
Mixed-Linkage Glucan Is the Main Carbohydrate Source and Starch Is an Alternative Source during Brachypodium Grain Germination

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Results
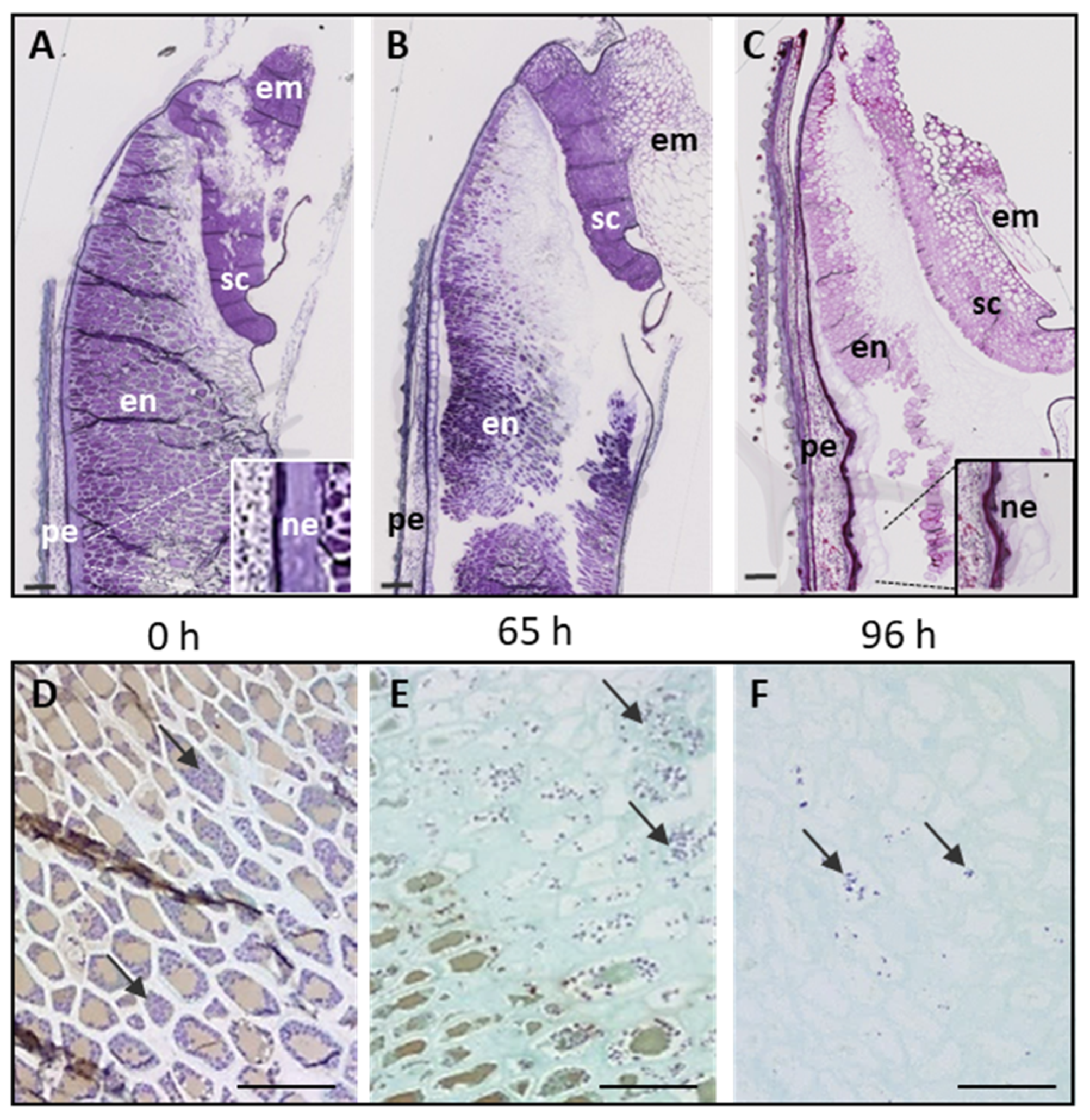
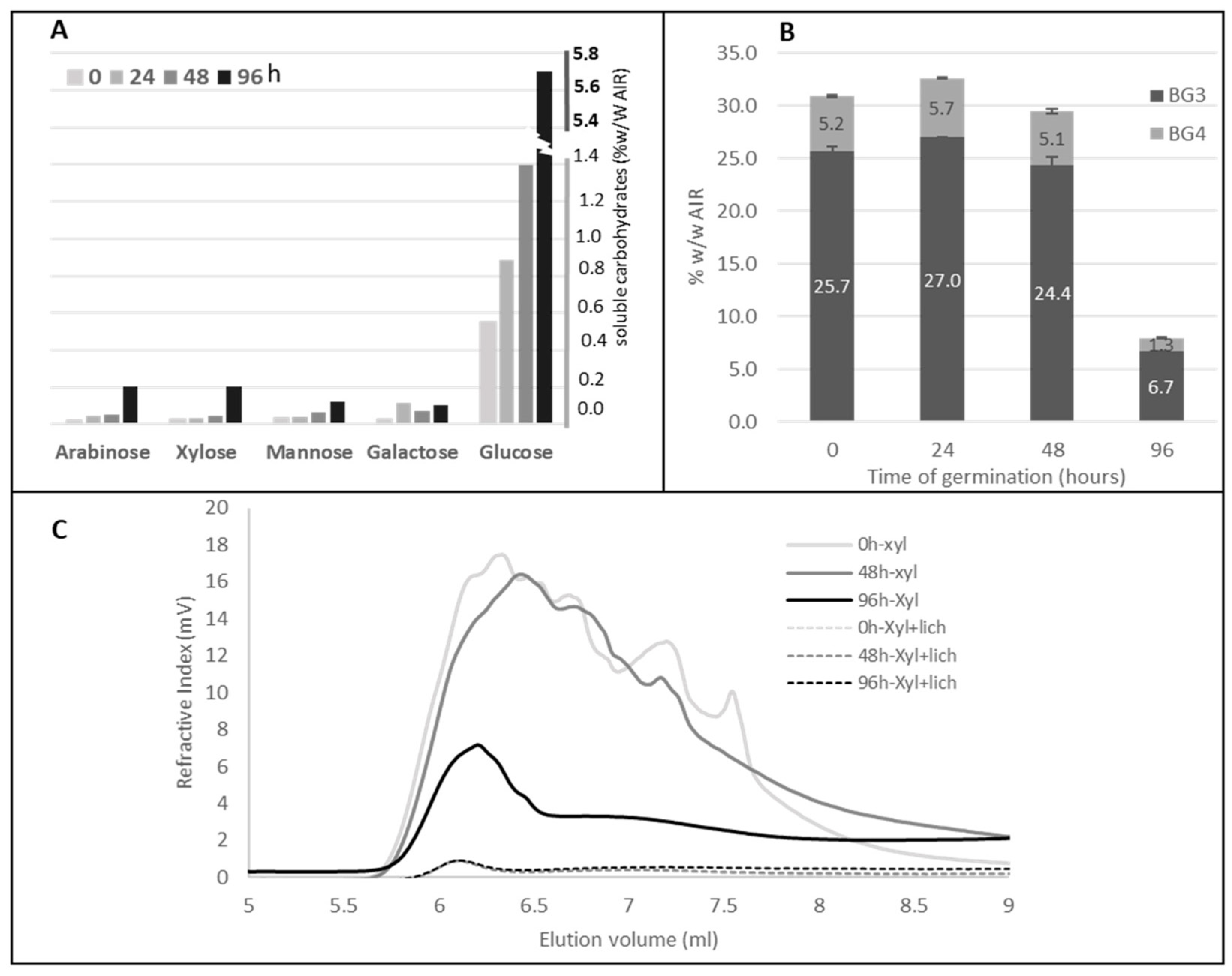
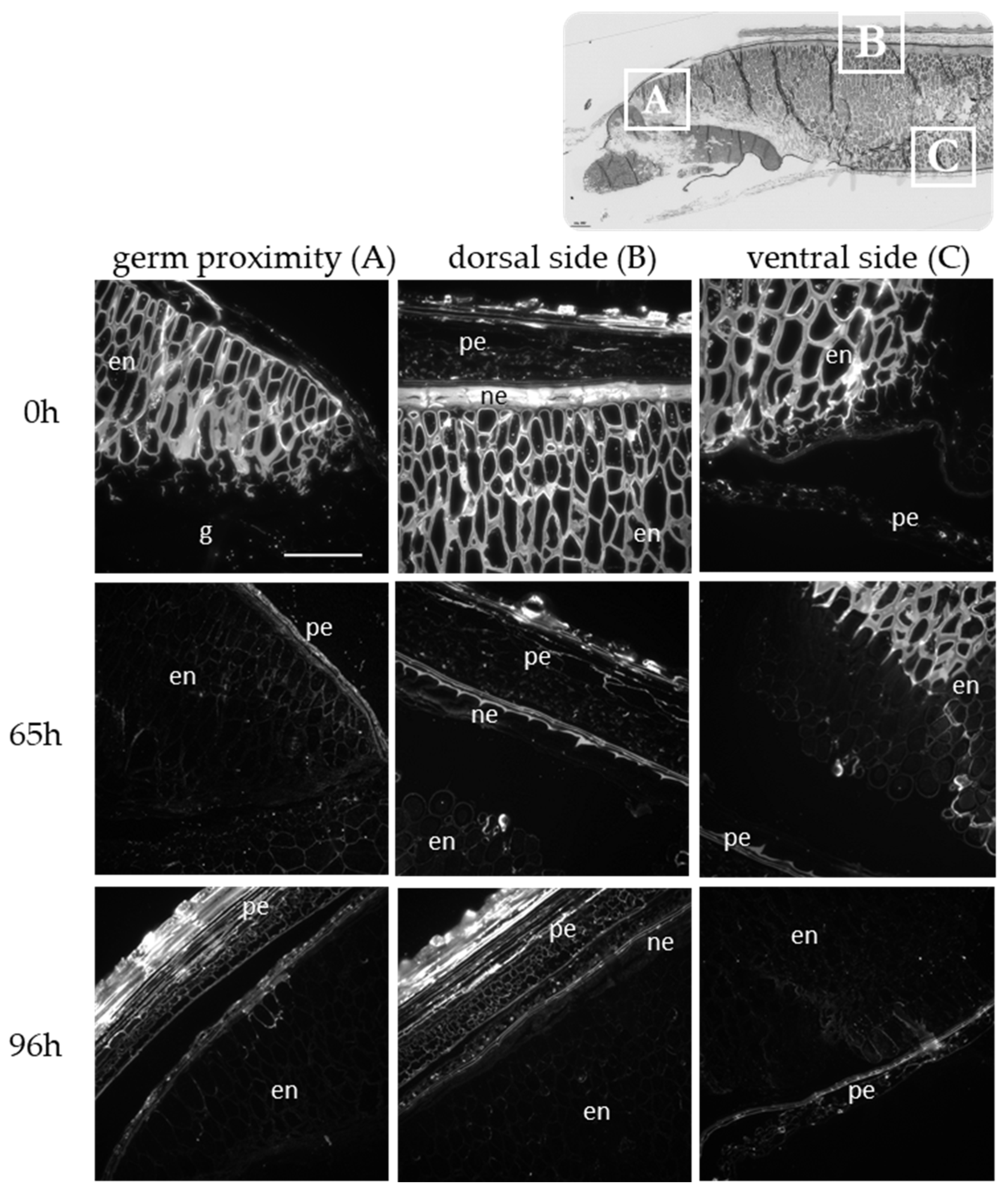
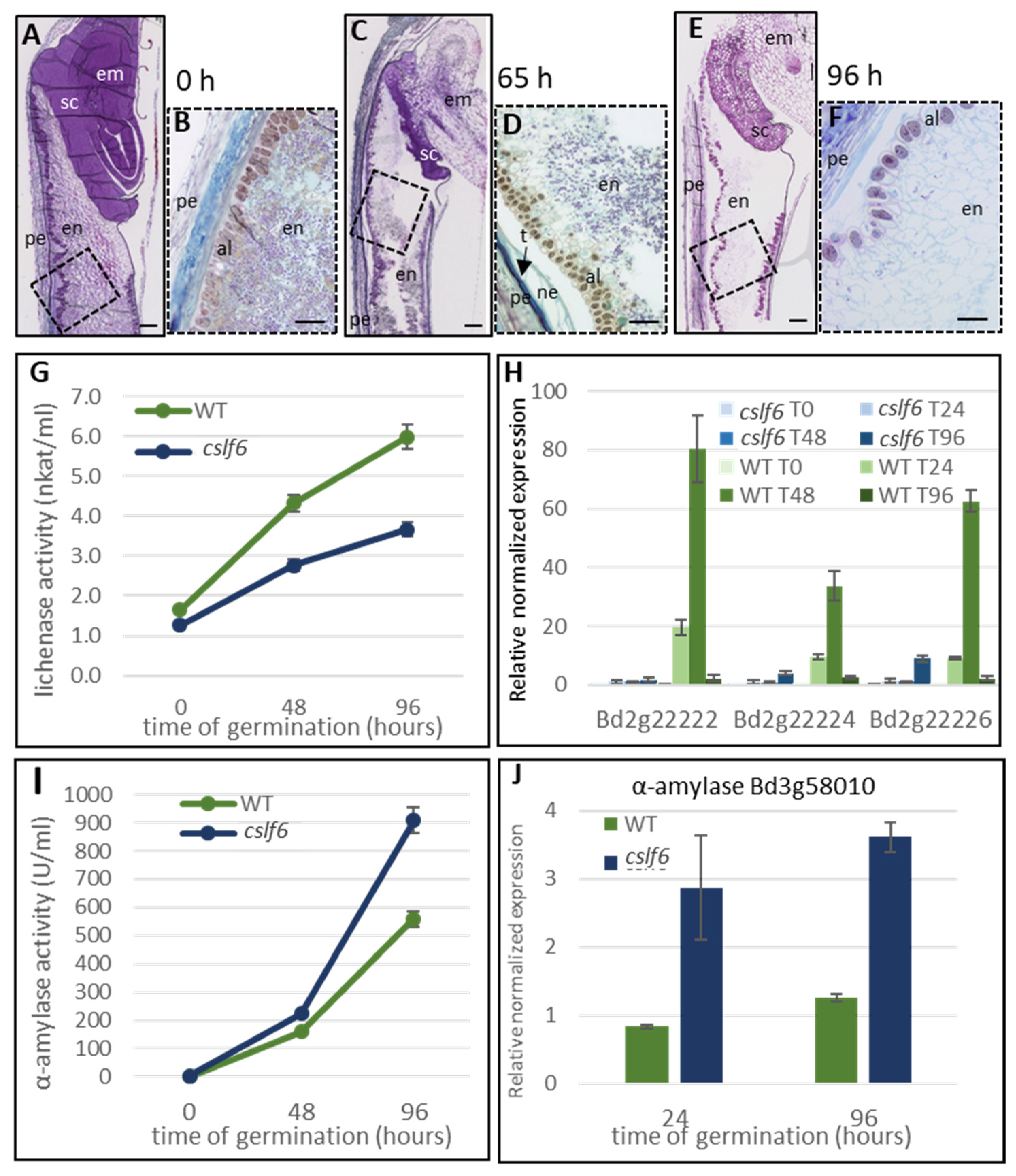
2.1. Histology and Polysaccharide Composition of Wild Type Brachypodium Grain during the Germination
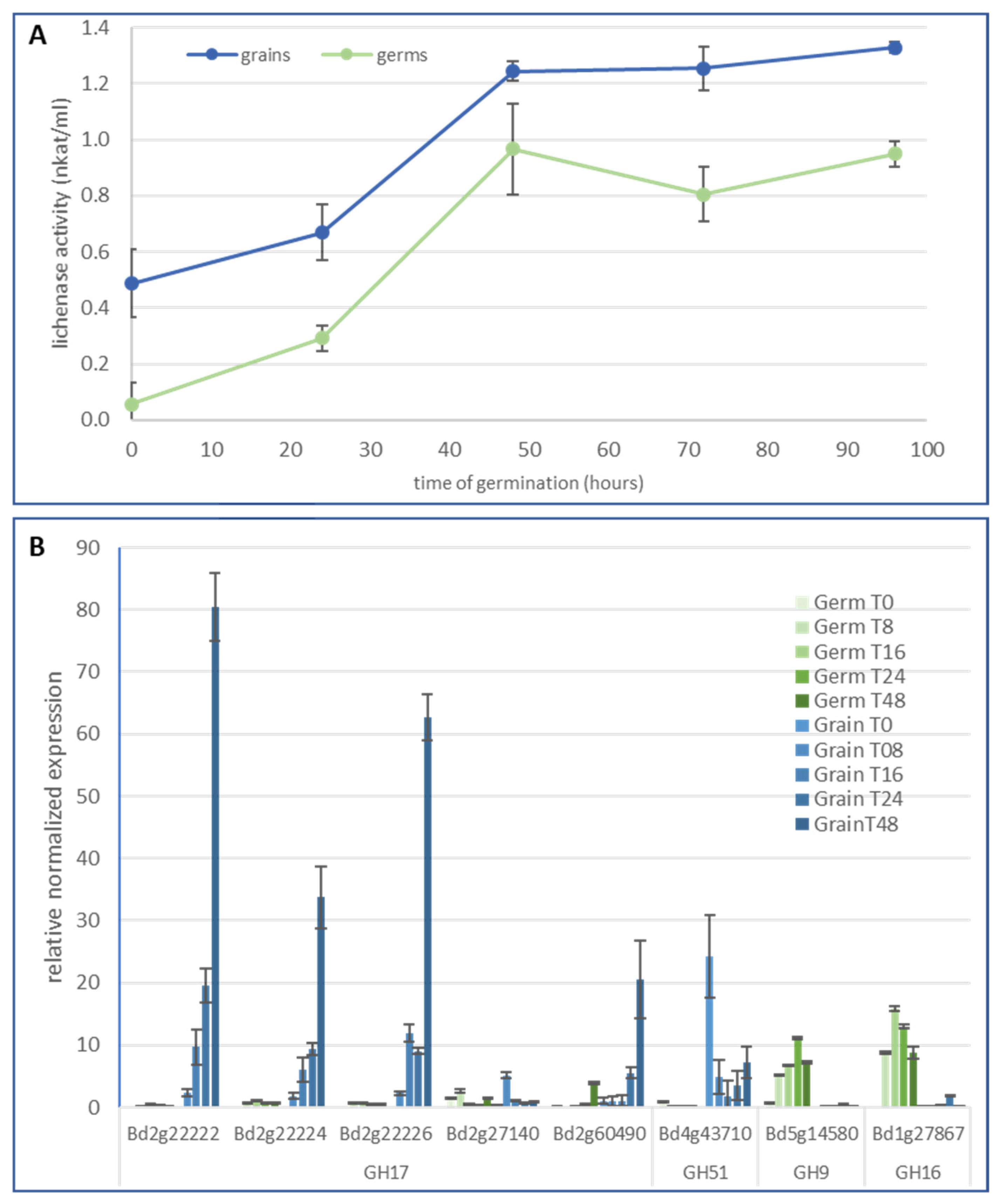
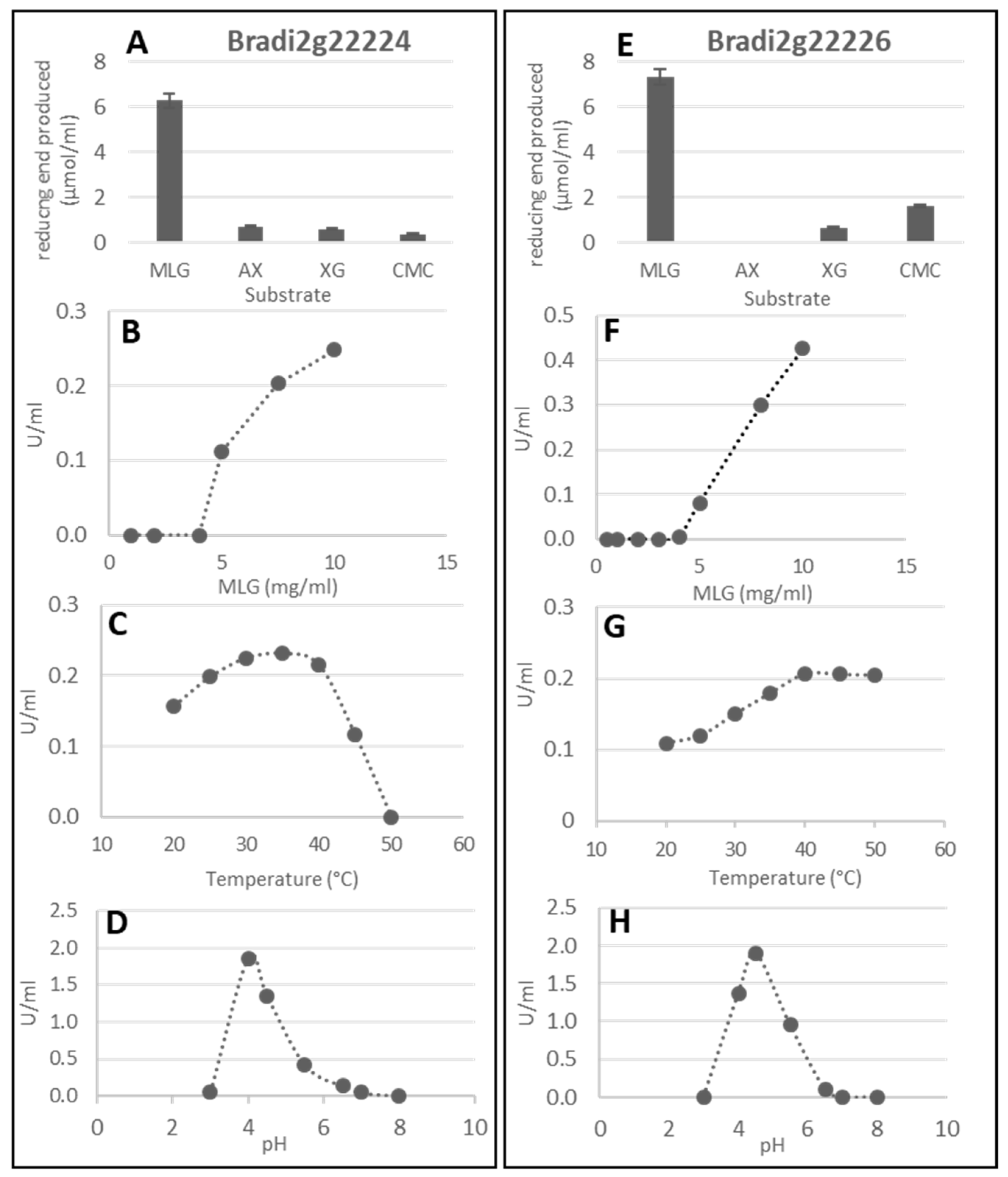
2.2. Lichenase Activity in Germinated Grains
2.3. Characterization of Brachypodium Mutants Affected in Genes Involved in MLG Biosynthesis
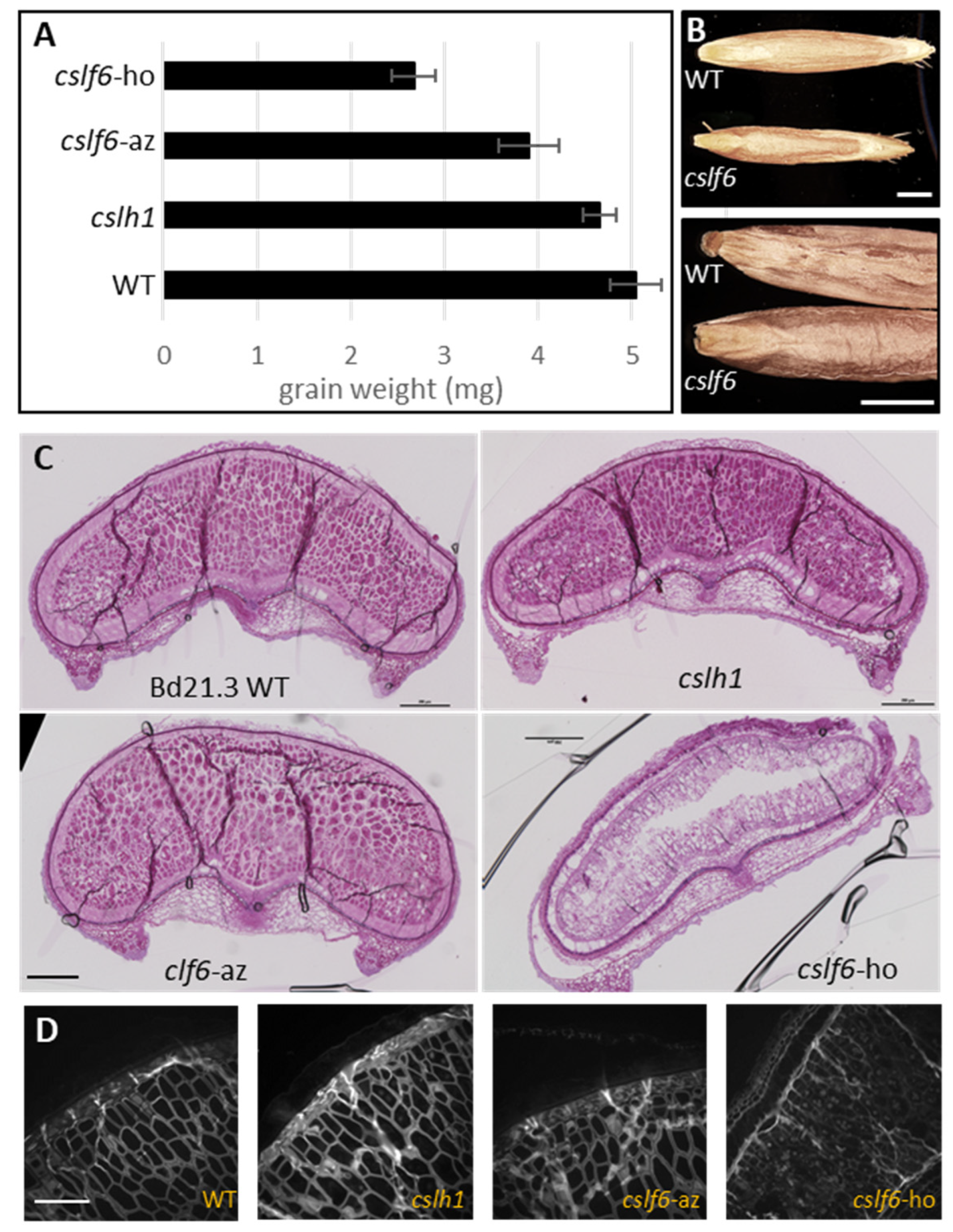
2.3.1. The cslf6 Mutant Grains Are Completely Devoid of MLG
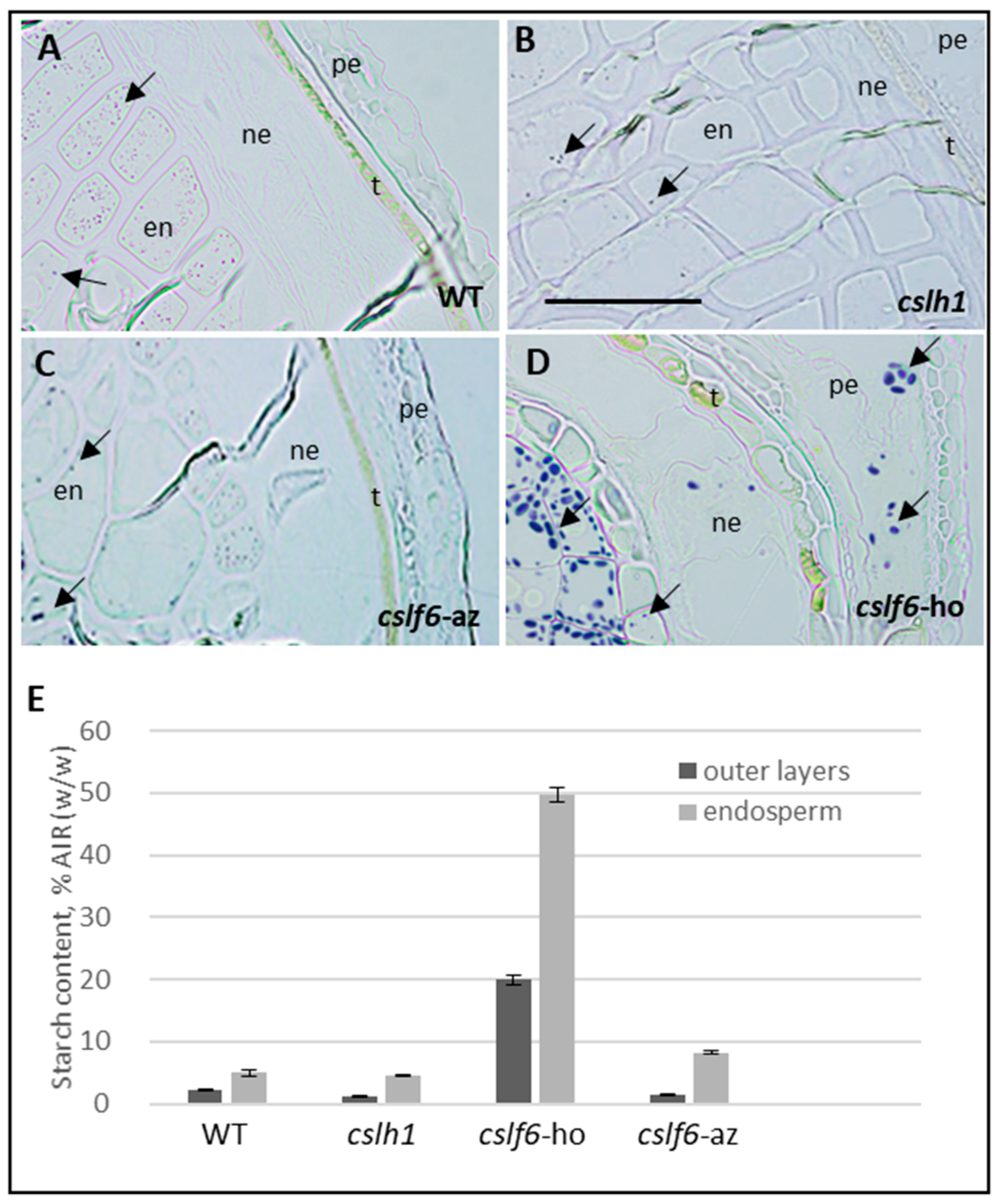
2.3.2. Loss of MLG Comes with an Increase of Starch in the cslf6 Mutant
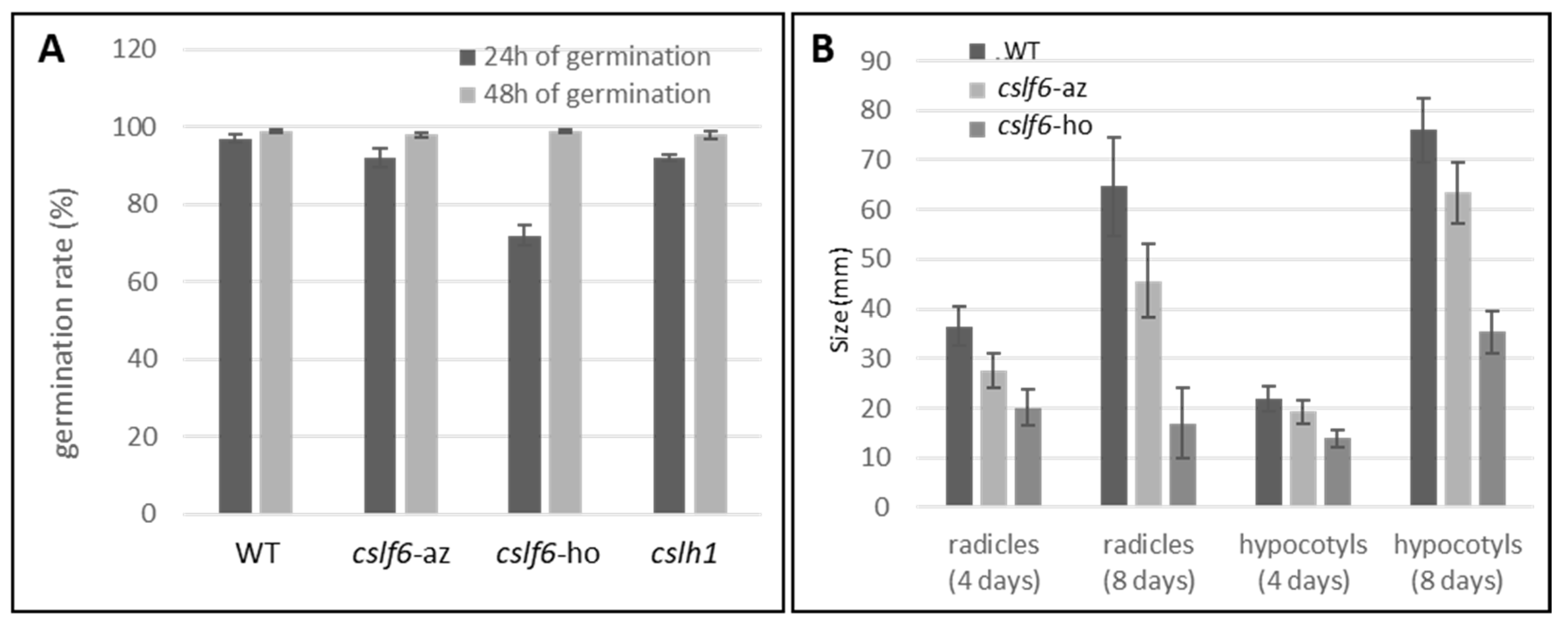
2.3.3. cslf6 Mutation Impact on Germination and Seedling Growth
3. Discussion
4. Material and Methods
4.1. Plant Culture
4.2. Seed Germination
4.3. Carbohydrate Analysis
4.3.1. Preparation of Alcohol-Insoluble Residue (AIR)
4.3.2. Neutral Sugar Determination
4.4. Starch and MLG Determination
4.5. HPSEC Analysis
4.6. Microscopic Analysis
4.6.1. Histological Staining
4.6.2. Immunolabeling
4.7. Cloning, Expression and Purification of Recombinant Lichenases
4.8. Protein Extraction
4.9. Enzymatic Activities
4.10. Quantitative RT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wolny, E.; Betekhtin, A.; Rojek, M.; Braszewska-Zalewska, A.; Lusinska, J.; Hasterok, R. Germination and the Early Stages of Seedling Development in Brachypodium distachyon. Int. J. Mol. Sci. 2018, 19, 2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, R.A.; Fincher, G.B. Current challenges in cell wall biology in the cereals and grasses. Front. Plant Sci. 2012, 3, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillon, F.; Larré, C.; Petipas, F.; Berger, A.; Moussawi, J.; Rogniaux, H.; Santoni, A.; Saulnier, L.; Jamme, F.; Miquel, M.; et al. A comprehensive overview of grain development in Brachypodium distachyon variety Bd21. J. Exp. Bot. 2012, 63, 739–755. [Google Scholar] [CrossRef] [Green Version]
- Trafford, K.; Haleux, P.; Henderson, M.; Parker, M.; Shirley, N.J.; Tucker, M.R.; Fincher, G.B.; Burton, R.A. Grain development in Brachypodium and other grasses: Possible interactions between cell expansion, starch deposition, and cell-wall synthesis. J. Exp. Bot. 2013, 64, 5033–5047. [Google Scholar] [CrossRef] [Green Version]
- Guillon, F.; Bouchet, B.; Jamme, F.; Robert, P.; Quemener, B.; Barron, C.; Larre, C.; Dumas, P.; Saulnier, L. Brachypodium distachyon grain: Characterization of endosperm cell walls. J. Exp. Bot. 2011, 62, 1001–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opanowicz, M.; Hands, P.; Betts, D.; Parker, M.L.; Toole, G.A.; Mills, E.N.C.; Doonan, J.H.; Drea, S. Endosperm development in Brachypodium distachyon. J. Exp. Bot. 2011, 62, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Hands, P.; Drea, S. A comparative view of grain development in Brachypodium distachyon. J. Cereal Sci. 2012, 56, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Jensen, J.K.; Zemelis-Durfee, S.; Kim, S.; Chan, J.; Beaudry, C.M.; Brandizzi, F.; Wilkerson, C.G. Disruption of Brachypodium Lichenase alters metabolism of mixed-linkage glucan and starch. Plant J. 2022, 109, 927–939. [Google Scholar] [CrossRef]
- Woodward, J.R.; Fincher, G.B. Substrate specificities and kinetic-properties of two (1→3),(1→4)-beta-D-glucan endo-hydrolases from germinating barley (Hordeum vulgare). Carbohydr. Res. 1982, 106, 111–122. [Google Scholar] [CrossRef]
- Fincher, G.B.; Lock, P.A.; Morgan, M.M.; Lingelbach, K.; Wettenhall, R.E.H.; Mercer, J.F.B.; Brandt, A.; Thomsen, K.K. Primary structure of the (1→3),(1→4)-beta-D-glucan 4-Glucanohydrolase from barley aleurone. Proc. Natl. Acad. Sci. USA 1986, 83, 2081–2085. [Google Scholar] [CrossRef] [Green Version]
- Litts, J.C.; Simmons, C.R.; Karrer, E.E.; Huang, N.; Rodriguez, R.L. The isolation and characterization of a barley 1,3-1,4-Beta-Glucanase gene. Eur. J. Biochem. 1990, 194, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Slakeski, N.; Baulcombe, D.C.; Devos, K.M.; Ahluwalia, B.; Doan, D.N.P.; Fincher, G.B. Structure and tissue-specific regulation of genes encoding Barley (1→3),(1→4)-beta-glucan endohydrolases. Mol. Gen. Genet. 1990, 224, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Slakeski, N.; Fincher, G.B. Developmental regulation of (1-3,1-4)beta-glucanase gene-expression in barley—Tissue-specific expression of individual isoenzymes. Plant Physiol. 1992, 99, 1226–1231. [Google Scholar] [CrossRef] [Green Version]
- Skendi, A.; Biliaderis, C.G.; Lazaridou, A.; Izydorczyk, M.S. Structure and rheological properties of water soluble beta-glucans from oat cultivars of Avena sativa and Avena bysantina. J. Cereal Sci. 2003, 38, 15–31. [Google Scholar] [CrossRef]
- Doblin, M.S.; Pettolino, F.A.; Wilson, S.M.; Campbell, R.; Burton, R.A.; Fincher, G.B.; Newbigin, E.; Bacic, A. A barley cellulose synthase-like CSLH gene mediates (1,3;1,4)-beta-D-glucan synthesis in transgenic Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 5996–6001. [Google Scholar] [CrossRef] [Green Version]
- Little, A.; Schwerdt, J.G.; Shirley, N.J.; Khor, S.F.; Neumann, K.; O’Donovan, L.A.; Lahnstein, J.; Collins, H.M.; Henderson, M.; Fincher, G.B.; et al. Revised Phylogeny of the Cellulose Synthase Gene Superfamily: Insights into Cell Wall Evolution. Plant Physiol. 2018, 177, 1124–1141. [Google Scholar] [CrossRef] [Green Version]
- Burton, R.A.; Jobling, S.A.; Harvey, A.J.; Shirley, N.J.; Mather, D.E.; Bacic, A.; Fincher, G.B. The genetics and transcriptional profiles of the cellulose synthase-like HvCslF gene family in barley. Plant Physiol. 2008, 146, 1821–1833. [Google Scholar] [CrossRef] [Green Version]
- Christensen, U.; Alonso-Simon, A.; Scheller, H.V.; Willats, W.G.T.; Harholt, J. Characterization of the primary cell walls of seedlings of Brachypodium distachyon—A potential model plant for temperate grasses. Phytochemistry 2010, 71, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Zemelis, S.; Keegstra, K.; Brandizzi, F. The cytoplasmic localization of the catalytic site of CSLF6 supports a channeling model for the biosynthesis of mixed-linkage glucan. Plant J. 2015, 81, 537–547. [Google Scholar] [CrossRef]
- Ermawar, R.A.; Collins, H.M.; Byrt, C.S.; Henderson, M.; O’Donovan, L.A.; Shirley, N.J.; Schwerdt, J.G.; Lahnstein, J.; Fincher, G.B.; Burton, R.A. Genetics and physiology of cell wall polysaccharides in the model C4 grass, Setaria viridis spp. BMC Plant Biol. 2015, 15, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, R.A.; Collins, H.M.; Kibble, N.A.J.; Smith, J.A.; Shirley, N.J.; Jobling, S.A.; Henderson, M.; Singh, R.R.; Pettolino, F.; Wilson, S.M.; et al. Over-expression of specific HvCslF cellulose synthase-like genes in transgenic barley increases the levels of cell wall (1,3;1,4)-β-d-glucans and alters their fine structure. Plant Biotechnol. J. 2011, 9, 117–135. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Zemelis-Durfee, S.; Jensen, J.K.; Wilkerson, C.G.; Keegstra, K.; Brandizzi, F. In the grass species Brachypodium distachyon, the production of mixed-linkage (1,3;1,4)-β-glucan (MLG) occurs in the Golgi apparatus. Plant J. 2018, 93, 1062–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonooka, T.; Aoki, E.; Yoshioka, T.; Taketa, S. A novel mutant gene for (1-3,1-4)-beta-D-glucanless grain on barley (Hordeum vulgare L.) chromosome 7H. Breed. Sci. 2009, 59, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Cory, A.T.; Baga, M.; Anyia, A.; Rossnagel, B.G.; Chibbar, R.N. Genetic markers for CslF6 gene associated with (1,3;1,4)-beta-glucan concentration in barley grain. J. Cereal Sci. 2012, 56, 332–339. [Google Scholar] [CrossRef]
- Taketa, S.; Yuo, T.; Tonooka, T.; Tsumuraya, Y.; Inagaki, Y.; Haruyama, N.; Larroque, O.; Jobling, S.A. Functional characterization of barley betaglucanless mutants demonstrates a unique role for CslF6 in (1,3;1,4)-beta-D-glucan biosynthesis. J. Exp. Bot. 2012, 63, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.S.; Burton, C.; Hong, Z.L.; Jackson, E. A mutation of the cellulose-synthase-like (CslF6) gene in barley (Hordeum vulgare L.) partially affects the beta-glucan content in grains. J. Cereal Sci. 2014, 59, 189–195. [Google Scholar] [CrossRef]
- Wong, S.C.; Shirley, N.J.; Little, A.; Khoo, K.H.P.; Schwerdt, J.; Fincher, G.B.; Burton, R.A.; Mather, D.E. Differential expression of the HvCslF6 gene late in grain development may explain quantitative differences in (1,3;1,4)-beta-glucan concentration in barley. Mol. Breed. 2015, 35, 20. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, C.; Freeman, J.; Jones, H.D.; Sparks, C.; Pellny, T.K.; Wilkinson, M.D.; Dunwell, J.; Andersson, A.A.M.; Aman, P.; Guillon, F.; et al. Down-regulation of the CSLF6 gene results in decreased (1,3;1,4)-beta-D-glucan in endosperm of wheat. Plant Physiol. 2010, 152, 1209–1218. [Google Scholar] [CrossRef] [Green Version]
- Vega-Sánchez, M.E.; Verhertbruggen, Y.; Christensen, U.; Chen, X.; Sharma, V.; Varanasi, P.; Jobling, S.A.; Talbot, M.; White, R.G.; Joo, M.; et al. Loss of Cellulose synthase-like F6 function affects mixed-linkage glucan deposition, cell wall mechanical properties, and defense responses in vegetative tissues of rice. Plant Physiol. 2012, 159, 56–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsia, M.M.; O’Malley, R.; Cartwright, A.; Nieu, R.; Gordon, S.P.; Kelly, S.; Williams, T.G.; Wood, D.F.; Zhao, Y.; Bragg, J.; et al. Sequencing and functional validation of the JGI Brachypodium distachyon T-DNA collection. Plant J. 2017, 91, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bain, M.; van de Meene, A.; Costa, R.; Doblin, M.S. Characterisation of Cellulose Synthase Like F6 (CslF6) Mutants Shows Altered Carbon Metabolism in β-D-(1,3;1,4)-Glucan Deficient Grain in Brachypodium distachyon. Front. Plant Sci. 2021, 11, 602850. [Google Scholar] [CrossRef] [PubMed]
- Vega-Sánchez, M.E.; Verhertbruggen, Y.; Scheller, H.V.; Ronald, P.C. Abundance of mixed linkage glucan in mature tissues and secondary cell walls of grasses. Plant Signal. Behav. 2013, 8, e23143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francin-Allami, M.; Alvarado, C.; Daniel, S.; Geairon, A.; Saulnier, L.; Guillon, F. Spatial and temporal distribution of cell wall polysaccharides during grain development of Brachypodium distachyon. Plant Sci. 2019, 280, 367–382. [Google Scholar] [CrossRef]
- Granier, F.; Lemaire, A.; Wang, Y.; LeBris, P.; Antelme, S.; Vogel, J.; Laudencia-Chingcuanco, D.; Sibout, R. Chemical and Radiation Mutagenesis: Induction and Detection by Whole Genome Sequencing BT. In Genetics and Genomics of Brachypodium; Vogel, J.P., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 155–170. ISBN 978-3-319-26944-3. [Google Scholar]
- Sibout, R.; Proost, S.; Hansen, B.O.; Vaid, N.; Giorgi, F.M.; Ho-Yue-Kuang, S.; Legee, F.; Cezart, L.; Bouchabke-Coussa, O.; Soulhat, C.; et al. Expression atlas and comparative coexpression network analyses reveal important genes involved in the formation of lignified cell wall in Brachypodium distachyon. New Phytol. 2017, 215, 1009–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmais, M.; Antelme, S.; Ho-Yue-Kuang, S.; Wang, Y.; Darracq, O.; d’Yvoire, M.B.; Cézard, L.; Légée, F.; Blondet, E.; Oria, N.; et al. A TILLING Platform for Functional Genomics in Brachypodium distachyon. PLoS ONE 2013, 8, e65503. [Google Scholar] [CrossRef] [Green Version]
- Tanackovic, V.; Svensson, J.T.; Jensen, S.L.; Buléon, A.; Blennow, A. The deposition and characterization of starch in Brachypodium distachyon. J. Exp. Bot. 2014, 65, 5179–5192. [Google Scholar] [CrossRef] [Green Version]
- Scholthof, K.-B.G.; Irigoyen, S.; Catalan, P.; Mandadi, K.K. Brachypodium: A Monocot Grass Model Genus for Plant Biology. Plant Cell 2018, 30, 1673–1694. [Google Scholar] [CrossRef] [Green Version]
- Hands, P.; Kourmpetli, S.; Sharples, D.; Harris, R.G.; Drea, S. Analysis of grain characters in temperate grasses reveals distinctive patterns of endosperm organization associated with grain shape. J. Exp. Bot. 2012, 63, 6253–6266. [Google Scholar] [CrossRef] [Green Version]
- Selvig, A.; Aarnes, H.; Lie, S. Cell-Wall Degradation in Endosperm of Barley During Germination. J. Inst. Brew. 1986, 92, 185–187. [Google Scholar] [CrossRef]
- Langenaeken, N.A.; Ieven, P.; Hedlund, E.G.; Kyomugasho, C.; van de Walle, D.; Dewettinck, K.; Van Loey, A.M.; Roeffaers, M.B.J.; Courtin, C.M. Arabinoxylan, β-glucan and pectin in barley and malt endosperm cell walls: A microstructure study using CLSM and cryo-SEM. Plant J. 2020, 103, 1477–1489. [Google Scholar] [CrossRef]
- Oliveira, L.A.; de Souza, G.A.; Silva, B.T.; Rocha, A.A.G.; Picoli, E.A.d.T.; Pereira, D.d.S.; Donzeles, S.M.L.; Ribeiro, M.d.F.; Ferreira, W.P.M. Histochemical approach of the mobilization of reserve compounds in germinating coffee seeds. Coffee Sci. 2020, 15, e151704. [Google Scholar] [CrossRef]
- Kosina, R.; Kaminska, K. The role of nucellar epidermisduring the germination of Brachypodium distachyon. Annu. Wheat Newsl. 2013, 59, 124. [Google Scholar]
- Chateigner-Boutin, A.L.; Suliman, M.; Bouchet, B.; Alvarado, C.; Lollier, V.; Rogniaux, H.; Guillon, F.; Larré, C. Endomembrane proteomics reveals putative enzymes involved in cell wall metabolism in wheat grain outer layers. J. Exp. Bot. 2015, 66, 2649–2658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerkovic, A.; Kriegel, A.M.; Bradner, J.R.; Atwell, B.J.; Roberts, T.H.; Willows, R.D. Strategic distribution of protective proteins within bran layers of wheat protects the nutrient-rich endosperm. Plant Physiol. 2010, 152, 1459–1470. [Google Scholar] [CrossRef] [Green Version]
- Buckeridge, M.S.; dos Santos, H.P.; Tine, M.A.S. Mobilisation of storage cell wall polysaccharides in seeds. Plant Physiol. Biochem. 2000, 38, 141–156. [Google Scholar] [CrossRef]
- Perrot, T.; Pauly, M.; Ramírez, V. Emerging Roles of β-Glucanases in Plant Development and Adaptative Responses. Plants 2022, 11, 1119. [Google Scholar] [CrossRef]
- Islam, M.Z.; An, H.-G.; Kang, S.-J.; Lee, Y.-T. Physicochemical and bioactive properties of a high β-glucan barley variety “Betaone” affected by germination processing. Int. J. Biol. Macromol. 2021, 177, 129–134. [Google Scholar] [CrossRef]
- Campbell, J.M.; Reid, J.S.G. Galactomannan Formation and Guanosine 5’-Diphosphate-Mannose–Galactomannan Mannosyltransferase in Developing Seeds of Fenugreek (Trigonella-Foenum-Graecum L., Leguminosae). Planta 1982, 155, 105–111. [Google Scholar] [CrossRef]
- Mccleary, B.V.; Clark, A.H.; Dea, I.C.M.; Rees, D.A. The Fine-Structures of Carob and Guar Galactomannans. Carbohydr. Res. 1985, 139, 237–260. [Google Scholar] [CrossRef]
- Kumar, C.S.; Bhattacharya, S. Tamarind seed: Properties, processing and utilization. Crit. Rev. Food Sci. Nutr. 2008, 48, 1–20. [Google Scholar] [CrossRef]
- Dervilly, G.; Leclercq, C.; Zimmermann, D.; Roue, C.; Thibault, J.F.; Saulnier, L. Isolation and characterization of high molar mass water-soluble arabinoxylans from barley and barley malt. Carbohydr. Polym. 2002, 47, 143–149. [Google Scholar] [CrossRef]
- Akiyama, T.; Jin, S.; Yoshida, M.; Hoshino, T.; Opassiri, R.; Ketudat Cairns, J.R. Expression of an endo-(1,3;1,4)-beta-glucanase in response to wounding, methyl jasmonate, abscisic acid and ethephon in rice seedlings. J. Plant Physiol. 2009, 166, 1814–1825. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.M.; Høj, P.B.; Fincher, G.B. Purification and characterization of (1→3, 1→4)-beta-glucan endohydrolases from germinated wheat (Triticum aestivum). Plant Mol. Biol. 1993, 22, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.J.; Martin, D.J.; Gengenbach, B.G.; Rines, H.W.; Somers, D.A. Sequence of a (1-3,1-4)-beta-glucanase cDNA from oat. Plant Physiol. 1993, 103, 295–296. [Google Scholar] [CrossRef]
- Kraemer, F.J.; Lunde, C.; Koch, M.; Kuhn, B.M.; Ruehl, C.; Brown, P.J.; Hoffmann, P.; Göhre, V.; Hake, S.; Pauly, M.; et al. A mixed-linkage (1,3;1,4)-β-D-glucan specific hydrolase mediates dark-triggered degradation of this plant cell wall polysaccharide. Plant Physiol. 2021, 185, 1559–1573. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Brandizzi, F. Advances in Cell Wall Matrix Research with a Focus on Mixed-Linkage Glucan. Plant Cell Physiol. 2021, 62, 1839–1846. [Google Scholar] [CrossRef]
- Purushotham, P.; Ho, R.; Yu, L.; Fincher, G.B.; Bulone, V.; Zimmer, J. Mechanism of mixed-linkage glucan biosynthesis by barley cellulose synthase-like CslF6 (1,3;1,4)-β-glucan synthase. Sci. Adv. 2022, 8, eadd1596. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.R.; Zhou, L.; Zhang, J.; Yu, H.; Xiong, F.; Wang, Z. Comparison of starch granule development and physicochemical properties of starches in wheat pericarp and endosperm. J. Sci. Food Agric. 2015, 95, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Dervilly, G.; Saulnier, L.; Roger, P.; Thibault, J.-F. Isolation of homogeneous fractions from wheat water-soluble arabinoxylans. Influence of the structure on their macromolecular characteristics. J. Agric. Food Chem. 2000, 48, 270–278. [Google Scholar] [CrossRef]
- Chateigner-Boutin, A.-L.; Bouchet, B.; Alvarado, C.; Bakan, B.; Guillon, F. The wheat grain contains pectic domains exhibiting specific spatial and development-associated distribution. PLoS ONE 2014, 9, e89620. [Google Scholar] [CrossRef]
- Guillon, F.; Tranquet, O.; Quillien, L.; Utille, J.-P.; Ordaz Ortiz, J.J.; Saulnier, L. Generation of polyclonal and monoclonal antibodies against arabinoxylans and their use for immunocytochemical location of arabinoxylans in cell walls of endosperm of wheat. J. Cereal Sci. 2004, 40, 167–182. [Google Scholar] [CrossRef]
- Sturgeon, R.J. Methods in Plant Biochemistry; Dey, P.M., Harb, J.B., Eds.; Academic Press: Cambridge, MA, USA, 1990; Volume 2, pp. 1–37. [Google Scholar]
- Molinari, H.B.C.; Pellny, T.K.; Freeman, J.; Shewry, P.R.; Mitchell, R.A.C. Grass cell wall feruloylation: Distribution of bound ferulate and candidate gene expression in Brachypodium distachyon. Front. Plant Sci. 2013, 4, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Niu, S.; Di, D.; Shi, L.; Liu, D.; Cao, X.; Miao, H.; Wang, X.; Han, C.; Yu, J.; et al. Selection of reference genes for gene expression studies in virus-infected monocots using quantitative real-time PCR. J. Biotechnol. 2013, 168, 7–14. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germination Time | ||||
|---|---|---|---|---|
| 0 h | 24 h | 48 h | 96 h | |
| Rhamnose | 0.3 +/− 0.0 | 0.2 +/− 0.1 | 0.2 +/− 0.0 | 0.2 +/− 0.1 |
| Fucose | ns | ns | ns | ns |
| Mannose | 0.5 +/− 0.1 | 0.5 +/− 0.1 | 0.4 +/− 0.2 | 0.5 +/− 0.1 |
| Galactose | 0.6 +/− 0.0 | 0.7 +/− 0.0 | 0.6 +/− 0.0 | 1.0 +/− 0.1 |
| Arabinose | 3.1 +/− 0.2 | 3.0 +/− 0.0 | 3.3 +/− 0.3 | 5.3 +/− 0.0 |
| Xylose | 8.8 +/− 0.3 | 6.0 +/− 0.3 | 7.5 +/− 0.5 | 12.9 +/− 0.7 |
| Glucose (total) | 48.9 +/− 0.4 | 46.4 +/− 2.0 | 42.1 +/− 0.7 | 32.1 +/− 2.6 |
| Starch | 5.0 +/− 0.1 | 5.3 +/− 0.1 | 5.2 +/− 0.0 | 3.0 +/− 0.1 |
| β-glucans | 30.9 +/− 0.5 | 32.6 +/− 0.1 | 29.5 +/− 1.0 | 7.9 +/− 0.0 |
| Total sugars | 62.2 +/− 0.3 | 56.8 +/− 2.4 | 53.8 +/− 0.2 | 52.0 +/− 3.5 |
| Mature Grain | Endosperm | Outer Layers | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| WT | cslf6-az | cslf6-ho | WT | cslf6-az | cslf6-ho | cslh1 | WT | cslf6-az | cslf6-ho | cslh1 | |
| Rhamnose | 0.3 +/− 0.1 | 0.3 +/− 0.0 | 0.2 +/− 0.0 | 0.4 +/− 0.1 | 0.3 +/− 0.1 | 0.3 +/− 0.0 | 0.3 +/− 0.0 | 0.3 +/− 0.1 | 0.5 +/− 0.1 | 0.2 +/− 0.0 | 0.4 +/− 0.0 |
| Fucose | 0.5 +/− 0.0 | 0.5 +/− 0.0 | 0.6 +/− 0.0 | 0.3 +/− 0.1 | 0.3 +/− 0.1 | 0.2 +/− 0.0 | ns | 0.3 +/− 0.0 | 0.4 +/− 0.0 | 0.3 +/− 0.1 | ns |
| Mannose | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Galactose | 0.6 +/− 0.1 | 0.7 +/− 0.2 | 1.6 +/− 0.1 | 1.2 +/− 0.1 | 0.7 +/− 0.3 | 1.5 +/− 0.3 | 0.8 +/− 0.2 | 1.0 +/− 0.2 | 0.8 +/− 0.2 | 1.9 +/− 0.1 | 0.6 +/− 0.0 |
| Arabinose | 2.9 +/− 0.3 | 3.0 +/− 0.1 | 7.3 +/− 0.1 | 2.2 +/− 0.2 | 2.2 +/− 0.2 | 3.5 +/− 0.1 | 2.5 +/− 0.0 | 4.3 +/− 0.1 | 4.3 +/− 0.4 | 8.5 +/− 0.3 | 4.1 +/− 0.0 |
| Xylose | 5.0 +/− 0.4 | 4.1 +/− 0.2 | 9.3 +/− 0.1 | 2.1 +/− 0.1 | 2.3 +/− 0.5 | 4.0 +/− 0.0 | 2.3 +/− 0.0 | 8.1 +/− 0.1 | 8.2 +/− 0.3 | 14.3 +/− 0.3 | 5.3 +/− 0.1 |
| Glucose (total) | 52.6 +/− 4.8 | 52.1 +/− 0.2 | 28.9 +/− 1.3 | 52.6 +/− 1.3 | 52.0 +/− 0.0 | 56.1 +/− 1.4 | 50.2 +/− 2.8 | 60.6 +/− 0.6 | 53.5 +/− 0.2 | 29.6 +/− 0.3 | 65.3 +/− 2.1 |
| β-glucans | 41.7 +/− 1.5 | 40.7 +/− 1.2 | ns | 46.8 +/− 0.1 | 48.5 +/− 1.1 | ns | 46.4 +/− 0.7 | 51.4 +/− 0.8 | 47.5 +/− 0.8 | ns | 50.0 +/− 0.2 |
| Ratio BG3/BG4 | 5.4 | 5.1 | ns | 5.4 | 5.5 | ns | 4.9 | 4 | 4 | ns | 3.9 |
| Starch | 5.3 +/− 0.1 | 5.4 +/− 0.0 | 26.1 +/− 0.8 | 5.0 +/− 0.6 | 8.3 +/− 0.2 | 49.8 +/− 1.2 | 4.5 +/− 0.1 | 2.2 +/− 0.1 | 1.4 +/− 0.0 | 19.9 +/− 0.7 | 1.2 +/− 0.0 |
| Total sugars | 62.3 +/− 5.2 | 60.7 +/− 1.5 | 47.9 +/− 0.0 | 53.4 +/− 4.2 | 57.8 +/− 1.9 | 65.7 +/− 1.0 | 56.4 +/− 3.0 | 74.3 +/− 3.0 | 67.8 +/− 1.0 | 55.0 +/− 0.9 | 76.0 +/− 2.3 |
| csf6-ho | ||
|---|---|---|
| 0 h | 96 h | |
| Rhamnose | 0.2 +/−0.0 | 0.2 +/−0.0 |
| Mannose | 0.7 +/−0.4 | 0.3 +/−0.2 |
| Fucose | 0.6 +/−0.0 | 0.6 +/−0.0 |
| Galactose | 1.5 +/−0.1 | 2.3 +/−0.1 |
| Arabinose | 6.4 +/−0.3 | 9.9 +/−0.1 |
| Xylose | 7.7 +/−0.3 | 18.0 +/−0.6 |
| Glucose (total) | 35.3 +/−5.0 | 21.5 +/−0.7 |
| Starch | 32.2 +/−0.2 | 7.2 +/−0.2 |
| β-glucans | ns | ns |
| Total sugars | 52.4 +/−4.8 | 52.9 +/−0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francin-Allami, M.; Bouder, A.; Geairon, A.; Alvarado, C.; Le-Bot, L.; Daniel, S.; Shao, M.; Laudencia-Chingcuanco, D.; Vogel, J.P.; Guillon, F.; et al. Mixed-Linkage Glucan Is the Main Carbohydrate Source and Starch Is an Alternative Source during Brachypodium Grain Germination. Int. J. Mol. Sci. 2023, 24, 6821. https://doi.org/10.3390/ijms24076821
Francin-Allami M, Bouder A, Geairon A, Alvarado C, Le-Bot L, Daniel S, Shao M, Laudencia-Chingcuanco D, Vogel JP, Guillon F, et al. Mixed-Linkage Glucan Is the Main Carbohydrate Source and Starch Is an Alternative Source during Brachypodium Grain Germination. International Journal of Molecular Sciences. 2023; 24(7):6821. https://doi.org/10.3390/ijms24076821
Chicago/Turabian StyleFrancin-Allami, Mathilde, Axelle Bouder, Audrey Geairon, Camille Alvarado, Lucie Le-Bot, Sylviane Daniel, Mingqin Shao, Debbie Laudencia-Chingcuanco, John P. Vogel, Fabienne Guillon, and et al. 2023. "Mixed-Linkage Glucan Is the Main Carbohydrate Source and Starch Is an Alternative Source during Brachypodium Grain Germination" International Journal of Molecular Sciences 24, no. 7: 6821. https://doi.org/10.3390/ijms24076821
APA StyleFrancin-Allami, M., Bouder, A., Geairon, A., Alvarado, C., Le-Bot, L., Daniel, S., Shao, M., Laudencia-Chingcuanco, D., Vogel, J. P., Guillon, F., Bonnin, E., Saulnier, L., & Sibout, R. (2023). Mixed-Linkage Glucan Is the Main Carbohydrate Source and Starch Is an Alternative Source during Brachypodium Grain Germination. International Journal of Molecular Sciences, 24(7), 6821. https://doi.org/10.3390/ijms24076821