
The Role of Paxillin Aberrant Expression in Cancer and Its Potential as a Target for Cancer Therapy


Abstract
:1. Introduction
2. Structure and Expression of Paxillin in Tumors
2.1. Structure of Paxillin
2.2. Expression of Paxillin in Tumors

{kind=link}
{kind=link}
{kind=link}
| System | Tumor Type | Expression | Level | Reference |
|---|---|---|---|---|
| Nervous system | Glioma | Upregulated | Tissue | [24] |
| Digestive system | GC | Upregulated | Tissue and cell | [7] |
| CRC | Upregulated | Tissue | [8] | |
| Esophageal cancer | Upregulated | Tissue | [28] | |
| LSCC | Upregulated | Tissue | [29] | |
| Pancreatic adenocarcinoma | Upregulated | Tissue | [30] | |
| Urinary system | CC | Upregulated | Tissue | [31] |
| PC | Upregulated | Tissue and cell | [32] | |
| Other | HNSCC | Upregulated | Tissue | [33] |
3. Clinical Prognostic Value
| Tumor Type | Sample Size | Prognostic Value | Clinical Significance | Reference |
|---|---|---|---|---|
| GC | 239 | Independent prognostic factors for patient survival | Positively related to distant metastasis and advanced tumor stage | [7] |
| CRC | 242 | Independent prognostic factors for patient survival | Positively related to tumor histological grade, tumor size, clinical TNM stage, and distant metastasis | [34] |
| PC | 386 | Independent risk factors for lymph node metastasis in prostate cancer | Positively related to preoperative prostate-specific antigen levels, clinical tumor stage, lymph node metastasis, and seminal vesicle invasion | [35] |
| CC | 430 | - | Positively related to tumor stage, poor differentiation, lymphovascular space invasion, and lymphovascular metastasis | [31] |
| SC/ASC | 46 | Independent prognostic factors for patient survival | Positively related to larger tumors, higher TNM stage, lymph node metastasis, and tumor aggressiveness | [37] |
| LSCC | 84 | Independent prognostic factors for overall patient survival | Positively related to histopathological grade, lymph node metastasis, and TNM stage | [29] |
| TSCC | 48 | Independent factors of survival in disease-free patients | Positively related to tumor differentiation, disease-free survival, lymph node metastasis, and depth of invasion | [38] |
| GBM | 325 | Independent prognostic factors for overall survival of patients | Positively related to tumor grade and overall survival of patients | [41] |
| HNSCC | 518 | Independent prognostic factors for patient survival | - | [33] |
4. Paxillin Expression and Tumor Cell Metastasis
4.1. Paxillin Regulates Tumor Cell Migration
4.2. Paxillin Regulates Tumor Cell Invasive Metastasis
5. Paxillin Affects Tumor Cell Survival and Angiogenesis
5.1. Tumor Cell Survival
5.2. Tumor Angiogenesis
6. The Signal Pathways and Regulatory Factors of Paxillin in Cancer
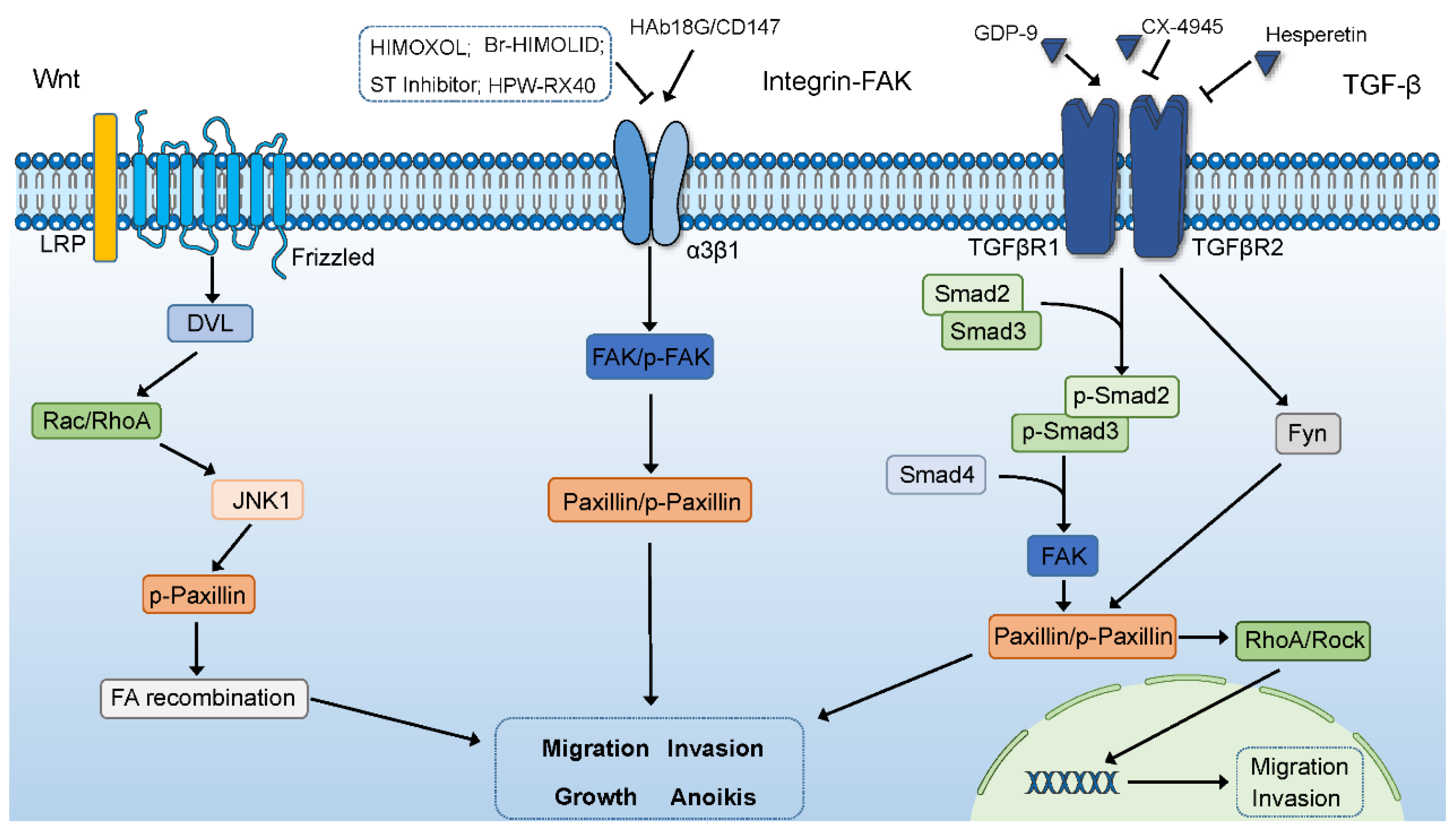
6.1. The Wnt Signaling Pathway
6.2. The Integrin-FAK Signaling Pathway
6.3. The TGF-B Signaling Pathway
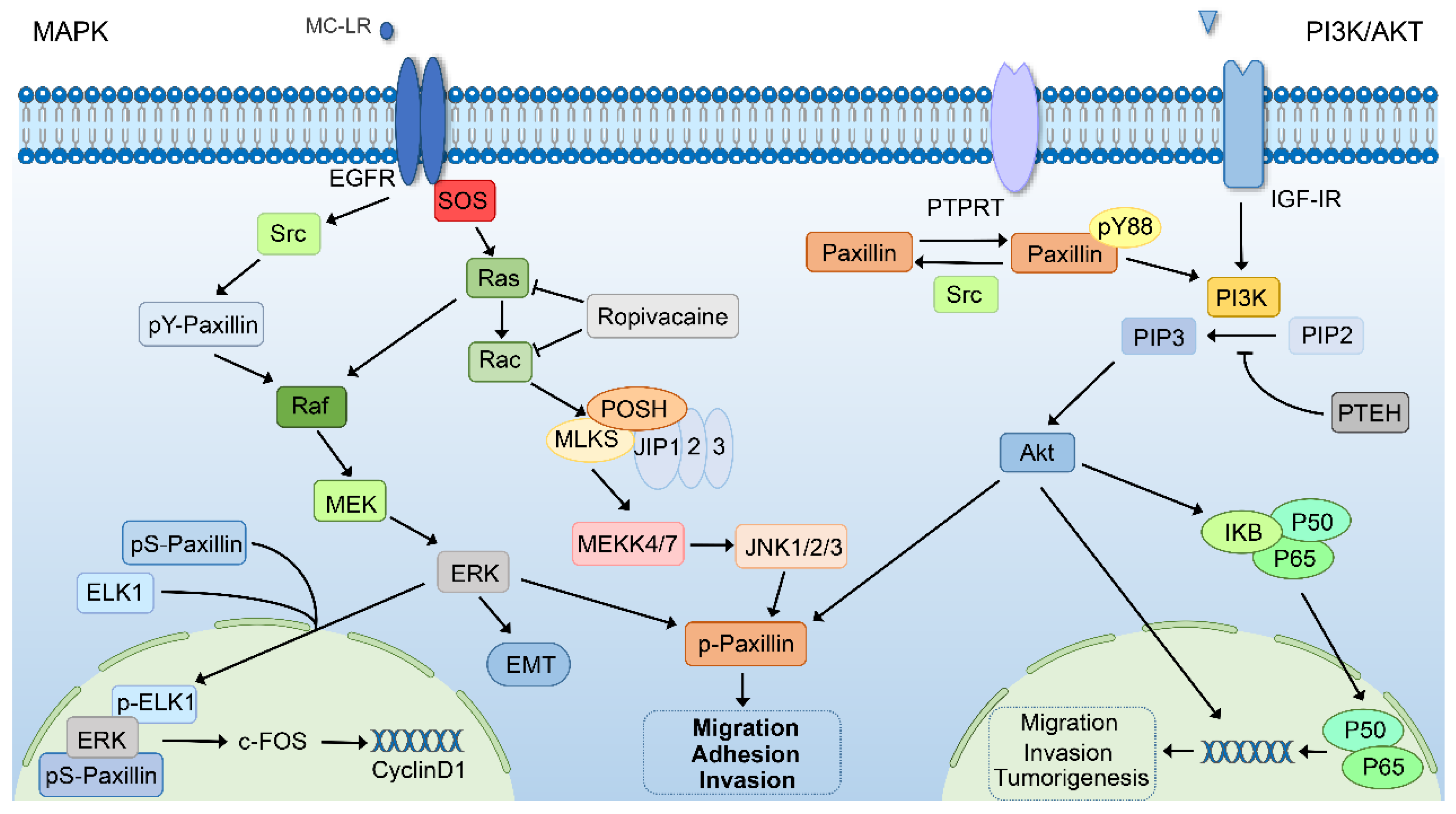
6.4. The MAPK Signaling Pathway
6.5. The PIK3/AKT Signaling Pathway
6.6. Other Regulatory Factors of Paxillin in Cancer
7. Therapeutic Potential
| Drugs | Cancer Types | Functions | Mechanisms | References |
|---|---|---|---|---|
| Paxillin antibody selective inhibitor | - | - | Inhibition of FAK-paxillin interaction | [166] |
| Magnolol | BC | Migration, invasion | Inhibition of FAK/Src/paxillin signaling pathway | [170] |
| Lith-O-Asp | Lung cancer | metastasis | Inhibition of FAK/paxillin signaling pathway | [171] |
| Mifepristone (RU486) | BC | Migration, adhesion | Inhibits the formation of FAK/Src/paxillin complex | [175] |
| SK228 | Lung and esophagus cancer | growth | Disruption of the F-actin cytoskeleton and FAK/paxillin signaling axis | [176] |
| Docetaxel | PC | Growth, apoptosis | Inhibits phosphorylation of paxillin | [177] |
| Deguelin | NSCLC | Metastasis | Inhibition of FAK/Src/paxillin signaling pathway | [178] |
| Cucurbitacin B | BC | Metastasis | Inhibition of FAK/paxillin signaling pathway | [179] |
| Hesperetin | BC | Migration, invasion | Inhibits phosphorylation of paxillin | [139] |
| D/PSP@CQ/CaP | BC | Metastasis, growth | Inhibition of autophagy-dependent degradation of paxillin | [185] |
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, M.C.; Turner, C.E. Paxillin: Adapting to change. Physiol. Rev. 2004, 84, 1315–1339. [Google Scholar] [CrossRef] [PubMed]
- Alpha, K.M.; Xu, W.; Turner, C.E. Paxillin family of focal adhesion adaptor proteins and regulation of cancer cell invasion. Int. Rev. Cell Mol. Biol. 2020, 355, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Colome, A.M.; Lee-Rivera, I.; Benavides-Hidalgo, R.; Lopez, E. Paxillin: A crossroad in pathological cell migration. J. Hematol. Oncol. 2017, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Hammes, S.R. Paxillin actions in the nucleus. Steroids 2018, 133, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Velazquez, M.A.; Salinas-Jazmin, N.; Mendoza-Patino, N.; Mandoki, J.J. Reduced paxillin expression contributes to the antimetastatic effect of 4-hydroxycoumarin on B16-F10 melanoma cells. Cancer Cell Int. 2008, 8, 8. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, T.; Tang, W.X.; Li, S.L. Expression of paxillin in breast cancer cell with high and low metastatic potentiality. Sichuan Da Xue Xue Bao Yi Xue Ban 2010, 41, 91–94. [Google Scholar]
- Chen, D.L.; Wang, Z.Q.; Ren, C.; Zeng, Z.L.; Wang, D.S.; Luo, H.Y.; Wang, F.; Qiu, M.Z.; Bai, L.; Zhang, D.S.; et al. Abnormal expression of paxillin correlates with tumor progression and poor survival in patients with gastric cancer. J. Transl. Med. 2013, 11, 277. [Google Scholar] [CrossRef]
- Yang, H.J.; Chen, J.Z.; Zhang, W.L.; Ding, Y.Q. Focal adhesion plaque associated cytoskeletons are involved in the invasion and metastasis of human colorectal carcinoma. Cancer Investig. 2010, 28, 127–134. [Google Scholar] [CrossRef]
- Tumbarello, D.A.; Brown, M.C.; Turner, C.E. The paxillin LD motifs. FEBS Lett. 2002, 513, 114–118. [Google Scholar] [CrossRef]
- Gehmlich, K.; Pinotsis, N.; Hayess, K.; van der Ven, P.F.; Milting, H.; El Banayosy, A.; Korfer, R.; Wilmanns, M.; Ehler, E.; Furst, D.O. Paxillin and ponsin interact in nascent costameres of muscle cells. J. Mol. Biol. 2007, 369, 665–682. [Google Scholar] [CrossRef]
- Nayal, A.; Webb, D.J.; Brown, C.M.; Schaefer, E.M.; Vicente-Manzanares, M.; Horwitz, A.R. Paxillin phosphorylation at Ser273 localizes a GIT1-PIX-PAK complex and regulates adhesion and protrusion dynamics. J. Cell Biol. 2006, 173, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Yan, D.P.; Ge, B.X. JNK regulates cell migration through promotion of tyrosine phosphorylation of paxillin. Cell. Signal. 2008, 20, 2002–2012. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Yano, H.; Uchida, H.; Hashimoto, S.; Schaefer, E.; Sabe, H. Tyrosine phosphorylation of paxillin alpha is involved in temporospatial regulation of paxillin-containing focal adhesion formation and F-actin organization in motile cells. J. Biol. Chem. 2000, 275, 27155–27164. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, T.; Nakata, A.; Mukai, M.; Shinkai, K.; Yano, H.; Sabe, H.; Schaefer, E.; Tatsuta, M.; Tsujimura, T.; Terada, N.; et al. Involvement of phosphorylation of Tyr-31 and Tyr-118 of paxillin in MM1 cancer cell migration. Int. J. Cancer 2002, 97, 330–335. [Google Scholar] [CrossRef]
- Petit, V.; Boyer, B.; Lentz, D.; Turner, C.E.; Thiery, J.P.; Valles, A.M. Phosphorylation of tyrosine residues 31 and 118 on paxillin regulates cell migration through an association with CRK in NBT-II cells. J. Cell Biol. 2000, 148, 957–970. [Google Scholar] [CrossRef]
- Abou Zeid, N.; Valles, A.M.; Boyer, B. Serine phosphorylation regulates paxillin turnover during cell migration. Cell Commun. Signal. 2006, 4, 8. [Google Scholar] [CrossRef]
- Cai, X.; Li, M.; Vrana, J.; Schaller, M.D. Glycogen synthase kinase 3- and extracellular signal-regulated kinase-dependent phosphorylation of paxillin regulates cytoskeletal rearrangement. Mol. Cell. Biol. 2006, 26, 2857–2868. [Google Scholar] [CrossRef]
- Woodrow, M.A.; Woods, D.; Cherwinski, H.M.; Stokoe, D.; McMahon, M. Ras-induced serine phosphorylation of the focal adhesion protein paxillin is mediated by the Raf→MEK→ERK pathway. Exp. Cell Res. 2003, 287, 325–338. [Google Scholar] [CrossRef]
- Perez-Alvarado, G.C.; Miles, C.; Michelsen, J.W.; Louis, H.A.; Winge, D.R.; Beckerle, M.C.; Summers, M.F. Structure of the carboxy-terminal LIM domain from the cysteine rich protein CRP. Nat. Struct. Biol. 1994, 1, 388–398. [Google Scholar] [CrossRef]
- Schmeichel, K.L.; Beckerle, M.C. The LIM domain is a modular protein-binding interface. Cell 1994, 79, 211–219. [Google Scholar] [CrossRef]
- Brown, M.C.; Perrotta, J.A.; Turner, C.E. Identification of LIM3 as the principal determinant of paxillin focal adhesion localization and characterization of a novel motif on paxillin directing vinculin and focal adhesion kinase binding. J. Cell Biol. 1996, 135, 1109–1123. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.C.; Turner, C.E. Roles for the tubulin- and PTP-PEST-binding paxillin LIM domains in cell adhesion and motility. Int. J. Biochem. Cell Biol. 2002, 34, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Jagadeeswaran, R.; Surawska, H.; Krishnaswamy, S.; Janamanchi, V.; Mackinnon, A.C.; Seiwert, T.Y.; Loganathan, S.; Kanteti, R.; Reichman, T.; Nallasura, V.; et al. Paxillin is a target for somatic mutations in lung cancer: Implications for cell growth and invasion. Cancer Res. 2008, 68, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Xia, L.; Xu, C.S.; Xiao, F.; Wang, Y.F. Paxillin functions as an oncogene in human gliomas by promoting cell migration and invasion. Onco Targets Ther. 2016, 9, 6935–6943. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Wu, D.W.; Lin, P.L.; Lee, H. Paxillin promotes colorectal tumor invasion and poor patient outcomes via ERK-mediated stabilization of Bcl-2 protein by phosphorylation at Serine 87. Oncotarget 2015, 6, 8698–8708. [Google Scholar] [CrossRef]
- Zhao, Y.; Scott, A.; Zhang, P.; Hao, Y.; Feng, X.; Somasundaram, S.; Khalil, A.M.; Willis, J.; Wang, Z. Regulation of paxillin-p130-PI3K-AKT signaling axis by Src and PTPRT impacts colon tumorigenesis. Oncotarget 2017, 8, 48782–48793. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, H.; Xiao, Y.; Shen, P.; Tan, L.; Zhang, S.; Liu, Q.; Gao, Z.; Zhao, J.; Zhao, Y.; et al. Pan-cancer analysis reveals an immunological role and prognostic potential of PXN in human cancer. Aging 2021, 13, 16248–16266. [Google Scholar] [CrossRef]
- Cai, H.X.; Yang, L.C.; Song, X.H.; Liu, Z.R.; Chen, Y.B.; Dong, G.K. Expression of paxillin and FAK mRNA and the related clinical significance in esophageal carcinoma. Mol. Med. Rep. 2012, 5, 469–472. [Google Scholar] [CrossRef]
- Li, L.; Wang, J.; Gao, L.; Gong, L. Expression of paxillin in laryngeal squamous cell carcinoma and its prognostic value. Int. J. Clin. Exp. Pathol. 2015, 8, 9232–9239. [Google Scholar]
- Lu, W.; Wei, W.; de Bock, G.H.; Zhou, H.; Li, Q.; Shen, X. The roles of Wnt5a, JNK and paxillin in the occurrence of metastasis of pancreatic adenocarcinoma. Int. J. Clin. Oncol. 2014, 19, 1011–1019. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, J.; Tang, M.; Chen, L.; Qi, X.; Li, J.; Yu, J.; Qiu, H.; Wang, Y. The overexpression of PXN promotes tumor progression and leads to radioresistance in cervical cancer. Future Oncol. 2018, 14, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Posadas, E.M.; Al-Ahmadie, H.; Robinson, V.L.; Jagadeeswaran, R.; Otto, K.; Kasza, K.E.; Tretiakov, M.; Siddiqui, J.; Pienta, K.J.; Stadler, W.M.; et al. FYN is overexpressed in human prostate cancer. BJU Int. 2009, 103, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Minemura, C.; Asai, S.; Kikkawa, N.; Kinoshita, T.; Oshima, S.; Koma, A.; Kasamatsu, A.; Hanazawa, T.; Uzawa, K.; et al. Identification of miR-199-5p and miR-199-3p Target Genes: Paxillin Facilities Cancer Cell Aggressiveness in Head and Neck Squamous Cell Carcinoma. Genes 2021, 12, 1910. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.J.; Du, S.K.; Dang, X.B.; Gong, M. Expression of Paxillin is Correlated with Clinical Prognosis in Colorectal Cancer Patients. Med. Sci. Monit. 2015, 21, 1989–1995. [Google Scholar] [CrossRef]
- Zheng, Q.S.; Chen, S.H.; Wu, Y.P.; Chen, H.J.; Chen, H.; Wei, Y.; Li, X.D.; Huang, J.B.; Xue, X.Y.; Xu, N. Increased Paxillin expression in prostate cancer is associated with advanced pathological features, lymph node metastases and biochemical recurrence. J. Cancer 2018, 9, 959–967. [Google Scholar] [CrossRef]
- Shi, J.; Wang, S.; Zhao, E.; Shi, L.; Xu, X.; Fang, M. Paxillin expression levels are correlated with clinical stage and metastasis in salivary adenoid cystic carcinoma. J. Oral. Pathol. Med. 2010, 39, 548–551. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, Z.; Jiang, S.; Zou, Q.; Yuan, Y.; Li, J.; Li, D.; Liang, L.; Chen, M.; Chen, S. Paxillin and carbonic anhydrase IX are prognostic markers in gallbladder squamous cell/adenosquamous carcinomas and adenocarcinomas. Histopathology 2014, 64, 921–934. [Google Scholar] [CrossRef]
- Theocharis, S.; Kotta-Loizou, I.; Giaginis, C.; Alexandrou, P.; Danas, E.; Tsourouflis, G.; Tsoukalas, N.; Coutts, R.H.; Tasoulas, J.; Klijanienko, J. Expression and Clinical Significance of Concomitant FAK/SRC and p-Paxillin in Mobile Tongue Squamous Cell Carcinoma. Anticancer Res. 2017, 37, 1313–1319. [Google Scholar] [CrossRef]
- Kratimenos, P.; Koutroulis, I.; Syriopoulou, V.; Michailidi, C.; Delivoria-Papadopoulos, M.; Klijanienko, J.; Theocharis, S. FAK-Src-paxillin system expression and disease outcome in human neuroblastoma. Pediatr. Hematol. Oncol. 2017, 34, 221–230. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, H.; Sun, D.; Liu, J. Identification of Paxillin as a Prognostic Factor for Glioblastoma via Integrated Bioinformatics Analysis. BioMed Res. Int. 2022, 2022, 7171126. [Google Scholar] [CrossRef]
- Sun, L.H.; Yang, F.Q.; Zhang, C.B.; Wu, Y.P.; Liang, J.S.; Jin, S.; Wang, Z.; Wang, H.J.; Bao, Z.S.; Yang, Z.X.; et al. Overexpression of Paxillin Correlates with Tumor Progression and Predicts Poor Survival in Glioblastoma. CNS Neurosci. Ther. 2017, 23, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Wu, D.W.; Shen, C.J.; Cheng, Y.M.; Wu, C.C.; Lee, H. Hepatitis B virus X protein promotes tumor invasion and poor prognosis in hepatocellular carcinoma via phosphorylation of paxillin at Serine 178 by activation of the c-Jun NH2-terminal kinase. Am. J. Cancer Res. 2020, 10, 275–283. [Google Scholar] [PubMed]
- Mackinnon, A.C.; Tretiakova, M.; Henderson, L.; Mehta, R.G.; Yan, B.C.; Joseph, L.; Krausz, T.; Husain, A.N.; Reid, M.E.; Salgia, R. Paxillin expression and amplification in early lung lesions of high-risk patients, lung adenocarcinoma and metastatic disease. J. Clin. Pathol. 2011, 64, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Zhang, C.; Feng, Y.; Chen, G.; Wen, S.; Huangfu, H.; Wang, B. Fascin-1, ezrin and paxillin contribute to the malignant progression and are predictors of clinical prognosis in laryngeal squamous cell carcinoma. PLoS ONE 2012, 7, e50710. [Google Scholar] [CrossRef]
- Yin, H.; Zhang, Q.; Wang, X.; Li, T.; Wan, Y.; Liu, Y.; Zhu, J. Role of paxillin in colorectal carcinoma and its relationship to clinicopathological features. Chin. Med. J. 2014, 127, 423–429. [Google Scholar]
- Andisheh-Tadbir, A.; Afshari, A.; Ashraf, M.J. Expression of Paxillin in Benign and Malignant Salivary Gland Tumors. J. Dent. 2019, 20, 249–254. [Google Scholar] [CrossRef]
- Li, B.Z.; Lei, W.; Zhang, C.Y.; Zhou, F.; Li, N.; Shi, S.S.; Feng, X.L.; Chen, Z.L.; Hang, J.; Qiu, B.; et al. Increased expression of paxillin is found in human oesophageal squamous cell carcinoma: A tissue microarray study. J. Int. Med. Res. 2008, 36, 273–278. [Google Scholar] [CrossRef]
- De Pascalis, C.; Etienne-Manneville, S. Single and collective cell migration: The mechanics of adhesions. Mol. Biol. Cell 2017, 28, 1833–1846. [Google Scholar] [CrossRef]
- Ye, X.; Weinberg, R.A. Epithelial-Mesenchymal Plasticity: A Central Regulator of Cancer Progression. Trends Cell Biol. 2015, 25, 675–686. [Google Scholar] [CrossRef]
- Friedl, P.; Gilmour, D. Collective cell migration in morphogenesis, regeneration and cancer. Nat. Rev. Mol. Cell Biol. 2009, 10, 445–457. [Google Scholar] [CrossRef]
- Sobkowicz, A.D.; Sanders, A.J.; Mason, M.D.; Jiang, W.G. Potential Implication of Paxillin in Cancer Establishment within the Bone Environment. Anticancer Res. 2017, 37, 4255–4268. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, H.; Li, N.; Sun, H.; Li, Q.; Shen, X. WNT5A/JNK signaling regulates pancreatic cancer cells migration by Phosphorylating Paxillin. Pancreatology 2013, 13, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Wu, Y.M.; Zhao, P.; Yang, X.M.; Jiang, J.L.; Chen, Z.N. Overexpression of HAb18G/CD147 promotes invasion and metastasis via alpha3beta1 integrin mediated FAK-paxillin and FAK-PI3K-Ca2+ pathways. Cell. Mol. Life Sci. 2008, 65, 2933–2942. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Mu, G.; Zhang, L.; Zhou, W.; Zhang, J.; Yu, H. Lysophosphatidic acid stimulates activation of focal adhesion kinase and paxillin and promotes cell motility, via LPA1-3, in human pancreatic cancer. Dig. Dis. Sci. 2013, 58, 3524–3533. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Bijian, K.; Qiu, D.; Su, J.; Saad, A.; Dahabieh, M.S.; Miller, W.H., Jr.; Alaoui-Jamali, M.A. Endosomal sorting and c-Cbl targeting of paxillin to autophagosomes regulate cell-matrix adhesion turnover in human breast cancer cells. Oncotarget 2017, 8, 31199–31214. [Google Scholar] [CrossRef]
- Sanchez, A.M.; Shortrede, J.E.; Vargas-Roig, L.M.; Flamini, M.I. Retinoic acid induces nuclear FAK translocation and reduces breast cancer cell adhesion through Moesin, FAK, and Paxillin. Mol. Cell. Endocrinol. 2016, 430, 1–11. [Google Scholar] [CrossRef]
- Kuo, C.H.; Liu, C.J.; Lu, C.Y.; Hu, H.M.; Kuo, F.C.; Liou, Y.S.; Yang, Y.C.; Hsieh, M.C.; Lee, O.K.; Wu, D.C.; et al. 17beta-estradiol inhibits mesenchymal stem cells-induced human AGS gastric cancer cell mobility via suppression of CCL5- Src/Cas/Paxillin signaling pathway. Int. J. Med. Sci. 2014, 11, 7–16. [Google Scholar] [CrossRef]
- Liu, C.J.; Kuo, F.C.; Hu, H.M.; Chen, C.Y.; Huang, Y.B.; Cheng, K.H.; Yokoyama, K.K.; Wu, D.C.; Hsieh, S.; Kuo, C.H. 17beta-Estradiol inhibition of IL-6-Src and Cas and paxillin pathway suppresses human mesenchymal stem cells-mediated gastric cancer cell motility. Transl. Res. 2014, 164, 232–243. [Google Scholar] [CrossRef]
- Das, J.; Agarwal, T.; Chakraborty, S.; Maiti, T.K. Compressive stress-induced autophagy promotes invasion of HeLa cells by facilitating protein turnover in vitro. Exp. Cell Res. 2019, 381, 201–207. [Google Scholar] [CrossRef]
- Katoh, K. Activation of Rho-kinase and focal adhesion kinase regulates the organization of stress fibers and focal adhesions in the central part of fibroblasts. PeerJ 2017, 5, e4063. [Google Scholar] [CrossRef]
- Hodge, R.G.; Ridley, A.J. Regulating Rho GTPases and their regulators. Nat. Rev. Mol. Cell. Biol. 2016, 17, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Raftopoulou, M.; Hall, A. Cell migration: Rho GTPases lead the way. Dev. Biol. 2004, 265, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Peng, X.; Zheng, Q. Ropivacaine inhibits the migration of esophageal cancer cells via sodium-channel-independent but prenylation-dependent inhibition of Rac1/JNK/paxillin/FAK. Biochem. Biophys. Res. Commun. 2018, 501, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Alpha, K.M.; Zehrbach, N.M.; Turner, C.E. Paxillin promotes breast tumor collective cell invasion through maintenance of adherens junction integrity. Mol. Biol. Cell 2022, 33, ar14. [Google Scholar] [CrossRef]
- Cao, H.; Xu, E.; Liu, H.; Wan, L.; Lai, M. Epithelial-mesenchymal transition in colorectal cancer metastasis: A system review. Pathol. Res. Pract. 2015, 211, 557–569. [Google Scholar] [CrossRef]
- Mizukoshi, K.; Okazawa, Y.; Haeno, H.; Koyama, Y.; Sulidan, K.; Komiyama, H.; Saeki, H.; Ohtsuji, N.; Ito, Y.; Kojima, Y.; et al. Metastatic seeding of human colon cancer cell clusters expressing the hybrid epithelial/mesenchymal state. Int. J. Cancer 2020, 146, 2547–2562. [Google Scholar] [CrossRef]
- Athanasopoulou, A.; Aroukatos, P.; Nakas, D.; Repanti, M.; Papadaki, H.; Bravou, V. Decreased ezrin and paxillin expression in human urothelial bladder tumors correlate with tumor progression. Urol. Oncol. Semin. Orig. Investig. 2013, 31, 836–842. [Google Scholar] [CrossRef]
- Zouq, N.K.; Keeble, J.A.; Lindsay, J.; Valentijn, A.J.; Zhang, L.; Mills, D.; Turner, C.E.; Streuli, C.H.; Gilmore, A.P. FAK engages multiple pathways to maintain survival of fibroblasts and epithelia: Differential roles for paxillin and p130Cas. J. Cell Sci. 2009, 122, 357–367. [Google Scholar] [CrossRef]
- Simpson, C.D.; Anyiwe, K.; Schimmer, A.D. Anoikis resistance and tumor metastasis. Cancer Lett. 2008, 272, 177–185. [Google Scholar] [CrossRef]
- Wen, L.; Zhang, X.; Zhang, J.; Chen, S.; Ma, Y.; Hu, J.; Yue, T.; Wang, J.; Zhu, J.; Wu, T.; et al. Paxillin knockdown suppresses metastasis and epithelial-mesenchymal transition in colorectal cancer via the ERK signalling pathway. Oncol. Rep. 2020, 44, 1105–1115. [Google Scholar] [CrossRef]
- Cai, F.F.; Xu, H.R.; Yu, S.H.; Li, P.; Lu, Y.Y.; Chen, J.; Bi, Z.Q.; Sun, H.S.; Cheng, J.; Zhuang, H.Q.; et al. ADT-OH inhibits malignant melanoma metastasis in mice via suppressing CSE/CBS and FAK/Paxillin signaling pathway. Acta Pharmacol. Sin. 2022, 43, 1829–1842. [Google Scholar] [CrossRef] [PubMed]
- Badowski, C.; Pawlak, G.; Grichine, A.; Chabadel, A.; Oddou, C.; Jurdic, P.; Pfaff, M.; Albiges-Rizo, C.; Block, M.R. Paxillin phosphorylation controls invadopodia/podosomes spatiotemporal organization. Mol. Biol. Cell 2008, 19, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Wen, J. Abnormal level of paxillin in cervical cancer cells is involved in tumor progression and invasion. Acta Biochim. Pol. 2021, 68, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Mousson, A.; Legrand, M.; Steffan, T.; Vauchelles, R.; Carl, P.; Gies, J.P.; Lehmann, M.; Zuber, G.; De Mey, J.; Dujardin, D.; et al. Inhibiting FAK-Paxillin Interaction Reduces Migration and Invadopodia-Mediated Matrix Degradation in Metastatic Melanoma Cells. Cancers 2021, 13, 1871. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.; Chen, J.; Zhang, L.; Gao, P.; Hui, Y.; Xu, P.; Zhang, X.; Liu, H. Bit1 knockdown contributes to growth suppression as well as the decreases of migration and invasion abilities in esophageal squamous cell carcinoma via suppressing FAK-paxillin pathway. Mol. Cancer 2016, 15, 23. [Google Scholar] [CrossRef]
- Du, T.; Qu, Y.; Li, J.; Li, H.; Su, L.; Zhou, Q.; Yan, M.; Li, C.; Zhu, Z.; Liu, B. Maternal embryonic leucine zipper kinase enhances gastric cancer progression via the FAK/Paxillin pathway. Mol. Cancer 2014, 13, 100. [Google Scholar] [CrossRef]
- Mekhdjian, A.H.; Kai, F.; Rubashkin, M.G.; Prahl, L.S.; Przybyla, L.M.; McGregor, A.L.; Bell, E.S.; Barnes, J.M.; DuFort, C.C.; Ou, G.; et al. Integrin-mediated traction force enhances paxillin molecular associations and adhesion dynamics that increase the invasiveness of tumor cells into a three-dimensional extracellular matrix. Mol. Biol. Cell 2017, 28, 1467–1488. [Google Scholar] [CrossRef]
- Wu, J.N.; Lin, L.; Luo, S.B.; Qiu, X.Z.; Zhu, L.Y.; Chen, D.; Wei, E.D.; Fu, Z.H.; Qin, M.B.; Liang, Z.H.; et al. SphK1-driven autophagy potentiates focal adhesion paxillin-mediated metastasis in colorectal cancer. Cancer Med. 2021, 10, 6010–6021. [Google Scholar] [CrossRef]
- Li, D.; Ding, J.; Wang, X.; Wang, C.; Wu, T. Fibronectin promotes tyrosine phosphorylation of paxillin and cell invasiveness in the gastric cancer cell line AGS. Tumori 2009, 95, 769–779. [Google Scholar] [CrossRef]
- Li, X.; Wang, J. Mechanical tumor microenvironment and transduction: Cytoskeleton mediates cancer cell invasion and metastasis. Int. J. Biol. Sci. 2020, 16, 2014–2028. [Google Scholar] [CrossRef]
- Condeelis, J.; Singer, R.H.; Segall, J.E. The great escape: When cancer cells hijack the genes for chemotaxis and motility. Annu. Rev. Cell Dev. Biol. 2005, 21, 695–718. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E. Mechanisms of cancer cell invasion. Curr. Opin. Genet. Dev. 2005, 15, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Wyckoff, J.; Condeelis, J. Cell migration in tumors. Curr. Opin. Cell Biol. 2005, 17, 559–564. [Google Scholar] [CrossRef]
- Parsons, J.T. Focal adhesion kinase: The first ten years. J. Cell Sci. 2003, 116, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.O.; Ahn, C.H.; Yang, I.H.; Han, J.M.; Shin, J.A.; Cho, S.D.; Hong, S.D. Norcantharidin Suppresses YD-15 Cell Invasion Through Inhibition of FAK/Paxillin and F-Actin Reorganization. Molecules 2019, 24, 1928. [Google Scholar] [CrossRef]
- Wang, S.; Li, F.; Fan, H. Interferon-inducible protein, IFIX, has tumor-suppressive effects in oral squamous cell carcinoma. Sci. Rep. 2021, 11, 19593. [Google Scholar] [CrossRef]
- Bendas, G.; Borsig, L. Cancer cell adhesion and metastasis: Selectins, integrins, and the inhibitory potential of heparins. Int. J. Cell Biol. 2012, 2012, 676731. [Google Scholar] [CrossRef]
- Wang, C.; Yoo, Y.; Fan, H.; Kim, E.; Guan, K.L.; Guan, J.L. Regulation of Integrin beta 1 recycling to lipid rafts by Rab1a to promote cell migration. J. Biol. Chem. 2010, 285, 29398–29405. [Google Scholar] [CrossRef]
- Farahani, E.; Patra, H.K.; Jangamreddy, J.R.; Rashedi, I.; Kawalec, M.; Rao Pariti, R.K.; Batakis, P.; Wiechec, E. Cell adhesion molecules and their relation to (cancer) cell stemness. Carcinogenesis 2014, 35, 747–759. [Google Scholar] [CrossRef]
- Lisiak, N.; Paszel-Jaworska, A.; Toton, E.; Rubis, B.; Pakula, M.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Rybczynska, M. Semisynthetic oleanane triterpenoids inhibit migration and invasion of human breast cancer cells through downregulated expression of the ITGB1/PTK2/PXN pathway. Chem. Biol. Interact. 2017, 268, 136–147. [Google Scholar] [CrossRef]
- Mori, S.; Chang, J.T.; Andrechek, E.R.; Matsumura, N.; Baba, T.; Yao, G.; Kim, J.W.; Gatza, M.; Murphy, S.; Nevins, J.R. Anchorage-independent cell growth signature identifies tumors with metastatic potential. Oncogene 2009, 28, 2796–2805. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Salgia, R.; Li, J.L.; Griffin, J.D.; Howley, P.M. The bovine papillomavirus E6 protein binds to the LD motif repeats of paxillin and blocks its interaction with vinculin and the focal adhesion kinase. J. Biol. Chem. 1997, 272, 33373–33376. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.M.; Latifkar, A.; Cerione, R.A.; Antonyak, M.A. Cool-associated Tyrosine-phosphorylated Protein 1 Is Required for the Anchorage-independent Growth of Cervical Carcinoma Cells by Binding Paxillin and Promoting AKT Activation. J. Biol. Chem. 2017, 292, 3947–3957. [Google Scholar] [CrossRef] [PubMed]
- Jun, Q.; Zhiwei, W.; Lilin, M.; Jing, K.; Qichao, N. Effects of paxillin on HCT-8 human colorectal cancer cells. Hepatogastroenterology 2011, 58, 1951–1955. [Google Scholar] [CrossRef]
- Ma, X.; Biswas, A.; Hammes, S.R. Paxillin regulated genomic networks in prostate cancer. Steroids 2019, 151, 108463. [Google Scholar] [CrossRef]
- Sheibani, N.; Tang, Y.; Sorenson, C.M. Paxillin’s LD4 motif interacts with bcl-2. J. Cell. Physiol. 2008, 214, 655–661. [Google Scholar] [CrossRef]
- Liang, Z.; Li, X.; Chen, J.; Cai, H.; Zhang, L.; Li, C.; Tong, J.; Hu, W. PRC1 promotes cell proliferation and cell cycle progression by regulating p21/p27-pRB family molecules and FAK-paxillin pathway in non-small cell lung cancer. Transl. Cancer Res. 2019, 8, 2059–2072. [Google Scholar] [CrossRef]
- Reinartz, S.; Schumann, T.; Finkernagel, F.; Wortmann, A.; Jansen, J.M.; Meissner, W.; Krause, M.; Schworer, A.M.; Wagner, U.; Muller-Brusselbach, S.; et al. Mixed-polarization phenotype of ascites-associated macrophages in human ovarian carcinoma: Correlation of CD163 expression, cytokine levels and early relapse. Int. J. Cancer 2014, 134, 32–42. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, L.; Ma, K.; Zhao, Y.; Liu, X.; Wang, Y.; Liu, M.; Liang, S.; Zhu, H.; Xu, N. Polarization of macrophages in the tumor microenvironment is influenced by EGFR signaling within colon cancer cells. Oncotarget 2016, 7, 75366–75378. [Google Scholar] [CrossRef]
- Zhang, L.L.; Zhang, L.F.; Shi, Y.B. Down-regulated paxillin suppresses cell proliferation and invasion by inhibiting M2 macrophage polarization in colon cancer. Biol. Chem. 2018, 399, 1285–1295. [Google Scholar] [CrossRef]
- Lee, J.H.; Wittki, S.; Brau, T.; Dreyer, F.S.; Kratzel, K.; Dindorf, J.; Johnston, I.C.; Gross, S.; Kremmer, E.; Zeidler, R.; et al. HIV Nef, paxillin, and Pak1/2 regulate activation and secretion of TACE/ADAM10 proteases. Mol. Cell 2013, 49, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Minamiguchi, K.; Kumagai, H.; Masuda, T.; Kawada, M.; Ishizuka, M.; Takeuchi, T. Thiolutin, an inhibitor of HUVEC adhesion to vitronectin, reduces paxillin in HUVECs and suppresses tumor cell-induced angiogenesis. Int. J. Cancer 2001, 93, 307–316. [Google Scholar] [CrossRef] [PubMed]
- German, A.E.; Mammoto, T.; Jiang, E.; Ingber, D.E.; Mammoto, A. Paxillin controls endothelial cell migration and tumor angiogenesis by altering neuropilin 2 expression. J. Cell Sci. 2014, 127, 1672–1683. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Hu, H.; Wang, X. Clinically relevant concentrations of lidocaine inhibit tumor angiogenesis through suppressing VEGF/VEGFR2 signaling. Cancer Chemother. Pharmacol. 2019, 83, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Hailat, M.M.; Ebrahim, H.Y.; Mohyeldin, M.M.; Goda, A.A.; Siddique, A.B.; El Sayed, K.A. The tobacco cembranoid (1S,2E,4S,7E,11E)-2,7,11-cembratriene-4,6-diol as a novel angiogenesis inhibitory lead for the control of breast malignancies. Bioorganic Med. Chem. 2017, 25, 3911–3921. [Google Scholar] [CrossRef]
- Niu, G.; Wright, K.L.; Huang, M.; Song, L.; Haura, E.; Turkson, J.; Zhang, S.; Wang, T.; Sinibaldi, D.; Coppola, D.; et al. Constitutive Stat3 activity up-regulates VEGF expression and tumor angiogenesis. Oncogene 2002, 21, 2000–2008. [Google Scholar] [CrossRef]
- Yang, X.; Li, S.; Zhong, J.; Zhang, W.; Hua, X.; Li, B.; Sun, H. CD151 mediates netrin-1-induced angiogenesis through the Src-FAK-Paxillin pathway. J. Cell. Mol. Med. 2017, 21, 72–80. [Google Scholar] [CrossRef]
- Sp, N.; Kang, D.Y.; Joung, Y.H.; Park, J.H.; Kim, W.S.; Lee, H.K.; Song, K.D.; Park, Y.M.; Yang, Y.M. Nobiletin Inhibits Angiogenesis by Regulating Src/FAK/STAT3-Mediated Signaling through PXN in ER(+) Breast Cancer Cells. Int. J. Mol. Sci. 2017, 18, 935. [Google Scholar] [CrossRef]
- Maniotis, A.J.; Folberg, R.; Hess, A.; Seftor, E.A.; Gardner, L.M.; Pe’er, J.; Trent, J.M.; Meltzer, P.S.; Hendrix, M.J. Vascular channel formation by human melanoma cells in vivo and in vitro: Vasculogenic mimicry. Am. J. Pathol. 1999, 155, 739–752. [Google Scholar] [CrossRef]
- Spiliopoulos, K.; Peschos, D.; Batistatou, A.; Ntountas, I.; Agnantis, N.; Kitsos, G. Vasculogenic mimicry: Lessons from melanocytic tumors. In Vivo 2015, 29, 309–317. [Google Scholar]
- Lu, X.S.; Sun, W.; Ge, C.Y.; Zhang, W.Z.; Fan, Y.Z. Contribution of the PI3K/MMPs/Ln-5gamma2 and EphA2/FAK/Paxillin signaling pathways to tumor growth and vasculogenic mimicry of gallbladder carcinomas. Int. J. Oncol. 2013, 42, 2103–2115. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, W.; Zhang, W.Z.; Ge, C.Y.; Zhang, J.T.; Liu, Z.Y.; Fan, Y.Z. Inhibition of tumor vasculogenic mimicry and prolongation of host survival in highly aggressive gallbladder cancers by norcantharidin via blocking the ephrin type a receptor 2/focal adhesion kinase/paxillin signaling pathway. PLoS ONE 2014, 9, e96982. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, J.H.; Li, S.; Li, L.L.; Huang, M.; Zhang, Y.H.; Liu, Y.; Yang, Y.T.; Ding, R.; Ke, Y.Q. Histone deacetylase 3 expression correlates with vasculogenic mimicry through the phosphoinositide3-kinase/ERK-MMP-laminin5gamma2 signaling pathway. Cancer Sci. 2015, 106, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Zang, M.; Zhang, Y.; Zhang, B.; Hu, L.; Li, J.; Fan, Z.; Wang, H.; Su, L.; Zhu, Z.; Li, C.; et al. CEACAM6 promotes tumor angiogenesis and vasculogenic mimicry in gastric cancer via FAK signaling. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2015, 1852, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, Z.; Yuan, J.; Sun, L.; Lin, L.; Huang, N.; Bin, J.; Liao, Y.; Liao, W. Long non-coding RNA MALAT1 promotes gastric cancer tumorigenicity and metastasis by regulating vasculogenic mimicry and angiogenesis. Cancer Lett. 2017, 395, 31–44. [Google Scholar] [CrossRef]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337.e310. [Google Scholar] [CrossRef]
- Gajos-Michniewicz, A.; Czyz, M. WNT Signaling in Melanoma. Int. J. Mol. Sci. 2020, 21, 4852. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, M.; Xu, F.; Jiang, S. Wnt signaling in breast cancer: Biological mechanisms, challenges and opportunities. Mol. Cancer 2020, 19, 165. [Google Scholar] [CrossRef]
- Hayat, R.; Manzoor, M.; Hussain, A. Wnt signaling pathway: A comprehensive review. Cell Biol. Int. 2022, 46, 863–877. [Google Scholar] [CrossRef]
- Cooper, J.; Giancotti, F.G. Integrin Signaling in Cancer: Mechanotransduction, Stemness, Epithelial Plasticity, and Therapeutic Resistance. Cancer Cell 2019, 35, 347–367. [Google Scholar] [CrossRef]
- Wang, Q.; Jiang, D.; Ye, Q.; Zhou, W.; Ma, J.; Wang, C.; Geng, Z.; Chu, M.; Zheng, J.; Chen, H.; et al. A widely expressed free immunoglobulin kappa chain with a unique Vkappa4-1/Jkappa3 pattern promotes colon cancer invasion and metastasis by activating the integrin beta1/FAK pathway. Cancer Lett. 2022, 540, 215720. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.B.; Dorsett, K.A.; Hjelmeland, A.B.; Bellis, S.L. The ST6Gal-I sialyltransferase protects tumor cells against hypoxia by enhancing HIF-1alpha signaling. J. Biol. Chem. 2018, 293, 5659–5667. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Dorsett, K.A.; Trummell, H.Q.; Yang, E.S.; Oliver, P.G.; Bonner, J.A.; Buchsbaum, D.J.; Bellis, S.L. ST6Gal-I sialyltransferase promotes chemoresistance in pancreatic ductal adenocarcinoma by abrogating gemcitabine-mediated DNA damage. J. Biol. Chem. 2018, 293, 984–994. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.C.; Shyr, Y.M.; Liao, W.Y.; Chen, T.H.; Wang, S.E.; Lu, P.C.; Lin, P.Y.; Chen, Y.B.; Mao, W.Y.; Han, H.Y.; et al. Elevation of beta-galactoside alpha2,6-sialyltransferase 1 in a fructoseresponsive manner promotes pancreatic cancer metastasis. Oncotarget 2017, 8, 7691–7709. [Google Scholar] [CrossRef]
- Perez-Garay, M.; Arteta, B.; Llop, E.; Cobler, L.; Pages, L.; Ortiz, R.; Ferri, M.J.; de Bolos, C.; Figueras, J.; de Llorens, R.; et al. alpha2,3-Sialyltransferase ST3Gal IV promotes migration and metastasis in pancreatic adenocarcinoma cells and tends to be highly expressed in pancreatic adenocarcinoma tissues. Int. J. Biochem. Cell Biol. 2013, 45, 1748–1757. [Google Scholar] [CrossRef]
- Fu, C.W.; Tsai, H.E.; Chen, W.S.; Chang, T.T.; Chen, C.L.; Hsiao, P.W.; Li, W.S. Sialyltransferase Inhibitors Suppress Breast Cancer Metastasis. J. Med. Chem. 2021, 64, 527–542. [Google Scholar] [CrossRef]
- Hsieh, P.W.; Chang, Y.T.; Chuang, W.Y.; Shih, H.C.; Chiang, S.Z.; Wu, C.C. The synthesis and biologic evaluation of anti-platelet and cytotoxic beta-nitrostyrenes. Bioorganic Med. Chem. 2010, 18, 7621–7627. [Google Scholar] [CrossRef]
- He, Y.; Varadarajan, S.; Munoz-Planillo, R.; Burberry, A.; Nakamura, Y.; Nunez, G. 3,4-methylenedioxy-beta-nitrostyrene inhibits NLRP3 inflammasome activation by blocking assembly of the inflammasome. J. Biol. Chem. 2014, 289, 1142–1150. [Google Scholar] [CrossRef]
- Chen, I.H.; Shih, H.C.; Hsieh, P.W.; Chang, F.R.; Wu, Y.C.; Wu, C.C. HPW-RX40 restores anoikis sensitivity of human breast cancer cells by inhibiting integrin/FAK signaling. Toxicol. Appl. Pharmacol. 2015, 289, 330–340. [Google Scholar] [CrossRef]
- Syed, V. TGF-beta Signaling in Cancer. J. Cell. Biochem. 2016, 117, 1279–1287. [Google Scholar] [CrossRef]
- Wang, J.; Xu, Z.; Wang, Z.; Du, G.; Lun, L. TGF-beta signaling in cancer radiotherapy. Cytokine 2021, 148, 155709. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Li, B.; Zhang, S. MIR503HG: A potential diagnostic and therapeutic target in human diseases. Biomed. Pharmacother. 2023, 160, 114314. [Google Scholar] [CrossRef]
- Akhurst, R.J.; Hata, A. Targeting the TGFbeta signalling pathway in disease. Nat. Rev. Drug Discov. 2012, 11, 790–811. [Google Scholar] [CrossRef] [PubMed]
- Bokobza, S.M.; Ye, L.; Kynaston, H.G.; Jiang, W.G. Growth and differentiation factor-9 promotes adhesive and motile capacity of prostate cancer cells by up-regulating FAK and Paxillin via Smad dependent pathway. Oncol. Rep. 2010, 24, 1653–1659. [Google Scholar] [CrossRef]
- Guerra, B.; Issinger, O.G. Protein kinase CK2 in human diseases. Curr. Med. Chem. 2008, 15, 1870–1886. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hwan Kim, S. CK2 inhibitor CX-4945 blocks TGF-beta1-induced epithelial-to-mesenchymal transition in A549 human lung adenocarcinoma cells. PLoS ONE 2013, 8, e74342. [Google Scholar] [CrossRef]
- Maeda, D.; Kubo, T.; Kiya, K.; Kawai, K.; Matsuzaki, S.; Kobayashi, D.; Fujiwara, T.; Katayama, T.; Hosokawa, K. Periostin is induced by IL-4/IL-13 in dermal fibroblasts and promotes RhoA/ROCK pathway-mediated TGF-beta1 secretion in abnormal scar formation. J. Plast. Surg. Hand Surg. 2019, 53, 288–294. [Google Scholar] [CrossRef]
- Yu, J.; Zhou, Z.; Wei, Z.; Wu, J.; OuYang, J.; Huang, W.; He, Y.; Zhang, C. FYN promotes gastric cancer metastasis by activating STAT3-mediated epithelial-mesenchymal transition. Transl. Oncol. 2020, 13, 100841. [Google Scholar] [CrossRef]
- Lu, Q.; Lai, Y.; Zhang, H.; Ren, K.; Liu, W.; An, Y.; Yao, J.; Fan, H. Hesperetin Inhibits TGF-beta1-Induced Migration and Invasion of Triple Negative Breast Cancer MDA-MB-231 Cells via Suppressing Fyn/Paxillin/RhoA Pathway. Integr. Cancer Ther. 2022, 21, 15347354221086900. [Google Scholar] [CrossRef]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef]
- Asl, E.R.; Amini, M.; Najafi, S.; Mansoori, B.; Mokhtarzadeh, A.; Mohammadi, A.; Lotfinejad, P.; Bagheri, M.; Shirjang, S.; Lotfi, Z.; et al. Interplay between MAPK/ERK signaling pathway and MicroRNAs: A crucial mechanism regulating cancer cell metabolism and tumor progression. Life Sci. 2021, 278, 119499. [Google Scholar] [CrossRef] [PubMed]
- Park, H.B.; Baek, K.H. E3 ligases and deubiquitinating enzymes regulating the MAPK signaling pathway in cancers. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2022, 1877, 188736. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; O’Malley, K.; Wang, Z.; Raj, G.V.; Defranco, D.B.; Hammes, S.R. Paxillin regulates androgen- and epidermal growth factor-induced MAPK signaling and cell proliferation in prostate cancer cells. J. Biol. Chem. 2010, 285, 28787–28795. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; De Castro, I.; Defranco, D.B.; Deng, F.M.; Melamed, J.; Kapur, P.; Raj, G.V.; Rossi, R.; Hammes, S.R. Paxillin mediates extranuclear and intranuclear signaling in prostate cancer proliferation. J. Clin. Investig. 2012, 122, 2469–2481. [Google Scholar] [CrossRef]
- Sun, Y.; Yu, X.; Li, M.; Liu, J. P44/42 MAPK signal pathway-mediated hyperphosphorylation of paxillin and redistribution of E-cadherin was involved in microcystin-LR-reduced cellular adhesion in a human liver cell line. Chemosphere 2018, 200, 594–602. [Google Scholar] [CrossRef]
- Aoki, M.; Fujishita, T. Oncogenic Roles of the PI3K/AKT/mTOR Axis. Viruses Genes Cancer 2017, 407, 153–189. [Google Scholar] [CrossRef]
- Tan, X.; Gong, L.; Li, X.; Zhang, X.; Sun, J.; Luo, X.; Wang, Q.; Chen, J.; Xie, L.; Han, S. Promethazine inhibits proliferation and promotes apoptosis in colorectal cancer cells by suppressing the PI3K/AKT pathway. Biomed. Pharmacother. 2021, 143, 112174. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Abak, A.; Tondro Anamag, F.; Shoorei, H.; Majidpoor, J.; Taheri, M. The emerging role of non-coding RNAs in the regulation of PI3K/AKT pathway in the carcinogenesis process. Biomed. Pharmacother. 2021, 137, 111279. [Google Scholar] [CrossRef]
- Metalli, D.; Lovat, F.; Tripodi, F.; Genua, M.; Xu, S.Q.; Spinelli, M.; Alberghina, L.; Vanoni, M.; Baffa, R.; Gomella, L.G.; et al. The insulin-like growth factor receptor I promotes motility and invasion of bladder cancer cells through Akt- and mitogen-activated protein kinase-dependent activation of paxillin. Am. J. Pathol. 2010, 176, 2997–3006. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chen, J.; He, L.; Stiles, B.L. PTEN: Tumor Suppressor and Metabolic Regulator. Front. Endocrinol. 2018, 9, 338. [Google Scholar] [CrossRef]
- Zhang, L.L.; Mu, G.G.; Ding, Q.S.; Li, Y.X.; Shi, Y.B.; Dai, J.F.; Yu, H.G. Phosphatase and Tensin Homolog (PTEN) Represses Colon Cancer Progression through Inhibiting Paxillin Transcription via PI3K/AKT/NF-kappaB Pathway. J. Biol. Chem. 2015, 290, 15018–15029. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Calin, G.A.; Lopez-Berestein, G.; Sood, A.K. miRNA Deregulation in Cancer Cells and the Tumor Microenvironment. Cancer Discov. 2016, 6, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xu, D.; Xu, X.; Xue, Q.; Gao, X.; Tang, C. MiR-216b regulates the tumorigenesis of gastric cancer by targeting PXN. Pathol. Res. Pract. 2021, 218, 153325. [Google Scholar] [CrossRef]
- Li, D.; Li, Z.; Xiong, J.; Gong, B.; Zhang, G.; Cao, C.; Jie, Z.; Liu, Y.; Cao, Y.; Yan, Y.; et al. MicroRNA-212 functions as an epigenetic-silenced tumor suppressor involving in tumor metastasis and invasion of gastric cancer through down-regulating PXN expression. Am. J. Cancer Res. 2015, 5, 2980–2997. [Google Scholar]
- Chen, D.L.; Wang, D.S.; Wu, W.J.; Zeng, Z.L.; Luo, H.Y.; Qiu, M.Z.; Ren, C.; Zhang, D.S.; Wang, Z.Q.; Wang, F.H.; et al. Overexpression of paxillin induced by miR-137 suppression promotes tumor progression and metastasis in colorectal cancer. Carcinogenesis 2013, 34, 803–811. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, H.; Hu, X.; Li, W. Knockdown of long non-coding RNA XIST inhibits cell viability and invasion by regulating miR-137/PXN axis in non-small cell lung cancer. Int. J. Biol. Macromol. 2018, 111, 623–631. [Google Scholar] [CrossRef]
- Li, P.; Wang, L.; Li, P.; Hu, F.; Cao, Y.; Tang, D.; Ye, G.; Li, H.; Wang, D. Silencing lncRNA XIST exhibits antiproliferative and proapoptotic effects on gastric cancer cells by up-regulating microRNA-132 and down-regulating PXN. Aging 2020, 13, 14469–14481. [Google Scholar] [CrossRef]
- Du, C.; Wang, Y.; Zhang, Y.; Zhang, J.; Zhang, L.; Li, J. LncRNA DLX6-AS1 Contributes to Epithelial-Mesenchymal Transition and Cisplatin Resistance in Triple-negative Breast Cancer via Modulating Mir-199b-5p/Paxillin Axis. Cell Transplant. 2020, 29, 963689720929983. [Google Scholar] [CrossRef]
- Wu, D.W.; Chuang, C.Y.; Lin, W.L.; Sung, W.W.; Cheng, Y.W.; Lee, H. Paxillin promotes tumor progression and predicts survival and relapse in oral cavity squamous cell carcinoma by microRNA-218 targeting. Carcinogenesis 2014, 35, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, R.; Okuzaki, D.; Okada, M.; Oneyama, C. MicroRNA-27b suppresses tumor progression by regulating ARFGEF1 and focal adhesion signaling. Cancer Sci. 2016, 107, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Liu, T.; Xu, H.; Wang, Y.; Yang, T.; Liu, L. LncRNA FIRRE promotes the proliferation and metastasis of hepatocellular carcinoma by regulating the expression of PXN through interacting with MBNL3. Biochem. Biophys. Res. Commun. 2022, 625, 188–195. [Google Scholar] [CrossRef]
- Xu, S.; Li, X.; Li, L.; Wang, Y.; Geng, C.; Guo, F.; Zhang, T.; Du, A.; Lu, Z.; Hui, H.; et al. CTCF-silenced miR-137 contributes to EMT and radioresistance in esophageal squamous cell carcinoma. Cancer Cell Int. 2021, 21, 155. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ding, X.; Liu, B.; Li, M.; Chang, Y.; Shen, H.; Xie, S.M.; Xing, L.; Li, Y. ETV4 overexpression promotes progression of non-small cell lung cancer by upregulating PXN and MMP1 transcriptionally. Mol. Carcinog. 2020, 59, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Nocula-Lugowska, M.; Lugowski, M.; Salgia, R.; Kossiakoff, A.A. Engineering Synthetic Antibody Inhibitors Specific for LD2 or LD4 Motifs of Paxillin. J. Mol. Biol. 2015, 427, 2532–2547. [Google Scholar] [CrossRef] [PubMed]
- Deramaudt, T.B.; Dujardin, D.; Noulet, F.; Martin, S.; Vauchelles, R.; Takeda, K.; Ronde, P. Altering FAK-paxillin interactions reduces adhesion, migration and invasion processes. PLoS ONE 2014, 9, e92059. [Google Scholar] [CrossRef]
- Lee, B.Y.; Timpson, P.; Horvath, L.G.; Daly, R.J. FAK signaling in human cancer as a target for therapeutics. Pharmacol. Ther. 2015, 146, 132–149. [Google Scholar] [CrossRef]
- Sulzmaier, F.J.; Jean, C.; Schlaepfer, D.D. FAK in cancer: Mechanistic findings and clinical applications. Nat. Rev. Cancer 2014, 14, 598–610. [Google Scholar] [CrossRef]
- Wu, G.S.; Song, Y.L.; Yin, Z.Q.; Guo, J.J.; Wang, S.P.; Zhao, W.W.; Chen, X.P.; Zhang, Q.W.; Lu, J.J.; Wang, Y.T. Ganoderiol A-enriched extract suppresses migration and adhesion of MDA-MB-231 cells by inhibiting FAK-SRC-paxillin cascade pathway. PLoS ONE 2013, 8, e76620. [Google Scholar] [CrossRef]
- Chen, J.Y.; Tang, Y.A.; Huang, S.M.; Juan, H.F.; Wu, L.W.; Sun, Y.C.; Wang, S.C.; Wu, K.W.; Balraj, G.; Chang, T.T.; et al. A novel sialyltransferase inhibitor suppresses FAK/paxillin signaling and cancer angiogenesis and metastasis pathways. Cancer Res. 2011, 71, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, J.; Shao, J.; Gao, Y.; Xu, J.; Yu, S.; Liu, Z.; Jia, L. The unique pharmacological characteristics of mifepristone (RU486): From terminating pregnancy to preventing cancer metastasis. Med. Res. Rev. 2014, 34, 979–1000. [Google Scholar] [CrossRef] [PubMed]
- Llaguno-Munive, M.; Medina, L.A.; Jurado, R.; Romero-Pina, M.; Garcia-Lopez, P. Mifepristone improves chemo-radiation response in glioblastoma xenografts. Cancer Cell Int. 2013, 13, 29. [Google Scholar] [CrossRef] [PubMed]
- Engman, M.; Varghese, S.; Lagerstedt Robinson, K.; Malmgren, H.; Hammarsjo, A.; Bystrom, B.; Lalitkumar, P.G.; Gemzell-Danielsson, K. GSTM1 gene expression correlates to leiomyoma volume regression in response to mifepristone treatment. PLoS ONE 2013, 8, e80114. [Google Scholar] [CrossRef]
- Yu, S.; Yang, X.; Zhu, Y.; Xie, F.; Lu, Y.; Yu, T.; Yan, C.; Shao, J.; Gao, Y.; Mo, F.; et al. Systems pharmacology of mifepristone (RU486) reveals its 47 hub targets and network: Comprehensive analysis and pharmacological focus on FAK-Src-Paxillin complex. Sci. Rep. 2015, 5, 7830. [Google Scholar] [CrossRef]
- Huang, S.M.; Hsu, P.C.; Chen, M.Y.; Li, W.S.; More, S.V.; Lu, K.T.; Wang, Y.C. The novel indole compound SK228 induces apoptosis and FAK/Paxillin disruption in tumor cell lines and inhibits growth of tumor graft in the nude mouse. Int. J. Cancer 2012, 131, 722–732. [Google Scholar] [CrossRef]
- Xiao, P.; Ma, T.; Zhou, C.; Xu, Y.; Liu, Y.; Zhang, H. Anticancer effect of docetaxel induces apoptosis of prostate cancer via the cofilin-1 and paxillin signaling pathway. Mol. Med. Rep. 2016, 13, 4079–4084. [Google Scholar] [CrossRef]
- Li, W.; Yu, X.; Ma, X.; Xie, L.; Xia, Z.; Liu, L.; Yu, X.; Wang, J.; Zhou, H.; Zhou, X.; et al. Deguelin attenuates non-small cell lung cancer cell metastasis through inhibiting the CtsZ/FAK signaling pathway. Cell. Signal. 2018, 50, 131–141. [Google Scholar] [CrossRef]
- Luo, W.W.; Zhao, W.W.; Lu, J.J.; Wang, Y.T.; Chen, X.P. Cucurbitacin B suppresses metastasis mediated by reactive oxygen species (ROS) via focal adhesion kinase (FAK) in breast cancer MDA-MB-231 cells. Chin. J. Nat. Med. 2018, 16, 10–19. [Google Scholar] [CrossRef]
- Wu, D.W.; Wu, T.C.; Wu, J.Y.; Cheng, Y.W.; Chen, Y.C.; Lee, M.C.; Chen, C.Y.; Lee, H. Phosphorylation of paxillin confers cisplatin resistance in non-small cell lung cancer via activating ERK-mediated Bcl-2 expression. Oncogene 2014, 33, 4385–4395. [Google Scholar] [CrossRef]
- Du, C.; Wang, X.; Zhang, J.; Liu, X.; Zhu, J.; Liu, Y. Paxillin is positively correlated with the clinicopathological factors of colorectal cancer, and knockdown of Paxillin improves sensitivity to cetuximab in colorectal cancer cells. Oncol. Rep. 2016, 35, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.W.; Chen, C.Y.; Chu, C.L.; Lee, H. Paxillin confers resistance to tyrosine kinase inhibitors in EGFR-mutant lung cancers via modulating BIM and Mcl-1 protein stability. Oncogene 2016, 35, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.W.; Huang, C.C.; Chang, S.W.; Chen, T.H.; Lee, H. Bcl-2 stabilization by paxillin confers 5-fluorouracil resistance in colorectal cancer. Cell Death Differ. 2015, 22, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Ashaie, M.A.; Islam, R.A.; Kamaruzman, N.I.; Ibnat, N.; Tha, K.K.; Chowdhury, E.H. Targeting Cell Adhesion Molecules via Carbonate Apatite-Mediated Delivery of Specific siRNAs to Breast Cancer Cells In Vitro and In Vivo. Pharmaceutics 2019, 11, 309. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yin, S.; Zhang, L.; Shi, K.; Tang, J.; Zhang, Z.; He, Q. A tumor-activatable particle with antimetastatic potential in breast cancer via inhibiting the autophagy-dependent disassembly of focal adhesion. Biomaterials 2018, 168, 1–9. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Huang, X.; Luo, W.; Liu, X.; Chen, W. The Role of Paxillin Aberrant Expression in Cancer and Its Potential as a Target for Cancer Therapy. Int. J. Mol. Sci. 2023, 24, 8245. https://doi.org/10.3390/ijms24098245
Liu W, Huang X, Luo W, Liu X, Chen W. The Role of Paxillin Aberrant Expression in Cancer and Its Potential as a Target for Cancer Therapy. International Journal of Molecular Sciences. 2023; 24(9):8245. https://doi.org/10.3390/ijms24098245
Chicago/Turabian StyleLiu, Weixian, Xinxian Huang, Weizhao Luo, Xinguang Liu, and Weichun Chen. 2023. "The Role of Paxillin Aberrant Expression in Cancer and Its Potential as a Target for Cancer Therapy" International Journal of Molecular Sciences 24, no. 9: 8245. https://doi.org/10.3390/ijms24098245
APA StyleLiu, W., Huang, X., Luo, W., Liu, X., & Chen, W. (2023). The Role of Paxillin Aberrant Expression in Cancer and Its Potential as a Target for Cancer Therapy. International Journal of Molecular Sciences, 24(9), 8245. https://doi.org/10.3390/ijms24098245