
Role of Sex Hormones in Prevalent Kidney Diseases

 , and
, and 
Abstract
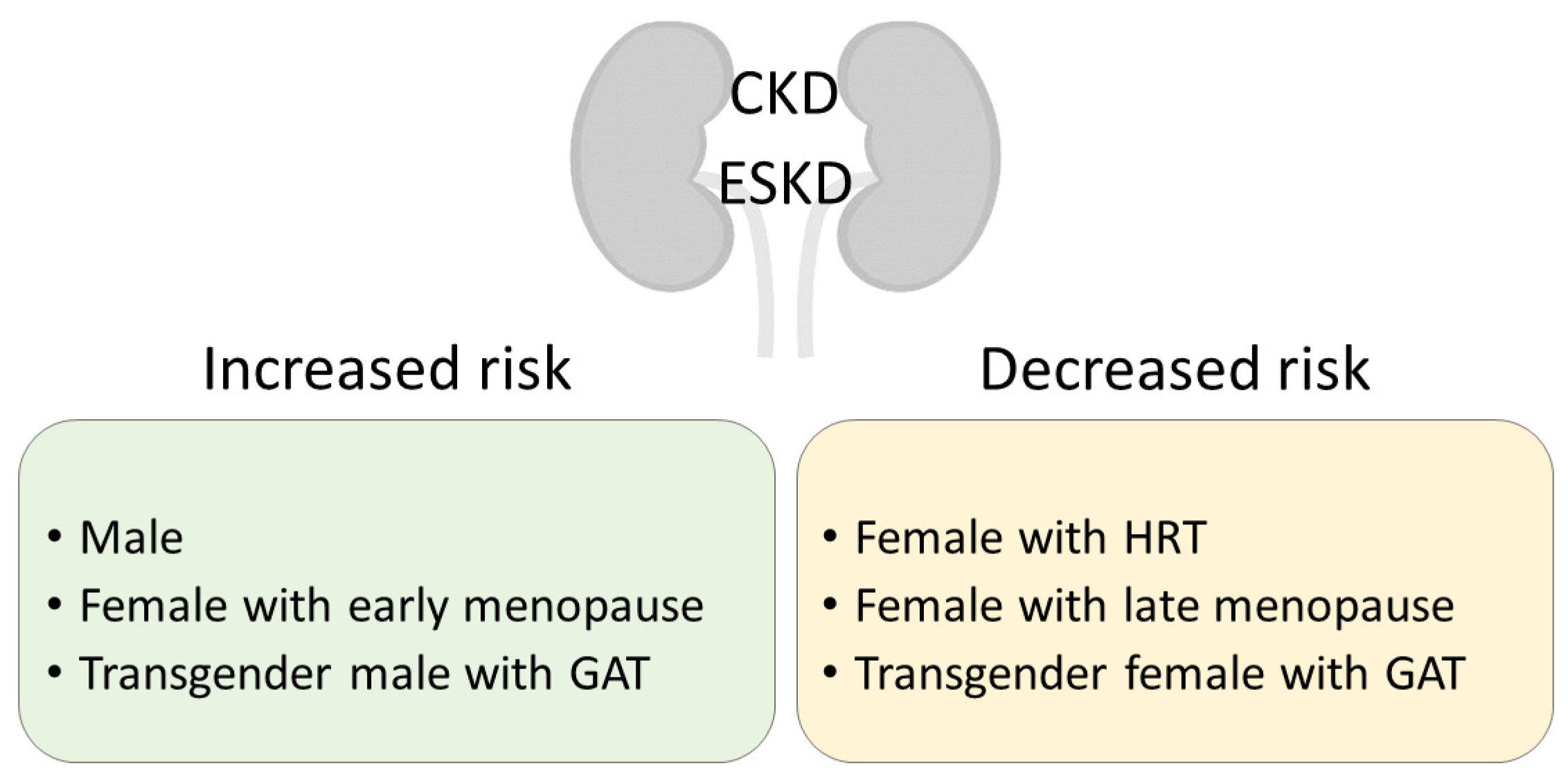
:1. Sex Disparities in CKD
2. Sex Hormones and Sex Hormone Receptors
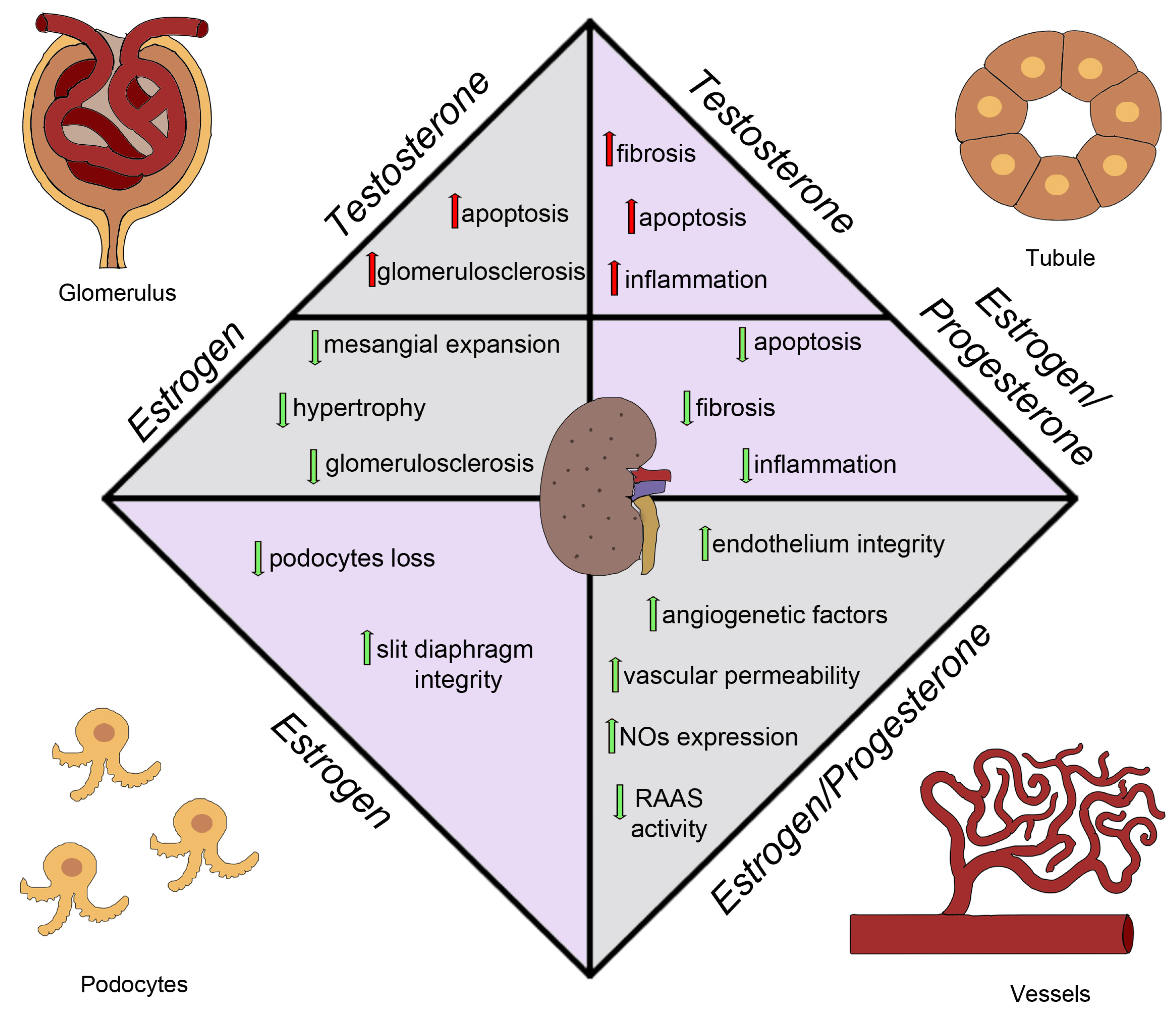
3. Experimental Models
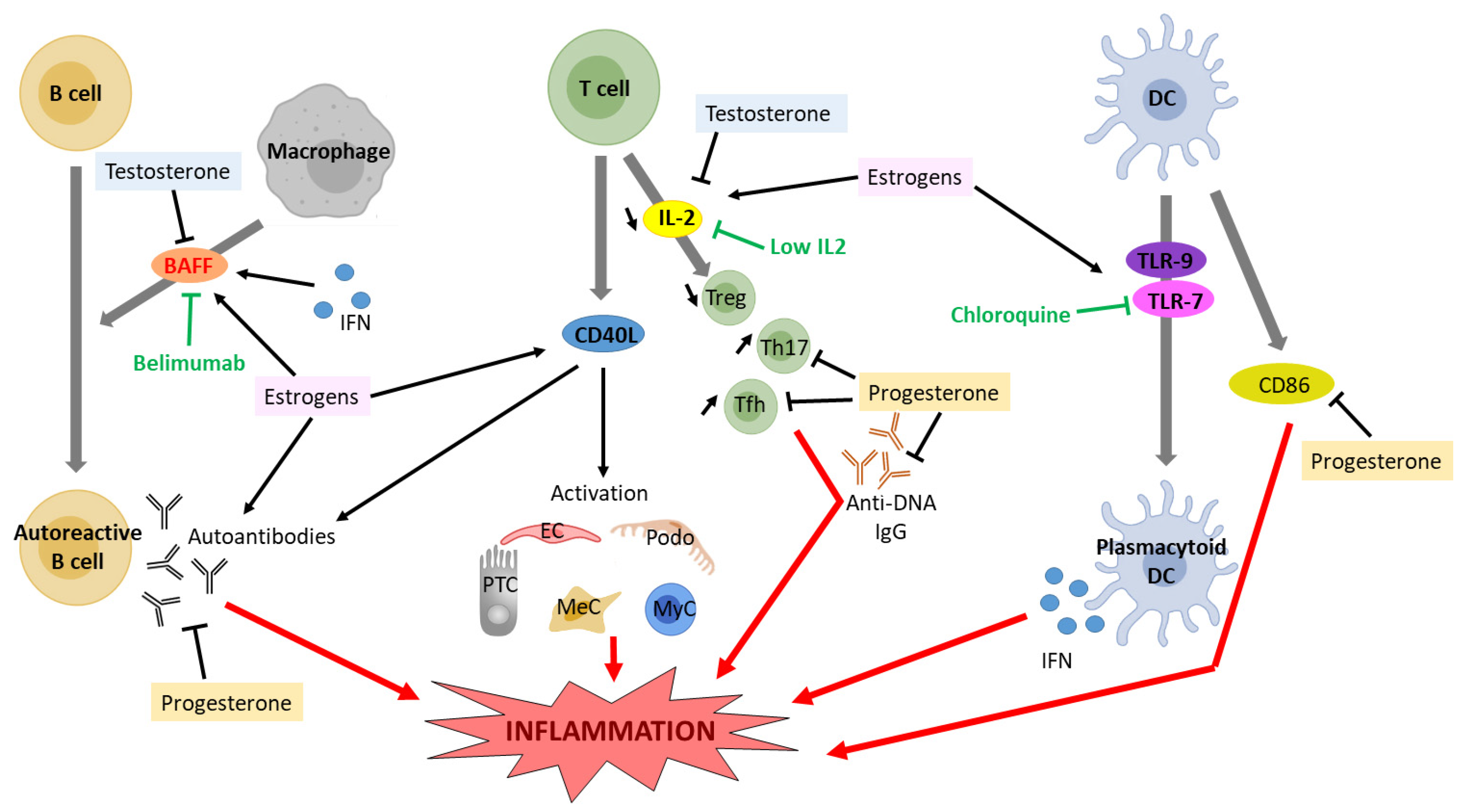
4. Lupus Nephritis
5. Diabetic Kidney Disease
6. IgA Nephropathy
7. Autosomal Dominant Polycystic Kidney Disease
8. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kovesdy, C.P. Epidemiology of chronic kidney disease: An update 2022. Kidney Int. Suppl. 2022, 12, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Foreman, K.J.; Marquez, N.; Dolgert, A.; Fukutaki, K.; Fullman, N.; McGaughey, M.; Pletcher, M.A.; Smith, A.E.; Tang, K.; Yuan, C.W.; et al. Forecasting life expectancy, years of life lost, and all-cause and cause-specific mortality for 250 causes of death: Reference and alternative scenarios for 2016–40 for 195 countries and territories. Lancet 2018, 392, 2052–2090. [Google Scholar] [CrossRef] [PubMed]
- Ke, C.; Liang, J.; Liu, M.; Liu, S.; Wang, C. Burden of chronic kidney disease and its risk-attributable burden in 137 low- and middle-income countries, 1990–2019: Results from the global burden of disease study 2019. BMC Nephrol. 2022, 23, 17. [Google Scholar] [CrossRef]
- Mills, K.T.; Xu, Y.; Zhang, W.; Bundy, J.D.; Chen, C.S.; Kelly, T.N.; Chen, J.; He, J. A systematic analysis of worldwide population-based data on the global burden of chronic kidney disease in 2010. Kidney Int. 2015, 88, 950–957. [Google Scholar] [CrossRef]
- Minutolo, R.; Gabbai, F.B.; Chiodini, P.; Provenzano, M.; Borrelli, S.; Garofalo, C.; Bellizzi, V.; Russo, D.; Conte, G.; De Nicola, L.; et al. Sex Differences in the Progression of CKD Among Older Patients: Pooled Analysis of 4 Cohort Studies. Am. J. Kidney Dis. 2020, 75, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Neugarten, J.; Golestaneh, L. Influence of Sex on the Progression of Chronic Kidney Disease. Mayo Clin. Proc. 2019, 94, 1339–1356. [Google Scholar] [CrossRef]
- Hannan, M.; Ansari, S.; Meza, N.; Anderson, A.H.; Srivastava, A.; Waikar, S.; Charleston, J.; Weir, M.R.; Taliercio, J.; Horwitz, E.; et al. Risk Factors for CKD Progression: Overview of Findings from the CRIC Study. Clin. J. Am. Soc. Nephrol. 2021, 16, 648–659. [Google Scholar] [CrossRef]
- Ricardo, A.C.; Yang, W.; Sha, D.; Appel, L.J.; Chen, J.; Krousel-Wood, M.; Manoharan, A.; Steigerwalt, S.; Wright, J.; Rahman, M.; et al. Sex-Related Disparities in CKD Progression. J. Am. Soc. Nephrol. 2019, 30, 137–146. [Google Scholar] [CrossRef]
- Yu, M.; Ryu, D.R.; Kim, S.J.; Choi, K.B.; Kang, D.H. Clinical implication of metabolic syndrome on chronic kidney disease depends on gender and menopausal status: Results from the Korean National Health and Nutrition Examination Survey. Nephrol. Dial. Transplant. 2010, 25, 469–477. [Google Scholar] [CrossRef]
- Qian, D.; Wang, Z.F.; Cheng, Y.C.; Luo, R.; Ge, S.W.; Xu, G. Early Menopause May Associate With a Higher Risk of CKD and All-Cause Mortality in Postmenopausal Women: An Analysis of NHANES, 1999–2014. Front. Med. 2022, 9, 823835. [Google Scholar] [CrossRef]
- Kattah, A.G.; Smith, C.Y.; Gazzuola Rocca, L.; Grossardt, B.R.; Garovic, V.D.; Rocca, W.A. CKD in Patients with Bilateral Oophorectomy. Clin. J. Am. Soc. Nephrol. 2018, 13, 1649–1658. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.C.; Jhee, J.H.; Joo, Y.S.; Lee, S.M.; Nam, K.H.; Yun, H.R.; Han, S.H.; Yoo, T.H.; Kang, S.W.; Park, J.T. Association of Reproductive Lifespan Duration and Chronic Kidney Disease in Postmenopausal Women. Mayo Clin. Proc. 2020, 95, 2621–2632. [Google Scholar] [CrossRef]
- Noh, J.H.; Koo, H. Older menarche age and short reproductive period linked to chronic kidney disease risk. Medicine 2019, 98, e15511. [Google Scholar] [CrossRef]
- Amiri, M.; Rahmati, M.; Farahmand, M.; Azizi, F.; Tehrani, F.R. Age at natural menopause in women with a history of chronic diseases-A population-based cohort study. Maturitas 2022, 158, 16–24. [Google Scholar] [CrossRef]
- Rytz, C.L.; Kochaksaraei, G.S.; Skeith, L.; Ronksley, P.E.; Dumanski, S.M.; Robert, M.; Ahmed, S.B. Menstrual Abnormalities and Reproductive Lifespan in Females with CKD: A Systematic Review and Meta-Analysis. Clin. J. Am. Soc. Nephrol. 2022, 17, 1742–1753. [Google Scholar] [CrossRef]
- Vellanki, K.; Hou, S. Menopause in CKD. Am. J. Kidney Dis. 2018, 71, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Kim, J.M. Klotho and Postmenopausal Hormone Replacement Therapy in Women with Chronic Kidney Disease. J. Menopausal Med. 2018, 24, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Marjoribanks, J.; Farquhar, C.; Roberts, H.; Lethaby, A. Long term hormone therapy for perimenopausal and postmenopausal women. Cochrane Database Syst. Rev. 2017, 1, CD004143. [Google Scholar] [CrossRef]
- Krupka, E.; Curtis, S.; Ferguson, T.; Whitlock, R.; Askin, N.; Millar, A.C.; Dahl, M.; Fung, R.; Ahmed, S.B.; Tangri, N.; et al. The Effect of Gender-Affirming Hormone Therapy on Measures of Kidney Function: A Systematic Review and Meta-Analysis. Clin. J. Am. Soc. Nephrol. 2022, 17, 1305–1315. [Google Scholar] [CrossRef]
- Hylander, B.; Lehtihet, M. Testosterone and gonadotropins but not SHBG vary with CKD stages in young and middle aged men. Basic Clin. Androl. 2015, 25, 9. [Google Scholar] [CrossRef]
- Dhindsa, S.; Reddy, A.; Karam, J.S.; Bilkis, S.; Chaurasia, A.; Mehta, A.; Raja, K.P.; Batra, M.; Dandona, P. Prevalence of subnormal testosterone concentrations in men with type 2 diabetes and chronic kidney disease. Eur. J. Endocrinol. 2015, 173, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, M.; Hoermann, R.; Ng Tang Fui, M.; Zajac, J.D.; Ierino, F.L.; Roberts, M.A. Sex steroids levels in chronic kidney disease and kidney transplant recipients: Associations with disease severity and prediction of mortality. Clin. Endocrinol. 2015, 82, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, W.; Kubber, H.; Dolff, S.; Benson, S.; Fuhrer, D.; Tan, S. Rapid recovery of hypogonadism in male patients with end stage renal disease after renal transplantation. Endocrine 2018, 60, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Lesniak, K.; Rymarz, A.; Sobol, M.; Dymus, J.; Wozniak-Kosek, A.; Niemczyk, S. Testosterone Deficiency and Nutritional Parameters as Predictors of All-Cause Mortality among Male Dialysis Patients. Nutrients 2022, 14, 4461. [Google Scholar] [CrossRef] [PubMed]
- Rymarz, A.; Skiba, R.; Matyjek, A.; Bartoszewicz, Z.; Niemczyk, S. Free testosterone levels and their association with body composition in women with chronic kidney disease. Pol. Merkur. Lek. 2021, 49, 329–333. [Google Scholar]
- Sikora-Grabka, E.; Adamczak, M.; Kuczera, P.; Wiecek, A. Serum sex hormones concentrations in young women in the early period after successful kidney transplantation. Endokrynol. Pol. 2018, 69, 150–155. [Google Scholar] [CrossRef]
- Lu, H.; Lei, X.; Klaassen, C. Gender differences in renal nuclear receptors and aryl hydrocarbon receptor in 5/6 nephrectomized rats. Kidney Int. 2006, 70, 1920–1928. [Google Scholar] [CrossRef]
- Fanelli, C.; Delle, H.; Cavaglieri, R.C.; Dominguez, W.V.; Noronha, I.L. Gender Differences in the Progression of Experimental Chronic Kidney Disease Induced by Chronic Nitric Oxide Inhibition. BioMed Res. Int. 2017, 2017, 2159739. [Google Scholar] [CrossRef]
- Antus, B.; Yao, Y.; Song, E.; Liu, S.; Lutz, J.; Heemann, U. Opposite effects of testosterone and estrogens on chronic allograft nephropathy. Transpl. Int. 2002, 15, 494–501. [Google Scholar] [CrossRef]
- Gross, M.L.; Adamczak, M.; Rabe, T.; Harbi, N.A.; Krtil, J.; Koch, A.; Hamar, P.; Amann, K.; Ritz, E. Beneficial Effects of Estrogens on Indices of Renal Damage in Uninephrectomized SHRsp Rats. J. Am. Soc. Nephrol. 2004, 15, 348–358. [Google Scholar] [CrossRef]
- Mercantepe, T.; Unal, D.; Selli, J.; Mercantepe, F.; Unal, B.; Karabiyik, T.N. Protective effects of estrogen and bortezomib in kidney tissue of post-menopausal rats: An ultrastructural study. Ren. Fail. 2016, 38, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Kasimay, O.; Sener, G.; Cakir, B.; Yuksel, M.; Cetinel, S.; Contuk, G.; Yegen, B.C. Estrogen protects against oxidative multiorgan damage in rats with chronic renal failure. Ren. Fail. 2009, 31, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Hutchens, M.P.; Fujiyoshi, T.; Komers, R.; Herson, P.S.; Anderson, S. Estrogen protects renal endothelial barrier function from ischemia-reperfusion in vitro and in vivo. Am. J. Physiol. Ren. Physiol. 2012, 303, F377–F385. [Google Scholar] [CrossRef]
- Diwan, V.; Small, D.; Kauter, K.; Gobe, G.C.; Brown, L. Gender differences in adenine-induced chronic kidney disease and cardiovascular complications in rats. Am. J. Physiol. Ren. Physiol. 2014, 307, F1169–F1178. [Google Scholar] [CrossRef]
- Maric, C.; Sandberg, K.; Hinojosa-Laborde, C. Glomerulosclerosis and tubulointerstitial fibrosis are attenuated with 17beta-estradiol in the aging Dahl salt sensitive rat. J. Am. Soc. Nephrol. 2004, 15, 1546–1556. [Google Scholar] [CrossRef] [PubMed]
- Anders, H.J.; Saxena, R.; Zhao, M.H.; Parodis, I.; Salmon, J.E.; Mohan, C. Lupus nephritis. Nat. Rev. Dis. Prim. 2020, 6, 7. [Google Scholar] [CrossRef]
- Shaharir, S.S.; Kadir, W.D.A.; Nordin, F.; Bakar, F.A.; Ting, M.W.H.; Jamil, A.; Mohd, R.; Wahab, A.A. Systemic lupus erythematosus among male patients in Malaysia: How are we different from other geographical regions? Lupus 2019, 28, 137–144. [Google Scholar] [CrossRef]
- Riveros Frutos, A.; Casas, I.; Rua-Figueroa, I.; Lopez-Longo, F.J.; Calvo-Alen, J.; Galindo, M.; Fernandez-Nebro, A.; Pego-Reigosa, J.M.; Olive Marques, A.; on behalf of the RELESSER Group, part of the Spanish Society of Rheumatology Systemic Autoimmune Diseases Study Group (EASSER). Systemic lupus erythematosus in Spanish males: A study of the Spanish Rheumatology Society Lupus Registry (RELESSER) cohort. Lupus 2017, 26, 698–706. [Google Scholar] [CrossRef]
- Peng, W.; Tang, Y.; Tan, L.; Qin, W. Clinicopathological study of male and female patients with lupus nephritis: A retrospective study. Int. Urol. Nephrol. 2018, 50, 313–320. [Google Scholar] [CrossRef]
- Okpechi, I.G.; Swanepoel, C.R.; Tiffin, N.; Duffield, M.; Rayner, B.L. Clinicopathological insights into lupus nephritis in South Africans: A study of 251 patients. Lupus 2012, 21, 1017–1024. [Google Scholar] [CrossRef]
- Ichinose, K.; Kitamura, M.; Sato, S.; Fujikawa, K.; Horai, Y.; Matsuoka, N.; Tsuboi, M.; Nonaka, F.; Shimizu, T.; Fukui, S.; et al. Factors predictive of long-term mortality in lupus nephritis: A multicenter retrospective study of a Japanese cohort. Lupus 2019, 28, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Resende, A.L.; Titan, S.M.; Barros, R.T.; Woronik, V. Worse renal outcome of lupus nephritis in male patients: A case-control study. Lupus 2011, 20, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Yoo, K.D.; Oh, Y.K.; Kim, D.K.; Oh, K.H.; Joo, K.W.; Kim, Y.S.; Ahn, C.; Han, J.S.; Lim, C.S. Predictors of Relapse in Adult-Onset Nephrotic Minimal Change Disease. Medicine 2016, 95, e3179. [Google Scholar] [CrossRef] [PubMed]
- Neugarten, J.; Acharya, A.; Silbiger, S.R. Effect of gender on the progression of nondiabetic renal disease: A meta-analysis. J. Am. Soc. Nephrol. 2000, 11, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Maric-Bilkan, C. Sex Differences in Diabetic Kidney Disease. Mayo Clin. Proc. 2020, 95, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Shepard, B.D. Sex differences in diabetes and kidney disease: Mechanisms and consequences. Am. J. Physiol. Ren. Physiol. 2019, 317, F456–F462. [Google Scholar] [CrossRef]
- Gall, M.A.; Hougaard, P.; Borch-Johnsen, K.; Parving, H.H. Risk factors for development of incipient and overt diabetic nephropathy in patients with non-insulin dependent diabetes mellitus: Prospective, observational study. BMJ 1997, 314, 783–788. [Google Scholar] [CrossRef]
- van Daalen, E.E.; Jennette, J.C.; McAdoo, S.P.; Pusey, C.D.; Alba, M.A.; Poulton, C.J.; Wolterbeek, R.; Nguyen, T.Q.; Goldschmeding, R.; Alchi, B.; et al. Predicting Outcome in Patients with Anti-GBM Glomerulonephritis. Clin. J. Am. Soc. Nephrol. 2018, 13, 63–72. [Google Scholar] [CrossRef]
- Nakamura, I.; Iwase, H.; Arai, K.; Nagai, Y.; Toma, K.; Katsumata, T.; Hiki, Y.; Kokubo, T.; Sano, T.; Kobayashi, Y. Detection of gender difference and epitope specificity of IgG antibody activity against IgA1 hinge portion in IgA nephropathy patients by using synthetic hinge peptide and glycopeptide probes. Nephrology 2004, 9, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Tan, X.; Zhou, Q.; Ai, Z.; Liu, W.; Chen, W.; Yu, X.; Yang, Q. Gender-related differences in clinicopathological characteristics and renal outcomes of Chinese patients with IgA nephropathy. BMC Nephrol. 2018, 19, 31. [Google Scholar] [CrossRef]
- Feng, Y.; Su, Y.; Ma, C.; Jing, Z.; Yang, X.; Zhang, D.; Xie, M.; Li, W.; Wei, J. 3′UTR variants of TNS3, PHLDB1, NTN4, and GNG2 genes are associated with IgA nephropathy risk in Chinese Han population. Int. Immunopharmacol. 2019, 71, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Cattran, D.C.; Reich, H.N.; Beanlands, H.J.; Miller, J.A.; Scholey, J.W.; Troyanov, S.; for the Genes, Gender and Glomerulonephritis Group. The impact of sex in primary glomerulonephritis. Nephrol. Dial. Transplant. 2008, 23, 2247–2253. [Google Scholar] [CrossRef] [PubMed]
- Fenton, A.; Smith, S.W.; Hewins, P. Adult minimal-change disease: Observational data from a UK centre on patient characteristics, therapies, and outcomes. BMC Nephrol. 2018, 19, 207. [Google Scholar] [CrossRef] [PubMed]
- Troyanov, S.; Wall, C.A.; Miller, J.A.; Scholey, J.W.; Cattran, D.C.; for the Toronto Glomerulonephritis Registry Group. Focal and segmental glomerulosclerosis: Definition and relevance of a partial remission. J. Am. Soc. Nephrol. 2005, 16, 1061–1068. [Google Scholar] [CrossRef]
- Johnson, A.M.; Gabow, P.A. Identification of patients with autosomal dominant polycystic kidney disease at highest risk for end-stage renal disease. J. Am. Soc. Nephrol. 1997, 8, 1560–1567. [Google Scholar] [CrossRef]
- Tampe, D.; Korsten, P.; Strobel, P.; Hakroush, S.; Tampe, B. Comprehensive Analysis of Sex Differences at Disease Manifestation in ANCA-Associated Glomerulonephritis. Front. Immunol. 2021, 12, 736638. [Google Scholar] [CrossRef]
- Scott, J.; Canepa, C.; Buettner, A.; Ryan, L.; Moloney, B.; Cormican, S.; Walsh, C.; White, A.; Salama, A.D.; Little, M.A. A cohort study to investigate sex-specific differences in ANCA-associated glomerulonephritis outcomes. Sci. Rep. 2021, 11, 13080. [Google Scholar] [CrossRef]
- Reynolds, W.F.; Stegeman, C.A.; Tervaert, J.W. −463 G/A myeloperoxidase promoter polymorphism is associated with clinical manifestations and the course of disease in MPO-ANCA-associated vasculitis. Clin. Immunol. 2002, 103, 154–160. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef]
- Holst, J.P.; Soldin, O.P.; Guo, T.; Soldin, S.J. Steroid hormones: Relevance and measurement in the clinical laboratory. Clin. Lab. Med. 2004, 24, 105–118. [Google Scholar] [CrossRef]
- Boonyaratanakornkit, V.; Edwards, D.P. Receptor mechanisms mediating non-genomic actions of sex steroids. Semin. Reprod. Med. 2007, 25, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.R. Integration of the extranuclear and nuclear actions of estrogen. Mol. Endocrinol. 2005, 19, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Valentina, C.; John, R.B.; Patrizia, P. Sex steroid hormone receptors, their ligands, and nuclear and non-nuclear pathways. AIMS Mol. Sci. 2015, 2, 294–310. [Google Scholar] [CrossRef]
- Ma, H.Y.; Chen, S.; Du, Y. Estrogen and estrogen receptors in kidney diseases. Ren. Fail. 2021, 43, 619–642. [Google Scholar] [CrossRef]
- Yasar, P.; Ayaz, G.; User, S.D.; Gupur, G.; Muyan, M. Molecular mechanism of estrogen-estrogen receptor signaling. Reprod. Med. Biol. 2017, 16, 4–20. [Google Scholar] [CrossRef]
- Davey, R.A.; Grossmann, M. Androgen Receptor Structure, Function and Biology: From Bench to Bedside. Clin. Biochem. Rev. 2016, 37, 3–15. [Google Scholar]
- Jacobsen, B.M.; Horwitz, K.B. Progesterone receptors, their isoforms and progesterone regulated transcription. Mol. Cell Endocrinol. 2012, 357, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.K.O.; Seelig, D.M.; Sharkey, L.C.; Choi, W.S.V.; Abdelgawad, I.Y.; Zordoky, B.N. Sexual dimorphism of acute doxorubicin-induced nephrotoxicity in C57Bl/6 mice. PLoS ONE 2019, 14, e0212486. [Google Scholar] [CrossRef]
- Lee, V.W.; Harris, D.C. Adriamycin nephropathy: A model of focal segmental glomerulosclerosis. Nephrology 2011, 16, 30–38. [Google Scholar] [CrossRef]
- Nangaku, M.; Kitching, A.R.; Boor, P.; Fornoni, A.; Floege, J.; Coates, P.T.; Himmelfarb, J.; Lennon, R.; Anders, H.-J.; Humphreys, B.D.; et al. International Society of Nephrology first consensus guidance for preclinical animal studies in translational nephrology. Kidney Int. 2023. [Google Scholar] [CrossRef]
- Diaz Brinton, R. Minireview: Translational Animal Models of Human Menopause: Challenges and Emerging Opportunities. Endocrinology 2012, 153, 3571–3578. [Google Scholar] [CrossRef] [PubMed]
- Couse, J.F.; Korach, K.S. Estrogen receptor null mice: What have we learned and where will they lead us? Endocr. Rev. 1999, 20, 358–417. [Google Scholar] [CrossRef] [PubMed]
- Krege, J.H.; Hodgin, J.B.; Couse, J.F.; Enmark, E.; Warner, M.; Mahler, J.F.; Sar, M.; Korach, K.S.; Gustafsson, J.A.; Smithies, O. Generation and reproductive phenotypes of mice lacking estrogen receptor beta. Proc. Natl. Acad. Sci. USA 1998, 95, 15677–15682. [Google Scholar] [CrossRef] [PubMed]
- Lubahn, D.B.; Moyer, J.S.; Golding, T.S.; Couse, J.F.; Korach, K.S.; Smithies, O. Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc. Natl. Acad. Sci. USA 1993, 90, 11162–11166. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Yeh, S.; Lee, S.O.; Chang, T.M. Androgen receptor (AR) pathophysiological roles in androgen-related diseases in skin, bone/muscle, metabolic syndrome and neuron/immune systems: Lessons learned from mice lacking AR in specific cells. Nucl. Recept. Signal. 2013, 11, e001. [Google Scholar] [CrossRef]
- Humphreys, R.C.; Lydon, J.P.; O’Malley, B.W.; Rosen, J.M. Use of PRKO mice to study the role of progesterone in mammary gland development. J. Mammary Gland Biol. Neoplasia 1997, 2, 343–354. [Google Scholar] [CrossRef]
- Igarashi, P. Kidney-specific gene targeting. J. Am. Soc. Nephrol. 2004, 15, 2237–2239. [Google Scholar] [CrossRef]
- Kohan, D.E. Progress in gene targeting: Using mutant mice to study renal function and disease. Kidney Int. 2008, 74, 427–437. [Google Scholar] [CrossRef]
- Guo, Z.M.; Xu, K.; Yue, Y.; Huang, B.; Deng, X.Y.; Zhong, N.Q.; Hong, X.; Chen, X.G.; Xiao, D. Temporal control of Cre recombinase-mediated in vitro DNA recombination by Tet-on gene expression system. Acta Biochim. Biophys. Sin. 2005, 37, 133–138. [Google Scholar] [CrossRef]
- Shimshek, D.R.; Kim, J.; Hubner, M.R.; Spergel, D.J.; Buchholz, F.; Casanova, E.; Stewart, A.F.; Seeburg, P.H.; Sprengel, R. Codon-improved Cre recombinase (iCre) expression in the mouse. Genesis 2002, 32, 19–26. [Google Scholar] [CrossRef]
- Furth, P.A.; St Onge, L.; Boger, H.; Gruss, P.; Gossen, M.; Kistner, A.; Bujard, H.; Hennighausen, L. Temporal control of gene expression in transgenic mice by a tetracycline-responsive promoter. Proc. Natl. Acad. Sci. USA 1994, 91, 9302–9306. [Google Scholar] [CrossRef]
- Arnold, A.P.; Chen, X. What does the “four core genotypes” mouse model tell us about sex differences in the brain and other tissues? Front. Neuroendocrinol. 2009, 30, 1–9. [Google Scholar] [CrossRef]
- Lovell-Badge, R.; Robertson, E. XY female mice resulting from a heritable mutation in the primary testis-determining gene, Tdy. Development 1990, 109, 635–646. [Google Scholar] [CrossRef]
- Mahadevaiah, S.K.; Odorisio, T.; Elliott, D.J.; Rattigan, A.; Szot, M.; Laval, S.H.; Washburn, L.L.; McCarrey, J.R.; Cattanach, B.M.; Lovell-Badge, R.; et al. Mouse homologues of the human AZF candidate gene RBM are expressed in spermatogonia and spermatids, and map to a Y chromosome deletion interval associated with a high incidence of sperm abnormalities. Hum. Mol. Genet. 1998, 7, 715–727. [Google Scholar] [CrossRef]
- Bajema, I.M.; Wilhelmus, S.; Alpers, C.E.; Bruijn, J.A.; Colvin, R.B.; Cook, H.T.; D’Agati, V.D.; Ferrario, F.; Haas, M.; Jennette, J.C.; et al. Revision of the International Society of Nephrology/Renal Pathology Society classification for lupus nephritis: Clarification of definitions, and modified National Institutes of Health activity and chronicity indices. Kidney Int. 2018, 93, 789–796. [Google Scholar] [CrossRef]
- Weening, J.J.; D’Agati, V.D.; Schwartz, M.M.; Seshan, S.V.; Alpers, C.E.; Appel, G.B.; Balow, J.E.; Bruijn, J.A.; Cook, T.; Ferrario, F.; et al. The classification of glomerulonephritis in systemic lupus erythematosus revisited. Kidney Int. 2004, 65, 521–530. [Google Scholar] [CrossRef]
- Izmirly, P.M.; Ferucci, E.D.; Somers, E.C.; Wang, L.; Lim, S.S.; Drenkard, C.; Dall’Era, M.; McCune, W.J.; Gordon, C.; Helmick, C.; et al. Incidence rates of systemic lupus erythematosus in the USA: Estimates from a meta-analysis of the Centers for Disease Control and Prevention national lupus registries. Lupus Sci. Med. 2021, 8, e000614. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.R.W.; Drenkard, C.; Falasinnu, T.; Hoi, A.; Mak, A.; Kow, N.Y.; Svenungsson, E.; Peterson, J.; Clarke, A.E.; Ramsey-Goldman, R. Global epidemiology of systemic lupus erythematosus. Nat. Rev. Rheumatol. 2021, 17, 515–532. [Google Scholar] [CrossRef]
- Van Griensven, M.; Bergijk, E.C.; Baelde, J.J.; De Heer, E.; Bruijn, J.A. Differential effects of sex hormones on autoantibody production and proteinuria in chronic graft-versus-host disease-induced experimental lupus nephritis. Clin. Exp. Immunol. 1997, 107, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Treurniet, R.A.; Bergijk, E.C.; Baelde, J.J.; De Heer, E.; Hoedemaeker, P.J.; Bruijn, J.A. Gender-related influences on the development of chronic graft-versus-host disease-induced experimental lupus nephritis. Clin. Exp. Immunol. 1993, 91, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Kim, H.A.; Suh, C.H.; Jung, J.Y. Sex hormones affect the pathogenesis and clinical characteristics of systemic lupus erythematosus. Front. Med. 2022, 9, 906475. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, S.K.; Bermas, B.; Costenbader, K.H. Sexual disparities in the incidence and course of SLE and RA. Clin. Immunol. 2013, 149, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.J.; Wallace, D.J.; Ishimori, M.L.; Scofield, R.H.; Weisman, M.H. Review: Male systemic lupus erythematosus: A review of sex disparities in this disease. Lupus 2010, 19, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Pontes, L.T.; Camilo, D.T.; De Bortoli, M.R.; Santos, R.S.S.; Luchi, W.M. New-onset lupus nephritis after male-to-female sex reassignment surgery. Lupus 2018, 27, 2166–2169. [Google Scholar] [CrossRef]
- Bassi, N.; Luisetto, R.; Ghirardello, A.; Gatto, M.; Valente, M.; Della Barbera, M.; Nalotto, L.; Punzi, L.; Doria, A. 17-beta-estradiol affects BLyS serum levels and the nephritogenic autoantibody network accelerating glomerulonephritis in NZB/WF1 mice. Lupus 2015, 24, 382–391. [Google Scholar] [CrossRef]
- Feng, F.; Silvin, C.J.; Fiore, N.C.; Stoll, M.L.; Price, K.E.; Shanley, P.S.; Silverstone, A.E.; Gavalchin, J. 17beta-Estradiol (E-2) administration to male (NZB × SWR)F1 mice results in increased IdLNF1-reactive memory T-lymphocytes and accelerated glomerulonephritis. Lupus 2012, 21, 288–301. [Google Scholar] [CrossRef]
- Feng, F.; Nyland, J.; Banyai, M.; Tatum, A.; Silverstone, A.E.; Gavalchin, J. The induction of the lupus phenotype by estrogen is via an estrogen receptor-alpha-dependent pathway. Clin. Immunol. 2010, 134, 226–236. [Google Scholar] [CrossRef]
- Svenson, J.L.; EuDaly, J.; Ruiz, P.; Korach, K.S.; Gilkeson, G.S. Impact of estrogen receptor deficiency on disease expression in the NZM2410 lupus prone mouse. Clin. Immunol. 2008, 128, 259–268. [Google Scholar] [CrossRef]
- Dao, K.H.; Bermas, B.L. Systemic Lupus Erythematosus Management in Pregnancy. Int. J. Womens Health 2022, 14, 199–211. [Google Scholar] [CrossRef]
- Zenclussen, A.C.; Kokeny, G.; Thimm, O.; Sollwedel, A.; Godo, M.; Casalis, P.A.; Zenclussen, M.L.; Volk, H.D.; Hamar, P. Mechanisms behind flare of renal lupus during murine pregnancy. Reprod. Biomed. Online 2008, 17, 114–126. [Google Scholar] [CrossRef]
- Pakpoor, J.; Goldacre, R.; Goldacre, M.J. Associations between clinically diagnosed testicular hypofunction and systemic lupus erythematosus: A record linkage study. Clin. Rheumatol. 2018, 37, 559–562. [Google Scholar] [CrossRef]
- Gordon, C.; Wallace, D.J.; Shinada, S.; Kalunian, K.C.; Forbess, L.; Braunstein, G.D.; Weisman, M.H. Testosterone patches in the management of patients with mild/moderate systemic lupus erythematosus. Rheumatology 2008, 47, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Petri, M.A.; Mease, P.J.; Merrill, J.T.; Lahita, R.G.; Iannini, M.J.; Yocum, D.E.; Ginzler, E.M.; Katz, R.S.; Gluck, O.S.; Genovese, M.C.; et al. Effects of prasterone on disease activity and symptoms in women with active systemic lupus erythematosus. Arthritis Rheum. 2004, 50, 2858–2868. [Google Scholar] [CrossRef] [PubMed]
- van Vollenhoven, R.F.; Park, J.L.; Genovese, M.C.; West, J.P.; McGuire, J.L. A double-blind, placebo-controlled, clinical trial of dehydroepiandrosterone in severe systemic lupus erythematosus. Lupus 1999, 8, 181–187. [Google Scholar] [CrossRef]
- Lahita, R.G.; Cheng, C.Y.; Monder, C.; Bardin, C.W. Experience with 19-nortestosterone in the therapy of systemic lupus erythematosus: Worsened disease after treatment with 19-nortestosterone in men and lack of improvement in women. J. Rheumatol. 1992, 19, 547–555. [Google Scholar] [PubMed]
- Grygiel-Gorniak, B.; Puszczewicz, M.J. The influence of endogenous and exogenous sex hormones on systemic lupus erythematosus in pre- and postmenopausal women. Prz. Menopauzalny 2014, 13, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Recalde, G.; Moreno-Sosa, T.; Yudica, F.; Quintero, C.A.; Sanchez, M.B.; Jahn, G.A.; Kalergis, A.M.; Mackern-Oberti, J.P. Contribution of sex steroids and prolactin to the modulation of T and B cells during autoimmunity. Autoimmun. Rev. 2018, 17, 504–512. [Google Scholar] [CrossRef]
- Hughes, G.C.; Martin, D.; Zhang, K.; Hudkins, K.L.; Alpers, C.E.; Clark, E.A.; Elkon, K.B. Decrease in glomerulonephritis and Th1-associated autoantibody production after progesterone treatment in NZB/NZW mice. Arthritis Rheum. 2009, 60, 1775–1784. [Google Scholar] [CrossRef]
- Moulton, V.R. Sex Hormones in Acquired Immunity and Autoimmune Disease. Front. Immunol. 2018, 9, 2279. [Google Scholar] [CrossRef]
- Devarapu, S.K.; Lorenz, G.; Kulkarni, O.P.; Anders, H.J.; Mulay, S.R. Cellular and Molecular Mechanisms of Autoimmunity and Lupus Nephritis. Int. Rev. Cell Mol. Biol. 2017, 332, 43–154. [Google Scholar] [CrossRef]
- Kovats, S. Estrogen receptors regulate innate immune cells and signaling pathways. Cell. Immunol. 2015, 294, 63–69. [Google Scholar] [CrossRef]
- Kassi, E.; Moutsatsou, P. Estrogen receptor signaling and its relationship to cytokines in systemic lupus erythematosus. J. Biomed. Biotechnol. 2010, 2010, 317452. [Google Scholar] [CrossRef]
- Hill, L.; Jeganathan, V.; Chinnasamy, P.; Grimaldi, C.; Diamond, B. Differential roles of estrogen receptors alpha and beta in control of B-cell maturation and selection. Mol. Med. 2011, 17, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Panchanathan, R.; Choubey, D. Murine BAFF expression is up-regulated by estrogen and interferons: Implications for sex bias in the development of autoimmunity. Mol. Immunol. 2013, 53, 15–23. [Google Scholar] [CrossRef]
- Samy, E.; Wax, S.; Huard, B.; Hess, H.; Schneider, P. Targeting BAFF and APRIL in systemic lupus erythematosus and other antibody-associated diseases. Int. Rev. Immunol. 2017, 36, 3–19. [Google Scholar] [CrossRef]
- Rider, V.; Jones, S.; Evans, M.; Bassiri, H.; Afsar, Z.; Abdou, N.I. Estrogen increases CD40 ligand expression in T cells from women with systemic lupus erythematosus. J. Rheumatol. 2001, 28, 2644–2649. [Google Scholar] [PubMed]
- Shao, M.; He, J.; Zhang, R.; Zhang, X.; Yang, Y.; Li, C.; Liu, X.; Sun, X.; Li, Z. Interleukin-2 Deficiency Associated with Renal Impairment in Systemic Lupus Erythematosus. J. Interferon Cytokine Res. 2019, 39, 117–124. [Google Scholar] [CrossRef] [PubMed]
- McMurray, R.W.; Ndebele, K.; Hardy, K.J.; Jenkins, J.K. 17-β-ESTRADIOL SUPPRESSES IL-2 AND IL-2 RECEPTOR. Cytokine 2001, 14, 324–333. [Google Scholar] [CrossRef]
- Suzuki, N.; Suzuki, T.; Sakane, T. Hormones and lupus: Defective dehydroepiandrosterone activity induces impaired interleukin-2 activity of T lymphocytes in patients with systemic lupus erythematosus. Ann. Med. Interne 1996, 147, 248–252. [Google Scholar]
- Humrich, J.Y.; Cacoub, P.; Rosenzwajg, M.; Pitoiset, F.; Pham, H.P.; Guidoux, J.; Leroux, D.; Vazquez, T.; Riemekasten, G.; Smolen, J.S.; et al. Low-dose interleukin-2 therapy in active systemic lupus erythematosus (LUPIL-2): A multicentre, double-blind, randomised and placebo-controlled phase II trial. Ann. Rheum. Dis. 2022, 81, 1685–1694. [Google Scholar] [CrossRef]
- Seillet, C.; Laffont, S.; Tremollieres, F.; Rouquie, N.; Ribot, C.; Arnal, J.F.; Douin-Echinard, V.; Gourdy, P.; Guery, J.C. The TLR-mediated response of plasmacytoid dendritic cells is positively regulated by estradiol in vivo through cell-intrinsic estrogen receptor alpha signaling. Blood 2012, 119, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Chakhtoura, M.; Sriram, U.; Caricchio, R.; Gallucci, S. Conventional DCs from Male and Female Lupus-Prone B6.NZM Sle1/Sle2/Sle3 Mice Express an IFN Signature and Have a Higher Immunometabolism That Are Enhanced by Estrogen. J. Immunol. Res. 2018, 2018, 1601079. [Google Scholar] [CrossRef] [PubMed]
- Tsakonas, E.; Joseph, L.; Esdaile, J.M.; Choquette, D.; Senecal, J.L.; Cividino, A.; Danoff, D.; Osterland, C.K.; Yeadon, C.; Smith, C.D. A long-term study of hydroxychloroquine withdrawal on exacerbations in systemic lupus erythematosus. The Canadian Hydroxychloroquine Study Group. Lupus 1998, 7, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Sakiani, S.; Olsen, N.J.; Kovacs, W.J. Gonadal steroids and humoral immunity. Nat. Rev. Endocrinol. 2013, 9, 56–62. [Google Scholar] [CrossRef]
- Wilhelmson, A.S.; Stubelius, A.; Borjesson, A.E.; Wu, J.; Stern, A.; Malin, S.; Martensson, I.L.; Ohlsson, C.; Carlsten, H.; Tivesten, A. Androgens regulate bone marrow B lymphopoiesis in male mice by targeting osteoblast-lineage cells. Endocrinology 2015, 156, 1228–1236. [Google Scholar] [CrossRef]
- Wilhelmson, A.S.; Lantero Rodriguez, M.; Stubelius, A.; Fogelstrand, P.; Johansson, I.; Buechler, M.B.; Lianoglou, S.; Kapoor, V.N.; Johansson, M.E.; Fagman, J.B.; et al. Testosterone is an endogenous regulator of BAFF and splenic B cell number. Nat. Commun. 2018, 9, 2067. [Google Scholar] [CrossRef]
- Wong, A.H.; Agrawal, N.; Hughes, G.C. Altered IgG autoantibody levels and CD4+ T cell subsets in lupus-prone Nba2 mice lacking the nuclear progesterone receptor. Autoimmunity 2015, 48, 389–401. [Google Scholar] [CrossRef]
- Bonner, R.; Albajrami, O.; Hudspeth, J.; Upadhyay, A. Diabetic Kidney Disease. Prim. Care 2020, 47, 645–659. [Google Scholar] [CrossRef]
- Anders, H.J.; Huber, T.B.; Isermann, B.; Schiffer, M. CKD in diabetes: Diabetic kidney disease versus nondiabetic kidney disease. Nat. Rev. Nephrol. 2018, 14, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Alicic, R.Z.; Rooney, M.T.; Tuttle, K.R. Diabetic Kidney Disease: Challenges, Progress, and Possibilities. Clin. J. Am. Soc. Nephrol. 2017, 12, 2032–2045. [Google Scholar] [CrossRef]
- van Blijderveen, J.C.; Straus, S.M.; Zietse, R.; Stricker, B.H.; Sturkenboom, M.C.; Verhamme, K.M. A population-based study on the prevalence and incidence of chronic kidney disease in the Netherlands. Int. Urol. Nephrol. 2014, 46, 583–592. [Google Scholar] [CrossRef]
- Yu, M.K.; Katon, W.; Young, B.A. Associations between sex and incident chronic kidney disease in a prospective diabetic cohort. Nephrology 2015, 20, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Maric, C.; Sullivan, S. Estrogens and the diabetic kidney. Gend. Med. 2008, 5 (Suppl. A), S103–S113. [Google Scholar] [CrossRef]
- Burns, K.A.; Korach, K.S. Estrogen receptors and human disease: An update. Arch. Toxicol. 2012, 86, 1491–1504. [Google Scholar] [CrossRef]
- Stringer, K.D.; Komers, R.; Osman, S.A.; Oyama, T.T.; Lindsley, J.N.; Anderson, S. Gender hormones and the progression of experimental polycystic kidney disease. Kidney Int. 2005, 68, 1729–1739. [Google Scholar] [CrossRef]
- Wells, C.C.; Riazi, S.; Mankhey, R.W.; Bhatti, F.; Ecelbarger, C.; Maric, C. Diabetic nephropathy is associated with decreased circulating estradiol levels and imbalance in the expression of renal estrogen receptors. Gend. Med. 2005, 2, 227–237. [Google Scholar] [CrossRef]
- Andersson, B.; Mattsson, L.A.; Hahn, L.; Marin, P.; Lapidus, L.; Holm, G.; Bengtsson, B.A.; Bjorntorp, P. Estrogen replacement therapy decreases hyperandrogenicity and improves glucose homeostasis and plasma lipids in postmenopausal women with noninsulin-dependent diabetes mellitus. J. Clin. Endocrinol. Metab. 1997, 82, 638–643. [Google Scholar] [CrossRef]
- Brussaard, H.E.; Gevers Leuven, J.A.; Frolich, M.; Kluft, C.; Krans, H.M. Short-term oestrogen replacement therapy improves insulin resistance, lipids and fibrinolysis in postmenopausal women with NIDDM. Diabetologia 1997, 40, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Szekacs, B.; Vajo, Z.; Varbiro, S.; Kakucs, R.; Vaslaki, L.; Acs, N.; Mucsi, I.; Brinton, E.A. Postmenopausal hormone replacement improves proteinuria and impaired creatinine clearance in type 2 diabetes mellitus and hypertension. BJOG 2000, 107, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Chin, M.; Isono, M.; Isshiki, K.; Araki, S.; Sugimoto, T.; Guo, B.; Sato, H.; Haneda, M.; Kashiwagi, A.; Koya, D. Estrogen and raloxifene, a selective estrogen receptor modulator, ameliorate renal damage in db/db mice. Am. J. Pathol. 2005, 166, 1629–1636. [Google Scholar] [CrossRef]
- Hadjadj, S.; Gourdy, P.; Zaoui, P.; Guerci, B.; Roudaut, N.; Gautier, J.F.; Chabin, M.; Mauco, G.; Ragot, S.; for the RADIAN (Raloxifene in Diabetic Nephropathy) Study Group. Effect of raloxifene—A selective oestrogen receptor modulator—On kidney function in post-menopausal women with Type 2 diabetes: Results from a randomized, placebo-controlled pilot trial. Diabet. Med. 2007, 24, 906–910. [Google Scholar] [CrossRef]
- Hu, X.; Liu, W.; Yan, Y.; Liu, H.; Huang, Q.; Xiao, Y.; Gong, Z.; Du, J. Vitamin D protects against diabetic nephropathy: Evidence-based effectiveness and mechanism. Eur. J. Pharmacol. 2019, 845, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Agmon-Levin, N.; Theodor, E.; Segal, R.M.; Shoenfeld, Y. Vitamin D in systemic and organ-specific autoimmune diseases. Clin. Rev. Allergy Immunol. 2013, 45, 256–266. [Google Scholar] [CrossRef]
- Cutolo, M.; Paolino, S.; Sulli, A.; Smith, V.; Pizzorni, C.; Seriolo, B. Vitamin D, steroid hormones, and autoimmunity. Ann. N. Y. Acad. Sci. 2014, 1317, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Santoro, D.; Gembillo, G.; Ando, G. Glomerular Filtration Rate as a Predictor of Outcome in Acute Coronary Syndrome Complicated by Atrial Fibrillation. J. Clin. Med. 2020, 9, 1466. [Google Scholar] [CrossRef]
- Gangula, P.R.; Dong, Y.L.; Al-Hendy, A.; Richard-Davis, G.; Montgomery-Rice, V.; Haddad, G.; Millis, R.; Nicholas, S.B.; Moseberry, D. Protective cardiovascular and renal actions of vitamin D and estrogen. Front. Biosci. 2013, 5, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, M. Aldose reductase and the role of the polyol pathway in diabetic nephropathy. Kidney Int. Suppl. 2000, 77, S3–S12. [Google Scholar] [CrossRef]
- Elliot, S.J.; Karl, M.; Berho, M.; Xia, X.; Pereria-Simon, S.; Espinosa-Heidmann, D.; Striker, G.E. Smoking induces glomerulosclerosis in aging estrogen-deficient mice through cross-talk between TGF-beta1 and IGF-I signaling pathways. J. Am. Soc. Nephrol. 2006, 17, 3315–3324. [Google Scholar] [CrossRef]
- Blush, J.; Lei, J.; Ju, W.; Silbiger, S.; Pullman, J.; Neugarten, J. Estradiol reverses renal injury in Alb/TGF-beta1 transgenic mice. Kidney Int. 2004, 66, 2148–2154. [Google Scholar] [CrossRef]
- Cai, W.; Ramdas, M.; Zhu, L.; Chen, X.; Striker, G.E.; Vlassara, H. Oral advanced glycation endproducts (AGEs) promote insulin resistance and diabetes by depleting the antioxidant defenses AGE receptor-1 and sirtuin 1. Proc. Natl. Acad. Sci. USA 2012, 109, 15888–15893. [Google Scholar] [CrossRef]
- Uribarri, J.; Cai, W.; Ramdas, M.; Goodman, S.; Pyzik, R.; Chen, X.; Zhu, L.; Striker, G.E.; Vlassara, H. Restriction of advanced glycation end products improves insulin resistance in human type 2 diabetes: Potential role of AGER1 and SIRT1. Diabetes Care 2011, 34, 1610–1616. [Google Scholar] [CrossRef]
- Conti, G.; Caccamo, D.; Siligato, R.; Gembillo, G.; Satta, E.; Pazzano, D.; Carucci, N.; Carella, A.; Campo, G.D.; Salvo, A.; et al. Association of Higher Advanced Oxidation Protein Products (AOPPs) Levels in Patients with Diabetic and Hypertensive Nephropathy. Medicina 2019, 55, 675. [Google Scholar] [CrossRef]
- Negulescu, O.; Bognar, I.; Lei, J.; Devarajan, P.; Silbiger, S.; Neugarten, J. Estradiol reverses TGF-beta1-induced mesangial cell apoptosis by a casein kinase 2-dependent mechanism. Kidney Int. 2002, 62, 1989–1998. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.R.; Clegg, D.J.; Prossnitz, E.R.; Barton, M. Obesity, insulin resistance and diabetes: Sex differences and role of oestrogen receptors. Acta Physiol. 2011, 203, 259–269. [Google Scholar] [CrossRef]
- Lin, Y.; Nakachi, K.; Ito, Y.; Kikuchi, S.; Tamakoshi, A.; Yagyu, K.; Watanabe, Y.; Inaba, Y.; Tajima, K.; JACC Study Group. Variations in serum transforming growth factor-beta1 levels with gender, age and lifestyle factors of healthy Japanese adults. Dis. Mrk. 2009, 27, 23–28. [Google Scholar] [CrossRef]
- Lee, H.S.; Song, C.Y. Differential role of mesangial cells and podocytes in TGF-beta-induced mesangial matrix synthesis in chronic glomerular disease. Histol. Histopathol. 2009, 24, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Miller, D.; Ruoslahti, E.; Border, W.A. Production of extracellular matrix by glomerular epithelial cells is regulated by transforming growth factor-beta 1. Kidney Int. 1992, 41, 1213–1221. [Google Scholar] [CrossRef]
- Ruggenenti, P.; Cravedi, P.; Remuzzi, G. The RAAS in the pathogenesis and treatment of diabetic nephropathy. Nat. Rev. Nephrol. 2010, 6, 319–330. [Google Scholar] [CrossRef]
- Bumke-Vogt, C.; Bahr, V.; Diederich, S.; Herrmann, S.M.; Anagnostopoulos, I.; Oelkers, W.; Quinkler, M. Expression of the progesterone receptor and progesterone- metabolising enzymes in the female and male human kidney. J. Endocrinol. 2002, 175, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C.B.; Flyvbjerg, A.; Bruun, J.M.; Forman, A.; Wogensen, L.; Thomsen, K. Decreases in renal functional reserve and proximal tubular fluid output in conscious oophorectomized rats: Normalization with sex hormone substitution. J. Am. Soc. Nephrol. 2003, 14, 3102–3110. [Google Scholar] [CrossRef] [PubMed]
- Sandhi, J.; Singh, J.P.; Kaur, T.; Ghuman, S.S.; Singh, A.P. Involvement of progesterone receptors in ascorbic acid-mediated protection against ischemia-reperfusion-induced acute kidney injury. J. Surg. Res. 2014, 187, 278–288. [Google Scholar] [CrossRef]
- Al-Trad, B.; Ashankyty, I.M.; Alaraj, M. Progesterone ameliorates diabetic nephropathy in streptozotocin-induced diabetic Rats. Diabetol. Metab. Syndr. 2015, 7, 97. [Google Scholar] [CrossRef]
- Maric, C.; Forsblom, C.; Thorn, L.; Waden, J.; Groop, P.H.; on behalf of FinnDiane Study Group. Association between testosterone, estradiol and sex hormone binding globulin levels in men with type 1 diabetes with nephropathy. Steroids 2010, 75, 772–778. [Google Scholar] [CrossRef]
- Matsushita, M.; Tamura, K.; Osada, S.; Kogo, H. Effect of troglitazone on the excess testosterone and LH secretion in thyroidectomized, insulin-resistant, type 2 diabetic Goto-Kakizaki rats. Endocrine 2005, 27, 301–305. [Google Scholar] [CrossRef]
- Inada, A.; Inada, O.; Fujii, N.L.; Nagafuchi, S.; Katsuta, H.; Yasunami, Y.; Matsubara, T.; Arai, H.; Fukatsu, A.; Nabeshima, Y.I. Adjusting the 17beta-Estradiol-to-Androgen Ratio Ameliorates Diabetic Nephropathy. J. Am. Soc. Nephrol. 2016, 27, 3035–3050. [Google Scholar] [CrossRef]
- Elliot, S.J.; Berho, M.; Korach, K.; Doublier, S.; Lupia, E.; Striker, G.E.; Karl, M. Gender-specific effects of endogenous testosterone: Female alpha-estrogen receptor-deficient C57Bl/6J mice develop glomerulosclerosis. Kidney Int. 2007, 72, 464–472. [Google Scholar] [CrossRef]
- Sakemi, T.; Toyoshima, H.; Morito, F. Testosterone eliminates the attenuating effect of castration on the progressive glomerular injury in hypercholesterolemic male Imai rats. Nephron 1994, 67, 469–476. [Google Scholar] [CrossRef]
- Manigrasso, M.B.; Sawyer, R.T.; Marbury, D.C.; Flynn, E.R.; Maric, C. Inhibition of estradiol synthesis attenuates renal injury in male streptozotocin-induced diabetic rats. Am. J. Physiol. Ren. Physiol. 2011, 301, F634–F640. [Google Scholar] [CrossRef]
- Rodrigues, J.C.; Haas, M.; Reich, H.N. IgA Nephropathy. Clin. J. Am. Soc. Nephrol. 2017, 12, 677–686. [Google Scholar] [CrossRef]
- Beckwith, H.; Lightstone, L.; McAdoo, S. Sex and Gender in Glomerular Disease. Semin. Nephrol. 2022, 42, 185–196. [Google Scholar] [CrossRef]
- Lai, K.N.; Tang, S.C.; Schena, F.P.; Novak, J.; Tomino, Y.; Fogo, A.B.; Glassock, R.J. IgA nephropathy. Nat. Rev. Dis. Prim. 2016, 2, 16001. [Google Scholar] [CrossRef]
- Huang, P.P.; Shu, D.H.; Su, Z.; Luo, S.N.; Xu, F.F.; Lin, F. Association between lifestyle, gender and risk for developing end-stage renal failure in IgA nephropathy: A case-control study within 10 years. Ren. Fail. 2019, 41, 914–920. [Google Scholar] [CrossRef]
- Donadio, J.V.; Bergstralh, E.J.; Grande, J.P.; Rademcher, D.M. Proteinuria patterns and their association with subsequent end-stage renal disease in IgA nephropathy. Nephrol. Dial. Transplant. 2002, 17, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.M.; Azcona-Olivera, J.I.; Pestka, J.J. Vomitoxin (deoxynivalenol)-induced IgA nephropathy in the B6C3F1 mouse: Dose response and male predilection. Toxicology 1994, 92, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.M.; Azcona-Olivera, J.I.; Murtha, J.M.; Pestka, J.J. Effects of dihydrotestosterone and estradiol on experimental IgA nephropathy induced by vomitoxin. Fundam. Appl. Toxicol. 1995, 26, 107–116. [Google Scholar] [CrossRef]
- Yamamoto, R.; Nagasawa, Y.; Shoji, T.; Katakami, N.; Ohtoshi, K.; Hayaishi-Okano, R.; Yamasaki, Y.; Yamauchi, A.; Tsubakihara, Y.; Imai, E.; et al. A candidate gene approach to genetic contributors to the development of IgA nephropathy. Nephrol. Dial. Transplant. 2012, 27, 1020–1030. [Google Scholar] [CrossRef]
- Hu, S.L.; Wang, D.; Yuan, F.L.; Lei, Q.F.; Zhang, Y.; Cheng, J.Z. Identification of key genes and pathways in IgA nephropathy using bioinformatics analysis. Medicine 2020, 99, e21372. [Google Scholar] [CrossRef]
- Yu, W.; Zhao, B.; Zhong, H.; Yao, G. Estrogen receptor alpha expression in renal tissue and its relationship with prognosis in immunoglobulin A nephropathy. Int. J. Clin. Exp. Pathol. 2020, 13, 2319–2325. [Google Scholar] [PubMed]
- Bergmann, C.; Guay-Woodford, L.M.; Harris, P.C.; Horie, S.; Peters, D.J.M.; Torres, V.E. Polycystic kidney disease. Nat. Rev. Dis. Prim. 2018, 4, 50. [Google Scholar] [CrossRef]
- Cowley, B.D., Jr.; Rupp, J.C.; Muessel, M.J.; Gattone, V.H., 2nd. Gender and the effect of gonadal hormones on the progression of inherited polycystic kidney disease in rats. Am. J. Kidney Dis. 1997, 29, 265–272. [Google Scholar] [CrossRef]
- Gretz, N.; Ceccherini, I.; Kranzlin, B.; Kloting, I.; Devoto, M.; Rohmeiss, P.; Hocher, B.; Waldherr, R.; Romeo, G. Gender-dependent disease severity in autosomal polycystic kidney disease of rats. Kidney Int. 1995, 48, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Talbi, K.; Cabrita, I.; Schreiber, R.; Kunzelmann, K. Gender-Dependent Phenotype in Polycystic Kidney Disease Is Determined by Differential Intracellular Ca2+ Signals. Int. J. Mol. Sci. 2021, 22, 6019. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.Y.; Wee, J.; Jung, J.; Jang, Y.; Lee, B.; Hong, G.S.; Chang, B.C.; Choi, Y.L.; Shin, Y.K.; Min, H.Y.; et al. Anoctamin 1 (TMEM16A) is essential for testosterone-induced prostate hyperplasia. Proc. Natl. Acad. Sci. USA 2015, 112, 9722–9727. [Google Scholar] [CrossRef]
- Sweezey, N.; Tchepichev, S.; Gagnon, S.; Fertuck, K.; O’Brodovich, H. Female gender hormones regulate mRNA levels and function of the rat lung epithelial Na channel. Am. J. Physiol. 1998, 274, C379–C386. [Google Scholar] [CrossRef]
- Saint-Criq, V.; Harvey, B.J. Estrogen and the cystic fibrosis gender gap. Steroids 2014, 81, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Migliore, L.; Nicoli, V.; Stoccoro, A. Gender Specific Differences in Disease Susceptibility: The Role of Epigenetics. Biomedicines 2021, 9, 652. [Google Scholar] [CrossRef]
- Plunk, E.C.; Richards, S.M. Epigenetic Modifications due to Environment, Ageing, Nutrition, and Endocrine Disrupting Chemicals and Their Effects on the Endocrine System. Int. J. Endocrinol. 2020, 2020, 9251980. [Google Scholar] [CrossRef]
- Veloso Pereira, B.M.; Charleaux de Ponte, M.; Malavolta Luz, A.P.; Thieme, K. DNA methylation enzymes in the kidneys of male and female BTBR ob/ob mice. Front. Endocrinol. 2023, 14, 1167546. [Google Scholar] [CrossRef]
- Wu, Z.; Sun, Y.; Mei, X.; Zhang, C.; Pan, W.; Shi, W. 17beta-oestradiol enhances global DNA hypomethylation in CD4-positive T cells from female patients with lupus, through overexpression of oestrogen receptor-alpha-mediated downregulation of DNMT1. Clin. Exp. Dermatol. 2014, 39, 525–532. [Google Scholar] [CrossRef]
- Ding, H.; Zhang, L.; Yang, Q.; Zhang, X.; Li, X. Epigenetics in kidney diseases. Adv. Clin. Chem. 2021, 104, 233–297. [Google Scholar] [CrossRef]
- Ransick, A.; Lindstrom, N.O.; Liu, J.; Zhu, Q.; Guo, J.J.; Alvarado, G.F.; Kim, A.D.; Black, H.G.; Kim, J.; McMahon, A.P. Single-Cell Profiling Reveals Sex, Lineage, and Regional Diversity in the Mouse Kidney. Dev. Cell 2019, 51, 399–413.e7. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Liao, J.; He, J.; Pan, S.; Zhang, H.; Yang, X.; Cheng, J.; Chen, Y.; Mo, Z. Single-cell profiling reveals sex diversity in human renal proximal tubules. Gene 2020, 752, 144790. [Google Scholar] [CrossRef]
- Chen, L.; Chou, C.L.; Yang, C.R.; Knepper, M.A. Multiomics Analyses Reveal Sex Differences in Mouse Renal Proximal Subsegments. J. Am. Soc. Nephrol. 2023, 34, 829–845. [Google Scholar] [CrossRef]
- Sapir-Pichhadze, R.; Oertelt-Prigione, S. P32: A sex- and gender-sensitive model for evidence-based precision medicine: From knowledge generation to implementation in the field of kidney transplantation. Kidney Int. 2023, 103, 674–685. [Google Scholar] [CrossRef]
- Idda, M.L.; Campesi, I.; Fiorito, G.; Vecchietti, A.; Urru, S.A.M.; Solinas, M.G.; Franconi, F.; Floris, M. Sex-Biased Expression of Pharmacogenes across Human Tissues. Biomolecules 2021, 11, 1206. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Kidney Disease | Sex-Specific Associations |
|---|---|
| Lupus nephritis |
|
| Minimal change nephropathy |
|
| Diabetic kidney disease |
|
| Anti-GBM disease | |
| IgA Nephropathy | |
| Focal segmental glomerulosclerosis |
|
| Autosomal dominant polycystic kidney disease |
|
| ANCA associated vasculitis |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conte, C.; Antonelli, G.; Melica, M.E.; Tarocchi, M.; Romagnani, P.; Peired, A.J. Role of Sex Hormones in Prevalent Kidney Diseases. Int. J. Mol. Sci. 2023, 24, 8244. https://doi.org/10.3390/ijms24098244
Conte C, Antonelli G, Melica ME, Tarocchi M, Romagnani P, Peired AJ. Role of Sex Hormones in Prevalent Kidney Diseases. International Journal of Molecular Sciences. 2023; 24(9):8244. https://doi.org/10.3390/ijms24098244
Chicago/Turabian StyleConte, Carolina, Giulia Antonelli, Maria Elena Melica, Mirko Tarocchi, Paola Romagnani, and Anna Julie Peired. 2023. "Role of Sex Hormones in Prevalent Kidney Diseases" International Journal of Molecular Sciences 24, no. 9: 8244. https://doi.org/10.3390/ijms24098244
APA StyleConte, C., Antonelli, G., Melica, M. E., Tarocchi, M., Romagnani, P., & Peired, A. J. (2023). Role of Sex Hormones in Prevalent Kidney Diseases. International Journal of Molecular Sciences, 24(9), 8244. https://doi.org/10.3390/ijms24098244