
Melatonin and Abiotic Stress Tolerance in Crop Plants


Abstract
:1. Introduction
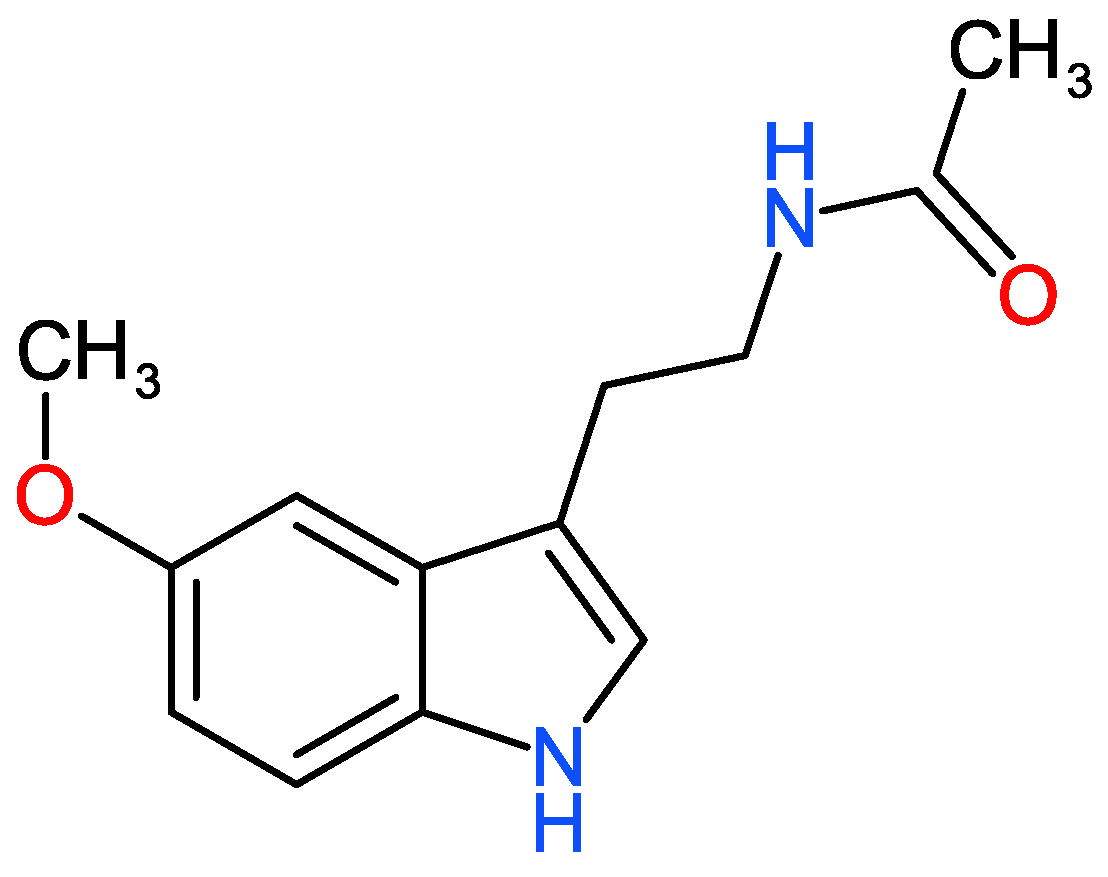
2. What Is Melatonin?
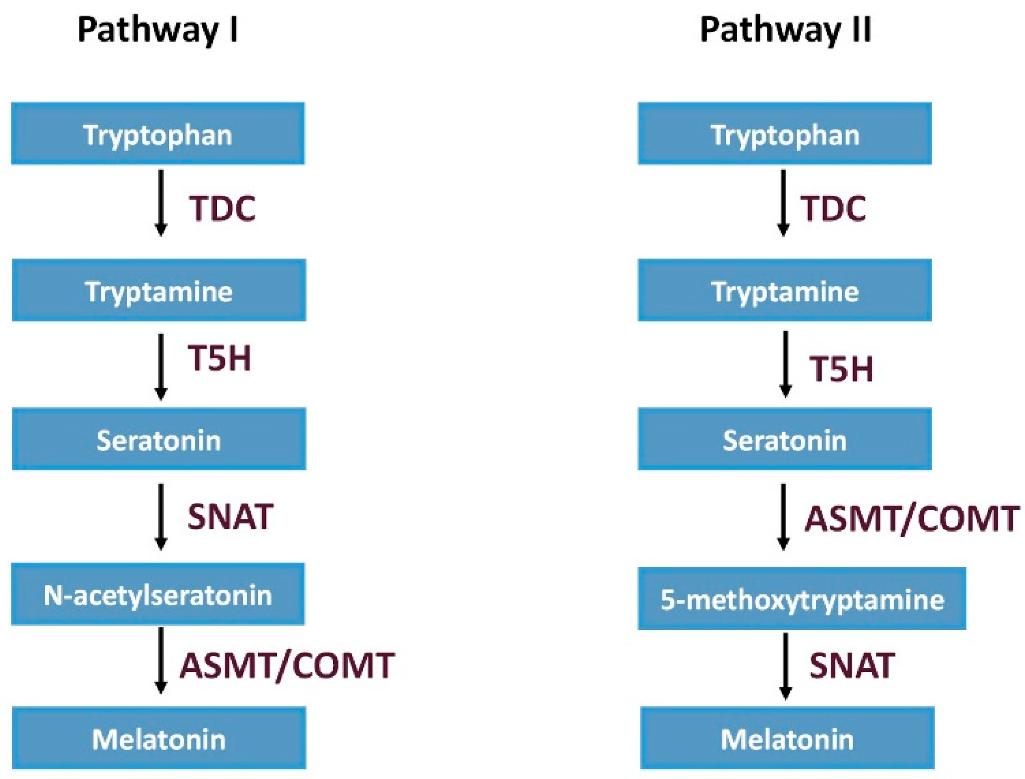
2.1. Melatonin Biosynthesis in Plants
2.2. Functions of Melatonin
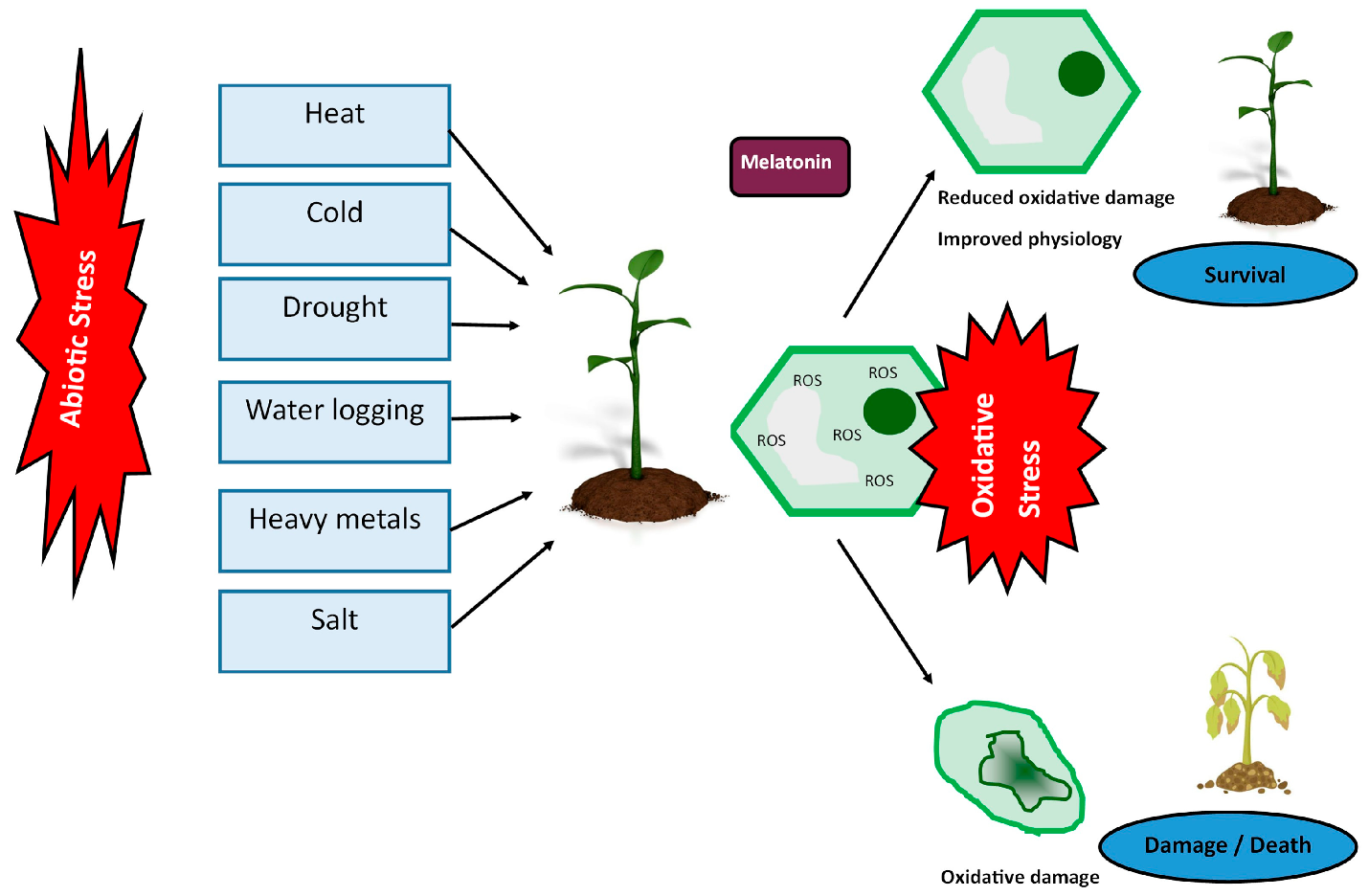
2.3. Endogenous Melatonin in Abiotic Stress
3. Exogenous Melatonin Application Enhances Abiotic Stress Tolerance
3.1. Melatonin Pre-Treatment for High-Temperature Stress
3.2. Melatonin Pre-Treatment Reduces Heat-Induced Damage during Reproduction
3.3. Melatonin Pre-Treatment for Water Stress
3.4. Melatonin for Heavy Metal Stress
3.5. Melatonin Pre-Treatment for Combined Stresses
4. Role of Melatonin in Cellular Function
4.1. ER Stress Response and Protein Homeostasis
4.2. Melatonin and Autophagy
4.3. Interaction of Melatonin with HSFs and HSPs
5. Melatonin Regulates Phytohormones under Abiotic Stress
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statements
Conflicts of Interest
References
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2021, 41, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. Biological Parts for Engineering Abiotic Stress Tolerance in Plants. BioDes. Res. 2022, 2022, 9819314. [Google Scholar] [CrossRef]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Gu, Q.; Xiao, Q.; Chen, Z.; Han, Y. Crosstalk between Melatonin and Reactive Oxygen Species in Plant Abiotic Stress Responses: An Update. Int. J. Mol. Sci. 2022, 23, 5666. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Nazir, F.; Fariduddin, Q.; Alam Khan, T. Hydrogen peroxide as a signalling molecule in plants and its crosstalk with other plant growth regulators under heavy metal stress. Chemosphere 2020, 252, 126486. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: The role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef]
- Gonzalez-Hunt, C.P.; Wadhwa, M.; Sanders, L.H. DNA damage by oxidative stress: Measurement strategies for two genomes. Curr. Opin. Toxicol. 2018, 7, 87–94. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS Homeostasis in Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef]
- Gechev, T.; Petrov, V. Reactive Oxygen Species and Abiotic Stress in Plants. Int. J. Mol. Sci. 2020, 21, 7433. [Google Scholar] [CrossRef]
- Chaki, M.; Begara-Morales, J.C.; Barroso, J.B. Oxidative Stress in Plants. Antioxidants 2020, 9, 481. [Google Scholar] [CrossRef]
- Lohani, N.; Golicz, A.A.; Singh, M.B.; Bhalla, P.L. Genome-wide analysis of the Hsf gene family in Brassica oleracea and a comparative analysis of the Hsf gene family in B. oleracea, B. rapa and B. napus. Funct. Integr. Genom. 2019, 19, 515–531. [Google Scholar] [CrossRef]
- Pardo-Hernández, M.; López-Delacalle, M.; Rivero, R. ROS and NO Regulation by Melatonin under Abiotic Stress in Plants. Antioxidants 2020, 9, 1078. [Google Scholar] [CrossRef]
- Yang, X.; Ren, J.; Li, J.; Lin, X.; Xia, X.; Yan, W.; Zhang, Y.; Deng, X.; Ke, Q. Meta-analysis of the effect of melatonin application on abiotic stress tolerance in plants. Plant Biotechnol. Rep. 2022, 17, 39–52. [Google Scholar] [CrossRef]
- Kołodziejczyk, I.; Posmyk, M.M. Melatonin—A new plant biostimulator? J. Elementol. 2016, 21, 1187–1198. [Google Scholar] [CrossRef]
- Qin, B.; Zou, J.; Cao, L.; Wang, M.; Zhang, Y.-X. Melatonin regulates material transport to reduce carbon emissions and increase yield under different nitrogen in rice. Agric. Ecosyst. Environ. 2023, 342, 108235. [Google Scholar] [CrossRef]
- Zou, J.; Jin, X.; Zhang, Y.; Ren, C.; Zhang, M.; Wang, M. Effects of melatonin on photosynthesis and soybean seed growth during grain filling under drought stress. Photosynthetica 2019, 57, 512–520. [Google Scholar] [CrossRef]
- Wei, W.; Li, Q.-T.; Chu, Y.-N.; Reiter, R.J.; Yu, X.-M.; Zhu, D.-H.; Zhang, W.-K.; Ma, B.; Lin, Q.; Zhang, J.-S.; et al. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.S.; Rasheed, M.; Saleem, M.H.; Ahmed, Z.I.; Hafeez, A.; Jilani, G.; Alamri, S.; Hashem, M.; Ali, S. Salt tolerance in maize with melatonin priming to achieve sustainability in yield on salt affected soils. Pak. J. Bot. 2022, 55, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Kamran, M.; Zhou, X.; Ahmad, I.; Meng, X.; Javed, T.; Iqbal, A.; Wang, G.; Su, W.; Wu, X.; et al. Melatonin improves the seed filling rate and endogenous hormonal mechanism in grains of summer maize. Physiol. Plant. 2021, 172, 1059–1072. [Google Scholar] [CrossRef]
- Lerner, A.B.; Case, J.D.; Takahashi, Y. Isolation of Melatonin and 5-Methoxyindole-3-acetic Acid from Bovine Pineal Glands. J. Biol. Chem. 1960, 235, 1992–1997. [Google Scholar] [CrossRef] [PubMed]
- Janas, K.M.; Posmyk, M.M. Melatonin, an underestimated natural substance with great potential for agricultural application. Acta Physiol. Plant. 2013, 35, 3285–3292. [Google Scholar] [CrossRef]
- Badria, F.A. Melatonin, Serotonin, and Tryptamine in Some Egyptian Food and Medicinal Plants. J. Med. Food 2002, 5, 153–157. [Google Scholar] [CrossRef]
- Blask, D.E.; Dauchy, R.T.; Sauer, L.A.; Krause, J.A. Melatonin uptake and growth prevention in rat hepatoma 7288CTC in response to dietary melatonin: Melatonin receptor-mediated inhibition of tumor linoleic acid metabolism to the growth signaling molecule 13-hydroxyoctadecadienoic acid and the potential role of phytomelatonin. Carcinogenesis 2004, 25, 951–960. [Google Scholar] [CrossRef]
- Shibaeva, T.G.; Markovskaya, E.F.; Mamaev, A.V. Phytomelatonin: A Review. Biol. Bull. Rev. 2018, 8, 375–388. [Google Scholar] [CrossRef]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front. Endocrinol. 2019, 10, 249. [Google Scholar] [CrossRef]
- Martin, W.; Kowallik, K. Annotated English translation of Mereschkowsky’s 1905 paper ‘Über Natur und Ursprung der Chromatophoren imPflanzenreiche’. Eur. J. Phycol. 1999, 34, 287–295. [Google Scholar] [CrossRef]
- Tan, D.-X.; Manchester, L.C.; Liu, X.; Rosales-Corral, S.A.; Acuna-Castroviejo, D.; Reiter, R.J. Mitochondria and chloroplasts as the original sites of melatonin synthesis: A hypothesis related to melatonin’s primary function and evolution in eukaryotes. J. Pineal Res. 2013, 54, 127–138. [Google Scholar] [CrossRef]
- Fan, J.; Xie, Y.; Zhang, Z.; Chen, L. Melatonin: A Multifunctional Factor in Plants. Int. J. Mol. Sci. 2018, 19, 1528. [Google Scholar] [CrossRef]
- Mannino, G.; Pernici, C.; Serio, G.; Gentile, C.; Bertea, C.M. Melatonin and Phytomelatonin: Chemistry, Biosynthesis, Metabolism, Distribution and Bioactivity in Plants and Animals—An Overview. Int. J. Mol. Sci. 2021, 22, 9996. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin in plants and other phototrophs: Advances and gaps concerning the diversity of functions. J. Exp. Bot. 2015, 66, 627–646. [Google Scholar] [CrossRef]
- Ye, T.; Yin, X.; Yu, L.; Zheng, S.-J.; Cai, W.-J.; Wu, Y.; Feng, Y.-Q. Metabolic analysis of the melatonin biosynthesis pathway using chemical labeling coupled with liquid chromatography-mass spectrometry. J. Pineal Res. 2019, 66, e12531. [Google Scholar] [CrossRef]
- Back, K.; Tan, D.-X.; Reiter, R.J. Melatonin biosynthesis in plants: Multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef]
- Hernández-Rui, M.B.A.A.J. The Physiological Function of Melatonin in Plants. Plant Signal. Behav. 2006, 1, 7. [Google Scholar]
- Wei, J.; Li, D.-X.; Zhang, J.-R.; Shan, C.; Rengel, Z.; Song, Z.-B.; Chen, Q. Phytomelatonin receptor PMTR1-mediated signaling regulates stomatal closure in Arabidopsis thaliana. J. Pineal Res. 2018, 65, e12500. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin as a regulatory hub of plant hormone levels and action in stress situations. Plant Biol. 2021, 23, 7–19. [Google Scholar] [CrossRef]
- Lee, H.Y.; Hwang, O.J.; Back, K. Phytomelatonin as a signaling molecule for protein quality control via chaperone, autophagy, and ubiquitin–proteasome systems in plants. J. Exp. Bot. 2022, 73, 5863–5873. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.-D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef]
- Khan, A.; Numan, M.; Khan, A.L.; Lee, I.-J.; Imran, M.; Asaf, S.; Al-Harrasi, A. Melatonin: Awakening the Defense Mechanisms during Plant Oxidative Stress. Plants 2020, 9, 407. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.A.; Shahid, R.; Ren, M.; Mora-Poblete, F.; Arnao, M.B.; Naz, S.; Anwar, M.; Shahid, S.; Shakoor, A.; Sohail, H.; et al. Phytomelatonin: An overview of the importance and mediating functions of melatonin against environmental stresses. Physiol. Plant. 2021, 172, 820–846. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Nawaz, G.; Cao, Q.; Xu, T. Melatonin is a potential target for improving horticultural crop resistance to abiotic stress. Sci. Hortic. 2021, 291, 110560. [Google Scholar] [CrossRef]
- Chang, J.; Guo, Y.; Yan, J.; Zhang, Z.; Yuan, L.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X.; et al. The role of watermelon caffeic acid O-methyltransferase (ClCOMT1) in melatonin biosynthesis and abiotic stress tolerance. Hortic. Res. 2021, 8, 210. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-D.; Sun, X.-S.; Liu, L.; Shi, H.-D.; Chen, S.-Y.; Zhao, D.-K. Overexpression of the Melatonin Synthesis-Related Gene SlCOMT1 Improves the Resistance of Tomato to Salt Stress. Molecules 2019, 24, 1514. [Google Scholar] [CrossRef]
- Yang, W.-J.; Du, Y.-T.; Zhou, Y.-B.; Chen, J.; Xu, Z.-S.; Ma, Y.-Z.; Chen, M.; Min, D.-H. Overexpression of TaCOMT Improves Melatonin Production and Enhances Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2019, 20, 652. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Xie, Q.; Liu, Y.; Lv, H.; Bai, R.; Ma, R.; Li, X.; Zhang, X.; Guo, Y.-D.; et al. SlSNAT Interacts with HSP40, a Molecular Chaperone, to Regulate Melatonin Biosynthesis and Promote Thermotolerance in Tomato. Plant Cell Physiol. 2020, 61, 909–921. [Google Scholar] [CrossRef]
- Gu, Q.; Chen, Z.; Yu, X.; Cui, W.; Pan, J.; Zhao, G.; Xu, S.; Wang, R.; Shen, W. Melatonin confers plant tolerance against cadmium stress via the decrease of cadmium accumulation and reestablishment of microRNA-mediated redox homeostasis. Plant Sci. 2017, 261, 28–37. [Google Scholar] [CrossRef]
- Wu, Y.; Fan, X.; Zhang, Y.; Jiang, J.; Sun, L.; Rahman, F.U.; Liu, C. VvSNAT1 overexpression enhances melatonin production and salt tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2021, 166, 485–494. [Google Scholar] [CrossRef]
- Yao, J.-W.; Ma, Z.; Ma, Y.-Q.; Zhu, Y.; Lei, M.-Q.; Hao, C.-Y.; Chen, L.-Y.; Xu, Z.-Q.; Huang, X. Role of melatonin in UV-B signaling pathway and UV-B stress resistance in Arabidopsis thaliana. Plant Cell Environ. 2021, 44, 114–129. [Google Scholar] [CrossRef]
- Qi, C.; Zhang, H.; Liu, Y.; Wang, X.; Dong, D.; Yuan, X.; Li, X.; Zhang, X.; Li, X.; Zhang, N.; et al. CsSNAT positively regulates salt tolerance and growth of cucumber by promoting melatonin biosynthesis. Environ. Exp. Bot. 2020, 175, 104036. [Google Scholar] [CrossRef]
- Xu, W.; Cai, S.; Zhang, Y.; Wang, Y.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Reiter, R.J.; et al. Melatonin enhances thermotolerance by promoting cellular protein protection in tomato plants. J. Pineal Res. 2016, 61, 457–469. [Google Scholar] [CrossRef]
- Zhuang, W.; Liu, T.; Shu, X.; Wang, H.; Wang, Z.; Wang, T.; Zhang, F.; Qu, S. Overexpression of MzASMT 1, a Gene from Malus zumi Mats, Enhances Salt Tolerance in Transgenic Tobacco. Front. Plant Sci. 2020, 11, 561903. [Google Scholar] [CrossRef]
- Zuo, B.; Zheng, X.; He, P.; Wang, L.; Lei, Q.; Feng, C.; Zhou, J.; Li, Q.; Han, Z.; Kong, J. Overexpression of MzASMT improves melatonin production and enhances drought tolerance in transgenic Arabidopsis thaliana plants. J. Pineal Res. 2014, 57, 408–417. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Mosaad, I.S.M.; Al-Ghamdi, A.A.; Abbasi, A.M.; Zhou, X.-B. Melatonin Application Alleviates Stress-Induced Photosynthetic Inhibition and Oxidative Damage by Regulating Antioxidant Defense System of Maize: A Meta-Analysis. Antioxidants 2022, 11, 512. [Google Scholar] [CrossRef]
- Manafi, H.; Baninasab, B.; Gholami, M.; Talebi, M.; Khanizadeh, S. Exogenous Melatonin Alleviates Heat-Induced Oxidative Damage in Strawberry (Fragaria × ananassa Duch. cv. Ventana) Plant. J. Plant Growth Regul. 2021, 41, 52–64. [Google Scholar] [CrossRef]
- Buttar, Z.; Wu, S.; Arnao, M.; Wang, C.; Ullah, I.; Wang, C. Melatonin Suppressed the Heat Stress-Induced Damage in Wheat Seedlings by Modulating the Antioxidant Machinery. Plants 2020, 9, 809. [Google Scholar] [CrossRef]
- Jahan, M.S.; Guo, S.; Baloch, A.R.; Sun, J.; Shu, S.; Wang, Y.; Ahammed, G.J.; Kabir, K.; Roy, R. Melatonin alleviates nickel phytotoxicity by improving photosynthesis, secondary metabolism and oxidative stress tolerance in tomato seedlings. Ecotoxicol. Environ. Saf. 2020, 197, 110593. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, X.; Zhang, Z.; Liu, N.; Li, D.; Hu, L. Melatonin Improved Waterlogging Tolerance in Alfalfa (Medicago sativa) by Reprogramming Polyamine and Ethylene Metabolism. Front. Plant Sci. 2019, 10, 44. [Google Scholar] [CrossRef]
- Rehaman, A.; Mishra, A.K.; Ferdose, A.; Per, T.S.; Hanief, M.; Jan, A.T.; Asgher, M. Melatonin in Plant Defense against Abiotic Stress. Forests 2021, 12, 1404. [Google Scholar] [CrossRef]
- Chen, Q.; Qi, W.-B.; Reiter, R.J.; Wei, W.; Wang, B.-M. Exogenously applied melatonin stimulates root growth and raises endogenous indoleacetic acid in roots of etiolated seedlings of Brassica juncea. J. Plant Physiol. 2009, 166, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Posmyk, M.M.; Kuran, H.; Marciniak, K.; Janas, K.M. Presowing seed treatment with melatonin protects red cabbage seedlings against toxic copper ion concentrations. J. Pineal Res. 2008, 45, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Değer, Ö.; Szafrańska, K.; Köklü, Ş.; Karaca, A.; Yakupoğlu, G.; Kocaçinar, F. Melatonin effects in enhancing chilling stress tolerance of pepper. Sci. Hortic. 2021, 289, 110434. [Google Scholar] [CrossRef]
- Li, X.; Wei, J.-P.; Scott, E.R.; Liu, J.-W.; Guo, S.; Li, Y.; Zhang, L.; Han, W.-Y. Exogenous Melatonin Alleviates Cold Stress by Promoting Antioxidant Defense and Redox Homeostasis in Camellia sinensis L. Molecules 2018, 23, 165. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.; Zhao, Y.; He, H.; Xi, Q.; Fu, J.; Zhao, Y. Exogenous melatonin improves growth in hulless barley seedlings under cold stress by influencing the expression rhythms of circadian clock genes. PeerJ 2021, 9, e10740. [Google Scholar] [CrossRef]
- Dai, L.; Li, J.; Harmens, H.; Zheng, X.; Zhang, C. Melatonin enhances drought resistance by regulating leaf stomatal behaviour, root growth and catalase activity in two contrasting rapeseed (Brassica napus L.) genotypes. Plant Physiol. Biochem. 2020, 149, 86–95. [Google Scholar] [CrossRef]
- Ibrahim, M.F.M.; Abd Elbar, O.H.; Farag, R.; Hikal, M.; El-Kelish, A.; El-Yazied, A.A.; Alkahtani, J.; Abd El-Gawad, H.G. Melatonin Counteracts Drought Induced Oxidative Damage and Stimulates Growth, Productivity and Fruit Quality Properties of Tomato Plants. Plants 2020, 9, 1276. [Google Scholar] [CrossRef]
- Roy, M.; Niu, J.; Irshad, A.; Kareem, H.A.; Hassan, M.U.; Xu, N.; Sui, X.; Guo, Z.; Amo, A.; Wang, Q. Exogenous melatonin protects alfalfa (Medicago sativa L.) seedlings from drought-induced damage by modulating reactive oxygen species metabolism, mineral balance and photosynthetic efficiency. Plant Stress 2021, 2, 100044. [Google Scholar] [CrossRef]
- Cherono, S.; Ntini, C.; Wassie, M.; Mollah, M.D.; Belal, M.A.; Ogutu, C.; Han, Y. Exogenous Application of Melatonin Improves Drought Tolerance in Coffee by Regulating Photosynthetic Efficiency and Oxidative Damage. J. Am. Soc. Hortic. Sci. 2021, 146, 24–32. [Google Scholar] [CrossRef]
- Imran, M.; Khan, A.L.; Shahzad, R.; Khan, M.A.; Bilal, S.; Khan, A.; Kang, S.-M.; Lee, I.-J. Exogenous melatonin induces drought stress tolerance by promoting plant growth and antioxidant defence system of soybean plants. AoB Plants 2021, 13, plab026. [Google Scholar] [CrossRef]
- Bai, Y.; Xiao, S.; Zhang, Z.; Zhang, Y.; Sun, H.; Zhang, K.; Wang, X.; Bai, Z.; Li, C.; Liu, L. Melatonin improves the germination rate of cotton seeds under drought stress by opening pores in the seed coat. PeerJ 2020, 8, e9450. [Google Scholar] [CrossRef]
- Wang, X.; Li, F.; Chen, Z.; Yang, B.; Komatsu, S.; Zhou, S. Proteomic analysis reveals the effects of melatonin on soybean root tips under flooding stress. J. Proteom. 2021, 232, 104064. [Google Scholar] [CrossRef]
- Gu, X.; Xue, L.; Lu, L.; Xiao, J.; Song, G.; Xie, M.; Zhang, H. Melatonin Enhances the Waterlogging Tolerance of Prunus persica by Modulating Antioxidant Metabolism and Anaerobic Respiration. J. Plant Growth Regul. 2020, 40, 2178–2190. [Google Scholar] [CrossRef]
- Ali, M.; Kamran, M.; Abbasi, G.H.; Saleem, M.H.; Ahmad, S.; Parveen, A.; Malik, Z.; Afzal, S.; Ahmar, S.; Dawar, K.M.; et al. Melatonin-Induced Salinity Tolerance by Ameliorating Osmotic and Oxidative Stress in the Seedlings of Two Tomato (Solanum lycopersicum L.) Cultivars. J. Plant Growth Regul. 2020, 40, 2236–2248. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Hoveizeh, N.F.; Gholami, R.; Abdelrahman, M.; Tran, L.P. Exogenous melatonin mitigates salinity-induced damage in olive seedlings by modulating ion homeostasis, antioxidant defense, and phytohormone balance. Physiol. Plant. 2021, 173, 1682–1694. [Google Scholar] [CrossRef]
- Cen, H.; Wang, T.; Liu, H.; Tian, D.; Zhang, Y. Melatonin Application Improves Salt Tolerance of Alfalfa (Medicago sativa L.) by Enhancing Antioxidant Capacity. Plants 2020, 9, 220. [Google Scholar] [CrossRef]
- Liu, J.; Shabala, S.; Zhang, J.; Ma, G.; Chen, D.; Shabala, L.; Zeng, F.; Chen, Z.; Zhou, M.; Venkataraman, G.; et al. Melatonin improves rice salinity stress tolerance by NADPH oxidase-dependent control of the plasma membrane K+ transporters and K+ homeostasis. Plant Cell Environ. 2020, 43, 2591–2605. [Google Scholar] [CrossRef]
- Mohamed, I.A.A.; Shalby, N.; El-Badri, A.M.A.; Saleem, M.H.; Khan, M.N.; Nawaz, M.A.; Qin, M.; Agami, R.A.; Kuai, J.; Wang, B.; et al. Stomata and Xylem Vessels Traits Improved by Melatonin Application Contribute to Enhancing Salt Tolerance and Fatty Acid Composition of Brassica napus L. Plants. Agronomy 2020, 10, 1186. [Google Scholar] [CrossRef]
- Ni, J.; Wang, Q.; Shah, F.A.; Liu, W.; Wang, D.; Huang, S.; Fu, S.; Wu, L. Exogenous Melatonin Confers Cadmium Tolerance by Counterbalancing the Hydrogen Peroxide Homeostasis in Wheat Seedlings. Molecules 2018, 23, 799. [Google Scholar] [CrossRef]
- Sun, C.; Lv, T.; Huang, L.; Liu, X.; Jin, C.; Lin, X. Melatonin ameliorates aluminum toxicity through enhancing aluminum exclusion and reestablishing redox homeostasis in roots of wheat. J. Pineal Res. 2020, 68, e12642. [Google Scholar] [CrossRef]
- Jahan, M.S.; Wang, Y.; Shu, S.; Hasan, M.M.; El-Yazied, A.A.; Alabdallah, N.M.; Hajjar, D.; Altaf, M.A.; Sun, J.S.; Guo, S. Melatonin Pretreatment Confers Heat Tolerance and Repression of Heat-Induced Senescence in Tomato through the Modulation of ABA- and GA-Mediated Pathways. Front. Plant Sci. 2021, 12, 650955. [Google Scholar] [CrossRef]
- Jia, C.; Yu, X.; Zhang, M.; Liu, Z.; Zou, P.; Ma, J.; Xu, Y. Application of Melatonin-Enhanced Tolerance to High-Temperature Stress in Cherry Radish (Raphanus sativus L. var. radculus pers). J. Plant Growth Regul. 2019, 39, 631–640. [Google Scholar] [CrossRef]
- Barman, D.; Ghimire, O.P.; Chinnusamy, V.; Kumar, R.R.; Arora, A. Amelioration of heat stress during reproductive stage in rice by melatonin. Indian J. Agric. Sci. 2019, 89, 7. [Google Scholar] [CrossRef]
- Qi, Z.-Y.; Wang, K.-X.; Yan, M.-Y.; Kanwar, M.K.; Li, D.-Y.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P.; Zhou, J. Melatonin Alleviates High Temperature-Induced Pollen Abortion in Solanum lycopersicum. Molecules 2018, 23, 386. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Khan, M.A.; Shahzad, R.; Bilal, S.; Khan, M.; Yun, B.-W.; Khan, A.L.; Lee, I.-J. Melatonin Ameliorates Thermotolerance in Soybean Seedling through Balancing Redox Homeostasis and Modulating Antioxidant Defense, Phytohormones and Polyamines Biosynthesis. Molecules 2021, 26, 5116. [Google Scholar] [CrossRef]
- Liang, D.; Gao, F.; Ni, Z.; Lin, L.; Deng, Q.; Tang, Y.; Wang, X.; Luo, X.; Xia, H. Melatonin Improves Heat Tolerance in Kiwifruit Seedlings through Promoting Antioxidant Enzymatic Activity and Glutathione S-Transferase Transcription. Molecules 2018, 23, 584. [Google Scholar] [CrossRef]
- Alam, M.N.; Zhang, L.; Yang, L.; Islam, R.; Liu, Y.; Luo, H.; Yang, P.; Wang, Q.; Chan, Z. Transcriptomic profiling of tall fescue in response to heat stress and improved thermotolerance by melatonin and 24-epibrassinolide. BMC Genom. 2018, 19, 224. [Google Scholar] [CrossRef]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. Short-term heat stress during flowering results in a decline in Canola seed productivity. J. Agron. Crops Sci. 2022, 208, 486–496. [Google Scholar] [CrossRef]
- Lohani, N.; Babaei, S.; Singh, M.; Bhalla, P. Genome-Wide In Silico Identification and Comparative Analysis of Dof Gene Family in Brassica napus. Plants 2021, 10, 709. [Google Scholar] [CrossRef]
- Arya, H.; Singh, M.B.; Bhalla, P.L. Towards Developing Drought-smart Soybeans. Front. Plant Sci. 2021, 12, 750664. [Google Scholar] [CrossRef]
- Babaei, S.; Singh, M.B.; Bhalla, P.L. Role of long non-coding RNAs in rice reproductive development. Front. Plant Sci. 2022, 13, 1040366. [Google Scholar] [CrossRef]
- Müller, F.; Rieu, I. Acclimation to high temperature during pollen development. Plant Reprod. 2016, 29, 107–118. [Google Scholar] [CrossRef]
- Xie, Z.; Wang, J.; Wang, W.; Wang, Y.; Xu, J.; Li, Z.; Zhao, X.; Fu, B. Integrated Analysis of the Transcriptome and Metabolome Revealed the Molecular Mechanisms Underlying the Enhanced Salt Tolerance of Rice Due to the Application of Exogenous Melatonin. Front. Plant Sci. 2021, 11, 618680. [Google Scholar] [CrossRef]
- Hoque, N.; Arif, T.U.; Hannan, A.; Sultana, N.; Akhter, S.; Hasanuzzaman, M.; Akter, F.; Hossain, S.; Sayed, A.; Hasan, T.; et al. Melatonin Modulates Plant Tolerance to Heavy Metal Stress: Morphological Responses to Molecular Mechanisms. Int. J. Mol. Sci. 2021, 22, 11445. [Google Scholar] [CrossRef]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to Stress Combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef]
- Kaya, C.; Higgs, D.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Integrative roles of nitric oxide and hydrogen sulfide in melatonin-induced tolerance of pepper (Capsicum annuum L.) plants to iron deficiency and salt stress alone or in combination. Physiol. Plant. 2020, 168, 256–277. [Google Scholar] [CrossRef]
- Li, J.; Ding, D.; Li, N.; Xie, J.; Yu, J.; Lyv, J.; Bakpa, E.P.; Zhang, J.; Wang, C.; Zhang, J. Melatonin enhances the low-temperature combined low-light tolerance of pepper (Capsicum annuum L.) seedlings by regulating photosynthesis, carotenoid, and hormone metabolism. Environ. Exp. Bot. 2022, 199, 104868. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Mangal, V.; Altaf, M.A.; Sharma, S.; Singh, B.; Kumar, M. Insight into melatonin-mediated response and signaling in the regulation of plant defense under biotic stress. Plant Mol. Biol. 2022, 109, 385–399. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, X.; Reiter, R.J.; Feng, S.; Wang, Y.; Liu, S.; Jin, L.; Li, Z.; Datla, R.; Ren, M. Melatonin Attenuates Potato Late Blight by Disrupting Cell Growth, Stress Tolerance, Fungicide Susceptibility and Homeostasis of Gene Expression in Phytophthora infestans. Front. Plant Sci. 2017, 8, 1993. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Z.; Lan, G.; Jiao, C.; Sun, Y. Effect ofexogenous melatonin on resistance of cucumber to downy mildew. Sci. Hortic. 2019, 255, 231–241. [Google Scholar] [CrossRef]
- Liu, Q.; Atta, U.R.; Wang, R.; Liu, K.; Ma, X.; Weng, Q. Defense-related hormone signaling coordinately controls the role of melatonin during Arabidopsis thaliana-Pseudomonas syringae interaction. Eur. J. Plant Pathol. 2021, 160, 707–716. [Google Scholar] [CrossRef]
- Zhao, H.; Xu, L.; Su, T.; Jiang, Y.; Hu, L.; Ma, F. Melatonin regulates carbohydrate metabolism and defenses against Pseudomonas syringae pv. tomato DC 3000 infection in Arabidopsis thaliana. J. Pineal Res. 2015, 59, 109–119. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. Melatonin is required for H2O2- and NO-mediated defense signaling through MAPKKK3 and OXI1 in Arabidopsis thaliana. J. Pineal Res. 2017, 62, e12379. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Du, C.; Wang, L.; Lai, M.; Fan, H. Exogenous melatonin improves the resistance to cucumber bacterial angular leaf spot caused by Pseudomonas syringae pv. Lachrymans. Physiol. Plant. 2022, 174, e13724. [Google Scholar] [CrossRef]
- Sofy, A.; Sofy, M.; Hmed, A.; Dawoud, R.; Refaey, E.; Mohamed, H.; El-Dougdoug, N. Molecular Characterization of the Alfalfa mosaic virus Infecting Solanum melongena in Egypt and the Control of Its Deleterious Effects with Melatonin and Salicylic Acid. Plants 2021, 10, 459. [Google Scholar] [CrossRef]
- Acosta, M.G.; Cano, A.; Hernández-Ruiz, J.; Arnao, M.B. Melatonin as a Possible Natural Safener in Crops. Plants 2022, 11, 890. [Google Scholar] [CrossRef]
- Zhang, L.; Jia, J.; Xu, Y.; Wang, Y.; Hao, J.; Li, T. Production of transgenic Nicotiana sylvestris plants expressing melatonin synthetase genes and their effect on UV-B-induced DNA damage. Vitr. Cell. Dev. Biol. Plant 2012, 48, 275–282. [Google Scholar] [CrossRef]
- Jinxiang, A.; Jiayi, S.; Zhenan, Y.; Zhichao, W.; Wenqian, C.; Yuhuan, W.; Yanyan, W.; Leilei, P.; Yutao, X.; Peng, L. Effects of exogenous melatonin on physiological response and DNA damage of Ardisia mamillata Hanceand and Ardisia crenata Sims under lead stress. Chin. Bull. Bot. 2022, 57, 171–181. [Google Scholar] [CrossRef]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. Rapid Transcriptional Reprogramming Associated with Heat Stress-Induced Unfolded Protein Response in Developing Brassica napus Anthers. Front. Plant Sci. 2022, 13, 905674. [Google Scholar] [CrossRef]
- Singh, M.B.; Lohani, N.; Bhalla, P.L. The Role of Endoplasmic Reticulum Stress Response in Pollen Development and Heat Stress Tolerance. Front. Plant Sci. 2021, 12, 661062. [Google Scholar] [CrossRef]
- Howell, S.H. Endoplasmic Reticulum Stress Responses in Plants. Annu. Rev. Plant Biol. 2013, 64, 477–499. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic Reticulum Stress and Oxidative Stress: A Vicious Cycle or a Double-Edged Sword? Antioxid. Redox Signal. 2007, 9, 2277–2293. [Google Scholar] [CrossRef]
- Lei, X.-Y.; Zhu, R.-Y.; Zhang, G.-Y.; Dai, Y.-R. Attenuation of cold-induced apoptosis by exogenous melatonin in carrot suspension cells: The possible involvement of polyamines. J. Pineal Res. 2004, 36, 126–131. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, K.; Zhou, X.; Xi, L.; Wang, Y.; Xu, H.; Pan, T.; Zou, Z. Melatonin alleviates chilling stress in cucumber seedlings by up-regulation of CsZat12 and modulation of polyamine and abscisic acid metabolism. Sci. Rep. 2017, 7, 4998. [Google Scholar] [CrossRef]
- Du, H.; Liu, G.; Hua, C.; Liu, D.; He, Y.; Liu, H.; Kurtenbach, R.; Ren, D. Exogenous melatonin alleviated chilling injury in harvested plum fruit via affecting the levels of polyamines conjugated to plasma membrane. Postharvest Biol. Technol. 2021, 179, 111585. [Google Scholar] [CrossRef]
- Marco, F.; Alcazar, R.; Tiburcio, A.F.; Carrasco, P. Interactions between Polyamines and Abiotic Stress Pathway Responses Unraveled by Transcriptome Analysis of Polyamine Overproducers. OMICS A J. Integr. Biol. 2011, 15, 775–781. [Google Scholar] [CrossRef]
- Sagor, G.H.M.; Chawla, P.; Kim, D.W.; Berberich, T.; Kojima, S.; Niitsu, M.; Kusano, T. The polyamine spermine induces the unfolded protein response via the MAPK cascade in Arabidopsis. Front. Plant Sci. 2015, 6, 687. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. Melatonin plays a pivotal role in conferring tolerance against endoplasmic reticulum stress via mitogen-activated protein kinases and bZIP60 in Arabidopsis thaliana. Melatonin Res. 2018, 1, 94–108. [Google Scholar] [CrossRef]
- Ozgur, R.; Uzilday, B.; Turkan, I.; Sekmen, A.H. The Effects of Melatonin on Transcriptional Profile of Unfolded Protein Response Genes under Endoplasmic Reticulum Stress in Arabidopsis thaliana. Plant Mol. Biol. Rep. 2016, 35, 188–202. [Google Scholar] [CrossRef]
- Avin-Wittenberg, T. Autophagy and its role in plant abiotic stress management. Plant Cell Environ. 2019, 42, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Sun, X.; Wang, N.; Tan, D.-X.; Ma, F. Melatonin enhances the occurrence of autophagy induced by oxidative stress in Arabidopsis seedlings. J. Pineal Res. 2015, 58, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Yu, X.; Lou, W.; Wei, S.; Wang, R.; Wan, Q.; Shen, W. Transgenic Arabidopsis overexpressing MsSNAT enhances salt tolerance via the increase in autophagy, and the reestablishment of redox and ion homeostasis. Environ. Exp. Bot. 2019, 164, 20–28. [Google Scholar] [CrossRef]
- Wei, Y.; Bai, Y.; Cheng, X.; Zhu, B.; Reiter, R.J.; Shi, H. The dual roles of melatonin biosynthesis enzymes in the coordination of melatonin biosynthesis and autophagy in cassava. J. Pineal Res. 2020, 69, e12652. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Cai, S.-Y.; Zhang, Y.; Xu, Y.-P.; Qi, Z.-Y.; Li, M.-Q.; Ahammed, G.J.; Xia, X.-J.; Shi, K.; Zhou, Y.-H.; Reiter, R.J.; et al. HsfA1a upregulates melatonin biosynthesis to confer cadmium tolerance in tomato plants. J. Pineal Res. 2017, 62, e12387. [Google Scholar] [CrossRef]
- Shi, H.; Tan, D.-X.; Reiter, R.J.; Ye, T.; Yang, F.; Chan, Z. Melatonin induces class A1 heat-shock factors (HSFA1s) and their possible involvement of thermotolerance in Arabidopsis. J. Pineal Res. 2015, 58, 335–342. [Google Scholar] [CrossRef]
- Xia, H.; Zhou, Y.; Deng, H.; Lin, L.; Deng, Q.; Wang, J.; Lv, X.; Zhang, X.; Liang, D. Melatonin improves heat tolerance in Actinidia deliciosa via carotenoid biosynthesis and heat shock proteins expression. Physiol. Plant. 2021, 172, 1582–1593. [Google Scholar] [CrossRef]
- Yao, L.; Cheng, X.; Gu, Z.; Huang, W.; Li, S.; Wang, L.; Wang, Y.-F.; Xu, P.; Ma, H.; Ge, X. The AWPM-19 Family Protein OsPM1 Mediates Abscisic Acid Influx and Drought Response in Rice. Plant Cell 2018, 30, 1258–1276. [Google Scholar] [CrossRef]
- Xing, X.; Ding, Y.; Jin, J.; Song, A.; Chen, S.; Chen, F.; Fang, W.; Jiang, J. Physiological and Transcripts Analyses Reveal the Mechanism by Which Melatonin Alleviates Heat Stress in Chrysanthemum Seedlings. Front. Plant Sci. 2021, 12, 673236. [Google Scholar] [CrossRef]
- Li, X.; Tan, D.-X.; Jiang, D.; Liu, F. Melatonin enhances cold tolerance in drought-primed wild-type and abscisic acid-deficient mutant barley. J. Pineal Res. 2016, 61, 328–339. [Google Scholar] [CrossRef]
- Fu, J.; Wu, Y.; Miao, Y.; Xu, Y.; Zhao, E.; Wang, J.; Sun, H.; Liu, Q.; Xue, Y.; Xu, Y.; et al. Improved cold tolerance in Elymus nutans by exogenous application of melatonin may involve ABA-dependent and ABA-independent pathways. Sci. Rep. 2017, 7, 39865. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin promotes adventitious- and lateral root regeneration in etiolated hypocotyls of Lupinus albus L. J. Pineal Res. 2007, 42, 147–152. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Bot. 2015, 66, 657–668. [Google Scholar] [CrossRef]
- Xu, L.; Yue, Q.; Bian, F.; Sun, H.; Zhai, H.; Yao, Y. Melatonin Enhances Phenolics Accumulation Partially via Ethylene Signaling and Resulted in High Antioxidant Capacity in Grape Berries. Front. Plant Sci. 2017, 8, 1426. [Google Scholar] [CrossRef]
- Onik, J.C.; Wai, S.C.; Li, A.; Lin, Q.; Sun, Q.; Wang, Z.; Duan, Y. Melatonin treatment reduces ethylene production and maintains fruit quality in apple during postharvest storage. Food Chem. 2021, 337, 127753. [Google Scholar] [CrossRef]
- Liu, J.; Yang, J.; Zhang, H.; Cong, L.; Zhai, R.; Yang, C.; Wang, Z.; Ma, F.; Xu, L. Melatonin Inhibits Ethylene Synthesis via Nitric Oxide Regulation to Delay Postharvest Senescence in Pears. J. Agric. Food Chem. 2019, 67, 2279–2288. [Google Scholar] [CrossRef]
- Xu, L.; Xiang, G.; Sun, Q.; Ni, Y.; Jin, Z.; Gao, S.; Yao, Y. Melatonin enhances salt tolerance by promoting MYB108A-mediated ethylene biosynthesis in grapevines. Hortic. Res. 2019, 6, 114. [Google Scholar] [CrossRef]
- Yang, L.; Bu, S.; Zhao, S.; Wang, N.; Xiao, J.; He, F.; Gao, X. Transcriptome and physiological analysis of increase in drought stress tolerance by melatonin in tomato. PLoS ONE 2022, 17, e0267594. [Google Scholar] [CrossRef]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. High temperature susceptibility of sexual reproduction in crop plants. J. Exp. Bot. 2020, 71, 555–568. [Google Scholar] [CrossRef]
- Jahan, M.S.; Guo, S.; Sun, J.; Shu, S.; Wang, Y.; El-Yazied, A.A.; Alabdallah, N.M.; Hikal, M.; Mohamed, M.H.; Ibrahim, M.F.; et al. Melatonin-mediated photosynthetic performance of tomato seedlings under high-temperature stress. Plant Physiol. Biochem. 2021, 167, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, H.-J.; Sun, Q.-Q.; Cao, Y.-Y.; Li, X.; Zhao, B.; Wu, P.; Guo, Y.-D. Proteomic analysis reveals a role of melatonin in promoting cucumber seed germination under high salinity by regulating energy production. Sci. Rep. 2017, 7, 503. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Liu, M.; Wang, X.; Liu, L.; Wu, H.; Chen, X.; Wang, H.; Shen, Q.; Chen, G.; Wang, Y. Seed-Soaking with Melatonin for the Improvement of Seed Germination, Seedling Growth, and the Antioxidant Defense System under Flooding Stress. Agronomy 2022, 12, 1918. [Google Scholar] [CrossRef]
- Zhang, M.; He, S.; Qin, B.; Jin, X.; Wang, M.; Ren, C.; Cao, L.; Zhang, Y. Exogenous melatonin reduces the inhibitory effect of osmotic stress on antioxidant properties and cell ultrastructure at germination stage of soybean. PLoS ONE 2020, 15, e0243537. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Deng, L.; Zhou, L.; Chen, G.; Wang, Y. Exogenous Melatonin Activates Antioxidant Systems to Increase the Ability of Rice Seeds to Germinate under High Temperature Conditions. Plants 2022, 11, 886. [Google Scholar] [CrossRef]
- Posmyk, M.M.; Bałabusta, M.; Wieczorek, M.; Sliwinska, E.; Janas, K.M. Melatonin applied to cucumber (Cucumis sativus L.) seeds improves germination during chilling stress. J. Pineal Res. 2009, 46, 214–223. [Google Scholar] [CrossRef]
- Celik, E.N.Y. Effects of Exogenous Melatonin Supplementations on Some Elemental Contents in Anatolian Black Pine (Pinus nigra J.F. Arnold. subsp. pallasiana (Lamb.) Holmboe) Seedling Tissues. BioResources 2021, 16, 5706–5720. [Google Scholar] [CrossRef]
- Song, R.; Ritonga, F.N.; Yu, H.; Ding, C.; Zhao, X. Effects of Exogenous Antioxidant Melatonin on Physiological and Biochemical Characteristics of Populus cathayana × canadansis ‘Xin Lin 1’ under Salt and Alkaline Stress. Forests 2022, 13, 1283. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Transgenic Plant | Transgenic Approach | Stress | Results | Reference |
|---|---|---|---|---|---|
| COMT | Arabidopsis (Arabidopsis thaliana) and Watermelon (Citrullus lanatus) | Overexpression—COMT gene (CICOMT from watermelon) driven by 35S promoter | Salt | Significant increase in melatonin content. Increased survival rate under salt, freeze and mannitol-induced drought stress. | [45] |
| Cold | |||||
| Drought | |||||
| Tomato (Solanum lycopersicum) | Overexpression—SICOMT1 driven by 35S promoter. | Salt | Elevated melatonin levels. Reduced drooping and wilting of leaves. Reduced superoxide and hydrogen peroxide levels. Increased proline levels. | [46] | |
| Arabidopsis (Arabidopsis thaliana) | Overexpression—COMT (TaCOMT from wheat) driven by 35S promoter | Drought | Elevated melatonin levels. Increased fresh weight, lateral root number, and total root length. | [47] | |
| SNAT | Tomato (Solanum lycopersicum) | Overexpression—SlSNAT gene driven by 35S promoter. | Heat | Elevated melatonin levels. Increased chlorophyll fluorescence (Fv/Fm). Reduced wilting after 1 day recovery. | [48] |
| Arabidopsis (Arabidopsis thaliana) | Overexpression—SNAT gene (MsSNAT from alfalfa) driven by 35S promoter. | Cadmium | Slightly elevated melatonin levels. Reduced root length decline and lowered cadmium deposition in roots. Reduced levels of miR398, indicating low oxidative stress | [49] | |
| Overexpression—SNAT gene (VvSNAT1 from grapes) driven by 35S promoter. | Salt | Significantly elevated melatonin content. Improved growth potential and greener leaves. | [50] | ||
| Overexpression—AtSNAT gene And SNAT mutant (SALK_032239) | UVB | Overexpression: Early and increased response in positive UV-B signalling regulatory genes. SALK_032239: Slow response under high and prolonged UVB treatment. | [51] | ||
| Cucumber (Cucumis sativus) | Overexpression—CsSNAT And Silencing—CsSNAT (RNAi) | Salt | Overexpression: Elevated melatonin content Less ROS accumulation and upregulation of antioxidant enzymes. Increased maximum photochemical efficiency of PSII (Fv/Fm). Silencing: Lowered melatonin content. Increased ROS accumulation and decreased antioxidant enzyme activity. Lowered maximum photochemical efficiency of PSII (Fv/Fm). | [52] | |
| ASMT | Tomato (Solanum lycopersicum) | Overexpression—SlASMT gene driven by 35S | Heat | Reduced wilting Reduced electrolyte leakage. Improved photosynthesis (Fv/Fm). Decreased insoluble and ubiquitinated proteins. Enhanced expression of heat shock protein genes and several autophagy genes. Decrease in accumulation of aggregated proteins. | [53] |
| Tobacco (Nicotiana tabacum) | Overexpression—ASMT (MzASMT1 from crab apple) driven by 35S promoter | Salt | Elevated melatonin contents. Lowered leaf wilting. Increased fresh weight, plant height, root length and relative water content. Higher chlorophyll content and improved photosynthesis (Fv/Fm). | [54] | |
| Arabidopsis (Arabidopsis thaliana) | MzASMT driven by 35S promoter has been transformed into Arabidopsis. | Drought | Elevated melatonin levels. Enhanced lateral roots. Increased fresh weight. | [55] |
| Stress | Plant | Growth Stage | Melatonin Application | Activity under Stress (Compared to Plants without Exogenous Melatonin Stressed) | Reference | |
|---|---|---|---|---|---|---|
| Optimised Concentration | Application Method | |||||
| Cold | Pepper (Capsicum annuum) | Seedling | 5 µM | Soil drench with 25 mL melatonin solution one day before chilling stress | Decreased hydrogen peroxide, malondialdehyde contents and membrane permeability. Increased photosynthesis, water relation and antioxidant enzyme activity. | [64] |
| Flowering | 5 µM | Soil drench with 100 mL melatonin solution | Increased the yield when applied during flowering. | [64] | ||
| Tea (Camellia sinensis) | Seedling (two-year-old) | 100 µM | Foliage spray—three times with five-day intervals. (last treatment—24 h before the stress) | Increased antioxidant enzyme activity and stimulate photosynthesis. | [65] | |
| Barley (Hordeum vulgare) | Seeds | 1 µM | Soaked the seeds for 12 h before germination. | Decreased malondialdehyde and soluble sugar content. Promoted seedling growth, increased chlorophyll content. | [66] | |
| Drought | Rapeseed (Brassica napus) | Seedling | 100 µM | Irrigation with 200 mL melatonin solution per pot each day for seven days. | Decreased malondialdehyde and hydrogen peroxide. Regulated leaf stomatal activity. Increased root growth and catalase activity. | [67] |
| Tomato (Solanum lycopersicum) | Young plants (5 weeks old) | 20 ppm | Foliar application | Decreased malondialdehyde and hydrogen peroxide. Increased yield and ascorbic acid content in fruits. | [68] | |
| Alfalfa (Medicago sativa) | Seedlings | 100 µM | Sprayed at dark, two days before the stress and repeated every three days up to 20 days. | Increased chlorophyll and carotenoid contents, photosynthetic rate and stomatal conductance, soluble sugar and proline content. Decreased malondialdehyde, hydrogen peroxide, electrolyte leakage and superoxide anion. | [69] | |
| Coffee (Coffea arabica) | Seedlings | 100 µM | Foliar spray of 20 mL and soil application of 30 mL three times per week. | Suppressed chlorophyll degradation and increased photosynthesis. Decreased malondialdehyde and electrolyte leakage. Increased enzymatic antioxidant activity. | [70] | |
| Soybean (Glycine max) | Seedlings | 100 µM | Rhizosphere application | Increased chlorophyll content, photosynthetic activity, shoot and root growth, enzymatic antioxidation. Increased salicylic and jasmonic acid content. Decreased malondialdehyde, electrolyte leakage and hydrogen peroxide. | [71] | |
| Cotton (Gossypium hirsutum) | Seeds | 100 µM | Soaked the seeds for 24 h prior to germination. | Increased soluble sugar and proline content. Increased stomatal regulation, germination rate, germination potential and fresh weight. Decreased the hydrogen peroxide, superoxide anion and malondialdehyde. | [72] | |
| Water Logging | Soybean (Glycine max) | Seedling | 10 µM | Root application at the same time of flood. | Increased root growth and development, increased root cell wall lignification. Increased alkaloid metabolism and ROS scavenging. | [73] |
| Alfalfa (Medicago sativa) | Seedling | 100 µM | Foliar spray one day before to the stress | Increased plant growth and photosynthesis. Increased endogenous melatonin levels. Increased polyamines and decreased ethylene. Decreased membrane damage and leaf senescence. | [60] | |
| Peach (Prunus persica) | seedlings | 200 µM | Applied to soil every other day during the stress. | Increased root and shoot development. Positive development in photosynthetic and stomatal apparatus. Increased antioxidant activities. Increased anaerobic respiration through enhanced aerenchyma. | [74] | |
| Salt | Tomato (Solanum lycopersicum) | Seedling | 1 µM | Applied to the medium mixed with saline treatment | Increased photosynthesis and antioxidant enzyme activity. Decreased malondialdehyde and hydrogen peroxide content. | [75] |
| Olive (Olea europaea) | Seedling | 100 µM | Foliar spray | Increased shoot and root growth, photosynthetic pigments, proline and soluble sugars. Increased enzymatic antioxidation. Decreased hydrogen peroxide, malondialdehyde and electrolyte leakage. | [76] | |
| Alfalfa (Medicago sativa) | Seeds | 10–100 µM | Seeds immersed and air-dried prior to germination | Increased seed germination, root length, seedling growth and enzymatic anti-oxidation. | [77] | |
| One-month-old plants | 50 µM | Foliar spray every night | Decreased hydrogen peroxide, malondialdehyde and electrolyte leakage | |||
| Rice (Oryza sativa) | Seedlings | 20 µM | Applied for 24 h prior to salt stress | Increased root and shoot growth. Increased the expression of stress-responsive genes. | [78] | |
| Rapeseed (Brassica napus) | Seeds | 50 µM | Seed primed for 8 h prior to germination | Decreased hydrogen peroxide and superoxide anions. Increase the regulation of antioxidant enzymes, chlorophyll content, photosynthetic rate and proline content. Improved the oil quality. | [79] | |
| Cadmium | Wheat (Triticum aestivum) | Seedling | 0.5–100 µM | Applied directly to the roots of seedlings growing in Petri dishes | Increased root and shoot growth, Increased enzymatic and non-enzymatic anti-oxidants. Decreased hydrogen peroxide content. | [80] |
| Aluminium | Wheat (Triticum aestivum) | Seedling | 10 µM | Treated for 12 h prior to the stress | Increased enzymatic and non-enzymatic antioxidant activity | [81] |
| Nickel | Tomato (Solanum lycopersicum | Seedling | 100 µM | Foliar sprayed with 80 mL solution with 3 days interval during the stress | Improved photosynthesis and gas exchange. Increased enzymatic anti-oxidation. Upregulation of stress-responsive genes. Decreased hydrogen peroxide, malondialdehyde and electrolyte leakage | [59] |
| Heat | Tomato (Solanum lycopersicum) | Seedling | 100 µM | Foliar sprayed every two days for seven days one week before the stress | Increased photosynthesis and stomatal activity. Decreased hydrogen peroxide, malondialdehyde and electrolyte leakage. Downregulation of genes encoding ROS accumulation. | [82] |
| Wheat (Triticum aestivum) | Seedling | 100 µM | Sprayed 80 mL of melatonin solution on leaves each day for seven days one week before the stress. | Increased chlorophyll content, enzymatic and non-enzymatic antioxidant activity and proline content. Decreased hydrogen peroxide and malondialdehyde. | [58] | |
| Cherry radish (Raphanus sativus) | Seedling | 29.0 mg/L | Applied on roots mixed with Hogland’s nutrient solution. | Increased chlorophyll, carotenoid content and enzymatic antioxidation. Decreased malondialdehyde. | [83] | |
| Strawberry (Fragaria × ananassa) | Young plants (3 weeks old) | 100 µM | Foliar spray three times at one-day intervals (last treatment—10 h prior to the stress) | Increased enzymatic and non-enzymatic antioxidant activity. Upregulation of stress-responsive genes. Decreased hydrogen peroxide, and malondialdehyde. | [57] | |
| Rice (Oryza sativa) | Flowering | 200 µM | Sprayed one day before the stress treatment. | Increased chlorophyll content and stomatal conductance. | [84] | |
| Tomato (Solanum lycopersicum | Flowering | 20 µM | Applied on roots on each day for 7 days followed by the heat stress. | Alleviated pollen abortion. Increased stability of tapetum cells and avoid pollen deformity by inducing stress-responsive genes. Increased ROS scavenging and enzymatic antioxidant activity. | [85] | |
| Soybean (Glycine max) | Seedling | 100 µM | Applied on root zone (30 mL) twice daily for 6 days prior to stress. | Increased chlorophyll content and non-enzymatic anti-oxidation. Decreased hydrogen peroxide, superoxide, malondialdehyde and electronic leakage. | [86] | |
| Kiwifruit (Actinidia deliciosa) | Seedling | 200 µM | Treated five times every two days prior to stress. | Increased proline, enzymatic and non-enzymatic antioxidant activity. Reduced hydrogen peroxide. | [87] | |
| Tall fescue (Festuca arundinacea Schreb.) | Seedling | 20 µM | Seedlings were transferred to MS medium containing melatonin two days prior to the stress. | Increased antioxidant enzyme activity and chlorophyll content. Changed stress-responsive gene regulation. Reduced hydrogen peroxide, superoxide anion, malondialdehyde and electronic leakage. | [88] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colombage, R.; Singh, M.B.; Bhalla, P.L. Melatonin and Abiotic Stress Tolerance in Crop Plants. Int. J. Mol. Sci. 2023, 24, 7447. https://doi.org/10.3390/ijms24087447
Colombage R, Singh MB, Bhalla PL. Melatonin and Abiotic Stress Tolerance in Crop Plants. International Journal of Molecular Sciences. 2023; 24(8):7447. https://doi.org/10.3390/ijms24087447
Chicago/Turabian StyleColombage, Roshira, Mohan B. Singh, and Prem L. Bhalla. 2023. "Melatonin and Abiotic Stress Tolerance in Crop Plants" International Journal of Molecular Sciences 24, no. 8: 7447. https://doi.org/10.3390/ijms24087447
APA StyleColombage, R., Singh, M. B., & Bhalla, P. L. (2023). Melatonin and Abiotic Stress Tolerance in Crop Plants. International Journal of Molecular Sciences, 24(8), 7447. https://doi.org/10.3390/ijms24087447