
Strategy for Pre-Clinical Development of Active Targeting MicroRNA Oligonucleotide Therapeutics for Unmet Medical Needs



Abstract
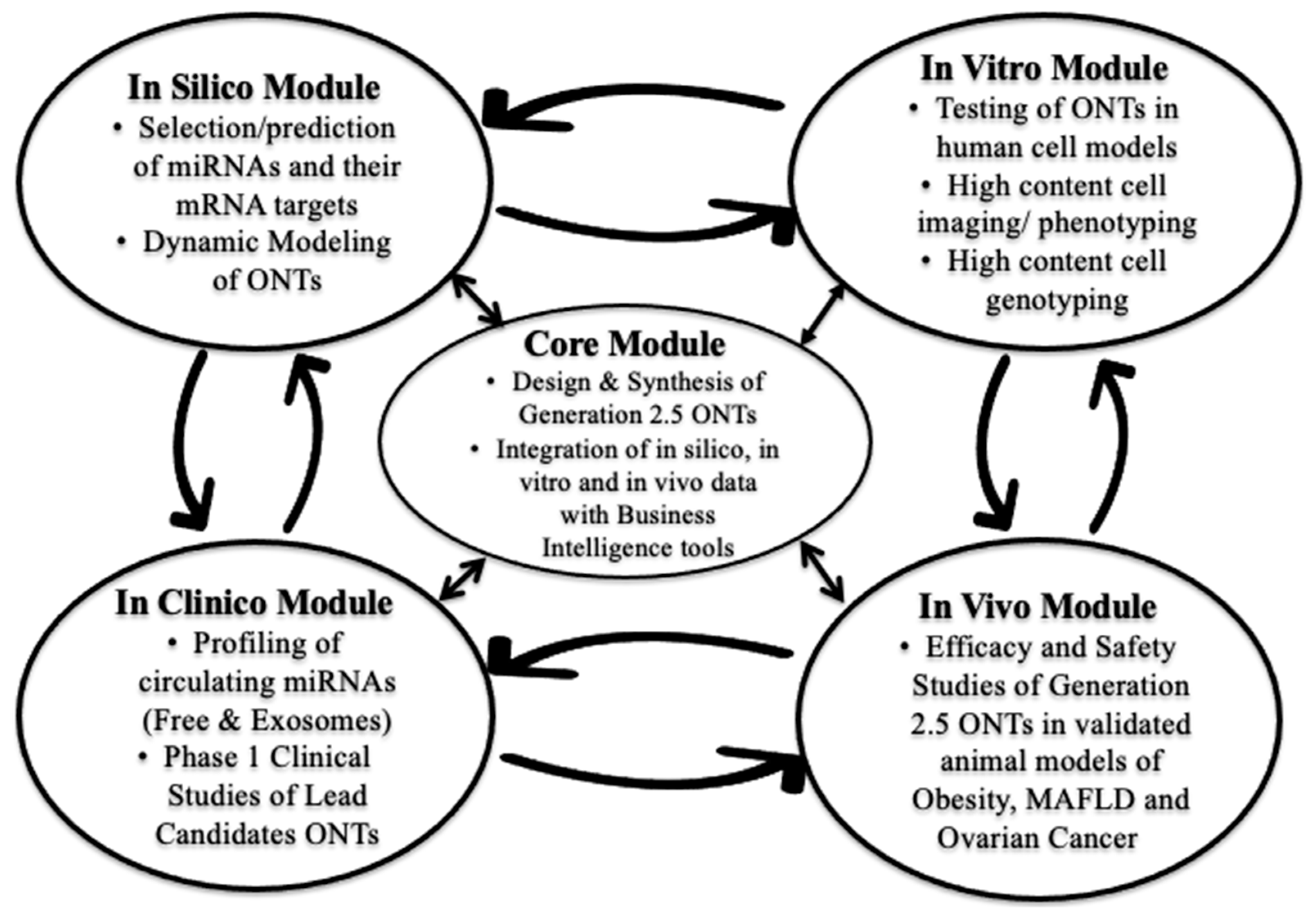
:1. Introduction
2. Results
2.1. Selection of MicroRNAs Involved in Adipose Tissue Functions to Treat Metabolic Pandemics
2.1.1. Background and Goal
2.1.2. Strategy
2.1.3. In Silico Search
2.1.4. Validation of miR-22-3p as a Metabolic Target
2.1.5. In Vitro and In Vivo Validation of miR-22-3p Antagonism
2.1.6. Safety Assessment
2.1.7. Design of Generation 2.5 ONTs for Active Targeted Delivery to Metabolic Tissues/Organs
- Eliminate potential toxicities by replacing PS and LNA modifications with a gamma PNA backbone;
- Maintain resistance to nucleases and proteases/peptidases;
- Avoid chirality;
- Limit binding to serum proteins;
- Optimize/simplify chemical synthesis;
- Conjugate ONT to a fatty acid or a short peptide for enhanced targeted delivery to adipocytes of a greatly reduced effective dose with an extended duration of action (mean residence time).
2.1.8. Selection of the Membrane Fatty Acid Translocase (FAT) Transporter for Active Targeted Delivery of ONTs to Adipocytes and Metabolic Organs
2.1.9. In Silico Modeling of Generation 2.5 ONTs
2.2. Selection and Targeting of MicroRNAs Involved in Ovarian Cancer Development and Spread
2.2.1. Background and Goal
2.2.2. Strategy
2.2.3. In Silico Search
2.2.4. Active Targeted Delivery of Generation 2.5 MiRNA-Based ONTs to Ovarian Cancer Cells via Folic Acid Receptor Alpha (FOLR1)
2.2.5. Active Targeted Delivery of Generation 2.5 miRNA-Based ONTs to the Adipocyte-Rich OC Tumor Microenvironment via FAT and FABP4 Transporters
2.2.6. In Vitro Testing of Generation 2.5 Candidate miRNA Agomirs and Antagomirs Will Be Conducted in Human Cells in Culture including the Following:
3. Discussion and Conclusions
4. Materials and Methods
4.1. In Vivo Experiments
4.2. MiR-22 Antagomir
4.3. Imaging Analyses
4.4. RNA Sequencing Analyses
4.5. Statistical Analyses
4.6. Studies Approval
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Patents
References
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grasser, F.A.; Lenhof, H.P.; et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Yedavilli, S.; Singh, A.D.; Singh, D.; Samal, R. Nano-Messengers of the Heart: Promising Theranostic Candidates for Cardiovascular Maladies. Front. Physiol. 2022, 13, 895322. [Google Scholar] [CrossRef]
- Henning, R.J. Cardiovascular Exosomes and MicroRNAs in Cardiovascular Physiology and Pathophysiology. J. Cardiovasc. Transl. Res. 2021, 14, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ni, Y.Q.; Xu, H.; Xiang, Q.Y.; Zhao, Y.; Zhan, J.K.; He, J.Y.; Li, S.; Liu, Y.S. Roles and mechanisms of exosomal non-coding RNAs in human health and diseases. Signal Transduct. Target. Ther. 2021, 6, 383. [Google Scholar] [CrossRef]
- Shu, Z.; Tan, J.; Miao, Y.; Zhang, Q. The role of microvesicles containing microRNAs in vascular endothelial dysfunction. J. Cell. Mol. Med. 2019, 23, 7933–7945. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Assmann, T.S.; Milagro, F.I.; Martinez, J.A. Crosstalk between microRNAs, the putative target genes and the lncRNA network in metabolic diseases. Mol. Med. Rep. 2019, 20, 3543–3554. [Google Scholar] [CrossRef]
- Bennett, C.F.; Kordasiewicz, H.B.; Cleveland, D.W. Antisense Drugs Make Sense for Neurological Diseases. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 831–852. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, M.; Condorelli, G.; Croce, C.M. MicroRNAs in diseases and drug response. Curr. Opin. Pharmacol. 2008, 8, 661–667. [Google Scholar] [CrossRef]
- Jae, N.; Dimmeler, S. Noncoding RNAs in Vascular Diseases. Circ. Res. 2020, 126, 1127–1145. [Google Scholar] [CrossRef]
- Chandan, K.; Gupta, M.; Sarwat, M. Role of Host and Pathogen-Derived MicroRNAs in Immune Regulation During Infectious and Inflammatory Diseases. Front. Immunol. 2019, 10, 3081. [Google Scholar] [CrossRef] [Green Version]
- Shah, V.; Shah, J. Recent trends in targeting miRNAs for cancer therapy. J. Pharm. Pharmacol. 2020, 72, 1732–1749. [Google Scholar] [CrossRef] [PubMed]
- Kousar, K.; Ahmad, T.; Abduh, M.S.; Kanwal, B.; Shah, S.S.; Naseer, F.; Anjum, S. miRNAs in Regulation of Tumor Microenvironment, Chemotherapy Resistance, Immunotherapy Modulation and miRNA Therapeutics in Cancer. Int. J. Mol. Sci. 2022, 23, 13822. [Google Scholar] [CrossRef] [PubMed]
- Veiga, R.N.; Zambalde, E.P.; Cox, L.; Jucoski, T.S.; Kohler, A.F.; Carvalho, T.M.; Rodrigues, A.C.; Ludwig, B.; Crowley, K.; de Oliveira, J.C.; et al. Regulation of Immune Cells by microRNAs and microRNA-Based Cancer Immunotherapy. Adv. Exp. Med. Biol. 2022, 1385, 75–108. [Google Scholar] [CrossRef] [PubMed]
- Otmani, K.; Rouas, R.; Lewalle, P. OncomiRs as noncoding RNAs having functions in cancer: Their role in immune suppression and clinical implications. Front. Immunol. 2022, 13, 913951. [Google Scholar] [CrossRef] [PubMed]
- Momin, M.Y.; Gaddam, R.R.; Kravitz, M.; Gupta, A.; Vikram, A. The Challenges and Opportunities in the Development of MicroRNA Therapeutics: A Multidisciplinary Viewpoint. Cells 2021, 10, 3097. [Google Scholar] [CrossRef] [PubMed]
- Bajan, S.; Hutvagner, G. RNA-Based Therapeutics: From Antisense Oligonucleotides to miRNAs. Cells 2020, 9, 137. [Google Scholar] [CrossRef] [Green Version]
- Corey, D.R.; Damha, M.J.; Manoharan, M. Challenges and Opportunities for Nucleic Acid Therapeutics. Nucleic Acid 2022, 32, 8–13. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.S. Therapeutic advances of miRNAs: A preclinical and clinical update. J. Adv. Res. 2021, 28, 127–138. [Google Scholar] [CrossRef]
- Bartolucci, D.; Pession, A.; Hrelia, P.; Tonelli, R. Precision Anti-Cancer Medicines by Oligonucleotide Therapeutics in Clinical Research Targeting Undruggable Proteins and Non-Coding RNAs. Pharmaceutics 2022, 14, 1453. [Google Scholar] [CrossRef]
- Taniguchi, H.; Suzuki, Y.; Imai, K.; Adachi, Y. Antitumoral RNA-targeted oligonucleotide therapeutics: The third pillar after small molecule inhibitors and antibodies. Cancer Sci. 2022, 113, 2952–2961. [Google Scholar] [CrossRef]
- Lamb, Y.N. Inclisiran: First Approval. Drugs 2021, 81, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.F.; Baker, B.F.; Pham, N.; Swayze, E.; Geary, R.S. Pharmacology of Antisense Drugs. Annu. Rev. Pharm. Toxicol. 2017, 57, 81–105. [Google Scholar] [CrossRef] [PubMed]
- Crooke, S.T.; Baker, B.F.; Crooke, R.M.; Liang, X.H. Antisense technology: An overview and prospectus. Nat. Rev. Drug Discov. 2021, 20, 427–453. [Google Scholar] [CrossRef]
- Crooke, S.T.; Liang, X.H.; Baker, B.F.; Crooke, R.M. Antisense Technology: A Review. J. Biol. Chem. 2021, 100416. [Google Scholar] [CrossRef]
- Alhamadani, F.; Zhang, K.; Parikh, R.; Wu, H.; Rasmussen, T.P.; Bahal, R.; Zhong, X.B.; Manautou, J.E. Adverse Drug Reactions and Toxicity of the Food and Drug Administration-Approved Antisense Oligonucleotide Drugs. Drug Metab. Dispos. 2022, 50, 879–887. [Google Scholar] [CrossRef]
- Migliorati, J.M.; Liu, S.; Liu, A.; Gogate, A.; Nair, S.; Bahal, R.; Rasmussen, T.P.; Manautou, J.E.; Zhong, X.B. Absorption, Distribution, Metabolism, and Excretion of US Food and Drug Administration-Approved Antisense Oligonucleotide Drugs. Drug Metab. Dispos. 2022, 50, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.M.; Tu, M.J. Deliver the promise: RNAs as a new class of molecular entities for therapy and vaccination. Pharm. Ther. 2022, 230, 107967. [Google Scholar] [CrossRef]
- Moumne, L.; Marie, A.C.; Crouvezier, N. Oligonucleotide Therapeutics: From Discovery and Development to Patentability. Pharmaceutics 2022, 14, 260. [Google Scholar] [CrossRef]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.J. Therapeutic siRNA: State of the art. Signal Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for microRNA Therapeutics and Clinical Research. Front. Genet. 2019, 10, 478. [Google Scholar] [CrossRef] [Green Version]
- Kara, G.; Arun, B.; Calin, G.A.; Ozpolat, B. miRacle of microRNA-Driven Cancer Nanotherapeutics. Cancers 2022, 14, 3818. [Google Scholar] [CrossRef] [PubMed]
- Suparpprom, C.; Vilaivan, T. Perspectives on conformationally constrained peptide nucleic acid (PNA): Insights into the structural design, properties and applications. RSC Chem. Biol. 2022, 3, 648–697. [Google Scholar] [CrossRef]
- Sahu, B.; Chenna, V.; Lathrop, K.L.; Thomas, S.M.; Zon, G.; Livak, K.J.; Ly, D.H. Synthesis of conformationally preorganized and cell-permeable guanidine-based gamma-peptide nucleic acids (gammaGPNAs). J. Org. Chem. 2009, 74, 1509–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaddam, R.R.; Dhuri, K.; Kim, Y.R.; Jacobs, J.S.; Kumar, V.; Li, Q.; Irani, K.; Bahal, R.; Vikram, A. gamma Peptide Nucleic Acid-Based miR-122 Inhibition Rescues Vascular Endothelial Dysfunction in Mice Fed a High-Fat Diet. J. Med. Chem. 2022, 65, 3332–3342. [Google Scholar] [CrossRef]
- Quijano, E.; Bahal, R.; Ricciardi, A.; Saltzman, W.M.; Glazer, P.M. Therapeutic Peptide Nucleic Acids: Principles, Limitations, and Opportunities. Yale J. Biol. Med. 2017, 90, 583–598. [Google Scholar] [PubMed]
- Smolarz, B.; Durczynski, A.; Romanowicz, H.; Szyllo, K.; Hogendorf, P. miRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [Google Scholar] [CrossRef]
- de Rooij, L.A.; Mastebroek, D.J.; Ten Voorde, N.; van der Wall, E.; van Diest, P.J.; Moelans, C.B. The microRNA Lifecycle in Health and Cancer. Cancers 2022, 14, 5748. [Google Scholar] [CrossRef]
- Dwivedi, S.K.D.; Rao, G.; Dey, A.; Mukherjee, P.; Wren, J.D.; Bhattacharya, R. Small Non-Coding-RNA in Gynecological Malignancies. Cancers 2021, 13, 1085. [Google Scholar] [CrossRef]
- Hashemipour, M.; Boroumand, H.; Mollazadeh, S.; Tajiknia, V.; Nourollahzadeh, Z.; Rohani Borj, M.; Pourghadamyari, H.; Rahimian, N.; Hamblin, M.R.; Mirzaei, H. Exosomal microRNAs and exosomal long non-coding RNAs in gynecologic cancers. Gynecol. Oncol. 2021, 161, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Beg, A.; Parveen, R.; Fouad, H.; Yahia, M.E.; Hassanein, A.S. Role of different non-coding RNAs as ovarian cancer biomarkers. J. Ovarian Res. 2022, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Liu, Y.; Shang, L.; Zhou, F.; Yang, S. EMT-associated microRNAs and their roles in cancer stemness and drug resistance. Cancer Commun. 2021, 41, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Shai, A.; Galouk, E.; Miari, R.; Tareef, H.; Sammar, M.; Zeidan, M.; Rayan, A.; Falah, M. Inhibiting mutant KRAS G12D gene expression using novel peptide nucleic acid-based antisense: A potential new drug candidate for pancreatic cancer. Oncol. Lett. 2022, 23, 130. [Google Scholar] [CrossRef]
- Tastan, B.; Tarakcioglu, E.; Birinci, Y.; Park, Y.; Genc, S. Role of Exosomal MicroRNAs in Cell-to-Cell Communication. Methods Mol. Biol. 2022, 2257, 269–292. [Google Scholar] [CrossRef]
- Sheiner, L.B. Learning versus confirming in clinical drug development. Clin. Pharmacol. Ther. 1997, 61, 275–291. [Google Scholar] [CrossRef]
- Cecchini, M. Use of healthcare services and expenditure in the US in 2025: The effect of obesity and morbid obesity. PLoS ONE 2018, 13, e0206703. [Google Scholar] [CrossRef]
- Okunogbe, A.; Nugent, R.; Spencer, G.; Ralston, J.; Wilding, J. Economic impacts of overweight and obesity: Current and future estimates for eight countries. BMJ Glob. Health 2021, 6, e006351. [Google Scholar] [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J.; International Consensus, P. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014 e1991. [Google Scholar] [CrossRef]
- Correa, L.H.; Heyn, G.S.; Magalhaes, K.G. The Impact of the Adipose Organ Plasticity on Inflammation and Cancer Progression. Cells 2019, 8, 662. [Google Scholar] [CrossRef] [Green Version]
- Kurylowicz, A. microRNAs in Human Adipose Tissue Physiology and Dysfunction. Cells 2021, 10, 3342. [Google Scholar] [CrossRef] [PubMed]
- Rakib, A.; Kiran, S.; Mandal, M.; Singh, U.P. MicroRNAs: A crossroad that connects obesity to immunity and aging. Immun. Ageing 2022, 19, 64. [Google Scholar] [CrossRef]
- Ma, L.; Gilani, A.; Yi, Q.; Tang, L. MicroRNAs as Mediators of Adipose Thermogenesis and Potential Therapeutic Targets for Obesity. Biology 2022, 11, 1657. [Google Scholar] [CrossRef] [PubMed]
- Traube, F.R.; Carell, T. The chemistries and consequences of DNA and RNA methylation and demethylation. RNA Biol. 2017, 14, 1099–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tateishi, K.; Okada, Y.; Kallin, E.M.; Zhang, Y. Role of Jhdm2a in regulating metabolic gene expression and obesity resistance. Nature 2009, 458, 757–761. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Li, X.; Shi, Z.; Bai, X.; Xia, Y.; Zheng, Y.; Xu, D.; Chen, F.; You, Y.; Fang, J.; et al. KDM3A Senses Oxygen Availability to Regulate PGC-1alpha-Mediated Mitochondrial Biogenesis. Mol. Cell 2019, 76, 885–895 e887. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]
- Keller, A.; Groger, L.; Tschernig, T.; Solomon, J.; Laham, O.; Schaum, N.; Wagner, V.; Kern, F.; Schmartz, G.P.; Li, Y.; et al. miRNATissueAtlas2: An update to the human miRNA tissue atlas. Nucleic Acids Res. 2022, 50, D211–D221. [Google Scholar] [CrossRef]
- Thibonnier, M.; Esau, C. Metabolic Benefits of MicroRNA-22 Inhibition. Nucleic Acid Ther. 2020, 30, 104–116. [Google Scholar] [CrossRef]
- Thibonnier, M.; Esau, C.; Ghosh, S.; Wargent, E.; Stocker, C. Metabolic and energetic benefits of microRNA-22 inhibition. BMJ Open Diabetes Res. Care 2020, 8, e001478. [Google Scholar] [CrossRef]
- Glatz, J.F.C.; Luiken, J. Dynamic role of the transmembrane glycoprotein CD36 (SR-B2) in cellular fatty acid uptake and utilization. J. Lipid Res. 2018, 59, 1084–1093. [Google Scholar] [CrossRef] [Green Version]
- Glatz, J.F.; Nabben, M.; Heather, L.C.; Bonen, A.; Luiken, J.J. Regulation of the subcellular trafficking of CD36, a major determinant of cardiac fatty acid utilization. Biochim. Biophys. Acta 2016, 1861, 1461–1471. [Google Scholar] [CrossRef]
- Jing, Z.; Qi, R.; Thibonnier, M.; Ren, P. Molecular Dynamics Study of the Hybridization between RNA and Modified Oligonucleotides. J. Chem. Theory Comput. 2019, 15, 6422–6432. [Google Scholar] [CrossRef]
- Jing, Z.; Ren, P. Molecular Dynamics Simulations of Protein RNA Complexes by Using an Advanced Electrostatic Model. J. Phys. Chem. B 2022, 126, 7343–7353. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Lu, C.; Jing, Z.; Wu, C.; Piquemal, J.P.; Ponder, J.W.; Ren, P. AMOEBA Polarizable Atomic Multipole Force Field for Nucleic Acids. J. Chem. Theory Comput. 2018, 14, 2084–2108. [Google Scholar] [CrossRef] [PubMed]
- Mosa, R.; Huang, L.; Wu, Y.; Fung, C.; Mallawakankanamalage, O.; LeRoith, D.; Chen, C. Hexarelin, a Growth Hormone Secretagogue, Improves Lipid Metabolic Aberrations in Nonobese Insulin-Resistant Male MKR Mice. Endocrinology 2017, 158, 3174–3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marechal, L.; Laviolette, M.; Rodrigue-Way, A.; Sow, B.; Brochu, M.; Caron, V.; Tremblay, A. The CD36-PPARgamma Pathway in Metabolic Disorders. Int. J. Mol. Sci. 2018, 19, 1529. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Lu, X.; Huang, L.; Zhang, C.; Veldhuis, J.D.; Cowley, M.A.; Chen, C. Stimulation of endogenous pulsatile growth hormone secretion by activation of growth hormone secretagogue receptor reduces the fat accumulation and improves the insulin sensitivity in obese mice. FASEB J. 2021, 35, e21269. [Google Scholar] [CrossRef]
- Webb, P.M.; Jordan, S.J. Epidemiology of epithelial ovarian cancer. Best Pract. Res. Clin. Obs. Gynaecol. 2017, 41, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Dinkelspiel, H.E.; Champer, M.; Hou, J.; Tergas, A.; Burke, W.M.; Huang, Y.; Neugut, A.I.; Ananth, C.V.; Hershman, D.L.; Wright, J.D. Long-term mortality among women with epithelial ovarian cancer. Gynecol. Oncol. 2015, 138, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Meng, F.; Zhong, Z. Emerging targeted drug delivery strategies toward ovarian cancer. Adv. Drug Deliv. Rev. 2021, 178, 113969. [Google Scholar] [CrossRef]
- Heh, E.; Allen, J.; Ramirez, F.; Lovasz, D.; Fernandez, L.; Hogg, T.; Riva, H.; Holland, N.; Chacon, J. Peptide Drug Conjugates and Their Role in Cancer Therapy. Int. J. Mol. Sci. 2023, 24, 829. [Google Scholar] [CrossRef]
- Yoshida, K.; Yokoi, A.; Kato, T.; Ochiya, T.; Yamamoto, Y. The clinical impact of intra- and extracellular miRNAs in ovarian cancer. Cancer Sci. 2020, 111, 3435–3444. [Google Scholar] [CrossRef]
- Zavesky, L.; Jandakova, E.; Weinberger, V.; Hanzikova, V.; Slanar, O.; Kohoutova, M. Ascites in ovarian cancer: MicroRNA deregulations and their potential roles in ovarian carcinogenesis. Cancer Biomark. 2022, 33, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Strumidlo, A.; Skiba, S.; Scott, R.J.; Lubinski, J. The potential role of miRNAs in therapy of breast and ovarian cancers associated with BRCA1 mutation. Hered. Cancer Clin. Pract. 2017, 15, 15. [Google Scholar] [CrossRef] [Green Version]
- Staicu, C.E.; Predescu, D.V.; Rusu, C.M.; Radu, B.M.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. Role of microRNAs as Clinical Cancer Biomarkers for Ovarian Cancer: A Short Overview. Cells 2020, 9, 169. [Google Scholar] [CrossRef] [Green Version]
- Segal, M.; Slack, F.J. Challenges identifying efficacious miRNA therapeutics for cancer. Expert Opin. Drug Discov. 2020, 15, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Rattanapan, Y.; Korkiatsakul, V.; Kongruang, A.; Siriboonpiputtana, T.; Rerkamnuaychoke, B.; Chareonsirisuthigul, T. MicroRNA Expression Profiling of Epithelial Ovarian Cancer Identifies New Markers of Tumor Subtype. Microrna 2020, 9, 289–294. [Google Scholar] [CrossRef]
- Liang, Z.; Lu, Z.; Zhang, Y.; Shang, D.; Li, R.; Liu, L.; Zhao, Z.; Zhang, P.; Lin, Q.; Feng, C.; et al. Targeting Membrane Receptors of Ovarian Cancer Cells for Therapy. Curr. Cancer Drug Targets 2019, 19, 449–467. [Google Scholar] [CrossRef]
- Mirahmadi, Y.; Nabavi, R.; Taheri, F.; Samadian, M.M.; Ghale-Noie, Z.N.; Farjami, M.; Samadi-Khouzani, A.; Yousefi, M.; Azhdari, S.; Salmaninejad, A.; et al. MicroRNAs as Biomarkers for Early Diagnosis, Prognosis, and Therapeutic Targeting of Ovarian Cancer. J. Oncol. 2021, 2021, 3408937. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, D.M.; Matulonis, U.A.; Birrer, M.J.; Castro, C.M.; Gilbert, L.; Vergote, I.; Martin, L.P.; Mantia-Smaldone, G.M.; Martin, A.G.; Bratos, R.; et al. Phase Ib study of mirvetuximab soravtansine, a folate receptor alpha (FRalpha)-targeting antibody-drug conjugate (ADC), in combination with bevacizumab in patients with platinum-resistant ovarian cancer. Gynecol. Oncol. 2020, 157, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Heo, Y.A. Etranacogene Dezaparvovec: First Approval. Drugs 2023, 83, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.; Oaknin, A.; Matulonis, U.A.; Mantia-Smaldone, G.M.; Lim, P.C.; Castro, C.M.; Provencher, D.; Memarzadeh, S.; Method, M.; Wang, J.; et al. Safety and efficacy of mirvetuximab soravtansine, a folate receptor alpha (FRalpha)-targeting antibody-drug conjugate (ADC), in combination with bevacizumab in patients with platinum-resistant ovarian cancer. Gynecol. Oncol. 2023, 170, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.; Dessi, J.; Self, C.H. Preclinical evaluation of light-activatable, bispecific anti-human CD3 antibody conjugates as anti-ovarian cancer therapeutics. MAbs 2009, 1, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Nam, S.M.; Moon, A. Antibody-drug conjugates and bispecific antibodies targeting cancers: Applications of click chemistry. Arch. Pharm. Res. 2023, 46, 131–148. [Google Scholar] [CrossRef]
- Chen, S.H.; Dominik, P.K.; Stanfield, J.; Ding, S.; Yang, W.; Kurd, N.; Llewellyn, R.; Heyen, J.; Wang, C.; Melton, Z.; et al. Dual checkpoint blockade of CD47 and PD-L1 using an affinity-tuned bispecific antibody maximizes antitumor immunity. J. Immunother. Cancer 2021, 9, e003464. [Google Scholar] [CrossRef]
- Bax, H.J.; Chauhan, J.; Stavraka, C.; Santaolalla, A.; Osborn, G.; Khiabany, A.; Grandits, M.; Lopez-Abente, J.; Palhares, L.; Chan Wah Hak, C.; et al. Folate receptor alpha in ovarian cancer tissue and patient serum is associated with disease burden and treatment outcomes. Br. J. Cancer 2022, 128, 342–353. [Google Scholar] [CrossRef]
- Kobel, M.; Madore, J.; Ramus, S.J.; Clarke, B.A.; Pharoah, P.D.; Deen, S.; Bowtell, D.D.; Odunsi, K.; Menon, U.; Morrison, C.; et al. Evidence for a time-dependent association between FOLR1 expression and survival from ovarian carcinoma: Implications for clinical testing. An Ovarian Tumour Tissue Analysis consortium study. Br. J. Cancer 2014, 111, 2297–2307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markert, S.; Lassmann, S.; Gabriel, B.; Klar, M.; Werner, M.; Gitsch, G.; Kratz, F.; Hasenburg, A. Alpha-folate receptor expression in epithelial ovarian carcinoma and non-neoplastic ovarian tissue. Anticancer Res. 2008, 28, 3567–3572. [Google Scholar]
- Liu, J.; Liu, L.; Antwi, P.A.; Luo, Y.; Liang, F. Identification and Validation of the Diagnostic Characteristic Genes of Ovarian Cancer by Bioinformatics and Machine Learning. Front. Genet. 2022, 13, 858466. [Google Scholar] [CrossRef]
- Scaranti, M.; Cojocaru, E.; Banerjee, S.; Banerji, U. Exploiting the folate receptor alpha in oncology. Nat. Rev. Clin. Oncol. 2020, 17, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Motohara, T.; Masuda, K.; Morotti, M.; Zheng, Y.; El-Sahhar, S.; Chong, K.Y.; Wietek, N.; Alsaadi, A.; Carrami, E.M.; Hu, Z.; et al. An evolving story of the metastatic voyage of ovarian cancer cells: Cellular and molecular orchestration of the adipose-rich metastatic microenvironment. Oncogene 2019, 38, 2885–2898. [Google Scholar] [CrossRef] [Green Version]
- Stocker, C.J.; Wargent, E.; O’Dowd, J.; Cornick, C.; Speakman, J.R.; Arch, J.R.; Cawthorne, M.A. Prevention of diet-induced obesity and impaired glucose tolerance in rats following administration of leptin to their mothers. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1810–R1818. [Google Scholar] [CrossRef]
- Arch, J.R.; Trayhurn, P. Detection of thermogenesis in rodents in response to anti-obesity drugs and genetic modification. Front. Physiol. 2013, 4, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M. KEGG Bioinformatics Resource for Plant Genomics and Metabolomics. Methods Mol. Biol. 2016, 1374, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Slenter, D.N.; Kutmon, M.; Hanspers, K.; Riutta, A.; Windsor, J.; Nunes, N.; Melius, J.; Cirillo, E.; Coort, S.L.; Digles, D.; et al. WikiPathways: A multifaceted pathway database bridging metabolomics to other omics research. Nucleic Acids Res. 2018, 46, D661–D667. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Final Score | Positive Combo |
|---|---|---|
| hsa-miR-22-3p | 12.11 | AKT1,BDNF,CDKN1A,CREB1,ESR1,HDAC4,HDAC6,KDM3A,KDM6B,KLF6,MAPK14,MECP2,PPARA,PPARGC1B,PRDM16,PTEN,RUNX2,SIRT1,SOD2,SP1,STAT3 |
| hsa-miR-520c-3p | 4.58 | AKT1,CDKN1A,ESR1,HDAC4,KDM6B,KLF6,MECP2,PPARA,PPARGC1B,PRDM16,PTEN,RUNX2,SP1,STAT3 |
| hsa-miR-10b-5p | 4.55 | BDNF,CDKN1A,CREB1,ESR1,HDAC4,KDM3A,PPARA,PPARGC1B,PRDM16,PTEN,SOD2,SP1 |
| hsa-miR-1470 | 4.35 | AKT1,CREB1,HDAC4,KDM6B,KLF6,MAPK14,MECP2,PPARA,PPARGC1B,PTEN,RUNX2,SOD2,STAT3 |
| hsa-miR-5089-3p | 4.29 | AKT1,CREB1,KLF6,MAPK14,MECP2,PRDM16,PTEN,SIRT1,SOD2,STAT3 |
| hsa-miR-7110-5p | 4.12 | CDKN1A,CREB1,KDM6B,KLF6,MECP2,PPARA,PPARGC1B,PRDM16,PTEN,RUNX2,SP1,STAT3 |
| hsa-miR-30a-5p | 4.1 | AKT1,BDNF,CREB1,KDM3A,KDM6B,KLF6,PPARGC1B,PRDM16,PTEN,RUNX2,SIRT1,SOD2 |
| hsa-miR-520a-3p | 3.95 | AKT1,CREB1,ESR1,HDAC4,KDM6B,KLF6,MECP2,PPARGC1B,PRDM16,PTEN,RUNX2,SOD2,STAT3 |
| hsa-miR-520b | 3.83 | AKT1,ESR1,HDAC4,KDM6B,KLF6,MECP2,PPARA,PPARGC1B,PRDM16,PTEN,RUNX2,SOD2,STAT3 |
| hsa-miR-365a-5p | 3.77 | CDKN1A,ESR1,HDAC4,KDM6B,KLF6,MECP2,PPARA,PPARGC1B,PRDM16,SOD2,STAT3 |
| Tool | Web Address | Function |
|---|---|---|
| Target Scan Human 8.0 | www.targetscan.org/vert_80 (accessed on 7 November 2022) | Search for predicted miRNA targets |
| metaMIR V 1.1.0 | http://rna.informatik.uni-freiburg.de/metaMIR/Input.jsp (accessed on 7 November 2022) | Predict interactions between miRNAs and clusters of genes in human |
| OncomiR | http://www.oncomir.org/ (accessed on 7 November 2022) | WashU Pan-Cancer miRNome Atlas exploring pan-cancer microRNA dysregulation |
| GeneNet V 3.0.2 | http://www.oncomir.org/ (accessed on 7 November 2022) | R package for learning high-dimensional dependency networks from genomic data |
| Cytoscape V3.7.1 | https://cytoscape.org/ (accessed on 7 November 2022) | Network data integration, analysis and visualization |
| DiffCorr V0.4.2 | https://sourceforge.net/projects/diffcorr/ (accessed on 7 November 2022) | R package to analyze differential correlations biological networks |
| STRING V11.5 | https://string-db.org/ (accessed on 7 November 2022) | Protein–protein interaction networks Functional enrichment analysis |
| Webgestalt V2019 | http://www.webgestalt.org/ (accessed on 7 November 2022) | Gene list over-representation analysis |
| ABCC3 | CALR | CUL4A | FZD2 | KLF9 | NRXN3 | RB1 | STK4 |
| ABL2 | CANX | CXCL1 | FZD6 | KLLN | NSD1 | RBBP8 | STMN2 |
| ACAP2 | CARD18 | CXCL10 | FZD8 | KRAS | NUAK1 | RHOBTB3 | STX17 |
| ACO2 | CASP10 | CXCL11 | GAB2 | LATS2 | OLA1 | RHOC | STXBP4 |
| ACSL4 | CASP8 | CXCL12 | GADD45B | LHX6 | OLFML3 | RNF44 | SUCO |
| ACTC1 | CCL5 | CXCL8 | GALNT1 | LIMK1 | OVOL1 | ROCK1 | SYNCRIP |
| ACTR1A | CCNB1 | CXCL9 | GALNT14 | LOX | P4HA1 | ROCK2 | TAGLN |
| ACTRT3 | CCND1 | CXCR3 | GALR1 | LPIN1 | PA1 | RUNX1 | TAP1 |
| ADAM12 | CCND2 | CYP1B1 | GCNT1 | LRRC15 | PAK2 | RUNX2 | TCF21 |
| ADAM17 | CCNE1 | CYTIP | GCNT2 | LRRK2 | PAPD7 | RUNX3 | TCF4 |
| ADAM19 | CCNG1 | DAAM1 | GCNT4 | LSG1 | PARP1 | S1PR1 | TCF7L1 |
| ADAMDEC1 | CCNG2 | DCN | GCOM1 | LUM | PAX7 | SALL2 | TEX261 |
| ADAMTS17 | CCR2 | DCTN5 | GCSAM | LZTS1 | PCDHA10 | SDC1 | TGFB1 |
| ADAMTS19 | CD1D | DCX | GEMIN4 | MACC1 | PCDHA3 | SEMA4D | THBS2 |
| ADAMTSL1 | CD2 | DDB2 | GFPT2 | MAP2 | PCDHA5 | SEMA6B | TIMM17A |
| AGO1 | CD247 | DICER1 | GM2A | MAP3K1 | PCDHGA10 | SEPTIN6 | TIMMDC1 |
| AKAP13 | CD27 | DKK1 | GNAI3 | MAP3K7 | PCNA | SET | TIMP2 |
| AKR1D1 | CD38 | DLG2 | GPR12 | MAPK1 | PDCD6 | SGCD | TIMP3 |
| AKT1 | CD3D | DLGAP2 | GPR124 | MAPK14 | PDE7A | SHROOM2 | TLN1 |
| AKT2 | CD3E | DNMT1 | GPR83 | MAPK3 | PDGFRA | SIK1 | TLR4 |
| AKT3 | CD44 | DTD2 | GRB7 | MCM2 | PDGFRB | SIK2 | TMEM239 |
| ALG2 | CD55 | DVL3 | HBEGF | MED12L | PDHB | SIRT1 | TMEM45A |
| ANKRD46 | CD68 | E2F2 | HDGF | MET | PDZK1IP1 | SIT1 | TP53 |
| ANXA8L1 | CD74 | E2F3 | HEPHL1 | MLIP | PHEX | SIX2 | TP53I11 |
| APAF1 | CD82 | E2F5 | HEYL | MLLT3 | PHLDB2 | SKAP2 | TRIM2 |
| APC2 | CD8A | EBF1 | HIF1A | MMP10 | PIEZO2 | SLA2 | TRIM27 |
| ARHGAP24 | CD97 | EFEMP1 | HLX | MMP16 | PIGH | SLAMF7 | TRIM31 |
| ARHGAP28 | CDC25A | EGFR | HMGA1 | MMP2 | PIK3CA | SLAMF8 | TRIM52 |
| ARID1A | CDC25B | EIF5A2 | HMGA2 | MMP9 | PKP1 | SLC24A4 | TSC1 |
| ARID3B | CDH1 | ELAVL1 | HMGB1 | MSH5 | PLAG1 | SLC2A3 | TTC14 |
| ARL5B | CDH2 | ELF5 | HNRNPC | MSN | PLAU | SLC31A1 | TUBB3 |
| ASXL3 | CDK1 | ELN | HOXA10 | MT-CO1 | PLD3 | SLC43A2 | TWIST1 |
| ATM | CDK12 | EML1 | HOXA13 | MT-ND2 | PLK1 | SLC4A4 | VAT1L |
| ATP5B | CDK2 | EPAS1 | HOXA9 | MTDH | PLS3 | SLC7A6 | VCAN |
| ATR | CDK4 | EPB41L3 | HOXB2 | MTFR1 | PMAIP1 | SMAD4 | VEGFA |
| AURKB | CDK6 | EPHA2 | ID1 | MTHFD1 | POSTN | SMAD7 | VEGFB |
| AXL | CDKN1A | EPHA4 | ID4 | MTSS1 | POTED | SMTNL2 | VEGFC |
| B3GNT5 | CDKN2A | ERBB2 | IGF1 | MUC1 | POU3F1 | SMURF1 | VIM |
| BAG5 | CEACAM1 | ERBB3 | IGF1R | MUC16 | PPP1R2 | SMYD1 | VTN |
| BAX | CHEK1 | ERBB4 | IGF2BP1 | MYC | PRDM16 | SNAI1 | WDR17 |
| BCL11B | CHEK2 | ESRRG | IGFBPL1 | MYCBP | PRKAA1 | SNAI2 | WNT1 |
| BCL2 | CHI3L1 | FAP | IL1A | MYCN | PROX1 | SOCS1 | WNT5A |
| BCL2L1 | CHST9 | FAR1 | IL2 | MYH9 | PTEN | SOCS2 | WSCD1 |
| BCR | CHSY1 | FBN1 | IL6R | MYO5A | PTGDR | SOD2 | XIAP |
| BIRC5 | COBLL1 | FBXO28 | INHBA | NEFL | PTHLH | SOS2 | XXYLT1 |
| BMF | COL11A1 | FCER1G | INSR | NEFM | PTPN12 | SOX11 | YAP1 |
| BMP3 | COL15A1 | FCRL1 | ITGA5 | NEUROD1 | PTPN4 | SOX12 | YOD1 |
| BMP4 | COL1A2 | FGF1 | ITGB1 | NEUROG1 | PTPRO | SOX4 | YY1 |
| BMP7 | COL3A1 | FGF2 | JAG1 | NF1 | PWWP2A | SOX9 | ZEB1 |
| BNIP3 | COL5A1 | FHL2 | JAG2 | NF2 | R3HDM4 | SPARC | ZEB2 |
| BRAF | COL5A2 | FN1 | JAKMIP2 | NFIX | RAB11FIP3 | SPHK1 | ZNF107 |
| BRCA1 | CPEB3 | FOSL2 | KCNA5 | NFKB1 | RAB22A | SPSB4 | ZNF138 |
| BRCA2 | CPNE3 | FOXA2 | KDR | NHS | RAB30 | SRC | ZNF181 |
| BTLA | CRISPLD2 | FOXD4L1 | KEAP1 | NOB1 | RAB5A | SREBF1 | ZNF346 |
| C10orf128 | CSF1R | FOXF2 | KIAA0101 | NOTCH1 | RACGAP1 | SREBF2 | ZNF423 |
| C11orf58 | CSMD3 | FOXM1 | KIAA0513 | NOTCH2 | RAD51 | SRSF1 | ZNF485 |
| C1orf105 | CTGF | FOXO3 | KLF12 | NOTCH3 | RAP1B | ST7L | ZNF521 |
| CACNA1C | CTNNB1 | FOXP1 | KLF15 | NREP | RARRES1 | STAT3 | ZNF697 |
| CACNG8 | CTSK | FUT4 | KLF4 | NRP1 | RASD1 | STK24 | ZNF706 |
| 20 to 29 Target Genes | 30 to 39 Target Genes | 40 to 59 Target Genes | |
|---|---|---|---|
| let-7a-5p | miR-3065-5p | let-7a-3p | miR-1275 |
| let-7b-5p | miR-30a-3p | miR-106a-5p | miR-15a-5p |
| let-7c-5p | miR-30d-3p | miR-106b-5p | miR-15b-5p |
| let-7d-5p | miR-30d-5p | miR-126-5p | miR-16-5p |
| let-7e-5p | miR-30e-3p | miR-129-5p | miR-195-5p |
| let-7f-5p | miR-30e-5p | miR-137 | miR-335-3p |
| let-7g-5p | miR-326 | miR-149-3p | miR-3607-3p |
| let-7i-5p | miR-330-5p | miR-17-5p | miR-373-5p |
| miR-105-5p | miR-340-5p | miR-181a-5p | miR-424-5p |
| miR-1253 | miR-34a-5p | miR-181b-5p | miR-497-5p |
| miR-124-3p | miR-363-3p | miR-181c-5p | miR-548a-5p |
| miR-128-3p | miR-377-3p | miR-181d-5p | miR-548b-5p |
| miR-1290 | miR-381-3p | miR-186-5p | miR-7-1-3p |
| miR-130a-5p | miR-486-3p | miR-200b-3p | miR-548d-3p |
| miR-130b-5p | miR-491-5p | miR-200c-3p | |
| miR-145-5p | miR-494-3p | miR-205-5p | |
| miR-182-5p | miR-506-3p | miR-20b-5p | |
| miR-185-5p | miR-511-5p | miR-30a-5p | |
| miR-1915-3p | miR-513a-3p | miR-30b-5p | |
| miR-200a-3p | miR-519a-3p | miR-30c-5p | |
| miR-204-5p | miR-519d-3p | miR-330-3p | |
| miR-20a-5p | miR-520b | miR-33a-3p | |
| miR-214-3p | miR-539-5p | miR-3688-3p | |
| miR-23a-3p | miR-551b-5p | miR-485-5p | |
| miR-23b-3p | miR-576-5p | miR-526b-3p | |
| miR-25-3p | miR-582-5p | miR-589-3p | |
| miR-26a-5p | miR-603 | miR-590-3p | |
| miR-26b-5p | miR-629-3p | miR-93-5p | |
| miR-27a-3p | miR-661 | miR-940 | |
| miR-27b-3p | miR-664a-3p | ||
| miR-29a-3p | miR-766-3p | ||
| miR-29b-1-5p | miR-92a-3p | ||
| miR-29b-2-5p | miR-92b-3p | ||
| miR-29b-3p | miR-96-5p | ||
| miR-29c-3p | miR-98-5p | ||
| miR-3065-3p | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thibonnier, M.; Ghosh, S. Strategy for Pre-Clinical Development of Active Targeting MicroRNA Oligonucleotide Therapeutics for Unmet Medical Needs. Int. J. Mol. Sci. 2023, 24, 7126. https://doi.org/10.3390/ijms24087126
Thibonnier M, Ghosh S. Strategy for Pre-Clinical Development of Active Targeting MicroRNA Oligonucleotide Therapeutics for Unmet Medical Needs. International Journal of Molecular Sciences. 2023; 24(8):7126. https://doi.org/10.3390/ijms24087126
Chicago/Turabian StyleThibonnier, Marc, and Sujoy Ghosh. 2023. "Strategy for Pre-Clinical Development of Active Targeting MicroRNA Oligonucleotide Therapeutics for Unmet Medical Needs" International Journal of Molecular Sciences 24, no. 8: 7126. https://doi.org/10.3390/ijms24087126
APA StyleThibonnier, M., & Ghosh, S. (2023). Strategy for Pre-Clinical Development of Active Targeting MicroRNA Oligonucleotide Therapeutics for Unmet Medical Needs. International Journal of Molecular Sciences, 24(8), 7126. https://doi.org/10.3390/ijms24087126