
Theaflavin Ameliorates Streptococcus suis-Induced Infection In Vitro and In Vivo

 ,
,
Abstract
:1. Introduction
2. Results
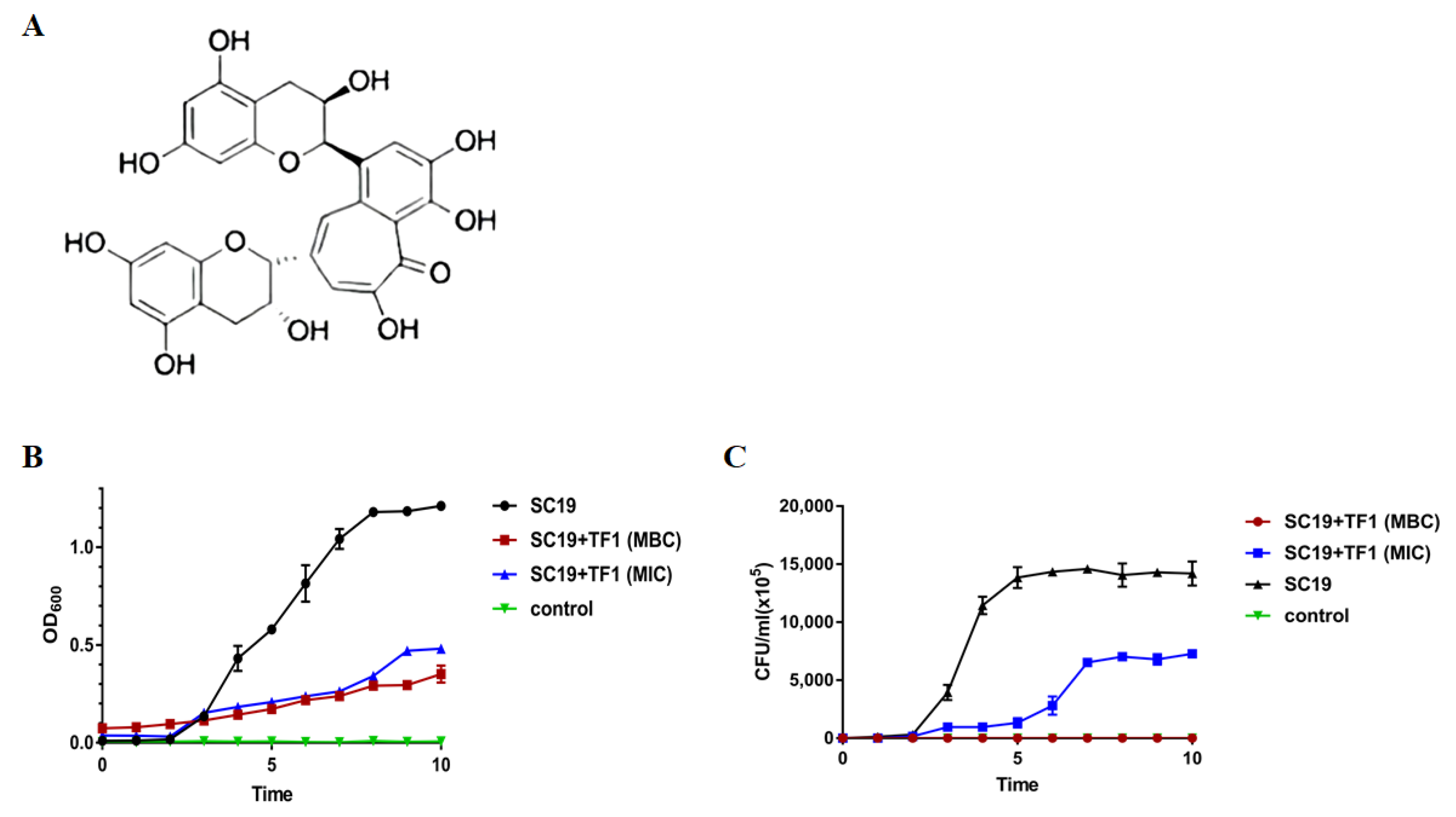
2.1. Inhibition of S. suis Growth by TF1
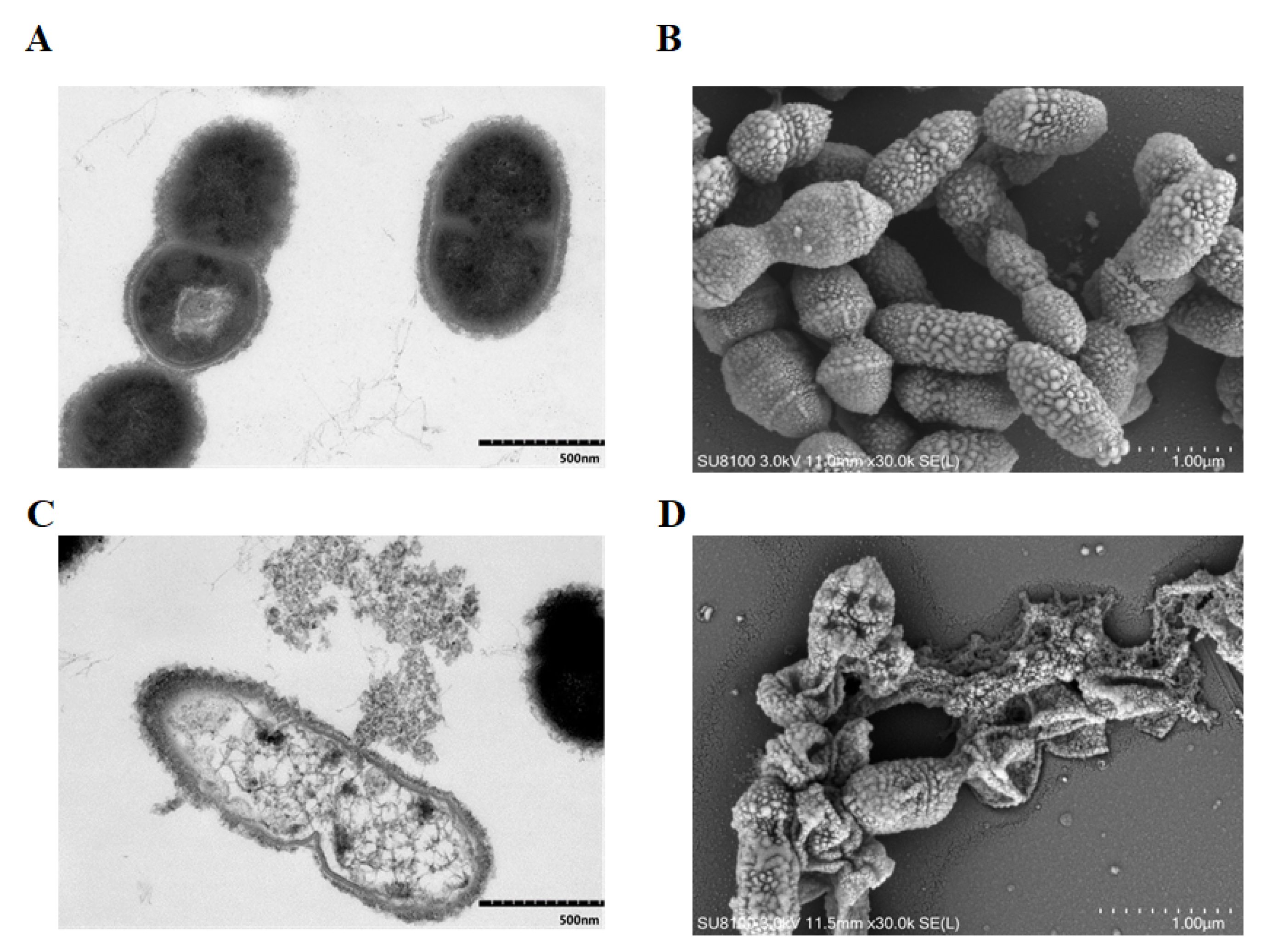
2.2. TF1 Causes Damage to the Internal and External Structure of S. suis
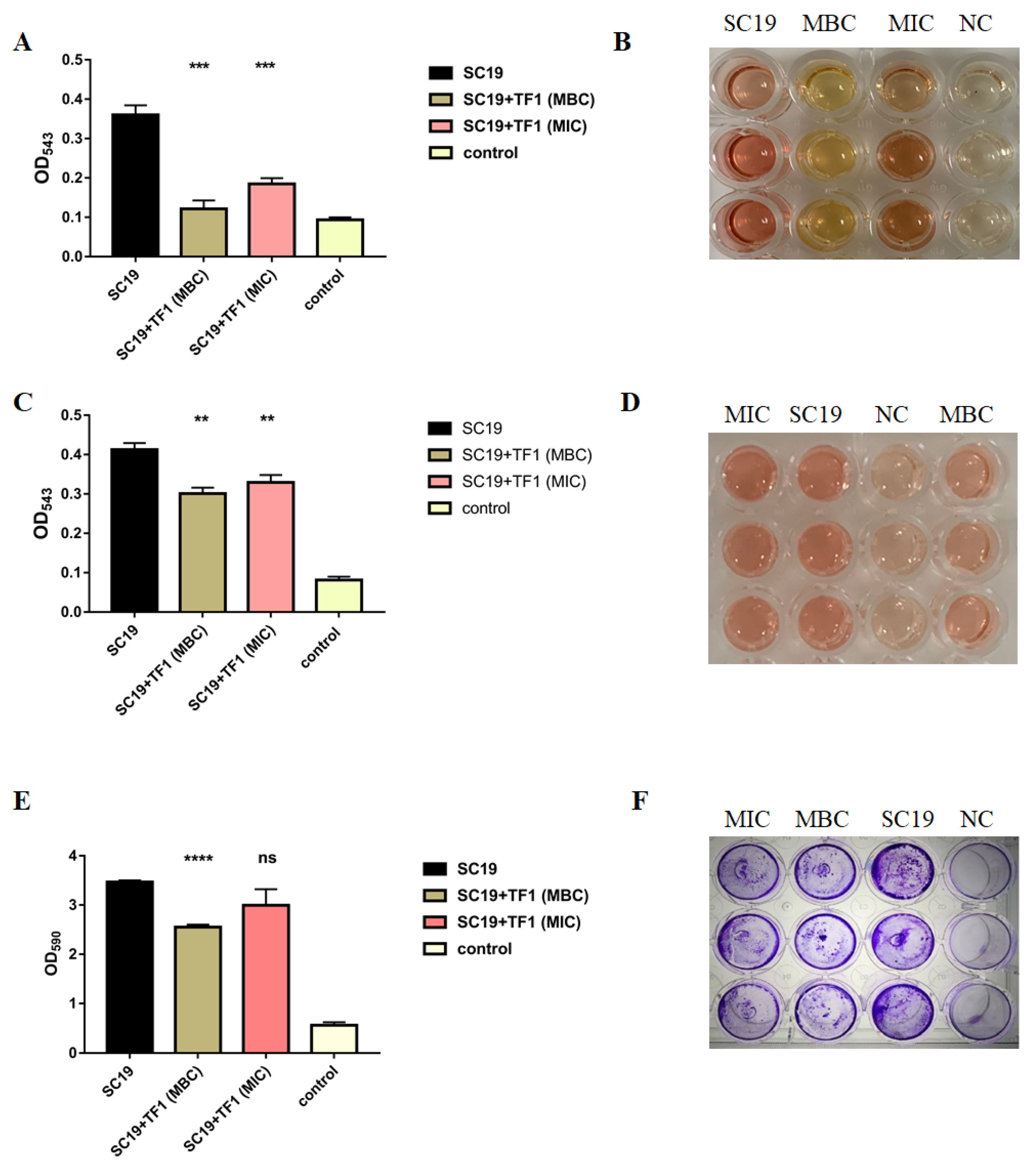
2.3. TF1 Inhibits Hemolytic Activity and Biofilm Information of S. suis
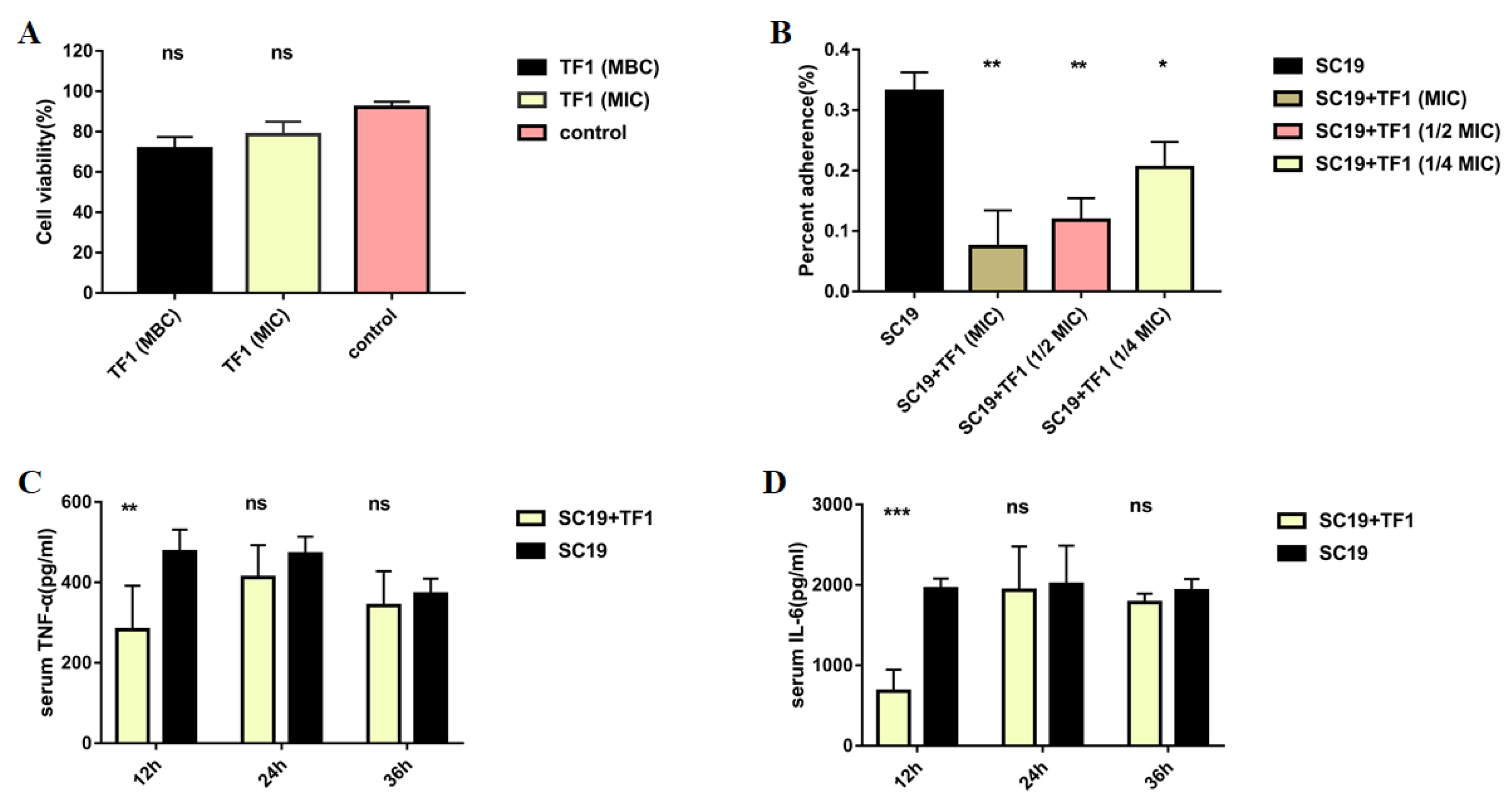
2.4. TF1 Decreased Adherent Activity of S. suis to Epithelial Cell Nptr
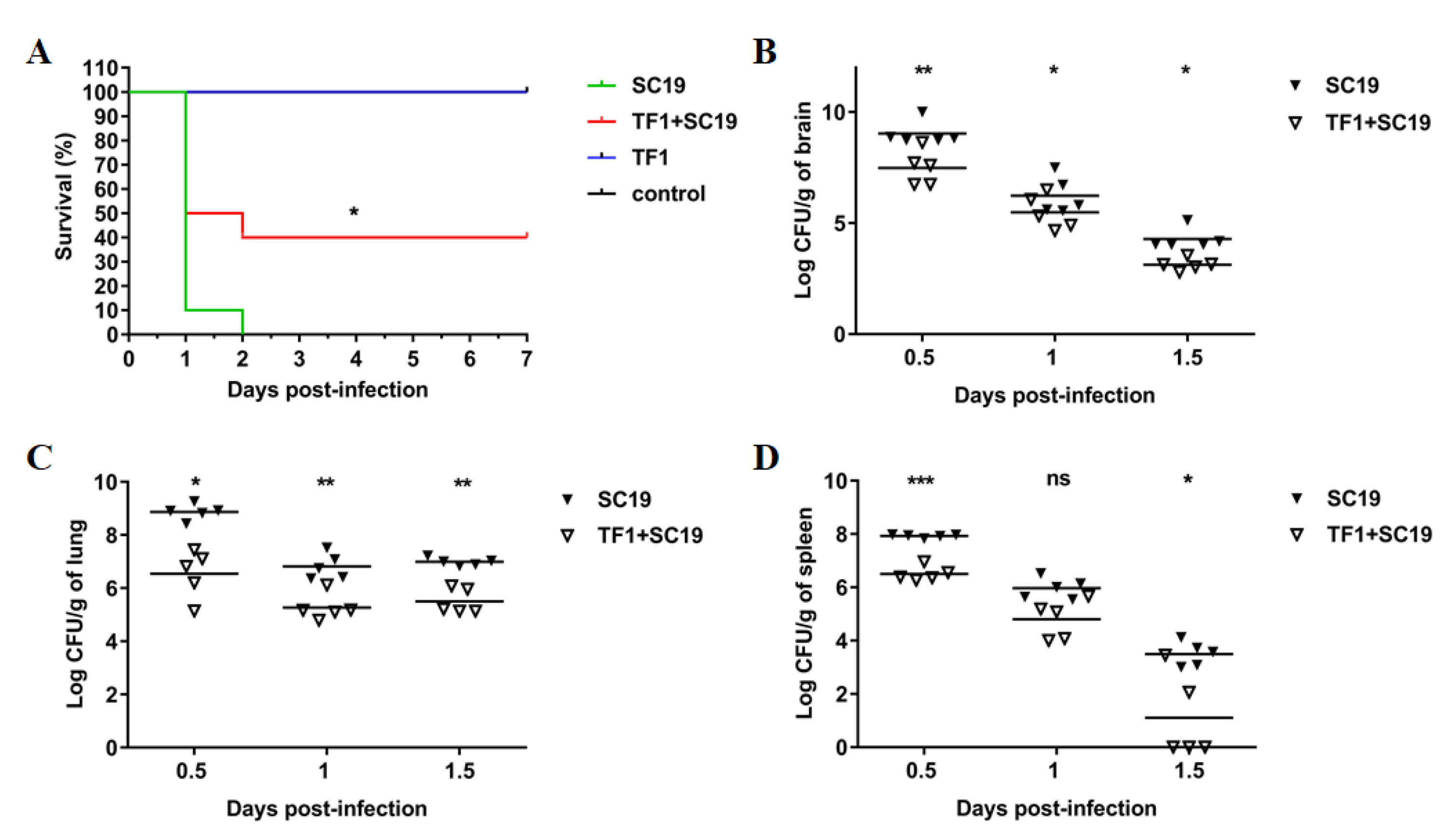
2.5. TF1 Protects Mice against SC19 and Decreased the Colonization Ability of SC19
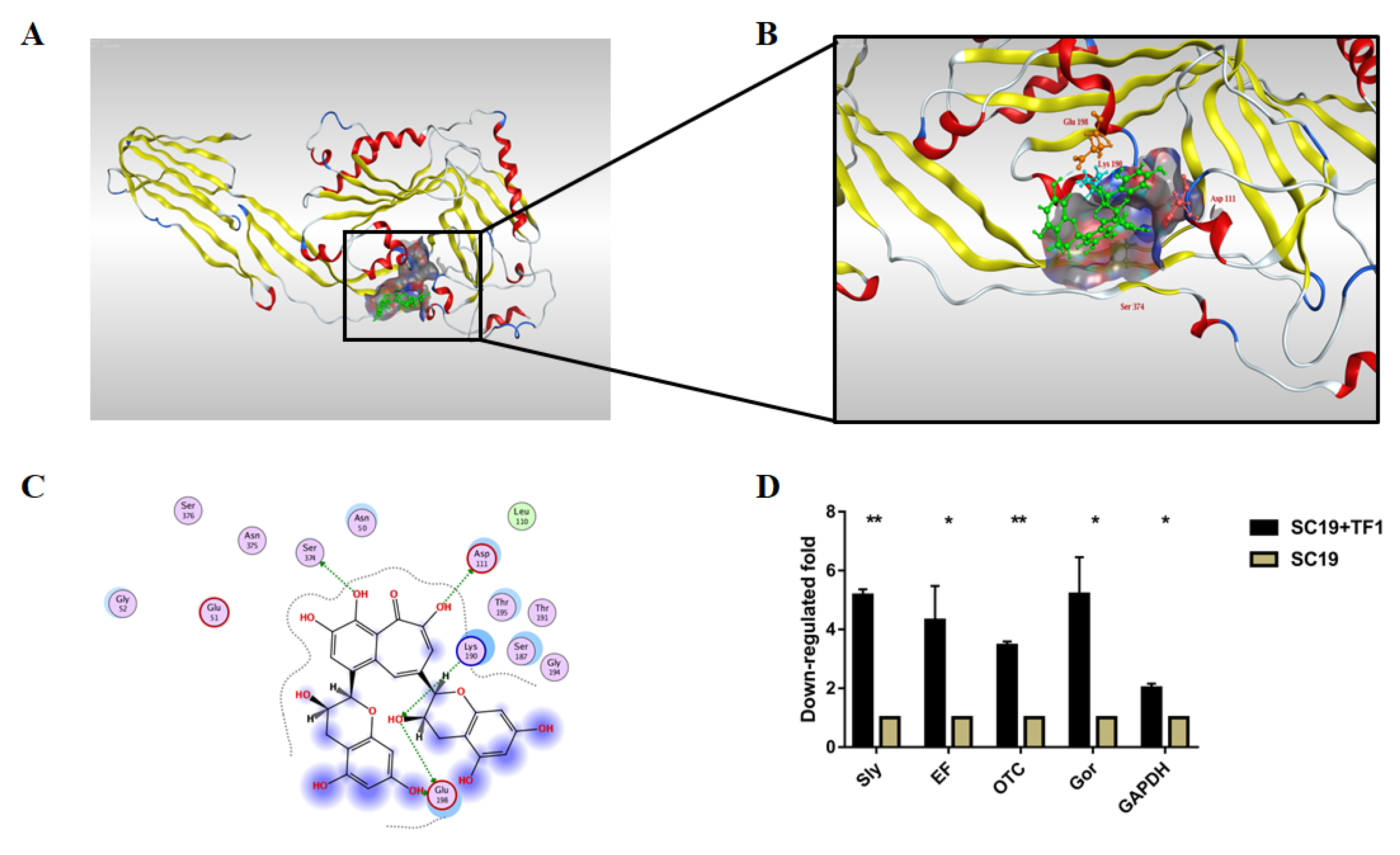
2.6. TF1 Binding Site on Sly Revealed by Molecular Docking
2.7. TF1 Inhibits the Expression of Virulence-Related Genes of SC19
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Growth Conditions and TF1
4.2. Susceptibility Testing and Time-Killing Curve of SC19 against TF1
4.3. Electron Microscope Observation of Bacterial Integrity
4.4. Effect of TF1 on Hemolytic Activity of Suilysin
4.5. Effect of TF1 on Biofilm Formation of SC19
4.6. Lactate Dehydrogenase (LDH) Cytotoxicity Assay of TF1
4.7. Test the Anti-Adhesion Activity of TF1 on SC19
4.8. Test the Anti-Infection Activity of TF1 on SC19
4.9. Molecular Docking Assay
4.10. qRT-PCR to Analyze the Cell Division and Virulent Related Genes at the mRNA Level
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jankun, J.; Selman, S.H.; Swiercz, R.; Skrzypczak-Jankun, E. Why drinking green tea could prevent cancer. Nature 1997, 387, 561. [Google Scholar] [CrossRef]
- Yussof, A.; Cammalleri, B.; Fayemiwo, O.; Lopez, S.; Chu, T. Antibacterial and Sporicidal Activity Evaluation of Theaflavin-3,3′-digallate. Int. J. Mol. Sci. 2022, 23, 2153. [Google Scholar] [CrossRef]
- Bag, S.; Mondal, A.; Majumder, A.; Banik, A. Tea and its phytochemicals: Hidden health benefits & modulation of signaling cascade by phytochemicals. Food Chem. 2021, 371, 131098. [Google Scholar]
- Tang, G.Y.; Meng, X.; Gan, R.Y.; Zhao, C.N.; Liu, Q.; Feng, Y.B.; Li, S.; Wei, X.L.; Atanasov, A.G.; Corke, H.; et al. Health Functions, and Related Molecular Mechanisms of Tea Components: An Update Review. Int. J. Mol. Sci. 2019, 20, 6196. [Google Scholar] [CrossRef]
- Oteiza, P.I.; Toyokuni, S. The new era for research on polyphenols and food factors. Arch. Biochem. Biophys. 2020, 696, 108678. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Qiao, X.; Gui, A.; Wang, S.; Liu, P.; Wang, X.; Teng, J.; Zheng, L.; Feng, L.; Han, H.; et al. Metabolomics Provides a Novel Interpretation of the Changes in Main Compounds during Black Tea Processing through Different Drying Methods. Molecules 2021, 26, 6739. [Google Scholar] [CrossRef]
- Tanaka, T.; Matsuo, Y. Production Mechanisms of Black Tea Polyphenols. Chem. Pharm. Bull. 2020, 68, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zhang, Y.; Wang, S. Inhibitory effects of theaflavin on planktonic and sessile bacterial cells of Streptococcus mutans in vitro. Shanghai Kou Qiang Yi Xue 2021, 30, 33–37. [Google Scholar] [PubMed]
- Sato, J.; Nakayama, M.; Tomita, A.; Sonoda, T.; Miyamoto, T. Difference in the antibacterial action of epigallocatechin gallate and theaflavin 3,3′-di-O-gallate on Bacillus coagulans. J. Appl. Microbiol. 2020, 129, 601–611. [Google Scholar] [CrossRef]
- Betts, J.W.; Hornsey, M.; Wareham, D.W.; La Ragione, R.M. In vitro and In vivo Activity of Theaflavin-Epicatechin Combinations versus Multidrug-Resistant Acinetobacter baumannii. Infect. Dis. Ther. 2017, 6, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Henika, P.R.; Levin, C.E.; Mandrell, R.E.; Kozukue, N. Antimicrobial activities of tea catechins and theaflavins and tea extracts against Bacillus cereus. J. Food Prot. 2006, 69, 354–361. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, H.; Wu, Z.; Wang, S.; Cao, M.; Hu, D.; Wang, C. Streptococcus suis infection: An emerging/reemerging challenge of bacterial infectious diseases? Virulence 2014, 5, 477–497. [Google Scholar] [CrossRef]
- Lun, Z.R.; Wang, Q.P.; Chen, X.G.; Li, A.X.; Zhu, X.Q. Streptococcus suis: An emerging zoonotic pathogen. Lancet Infect. Dis. 2007, 7, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Wang, C.; Feng, Y.; Yang, W.; Song, H.; Chen, Z.; Yu, H.; Pan, X.; Zhou, X.; Wang, H.; et al. Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med. 2006, 3, e151. [Google Scholar]
- Kerdsin, A.; Segura, M.; Fittipaldi, N.; Gottschalk, M. Sociocultural Factors Influencing Human Streptococcus suis Disease in Southeast Asia. Foods 2022, 11, 1190. [Google Scholar] [CrossRef] [PubMed]
- Lunha, K.; Chumpol, W.; Samngamnim, S.; Jiemsup, S.; Assavacheep, P.; Yongkiettrakul, S. Antimicrobial Susceptibility of Streptococcus suis Isolated from Diseased Pigs in Thailand, 2018–2020. Antibiotics 2022, 11, 410. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Lu, W.; Zhu, Y.; Wang, C.; Shi, L.; Li, X.; Wu, Z.; Wang, G.; Dong, W.; Tan, C.; et al. Auranofin Has Advantages over First-Line Drugs in the Treatment of Severe Streptococcus suis Infections. Antibiotics 2020, 10, 26. [Google Scholar] [CrossRef]
- Mascellino, M.T. Multi-Drug-Resistant Gram-Negative Microorganisms: Epidemiology, Treatment and Alternative Approach. Antibiotics 2022, 11, 678. [Google Scholar] [CrossRef]
- Mateo, E.M.; Jiménez, M. Silver Nanoparticle-Based Therapy: Can It Be Useful to Combat Multi-Drug Resistant Bacteria? Antibiotics 2022, 11, 1205. [Google Scholar] [CrossRef]
- Nedbalcova, K.; Kucharovicova, I.; Zouharova, M.; Matiaskova, K.; Kralova, N.; Brychta, M.; Simek, B.; Pecha, T.; Plodkova, H.; Matiasovic, J. Resistance of Streptococcus suis Isolates from the Czech Republic during 2018–2022. Antibiotics 2022, 11, 1214. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.F.; Tan, J.; Zeng, Y.B.; Li, H.Q.; Yang, Q.; Zhou, R. Antimicrobial resistance phenotypes and genotypes of Streptococcus suis isolated from clinically healthy pigs from 2017 to 2019 in Jiangxi Province, China. J. Appl. Microbiol. 2021, 130, 797–806. [Google Scholar] [CrossRef]
- Shen, X.; Niu, X.; Li, G.; Deng, X.; Wang, J. Amentoflavone Ameliorates Streptococcus suis-Induced Infection In Vitro and In Vivo. Appl. Environ. Microbiol. 2018, 84, e01804-18. [Google Scholar] [CrossRef]
- Zhang, Y.; Zong, B.; Wang, X.; Zhu, Y.; Hu, L.; Li, P.; Zhang, A.; Chen, H.; Liu, M.; Tan, C. Fisetin Lowers Streptococcus suis serotype 2 Pathogenicity in Mice by Inhibiting the Hemolytic Activity of Suilysin. Front. Microbiol. 2018, 9, 1723. [Google Scholar] [CrossRef]
- Niu, X.; Sun, L.; Wang, G.; Gao, Y.; Yang, Y.; Wang, X.; Wang, H. Investigation of the inhibition effect and mechanism of myricetin to Suilysin by molecular modeling. Sci. Rep. 2017, 7, 11748. [Google Scholar] [CrossRef]
- Gao, T.; Ye, F.; Yuan, F.; Liu, Z.; Liu, W.; Zhou, D.; Yang, K.; Guo, R.; Wang, N.; Zhang, T.; et al. Green tea polyphenols inhibit growth, pathogenicity and metabolomics profiles of Streptococcus suis. Microb. Pathog. 2022, 164, 105421. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Liu, Y.; Li, T.; Liu, X.; Hao, Z.; Ding, S.; Panichayupakaranant, P.; Zhu, K.; Shen, J. Plant Natural Flavonoids Against Multidrug Resistant Pathogens. Adv. Sci. 2021, 8, e2100749. [Google Scholar] [CrossRef]
- Gašparová, R. Natural Compounds with Oxepinochromene Scaffold. Structure, Source, Biological Activity and Synthesis. Chem. Biodivers. 2022, 19, e202200507. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Ge, X.; Xua, H.; Ma, K.; Zhang, W.; Zan, Y.; Efferth, T.; Xue, Z.; Hua, X. Phytochemicals with activity against methicillin-resistant Staphylococcus aureus. Phytomedicine 2022, 100, 154073. [Google Scholar] [CrossRef]
- Sato, J.; Tomita, A.; Sonoda, T.; Miyamoto, T. Theaflavin and its derivatives exert antibacterial action against Bacillus coagulans, through adsorption to cell surface phospholipids. J. Appl. Microbiol. 2022, 133, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef]
- Xu, L.; Lin, L.; Lu, X.; Xiao, P.; Liu, R.; Wu, M.; Jin, M.; Zhang, A. Acquiring high expression of suilysin enable non-epidemic Streptococccus suis to cause streptococcal toxic shock-like syndrome (STSLS) through NLRP3 inflammasome hyperactivation. Emerg. Microbes Infect. 2021, 10, 1309–1319. [Google Scholar] [CrossRef]
- Tenenbaum, T.; Asmat, T.M.; Seitz, M.; Schroten, H.; Schwerk, C. Biological activities of suilysin: Role in Streptococcus suis pathogenesis. Future Microbiol. 2016, 11, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lu, H.; Liu, M.; Wang, G.; Li, X.; Lu, W.; Dong, W.; Wang, X.; Chen, H.; Tan, C. Effective Antibacterial and Antihemolysin Activities of Ellipticine Hydrochloride against Streptococcus suis in a Mouse Model. Appl. Environ. Microbiol. 2021, 87, e03165-20. [Google Scholar] [CrossRef]
- Norton, P.M.; Rolph, C.; Ward, P.N.; Bentley, R.W.; Leigh, J.A. Epithelial invasion and cell lysis by virulent strains of Streptococcus, suis is enhanced by the presence of suilysin. FEMS Immunol. Med. Microbiol. 1999, 26, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Hou, Y.; Zhou, H.; Li, Y.; Xue, Z.; Xue, X.; Huang, G.; Huang, K.; He, X.; Xu, W. Hypolipidemic, anti-inflammatory, and anti-atherosclerotic effects of tea before and after microbial fermentation. Food Sci. Nutr. 2021, 9, 1160–1170. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, A.; Zhou, J.; Zhang, Y.; Zhou, F.; Chen, H.; Liu, Q.; Zhang, S.; Huang, J.; Liu, Z. Role and Mechanism of Theaflavins in Regulating Skeletal Muscle Inflammation. J. Agric. Food Chem. 2022, 70, 13233–13250. [Google Scholar] [CrossRef]
- Fu, G.; Wang, H.; Cai, Y.; Zhao, H.; Fu, W. Theaflavin Alleviates Inflammatory Response and Brain Injury Induced by Cerebral Hemorrhage via Inhibiting the Nuclear Transcription Factor Kappa β-Related Pathway in Rats [Retraction]. Drug Des. Devel. Ther. 2022, 16, 2341–2342. [Google Scholar] [CrossRef]
- Lun, S.; Perez-Casal, J.; Connor, W.; Willson, P.J. Role of suilysin in pathogenesis of Streptococcus suis capsular serotype 2. Microb. Pathog. 2003, 34, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, L.; Chen, H.; Zhou, R. Identification of Streptococcus suis genes preferentially expressed under iron starvation by selective capture of transcribed sequences. FEMS Microbiol. Lett. 2009, 292, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing. In CLSI Supplement M100, 27th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Morin, M.P.; Bedran, T.B.; Fournier-Larente, J.; Haas, B.; Azelmat, J.; Grenier, D. Green tea extract and its major constituent epigallocatechin-3-gallate inhibit growth and halitosis-related properties of Solobacterium moorei. BMC Complement Altern. Med. 2015, 15, 48. [Google Scholar] [CrossRef]
- Tan, M.F.; Hu, Q.; Hu, Z.; Zhang, C.Y.; Liu, W.Q.; Gao, T.; Zhang, L.S.; Yao, L.; Li, H.Q.; Zeng, Y.B.; et al. Streptococcus suis MsmK: Novel Cell Division Protein Interacting with FtsZ and Maintaining Cell Shape. mSphere 2021, 6, e00119-21. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Tan, M.; Liu, W.; Zhang, C.; Zhang, T.; Zheng, L.; Zhu, J.; Li, L.; Zhou, R. GidA, a tRNA Modification Enzyme, Contributes to the Growth, and Virulence of Streptococcus suis Serotype 2. Front. Cell. Infect. Microbiol. 2016, 6, 44. [Google Scholar] [CrossRef]
- Gao, T.; Ye, F.; Tan, Y.; Peng, M.; Yuan, F.; Liu, Z.; Zhou, D.; Yang, K.; Liu, W.; Guo, R.; et al. Metabolomics and proteomics analyses revealed mechanistic insights on the antimicrobial activity of epigallocatechin gallate against Streptococcus suis. Front. Cell. Infect. Microbiol. 2022, 12, 973282. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Yuan, F.; Liu, Z.; Liu, W.; Zhou, D.; Yang, K.; Duan, Z.; Guo, R.; Liang, W.; Hu, Q.; et al. MnmE, a Central tRNA-Modifying GTPase, Is Essential for the Growth, Pathogenicity, and Arginine Metabolism of Streptococcus suis Serotype 2. Front. Cell. Infect. Microbiol. 2019, 9, 173. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Xie, X.; Yi, P.; Kang, J.; Liao, J.; Li, W.; Li, F. Integrating Network Pharmacology with Molecular Docking to Unravel the Active Compounds and Potential Mechanism of Simiao Pill Treating Rheumatoid Arthritis. Evid. Based Complement Altern. Med. 2020, 2020, 5786053. [Google Scholar] [CrossRef]
- Mostafa, I.; Mohamed, N.H.; Mohamed, B.; Almeer, R.; Abulmeaty, M.M.A.; Bungau, S.G.; El-Shazly, A.M.; Yahya, G. In-silico screening of naturally derived phytochemicals against SARS-CoV Main protease. Environ. Sci. Pollut. Res. Int. 2022, 29, 26775–26791. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Li, J.; Fan, Q.; Mao, C.; Jin, M.; Liu, Y.; Sun, L.; Grenier, D.; Wang, Y. The otc gene of Streptococcus suis plays an important role in biofilm formation, adhesion, and virulence in a murine model. Vet. Microbiol. 2020, 251, 108925. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | MIC μg/mL | MBC μg/mL |
|---|---|---|
| Escherichia coli, K99 | 2048 | 4096 |
| Escherichia coli, K88 | 2048 | 4096 |
| Escherichia coli, 987P | 2048 | 4096 |
| Escherichia coli, ATCC25922 | 4096 | 4096 |
| Streptococcus suis, serotype 2 | 512 | 2048 |
| Streptococcus suis, serotype 7 | 512 | 2048 |
| S.equi subsp. Zooepidimicus | 512 | 2048 |
| Site | Size | PLB | Hyd | Side | Residues |
|---|---|---|---|---|---|
| 1 | 68 | 3.37 | 16 | 40 | 1:(ANS50 GLU51 GLY52 ASN82 ASN83 SER84 ASP86 ILE87 GLN107 LEU109 LEU110 ASP111 ASN112 SER187 LYS190 THR191 LYS192 PHE193 GLY194 THR195 ILE372 LEU373 SER374 ASN375 SER376) |
| 2 | 36 | 2.20 | 20 | 30 | 1:(TYR54 ILE55 TYR184 SER185 MET186 SER187 PHE208 APS209 VAL211 ASN212 GLU377 TYR378 ILE379 THR381) |
| 3 | 57 | 1.40 | 14 | 38 | 1:(MET182 PYR184 VAL211 GLU214 GLU215 LYS216 GLN217 SER281 SER282 ARG283 SER284 THR285 GLN286 VAL287 GLN288 ALA289 GLU380 THR381 THR382 SER383) |
| 4 | 21 | 1.16 | 10 | 17 | 1:(THR62 GLU63 LEU65 PHE70 GLU409 VEL410 SER411 TYR412 VEL419 GLU421 ASN445) |
| 5 | 57 | 0.96 | 25 | 36 | 1:(ILE97 TYR98 PRO99 ARG147 VAL150 ARG151 VAL154 ASN155 LEU158 TYR228 TYR270 GLY325 GLY354 VAL355 PRO356) |
| 6 | 11 | 0.83 | 8 | 27 | 1:(PHE70 VAL72 ARG74 THR384 HIS36 TYR412 GLU418 ARG447) |
| 7 | 43 | 0.7616 | 16 | 36 | 1:(ASN82 SER84 ASP86 ILE87 ALA88 ILE90 ASN112 GLM177 ASP179 LYS192 PHE193 6LY194 THR195 SER196 ASN22 L’S24 PHE275) |
| 8 | 27 | 0.59 | 13 | 23 | 1:(ARG272 MET274 ILE319 GLY322 ASP323 LYS340 ILE341 GLU344 GLY345 ALA346 TYR348 GLY349) |
| 9 | 51 | 0.38 | 21 | 37 | 1:(VAL89 ILE90 ASP91 ALA94 ALA95 ILE97 ASP111 ASN112 ASN113 LYS190 THR195 SER196 GLU198 LYS199 TYR359) |
| 10 | 22 | 0.21 | 10 | 14 | 1:(LEU128 ASN129 LEU130 PRO131 GLY132 LEU133 ALA134 ASN135GLY136 ASP137 TRP161) |
| 11 | 32 | 0.18 | 11 | 32 | 1:(ILE80 GLU180 THR181 MET182 TYR184GLN188 LYS192 GLN288 TYR378 GLU380) |
| 12 | 16 | 0.14 | 5 | 20 | 1:(ARG69 SER281 SER282 ARG283 TYR306 VAL385 ASN387) |
| 13 | 29 | −0.11 | 6 | 15 | 1:(HIS386 ASN387 SER388 SER389 ILE442 PRO443 GLY444 ASN445 ALA446 ARG447 LEU474 VAL475 GLV476) |
| 14 | 19 | −0.12 | 9 | 23 | 1:(GLU66 ASN67 ARG69 VAL71 GLY214 GL0215 LYS216 SER281 ARG283 VAL385) |
| 15 | 11 | −0.14 | 6 | 18 | 1:(TYR98 LEU102 LEU116 ILE117SER118 ILE119 ARG121 PRO145 THR146 ARG147) |
| 16 | 55 | −0.16 | 19 | 32 | 1:(GLY170 ASN171 THR172 GLN173 ALA174 LEU176 PHE223 GLN225 TYR2271LE295 LYS296 GLV297 ILE342 GLU343 ALA345 ARG347) |
| 17 | 21 | −0.19 | 18 | 21 | 1:(THR285GLN286 ALA289 ALA290 ILE300 ALA304 GLU305 TYR306 GLN307 ILE309 LEU310) |
| 18 | 18 | −0.26 | 9 | 12 | 1:(ARG447ASN448 LEU 449 ASP 471 LEU 472 PRO473 LEU 474) |
| 19 | 28 | −0.44 | 19 | 23 | 1:(ACE31 ASP32 ILE33 TYR36 VAL248 GLU249 LEU251 LYS252) |
| 20 | 26 | −0.52 | 6 | 19 | 1:(ASN67 GLY68 ARG69 ASN387SER388 SER389 ALA390 GLN441 GLY476 GLN477 LEU498) |
| 21 | 9 | −0.64 | 5 | 9 | 1:(GLU46 ILE47 LEU48 THR49 ASP106 GLN107 LEU110) |
| 22 | 17 | −0.64 | 10 | 20 | 1:(GLU175 LEU176GLN177 LYS224 ILE226 THR229 SER269 SER358) |
| 23 | 10 | −0.65 | 9 | 14 | 1:(ALA293 ALA294 ILE295 GLY297 ASP299 ILE300 SER301 LYs339 ILE342 GLU343) |
| Mol | S | rmsd_refine | E_conf | E_place | E_score1 | E_refine | |
|---|---|---|---|---|---|---|---|
| 1 | 135403798 | −6.3573 | 1.7533 | 101.7029 | −88.9436 | −13.1194 | −26.4553 |
| 2 | 135403798 | −6.2809 | 2.1405 | 101.6749 | −124.5373 | −12.5693 | −21.9376 |
| 3 | 135403798 | −6.1305 | 2.0692 | 103.7832 | −92.5910 | −12.0843 | −19.3180 |
| 4 | 135403798 | −6.1281 | 2.9734 | 101.0886 | −84.2101 | −13.3046 | −23.3519 |
| 5 | 135403798 | −6.0669 | 4.2344 | 102.9650 | −85.8538 | −15.1033 | −20.1235 |
| Primers | Primers Sequence (5′-3′) | Reference or Source |
|---|---|---|
| 16S rRNA-F | CATCCATAACAGCCATACCAG | [48] |
| 16S rRNA-R | TAAACCACATGCTCCACCGC | |
| gapdh-F | GCTGAAGAAGTAAACGCTGCT | [48] |
| gapdh-R | GTCGCATCAAACAATGAACC | |
| sly-F | AGTCAGTTTGGCACTCGTAGG | This work |
| sly-R | TTGTGGCTCGTAAGTCAAGC | |
| ef-F | TCCAATCACAGATCCAGATAGCG | [48] |
| ef-R | CTGACCCATTTGGACCATCTAAG | |
| gor-F | GTTCACGCGCATCCTACG | [48] |
| gor-R | TACCAGGAATAGCAGGGAC | |
| otc-F | TTGCCCTCTTGAAGCCATACCA | This work |
| otc-R | TTCCATTTCTTCTACGCCGAAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, T.; Tan, Y.; Wang, Y.; Yuan, F.; Liu, Z.; Yang, K.; Liu, W.; Guo, R.; Li, C.; Tian, Y.; et al. Theaflavin Ameliorates Streptococcus suis-Induced Infection In Vitro and In Vivo. Int. J. Mol. Sci. 2023, 24, 7442. https://doi.org/10.3390/ijms24087442
Gao T, Tan Y, Wang Y, Yuan F, Liu Z, Yang K, Liu W, Guo R, Li C, Tian Y, et al. Theaflavin Ameliorates Streptococcus suis-Induced Infection In Vitro and In Vivo. International Journal of Molecular Sciences. 2023; 24(8):7442. https://doi.org/10.3390/ijms24087442
Chicago/Turabian StyleGao, Ting, Yiqing Tan, Yanjun Wang, Fangyan Yuan, Zewen Liu, Keli Yang, Wei Liu, Rui Guo, Chang Li, Yongxiang Tian, and et al. 2023. "Theaflavin Ameliorates Streptococcus suis-Induced Infection In Vitro and In Vivo" International Journal of Molecular Sciences 24, no. 8: 7442. https://doi.org/10.3390/ijms24087442
APA StyleGao, T., Tan, Y., Wang, Y., Yuan, F., Liu, Z., Yang, K., Liu, W., Guo, R., Li, C., Tian, Y., & Zhou, D. (2023). Theaflavin Ameliorates Streptococcus suis-Induced Infection In Vitro and In Vivo. International Journal of Molecular Sciences, 24(8), 7442. https://doi.org/10.3390/ijms24087442