
Characterization of Porcine Ocular Surface Epithelial Microenvironment

 ,
, 
 and
and
Abstract
:1. Introduction
2. Results and Discussion
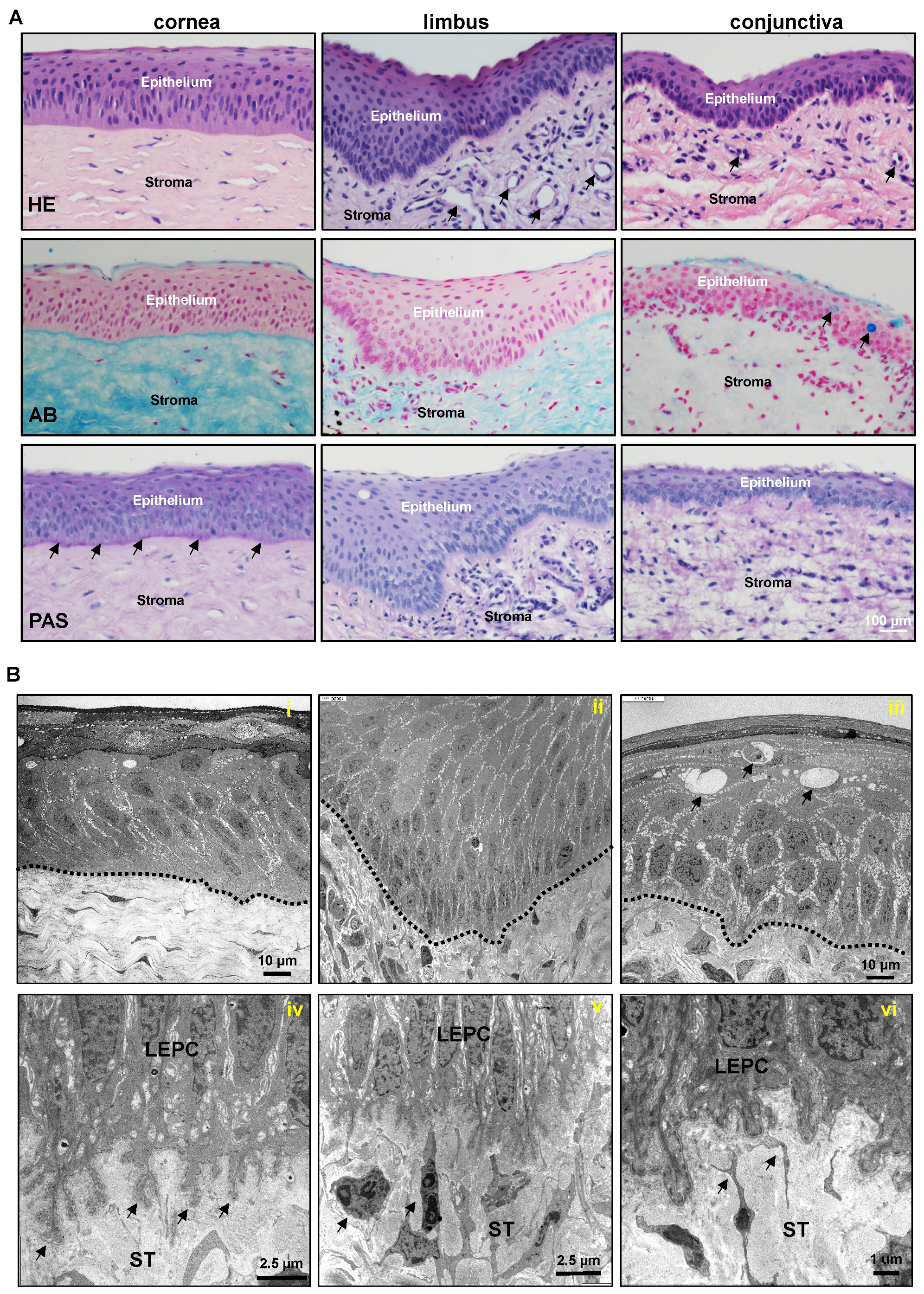
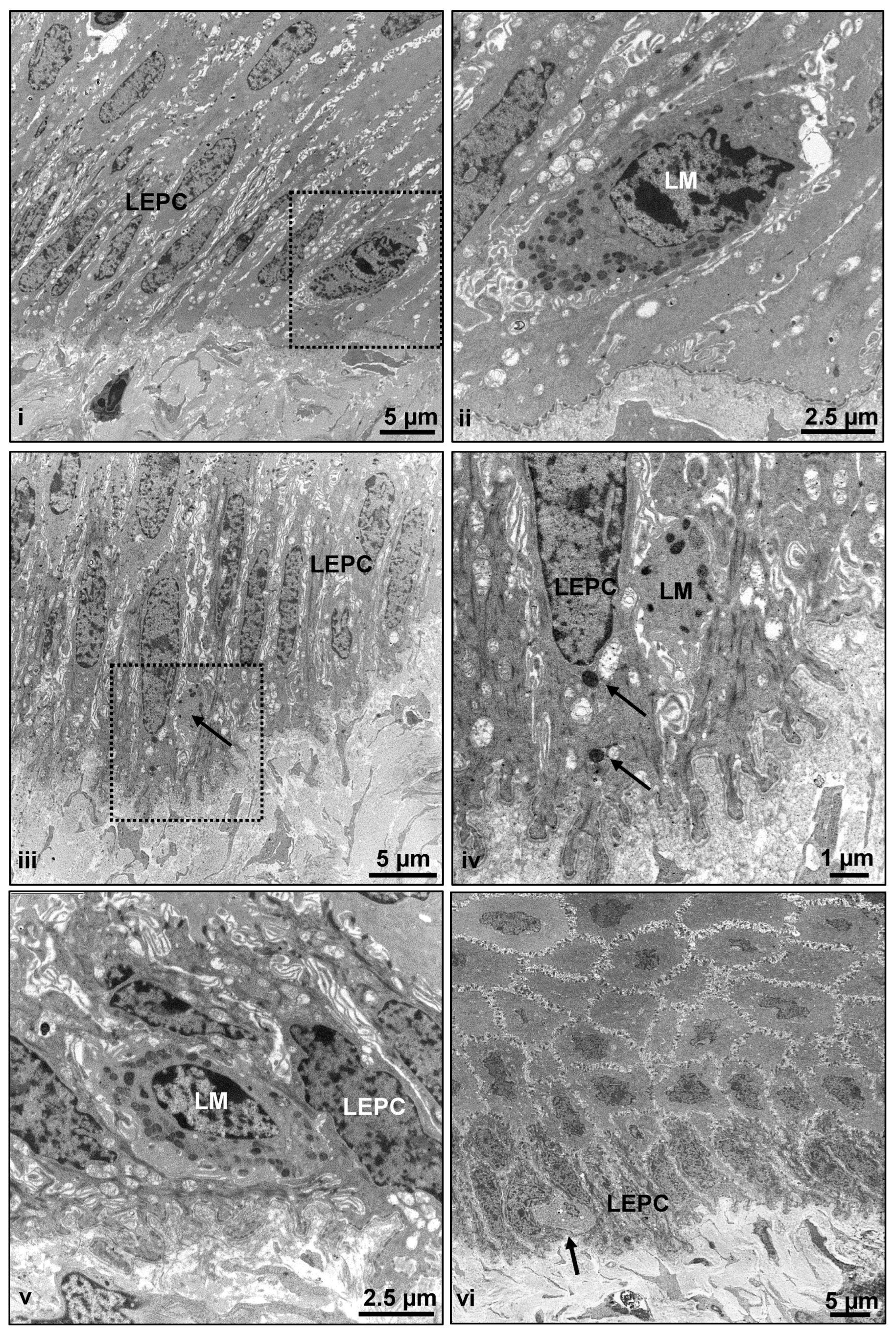
2.1. Architecture of Porcine Ocular Surface
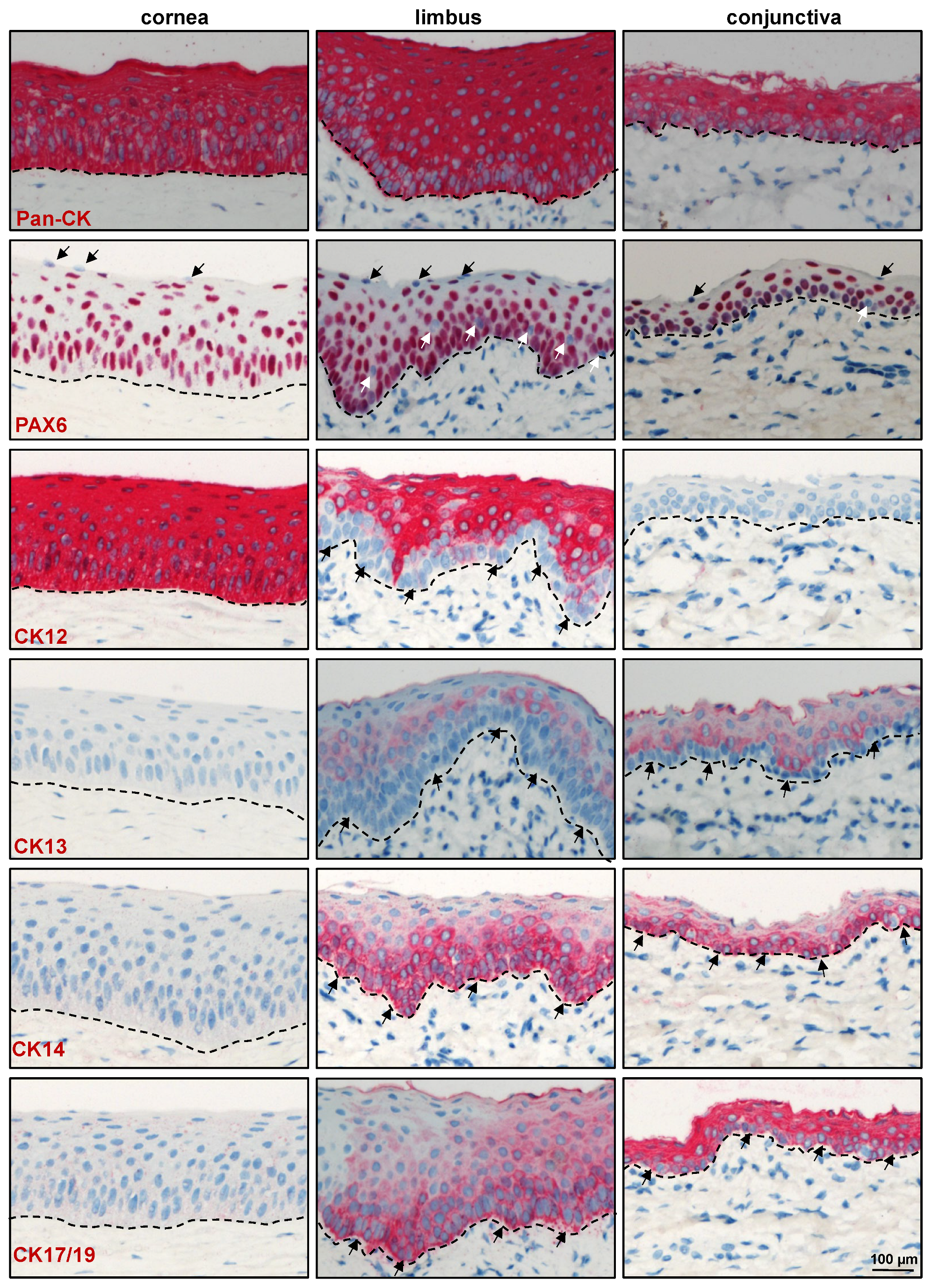
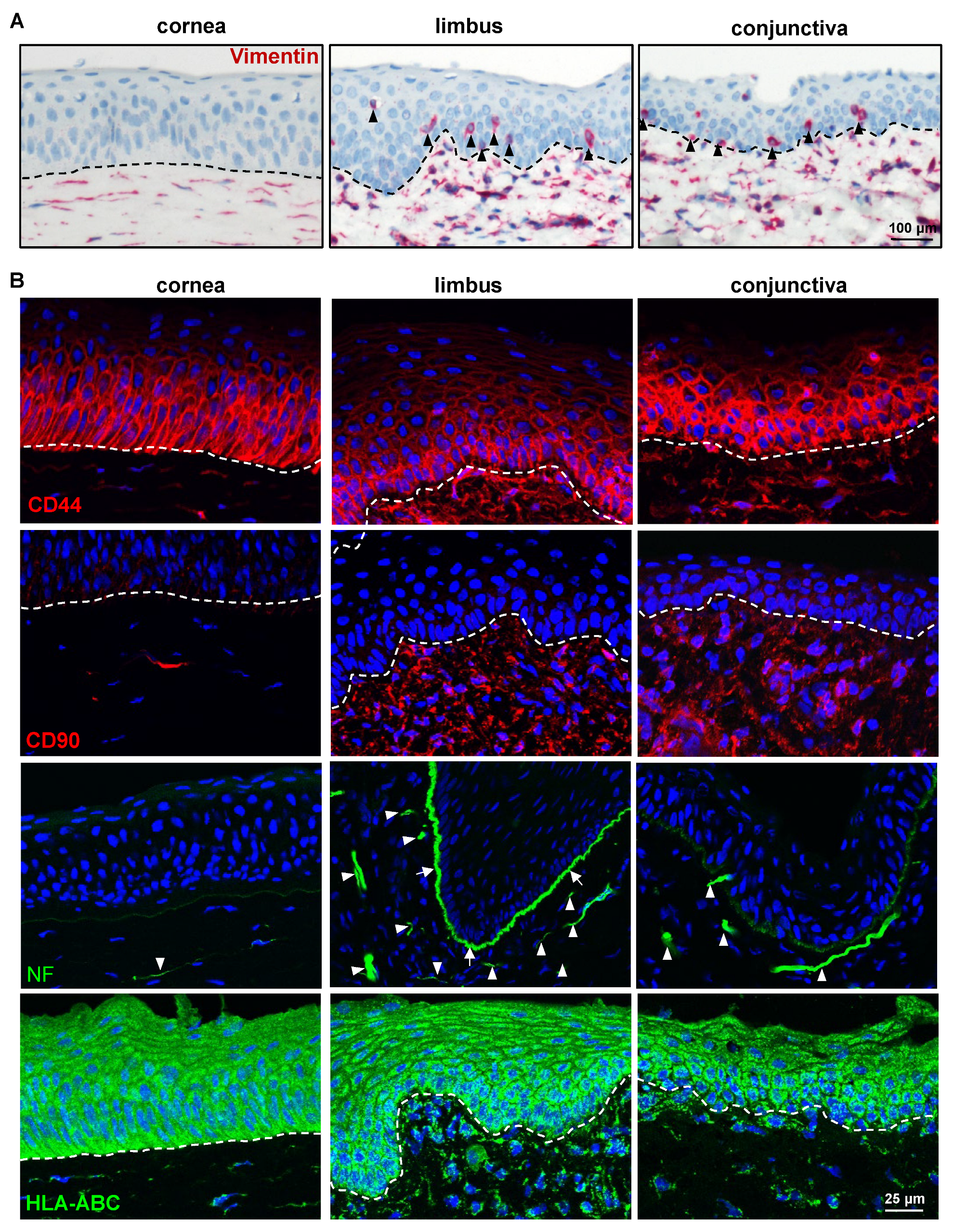
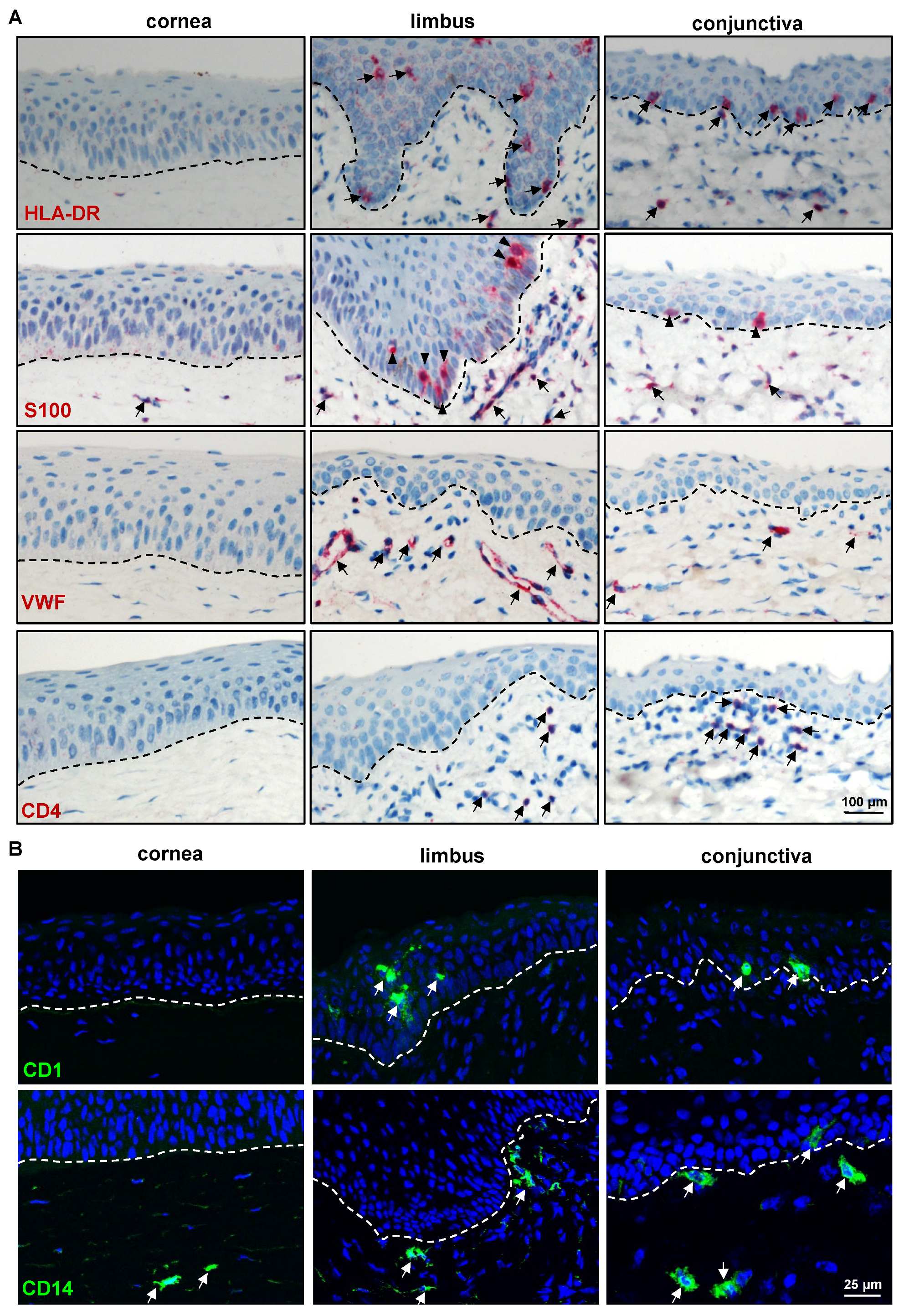
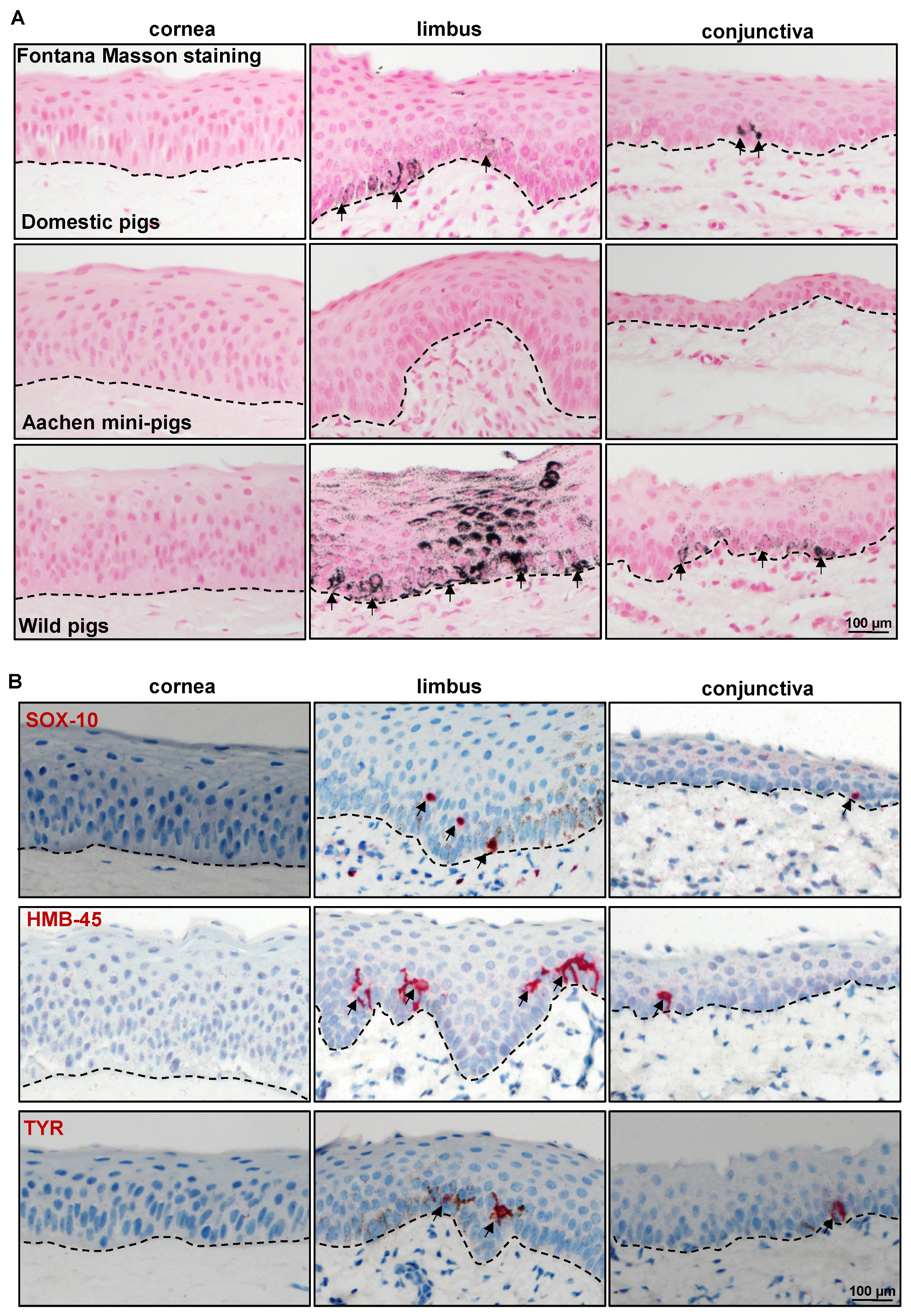
2.2. Immunohistochemical Characterization of the Porcine Ocular Surface
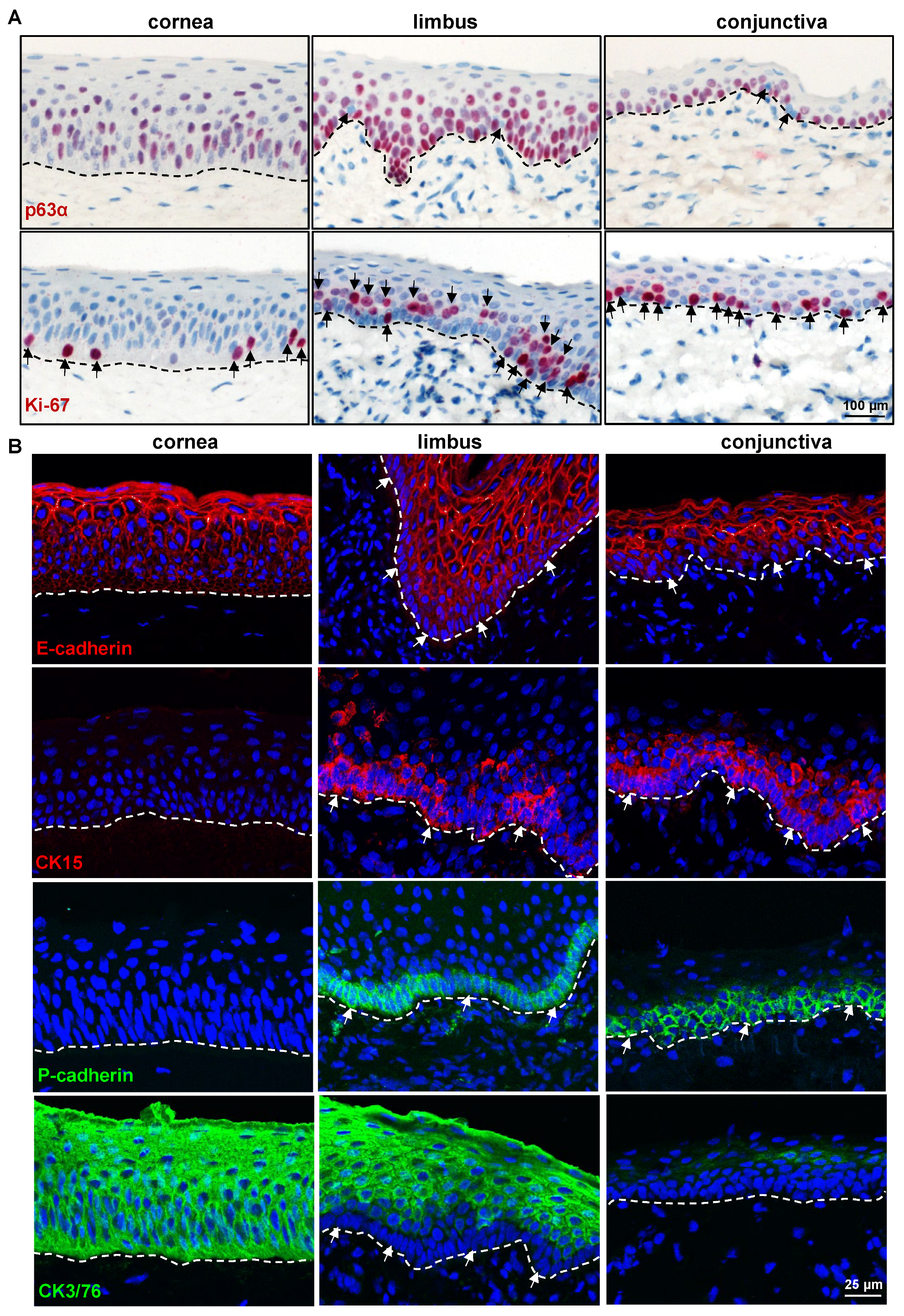
2.2.1. Epithelial Progenitor/Differentiation Markers
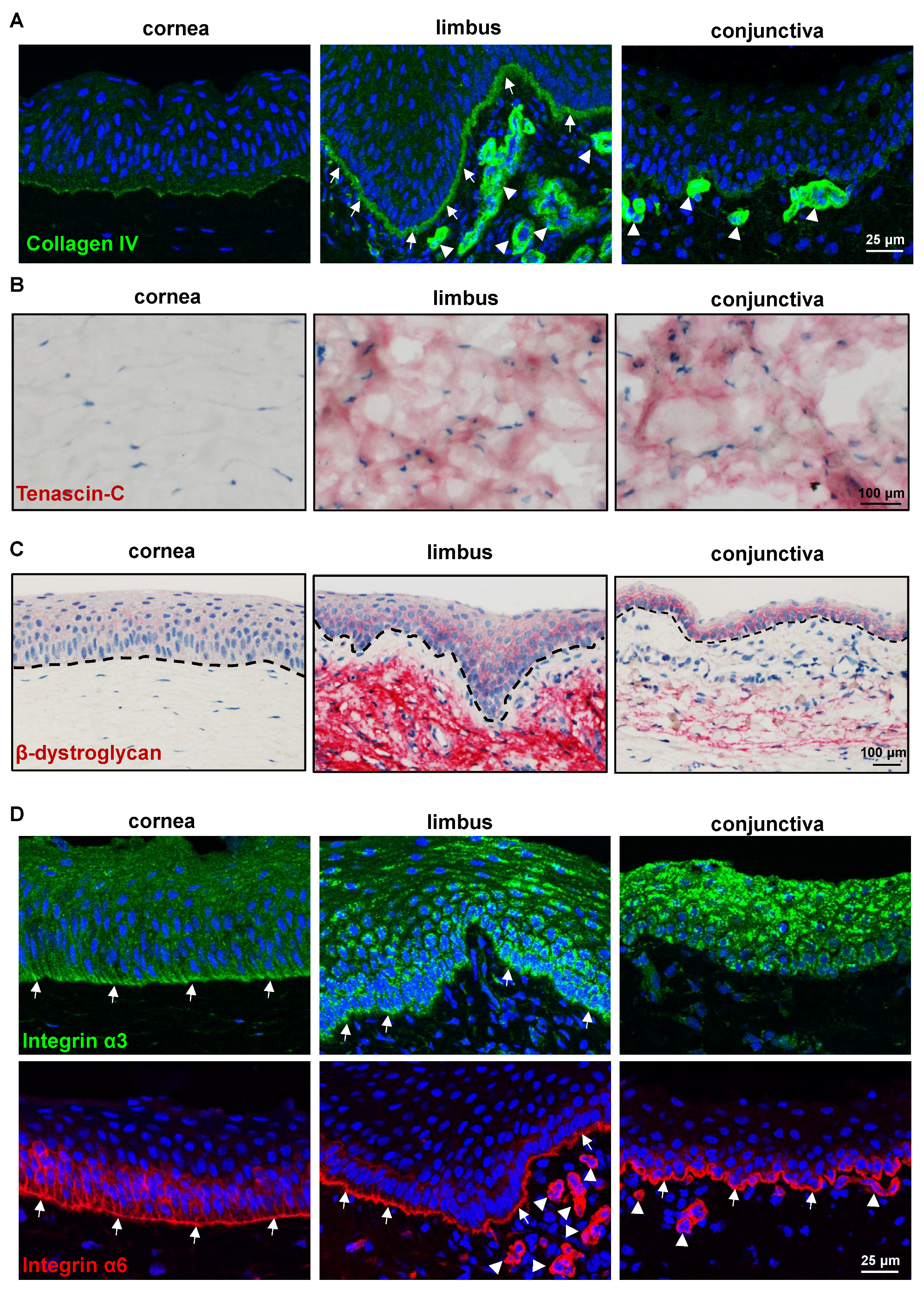
2.2.2. ECM and Its Associated Markers
2.2.3. Limbal Niche Cell Markers
3. Materials and Methods
3.1. Porcine Eyes
3.2. Tissue Preparation and Histology
3.3. Immunostaining of Frozen Sections
3.4. Immunostaining of Paraffin Sections
3.5. Transmission Electron Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hodges, R.R.; Dartt, D.A. Tear Film Mucins: Front Line Defenders of the Ocular Surface; Comparison with Airway and Gastrointestinal Tract Mucins. Exp. Eye Res. 2013, 117, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Cotsarelis, G.; Cheng, S.Z.; Dong, G.; Sun, T.T.; Lavker, R.M. Existence of Slow-Cycling Limbal Epithelial Basal Cells That Can Be Preferentially Stimulated to Proliferate: Implications on Epithelial Stem Cells. Cell 1989, 57, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.F.; Bron, A.J. Limbal Palisades of Vogt. Trans. Am. Ophthalmol. Soc. 1982, 80, 155–171. [Google Scholar] [PubMed]
- Shortt, A.J.; Secker, G.A.; Munro, P.M.; Khaw, P.T.; Tuft, S.J.; Daniels, J.T. Characterization of the Limbal Epithelial Stem Cell Niche: Novel Imaging Techniques Permit in Vivo Observation and Targeted Biopsy of Limbal Epithelial Stem Cells. Stem Cells 2007, 25, 1402–1409. [Google Scholar] [CrossRef]
- Schlötzer-Schrehardt, U.; Kruse, F.E. Identification and Characterization of Limbal Stem Cells. Exp. Eye Res. 2005, 81, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Stewart, R.M.K.; Sheridan, C.M.; Hiscott, P.S.; Czanner, G.; Kaye, S.B. Human Conjunctival Stem Cells Are Predominantly Located in the Medial Canthal and Inferior Forniceal Areas. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2021–2030. [Google Scholar] [CrossRef]
- Kate, A.; Basu, S. A Review of the Diagnosis and Treatment of Limbal Stem Cell Deficiency. Front. Med. 2022, 9, 836009. [Google Scholar] [CrossRef] [PubMed]
- Rama, P.; Matuska, S.; Paganoni, G.; Spinelli, A.; De Luca, M.; Pellegrini, G. Limbal Stem-Cell Therapy and Long-Term Corneal Regeneration. N. Engl. J. Med. 2010, 363, 147–155. [Google Scholar] [CrossRef]
- Elhusseiny, A.M.; Soleimani, M.; Eleiwa, T.K.; ElSheikh, R.H.; Frank, C.R.; Naderan, M.; Yazdanpanah, G.; Rosenblatt, M.I.; Djalilian, A.R. Current and Emerging Therapies for Limbal Stem Cell Deficiency. Stem Cells Transl. Med. 2022, 11, 259–268. [Google Scholar] [CrossRef]
- Li, J.; Xiao, Y.; Coursey, T.G.; Chen, X.; Deng, R.; Lu, F.; Pflugfelder, S.C.; Li, D.-Q. Identification for Differential Localization of Putative Corneal Epithelial Stem Cells in Mouse and Human. Sci. Rep. 2017, 7, 5169. [Google Scholar] [CrossRef]
- Delic, N.C.; Cai, J.R.; Watson, S.L.; Downie, L.E.; Di Girolamo, N. Evaluating the Clinical Translational Relevance of Animal Models for Limbal Stem Cell Deficiency: A Systematic Review. Ocul. Surf. 2022, 23, 169–183. [Google Scholar] [CrossRef]
- Kethiri, A.R.; Raju, E.; Bokara, K.K.; Mishra, D.K.; Basu, S.; Rao, C.M.; Sangwan, V.S.; Singh, V. Inflammation, Vascularization and Goblet Cell Differences in LSCD: Validating Animal Models of Corneal Alkali Burns. Exp. Eye Res. 2019, 185, 107665. [Google Scholar] [CrossRef] [PubMed]
- Majo, F.; Rochat, A.; Nicolas, M.; Jaoudé, G.A.; Barrandon, Y. Oligopotent Stem Cells Are Distributed throughout the Mammalian Ocular Surface. Nature 2008, 456, 250–254. [Google Scholar] [CrossRef]
- Notara, M.; Schrader, S.; Daniels, J.T. The Porcine Limbal Epithelial Stem Cell Niche as a New Model for the Study of Transplanted Tissue-Engineered Human Limbal Epithelial Cells. Tissue Eng. Part A 2011, 17, 741–750. [Google Scholar] [CrossRef]
- Crespo-Moral, M.; García-Posadas, L.; López-García, A.; Diebold, Y. Histological and Immunohistochemical Characterization of the Porcine Ocular Surface. PLoS ONE 2020, 15, e0227732. [Google Scholar] [CrossRef]
- A Validated Porcine Corneal Organ Culture Model to Study the Limbal Response to Corneal Epithelial Injury—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/32417262/ (accessed on 13 April 2023).
- Menduni, F.; Davies, L.N.; Madrid-Costa, D.; Fratini, A.; Wolffsohn, J.S. Characterisation of the Porcine Eyeball as an In-Vitro Model for Dry Eye. Contact Lens Anterior Eye 2018, 41, 13–17. [Google Scholar] [CrossRef]
- Patruno, M.; Perazzi, A.; Martinello, T.; Blaseotto, A.; Di Iorio, E.; Iacopetti, I. Morphological Description of Limbal Epithelium: Searching for Stem Cells Crypts in the Dog, Cat, Pig, Cow, Sheep and Horse. Vet. Res. Commun. 2017, 41, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Kitazawa, K.; Hikichi, T.; Nakamura, T.; Sotozono, C.; Kinoshita, S.; Masui, S. PAX6 Regulates Human Corneal Epithelium Cell Identity. Exp. Eye Res. 2017, 154, 30–38. [Google Scholar] [CrossRef]
- Sunny, S.S.; Lachova, J.; Dupacova, N.; Kozmik, Z. Multiple Roles of Pax6 in Postnatal Cornea Development. Dev. Biol. 2022, 491, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, Y.-T.; Hayashida, Y.; Blanco, G.; Kheirkah, A.; He, H.; Chen, S.-Y.; Liu, C.-Y.; Tseng, S.C.G. Down-Regulation of Pax6 Is Associated with Abnormal Differentiation of Corneal Epithelial Cells in Severe Ocular Surface Diseases. J. Pathol. 2008, 214, 114–122. [Google Scholar] [CrossRef]
- Polisetti, N.; Schlunck, G.; Reinhard, T. PAX6 Expression Patterns in the Adult Human Limbal Stem Cell Niche. Cells 2023, 12, 400. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, B.B.; Tighe, P.J.; Mohammed, I.; Yeung, A.M.; Powe, D.G.; Hopkinson, A.; Shanmuganathan, V.A.; Dua, H.S. Comparative Transcriptional Profiling of the Limbal Epithelial Crypt Demonstrates Its Putative Stem Cell Niche Characteristics. BMC Genom. 2010, 11, 526. [Google Scholar] [CrossRef]
- Ramirez-Miranda, A.; Nakatsu, M.N.; Zarei-Ghanavati, S.; Nguyen, C.V.; Deng, S.X. Keratin 13 Is a More Specific Marker of Conjunctival Epithelium than Keratin 19. Mol. Vis. 2011, 17, 1652–1661. [Google Scholar] [PubMed]
- Ligocki, A.J.; Fury, W.; Gutierrez, C.; Adler, C.; Yang, T.; Ni, M.; Bai, Y.; Wei, Y.; Lehmann, G.L.; Romano, C. Molecular Characteristics and Spatial Distribution of Adult Human Corneal Cell Subtypes. Sci. Rep. 2021, 11, 16323. [Google Scholar] [CrossRef]
- Merjava, S.; Neuwirth, A.; Tanzerova, M.; Jirsova, K. The Spectrum of Cytokeratins Expressed in the Adult Human Cornea, Limbus and Perilimbal Conjunctiva. Histol. Histopathol. 2011, 26, 323–331. [Google Scholar] [CrossRef]
- Pellegrini, G.; Dellambra, E.; Golisano, O.; Martinelli, E.; Fantozzi, I.; Bondanza, S.; Ponzin, D.; McKeon, F.; De Luca, M. P63 Identifies Keratinocyte Stem Cells. Proc. Natl. Acad. Sci. USA 2001, 98, 3156–3161. [Google Scholar] [CrossRef] [PubMed]
- Ramaesh, T.; Collinson, J.M.; Ramaesh, K.; Kaufman, M.H.; West, J.D.; Dhillon, B. Corneal Abnormalities in Pax6+/- Small Eye Mice Mimic Human Aniridia-Related Keratopathy. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1871–1878. [Google Scholar] [CrossRef]
- Senoo, M.; Pinto, F.; Crum, C.P.; McKeon, F. P63 Is Essential for the Proliferative Potential of Stem Cells in Stratified Epithelia. Cell 2007, 129, 523–536. [Google Scholar] [CrossRef]
- Polisetti, N.; Sharaf, L.; Martin, G.; Schlunck, G.; Reinhard, T. P-Cadherin Is Expressed by Epithelial Progenitor Cells and Melanocytes in the Human Corneal Limbus. Cells 2022, 11, 1975. [Google Scholar] [CrossRef] [PubMed]
- Polisetti, N.; Zenkel, M.; Menzel-Severing, J.; Kruse, F.E.; Schlötzer-Schrehardt, U. Cell Adhesion Molecules and Stem Cell-Niche-Interactions in the Limbal Stem Cell Niche. Stem Cells 2016, 34, 203–219. [Google Scholar] [CrossRef]
- Polisetti, N. (University of Freiburg, Freiburg, Germany) N-Cadherin Staining on Porcine Limbal and Conjunctival Basal Epithelium. 2023. [Google Scholar]
- Morrison, S.J.; Spradling, A.C. Stem Cells and Niches: Mechanisms That Promote Stem Cell Maintenance throughout Life. Cell 2008, 132, 598–611. [Google Scholar] [CrossRef]
- Ljubimov, A.V.; Burgeson, R.E.; Butkowski, R.J.; Michael, A.F.; Sun, T.T.; Kenney, M.C. Human Corneal Basement Membrane Heterogeneity: Topographical Differences in the Expression of Type IV Collagen and Laminin Isoforms. Lab. Investig. 1995, 72, 461–473. [Google Scholar]
- Schlötzer-Schrehardt, U.; Dietrich, T.; Saito, K.; Sorokin, L.; Sasaki, T.; Paulsson, M.; Kruse, F.E. Characterization of Extracellular Matrix Components in the Limbal Epithelial Stem Cell Compartment. Exp. Eye Res. 2007, 85, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Mei, H.; Gonzalez, S.; Deng, S.X. Extracellular Matrix Is an Important Component of Limbal Stem Cell Niche. J. Funct. Biomater. 2012, 3, 879–894. [Google Scholar] [CrossRef] [PubMed]
- Schlötzer-Schrehardt, U.; Latta, L.; Gießl, A.; Zenkel, M.; Fries, F.N.; Käsmann-Kellner, B.; Kruse, F.E.; Seitz, B. Dysfunction of the Limbal Epithelial Stem Cell Niche in Aniridia-Associated Keratopathy. Ocul. Surf. 2021, 21, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Polisetti, N. (University of Freiburg, Freiburg, Germany) Tenascin C Staining on Anterior Limbus and Conjunctiva. 2023. [Google Scholar]
- Polisetti, N. (University of Freiburg, Freiburg, Germany) Laminin Alpha 3 and Laminin Alpha 5 Staining on Porcine Ocular Surface. 2023. [Google Scholar]
- Moore, C.J.; Winder, S.J. Dystroglycan Versatility in Cell Adhesion: A Tale of Multiple Motifs. Cell Commun. Signal. 2010, 8, 3. [Google Scholar] [CrossRef]
- Ordonez, P.; Di Girolamo, N. Limbal Epithelial Stem Cells: Role of the Niche Microenvironment. Stem Cells 2012, 30, 100–107. [Google Scholar] [CrossRef]
- Ellis, S.J.; Tanentzapf, G. Integrin-Mediated Adhesion and Stem-Cell-Niche Interactions. Cell Tissue Res. 2010, 339, 121–130. [Google Scholar] [CrossRef]
- Li, W.; Hayashida, Y.; Chen, Y.-T.; Tseng, S.C.G. Niche Regulation of Corneal Epithelial Stem Cells at the Limbus. Cell Res. 2007, 17, 26–36. [Google Scholar] [CrossRef]
- Polisetti, N.; Gießl, A.; Li, S.; Sorokin, L.; Kruse, F.E.; Schlötzer-Schrehardt, U. Laminin-511-E8 Promotes Efficient in Vitro Expansion of Human Limbal Melanocytes. Sci. Rep. 2020, 10, 11074. [Google Scholar] [CrossRef]
- Polisetti, N.; Gießl, A.; Zenkel, M.; Heger, L.; Dudziak, D.; Naschberger, E.; Stich, L.; Steinkasserer, A.; Kruse, F.E.; Schlötzer-Schrehardt, U. Melanocytes as Emerging Key Players in Niche Regulation of Limbal Epithelial Stem Cells. Ocul. Surf. 2021, 22, 172–189. [Google Scholar] [CrossRef]
- Amin, S.; Jalilian, E.; Katz, E.; Frank, C.; Yazdanpanah, G.; Guaiquil, V.H.; Rosenblatt, M.I.; Djalilian, A.R. The Limbal Niche and Regenerative Strategies. Vision 2021, 5, 43. [Google Scholar] [CrossRef]
- Fernández-Pérez, J.; Binner, M.; Werner, C.; Bray, L.J. Limbal Stromal Cells Derived from Porcine Tissue Demonstrate Mesenchymal Characteristics in Vitro. Sci. Rep. 2017, 7, 6377. [Google Scholar] [CrossRef]
- Zhu, S.N.; Nölle, B.; Duncker, G. Expression of Adhesion Molecule CD44 on Human Corneas. Br. J. Ophthalmol. 1997, 81, 80–84. [Google Scholar] [CrossRef]
- Polisetti, N.; Sharaf, L.; Schlötzer-Schrehardt, U.; Schlunck, G.; Reinhard, T. Efficient Isolation and Functional Characterization of Niche Cells from Human Corneal Limbus. Int. J. Mol. Sci. 2022, 23, 2750. [Google Scholar] [CrossRef]
- Debeer, S.; Le Luduec, J.-B.; Kaiserlian, D.; Laurent, P.; Nicolas, J.-F.; Dubois, B.; Kanitakis, J. Comparative Histology and Immunohistochemistry of Porcine versus Human Skin. Eur. J. Dermatol. 2013, 23, 456–466. [Google Scholar] [CrossRef]
- Gillette, T.E.; Chandler, J.W.; Greiner, J.V. Langerhans Cells of the Ocular Surface. Ophthalmology 1982, 89, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Song, P.I.; Abraham, T.A.; Park, Y.; Zivony, A.S.; Harten, B.; Edelhauser, H.F.; Ward, S.L.; Armstrong, C.A.; Ansel, J.C. The Expression of Functional LPS Receptor Proteins CD14 and Toll-like Receptor 4 in Human Corneal Cells. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2867–2877. [Google Scholar]
- Polisetti, N. (University of Freiburg, Freiburg, Germany) Tyrosinase Related Protein 1 Expression on Porcine Ocular Surface. 2023. [Google Scholar]
- Polisetti, N. (University of Freiburg, Freiburg, Germany) Melanocyte Markers Expression on Achen Mini Pigs Ocular Surface. 2023. [Google Scholar]
- Polisetti, N.; Schmid, A.; Schlötzer-Schrehardt, U.; Maier, P.; Lang, S.J.; Steinberg, T.; Schlunck, G.; Reinhard, T. A Decellularized Human Corneal Scaffold for Anterior Corneal Surface Reconstruction. Sci. Rep. 2021, 11, 2992. [Google Scholar] [CrossRef] [PubMed]
- Polisetti, N.; Roschinski, B.; Schlötzer-Schrehardt, U.; Maier, P.; Schlunck, G.; Reinhard, T. A Decellularized Human Limbal Scaffold for Limbal Stem Cell Niche Reconstruction. Int. J. Mol. Sci. 2021, 22, 10067. [Google Scholar] [CrossRef]
- Polisetti, N.; Rapp, J.; Liang, P.; Dettmer-Monaco, V.; Bucher, F.; Pruszak, J.; Schlötzer-Schrehardt, U.; Cathomen, T.; Schlunck, G.; Reinhard, T. Transcriptomic Landscape and Functional Characterization of Human Induced Pluripotent Stem Cell-Derived Limbal Epithelial Progenitor Cells. Cells 2022, 11, 3752. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cornea | Limbus | Conjunctiva | ||||
|---|---|---|---|---|---|---|
| Basal | Suprabasal | Basal | Suprabasal | Basal | Suprabasal | |
| CK3 | ++ | ++ | − | ++ | − | − |
| CK12 | ++ | ++ | − | ++ | − | − |
| CK13 | − | − | − | (+) or + | − | ++ |
| CK14 | − | − | ++ | - | ++ | − |
| CK15 | − | − | ++ | - | ++ | − |
| CK17/19 | − | − | ++ | - | ++ | + |
| E-cadherin | + | ++ | − or + | ++ | − or + | ++ |
| Epithelial keratins (Pan-CK) | ++ | ++ | ++ | ++ | ++ | ++ |
| Integrin α3 | + | (+) | + | (+) | + | + |
| Integrin α6 | ++ | − | ++ | − | ++ | − |
| p63α | (+) | − | ++ | − | ++ | − |
| Pax6 | ++ | ++ | ++ | ++ | ++ | ++ |
| P-cadherin | − | − | + | − | + | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polisetti, N.; Martin, G.; Cristina Schmitz, H.R.; Schlötzer-Schrehardt, U.; Schlunck, G.; Reinhard, T. Characterization of Porcine Ocular Surface Epithelial Microenvironment. Int. J. Mol. Sci. 2023, 24, 7543. https://doi.org/10.3390/ijms24087543
Polisetti N, Martin G, Cristina Schmitz HR, Schlötzer-Schrehardt U, Schlunck G, Reinhard T. Characterization of Porcine Ocular Surface Epithelial Microenvironment. International Journal of Molecular Sciences. 2023; 24(8):7543. https://doi.org/10.3390/ijms24087543
Chicago/Turabian StylePolisetti, Naresh, Gottfried Martin, Heidi R. Cristina Schmitz, Ursula Schlötzer-Schrehardt, Günther Schlunck, and Thomas Reinhard. 2023. "Characterization of Porcine Ocular Surface Epithelial Microenvironment" International Journal of Molecular Sciences 24, no. 8: 7543. https://doi.org/10.3390/ijms24087543
APA StylePolisetti, N., Martin, G., Cristina Schmitz, H. R., Schlötzer-Schrehardt, U., Schlunck, G., & Reinhard, T. (2023). Characterization of Porcine Ocular Surface Epithelial Microenvironment. International Journal of Molecular Sciences, 24(8), 7543. https://doi.org/10.3390/ijms24087543