before the Exposure
2.3.2. First Day of Exposure
2.3.3. Third Day of Exposure
2.3.4. Seventh Day of Exposure
2.3.5. 14th Day of Exposure
2.3.6. 21st Day of Exposure
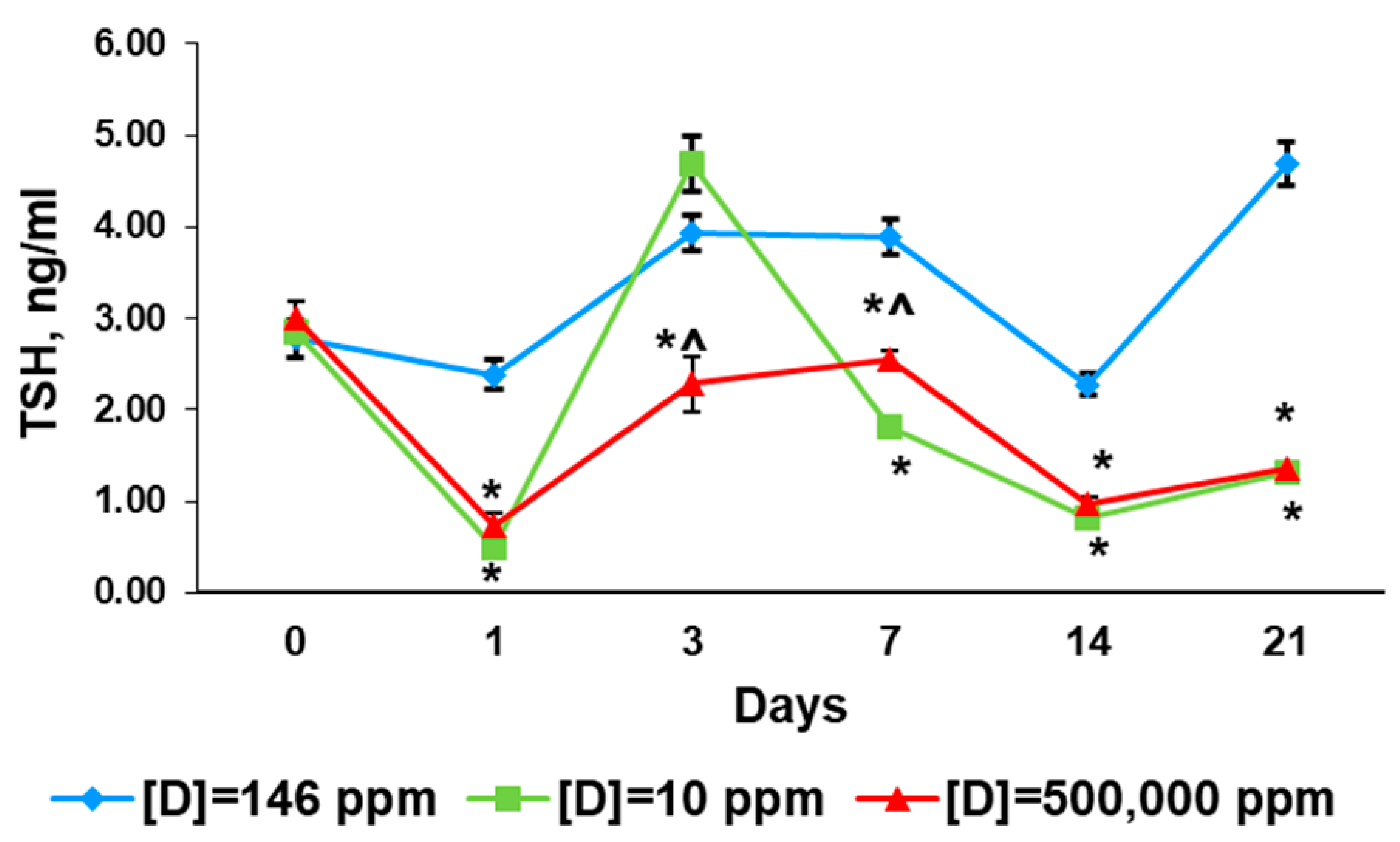
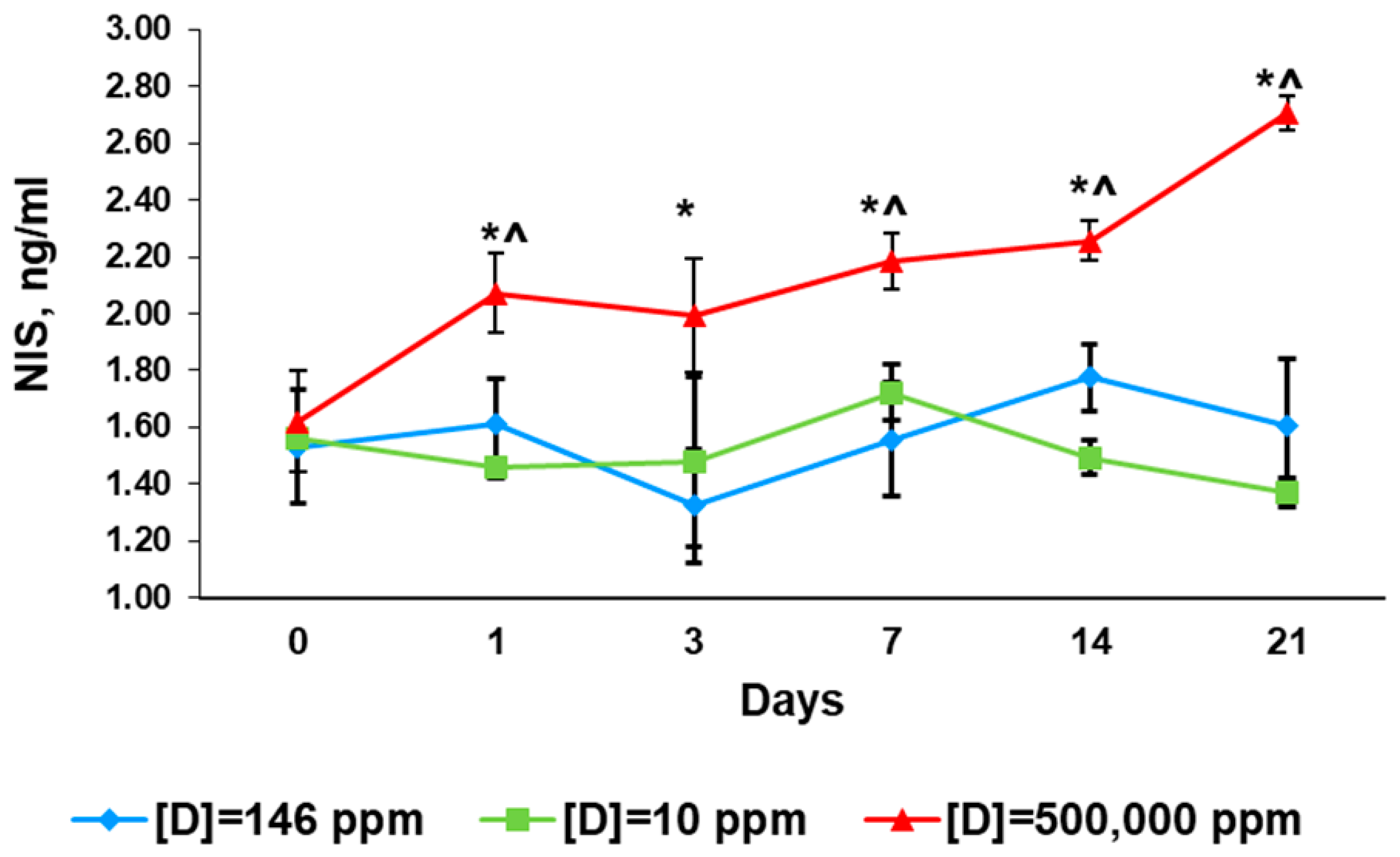
2.4. Serum Levels of Sodium/Iodide Symporter
2.4.1. Before the Exposure
2.4.2. First Day of Exposure
2.4.3. Third Day of Exposure
2.4.4. Seventh Day of Exposure
2.4.5. 14th Day of Exposure
2.4.6. 21st Day of Exposure
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Reagents
4.3. Ethical Approval
4.4. Thyroid Profile Assessment
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Podlesak, D.W.; Torregrossa, A.-M.; Ehleringer, J.R.; Dearing, M.D.; Passey, B.H.; Cerling, T.E. Turnover of oxygen and hydrogen isotopes in the body water, CO2, hair, and enamel of a small mammal. Geochim. Cosmochim. Acta 2008, 72, 19–35. [Google Scholar] [CrossRef]
- Shchepinov, M.S. Do “heavy’ eaters live longer? Bioessays 2007, 29, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Somlyai, G.; Javaheri, B.; Davari, H.; Gyöngyi, Z.; Somlyai, I.; Tamaddon, K.A.; Boros, L.G. Pre-Clinical and Clinical Data Confirm the Anticancer Effect of Deuterium Depletion. Biomacromol. J. 2016, 2, 1–7. [Google Scholar] [CrossRef]
- Boros, L.G.; Somlyai, I.; Kovács, B.Z.; Puskás, L.G.; Nagy, L.I.; Dux, L.; Farkas, G.; Somlyai, G. Deuterium Depletion Inhibits Cell Proliferation, RNA and Nuclear Membrane Turnover to Enhance Survival in Pancreatic Cancer. Cancer Control 2021, 28, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, J.; Zubarev, R.A. Slight Deuterium Enrichment in Water Acts as an Antioxidant: Is Deuterium a Cell Growth Regulator? Mol. Cell. Proteom. 2020, 19, 1790–1804. [Google Scholar] [CrossRef]
- Cong, F.S.; Zhang, Y.R.; Sheng, H.C.; Ao, Z.H.; Zhang, S.Y.; Wang, J.Y. Deuterium-depleted water inhibits human lung carcinoma cell growth by apoptosis. Exp. Ther. Med. 2010, 1, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Kravtsov, A.; Kozin, S.; Basov, A.; Butina, E.; Baryshev, M.; Malyshko, V.; Moiseev, A.; Elkina, A.; Dzhimak, S. Reduction of Deuterium Level Supports Resistance of Neurons to Glucose Deprivation and Hypoxia: Study in Cultures of Neurons and on Animals. Molecules 2022, 27, 243. [Google Scholar] [CrossRef]
- Yaglova, N.V.; Timokhina, E.P.; Obernikhin, S.S.; Yaglov, V.V. Emerging Role of Deuterium/Protium Disbalance in Cell Cycle and Apoptosis. Int. J. Mol. Sci. 2023, 24, 3107. [Google Scholar] [CrossRef]
- Dzhimak, S.S.; Basov, A.A.; Baryshev, M.G. Content of deuterium in biological fluids and organs: Influence of deuterium depleted water on D/H gradient and the process of adaptation. Dokl. Biochem. Biophys. 2015, 465, 370–373. [Google Scholar] [CrossRef]
- Yaglova, N.V.; Obernikhin, S.S.; Yaglov, V.V.; Nazimova, S.V. Time-dependent effect of deuterium depletion on tumor growth and metastasis. Russ. Open Med. J. 2020, 9, e0210. [Google Scholar] [CrossRef]
- Jancso, G. Isotope effects. In Handbook of Nuclear Chemistry; Vértes, A., Nagy, S., Klencsar, Z., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherland, 2013; Volume 2, pp. 85–116. [Google Scholar]
- Kumar, K.V.S.H.; Patnaik, S.K. Incidence of Endocrine Disorders in Indian Adult Male Population. Indian J. Endocrinol. Metab. 2017, 21, 809–811. [Google Scholar] [CrossRef] [PubMed]
- Crafa, A.; Calogero, A.E.; Cannarella, R.; Mongioi, L.M.; Condorelli, R.A.; Greco, E.A.; Aversa, A.; La Vignera, S. The Burden of Hormonal Disorders: A Worldwide Overview with a Particular Look in Italy. Front. Endocrinol. 2021, 12, 694325. [Google Scholar] [CrossRef] [PubMed]
- Yaglova, N.V.; Sledneva, Y.P.; Nazimova, S.V.; Obernikhin, S.S.; Yaglov, V.V. Sex Differences in the Production of SLC5A5, Thyroid Peroxidase, and Thyroid Hormones in Pubertal Rats Exposed to Endocrine Disruptor Dichlorodiphenyltrichloroethane (DDT) during Postnatal Ontogeny. Bull. Exp. Biol. Med. 2018, 164, 430–433. [Google Scholar] [CrossRef]
- Sun, M.; Cao, X.; Wu, Y.; Shen, L.; Wei, G. Prenatal exposure to endocrine-disrupting chemicals and thyroid function in neonates: A systematic review and meta-analysis. Ecotoxicol. Environ. Saf. 2022, 231, 113215. [Google Scholar] [CrossRef]
- Reiners, C.; Drozd, V.; Yamashita, S. Hypothyroidism after radiation exposure: Brief narrative review. J. Neural. Transm. 2020, 127, 1455–1466. [Google Scholar] [CrossRef]
- Zhai, R.; Lyu, Y.; Ni, M.; Kong, F.; Du, C.; Hu, C.; Ying, H. Predictors of radiation-induced hypothyroidism in nasopharyngeal carcinoma survivors after intensity-modulated radiotherapy. Radiat. Oncol. 2022, 17, 57. [Google Scholar] [CrossRef]
- Hatch-McChesney, A.; Lieberman, H.R. Iodine and Iodine Deficiency: A Comprehensive Review of a Re-Emerging Issue. Nutrients 2022, 14, 3474. [Google Scholar] [CrossRef]
- Mosin, O.; Shvets, V.; Skladnev, D.; Ignatov, I. Studying of microbic synthesis of deuteriumlabelled l-phenylalanine by methylotrophic bacterium brevibacterium methylicum on media with differeny content of heavy water. Russ. J. Biopharmaceut. 2012, 4, 11–22. [Google Scholar]
- Lobyshev, V.I.; Tverdislov, V.A.; Vogel, J.; Iakovenko, L.V. Activation of Na,K-ATPase by small concentrations of D2O, inhibition by high concentrations. Biofizika 1978, 23, 390–391. [Google Scholar] [PubMed]
- Lobyshev, V.I.; Fogel, I.; Iakovenko, L.V.; Rezaeva, M.N.; Tverdislov, V.A. D2O as a modifier of ionic specificity of Na, K-ATPase. Biofizika 1982, 27, 595–603. [Google Scholar]
- Li, X.; Snyder, M. Can heavy isotopes increase lifespan? Studies of relative abundance in various organisms reveal chemical perspectives on aging. Bioessays 2016, 38, 1093–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosenkov, A.V.; Gulyaev, M.V.; Lobyshev, V.I.; Yusubalieva, G.M.; Baklaushev, V.P. The reversible effect of deuteration on tissue fluid and biopolymers in normal and tumor tissues of mice. Biophysics 2018, 63, 820–824. [Google Scholar] [CrossRef]
- Yaglova, N.V.; Obernikhin, S.S.; Timokhina, E.P.; Yaglov, V.V. Response of Pituitary-Thyroid Axis to a Short-Term Shift in Deuterium Content in the Body. Bull. Exp. Biol. Med. 2021, 171, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gaetani, M.; Chernobrovkin, A.; Zubarev, R.A. Anticancer Effect of Deuterium Depleted Water—Redox Disbalance Leads to Oxidative Stress. Mol. Cell. Proteom. 2019, 18, 2373–2387. [Google Scholar] [CrossRef] [Green Version]
- Somlyai, G.; Jancso, G.; Jakli, G.; Vass, K.; Barna, B.; Lakics, V.; Gaal, T. Naturally occurring deuterium is essential for the normal growth rate of cells. FEBS Lett. 1993, 317, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Massart, C.; Hoste, C.; Virion, A.; Ruf, J.; Dumont, J.E.; Van Sande, J. Cell biology of H2O2 generation in the thyroid: Investigation of the control of dual oxidases (DUOX) activity in intact ex vivo thyroid tissue and cell lines. Mol. Cell. Endocrinol. 2011, 343, 32–44. [Google Scholar] [CrossRef]
- Carvalho, D.P.; Dupuy, C.; Gorin, Y.; Legue, O.; Pommier, J.; Haye, B.; Virion, A. The Ca2+- and reduced nicotinamide adenine dinucleotide phosphate-dependent hydrogen peroxide generating system is induced by thyrotropin in porcine thyroid cells. Endocrinology 1996, 137, 1007–1012. [Google Scholar] [CrossRef]
- Simmons, E.M.; Hartwig, J.F. On the interpretation of deuterium kinetic isotope effects in C-H bond functionalizations by transition-metal complexes. Angew. Chem. Int. Ed. Engl. 2012, 51, 3066–3072. [Google Scholar] [CrossRef]
- Sestili, P.; Brigotti, M.; Calcabrini, C.; Turrini, E.; Arfilli, V.; Carnicelli, D.; Lucarini, M.; Mazzanti, A.; Milelli, A.; Righi, V.; et al. Deuterium Incorporation Protects Cells from Oxidative Damage. Oxid. Med. Cell. Longev. 2019, 2019, 6528106. [Google Scholar] [CrossRef] [Green Version]
- Fahrenkrug, J.; Georg, B.; Hannibal, J.; Jorgensen, H.L. Hypophysectomy abolishes rhythms in rat thyroid hormones but not in the thyroid clock. J. Endocrinol. 2017, 233, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Gamble, K.L.; Berry, R.; Frank, S.J.; Young, M.E. Circadian clock control of endocrine factors. Nat. Rev. Endocrinol. 2014, 10, 466–475. [Google Scholar] [CrossRef] [Green Version]
- Diatroptov, M.E.; Simonova, E.Y.; Diatroptova, M.A. The infradian rthythms of cortisol and thyroid hormones at woman of reproductive age. I.P. Pavlov Russ. Med. Biol. Her. 2013, 21, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Mendel, C.M.; Weisinger, R.A.; Jones, A.L.; Cavalieri, R.R. Thyroid hormone-binding proteins in plasma facilitate uniform distribution of thyroxine within tissues: A perfused rat liver study. Endocrinology 1987, 120, 1742–1749. [Google Scholar] [CrossRef]
- Yamauchi, K.; Ishihara, A. Evolutionary changes to transthyretin: Developmentally regulated and tissue-specific gene expression. FEBS J. 2009, 276, 5357–5366. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, N. Iodide transport in the thyroid gland. Biochim. Biophys. Acta 1993, 1154, 65–82. [Google Scholar] [CrossRef]
- Dohan, O.; De la Vieja, A.; Paroder, V.; Riedel, C.; Artani, M.; Reed, M.; Ginter, C.S.; Carrasco, N. The sodium/iodide Symporter (NIS): Characterization, regulation, and medical significance. Endocr. Rev. 2003, 24, 48–77. [Google Scholar] [CrossRef] [Green Version]
- Portulano, C.; Paroder-Belenitsky, M.; Carrasco, N. The Na+/I− Symporter (NIS): Mechanism and Medical Impact. Endocr. Rev. 2014, 35, 106–149. [Google Scholar] [CrossRef] [Green Version]
- Yaglova, N.V.; Yaglov, V.V. Mechanisms of Disruptive Action of Dichlorodiphenyltrichloroethane (DDT) on the Function of Thyroid Follicular Epitheliocytes. Bull. Exp. Biol. Med. 2015, 160, 231–233. [Google Scholar] [CrossRef]
- Pesce, L.; Kopp, P. Iodide transport: Implications for health and disease. Int. J. Pediatr. Endocrinol. 2014, 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olgun, A. Biological effects of deuteronation: ATP synthase as an example. Theor. Biol. Med. Model. 2007, 22, 4–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaglova, N.V.; Obernikhin, S.S.; Timokhina, E.P.; Yaglov, V.V.; Tsomartova, D.A.; Nazimova, S.V.; Tsomartova, E.S.; Ivanova, M.Y.; Chereshneva, E.V.; Lomanovskaya, T.A. Bilateral Shifts in Deuterium Supply Similarly Change Physiology of the Pituitary–Thyroid Axis, but Differentially Influence Na+/I− Symporter Production. Int. J. Mol. Sci. 2023, 24, 6803. https://doi.org/10.3390/ijms24076803
Yaglova NV, Obernikhin SS, Timokhina EP, Yaglov VV, Tsomartova DA, Nazimova SV, Tsomartova ES, Ivanova MY, Chereshneva EV, Lomanovskaya TA. Bilateral Shifts in Deuterium Supply Similarly Change Physiology of the Pituitary–Thyroid Axis, but Differentially Influence Na+/I− Symporter Production. International Journal of Molecular Sciences. 2023; 24(7):6803. https://doi.org/10.3390/ijms24076803
Chicago/Turabian StyleYaglova, Nataliya V., Sergey S. Obernikhin, Ekaterina P. Timokhina, Valentin V. Yaglov, Dibakhan A. Tsomartova, Svetlana V. Nazimova, Elina S. Tsomartova, Marina Y. Ivanova, Elizaveta V. Chereshneva, and Tatiana A. Lomanovskaya. 2023. "Bilateral Shifts in Deuterium Supply Similarly Change Physiology of the Pituitary–Thyroid Axis, but Differentially Influence Na+/I− Symporter Production" International Journal of Molecular Sciences 24, no. 7: 6803. https://doi.org/10.3390/ijms24076803
APA StyleYaglova, N. V., Obernikhin, S. S., Timokhina, E. P., Yaglov, V. V., Tsomartova, D. A., Nazimova, S. V., Tsomartova, E. S., Ivanova, M. Y., Chereshneva, E. V., & Lomanovskaya, T. A. (2023). Bilateral Shifts in Deuterium Supply Similarly Change Physiology of the Pituitary–Thyroid Axis, but Differentially Influence Na+/I− Symporter Production. International Journal of Molecular Sciences, 24(7), 6803. https://doi.org/10.3390/ijms24076803