
Pre-Diagnostic Saliva Microbiota of School-Aged Children Who Developed Type 1 Diabetes or Inflammatory Bowel Diseases

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Background Data of Obtained Saliva Samples
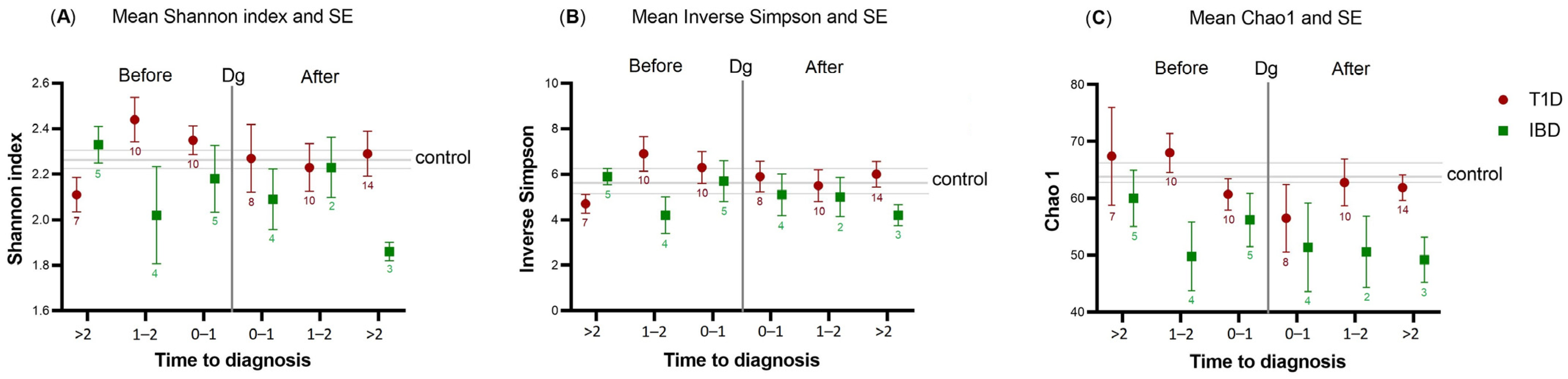
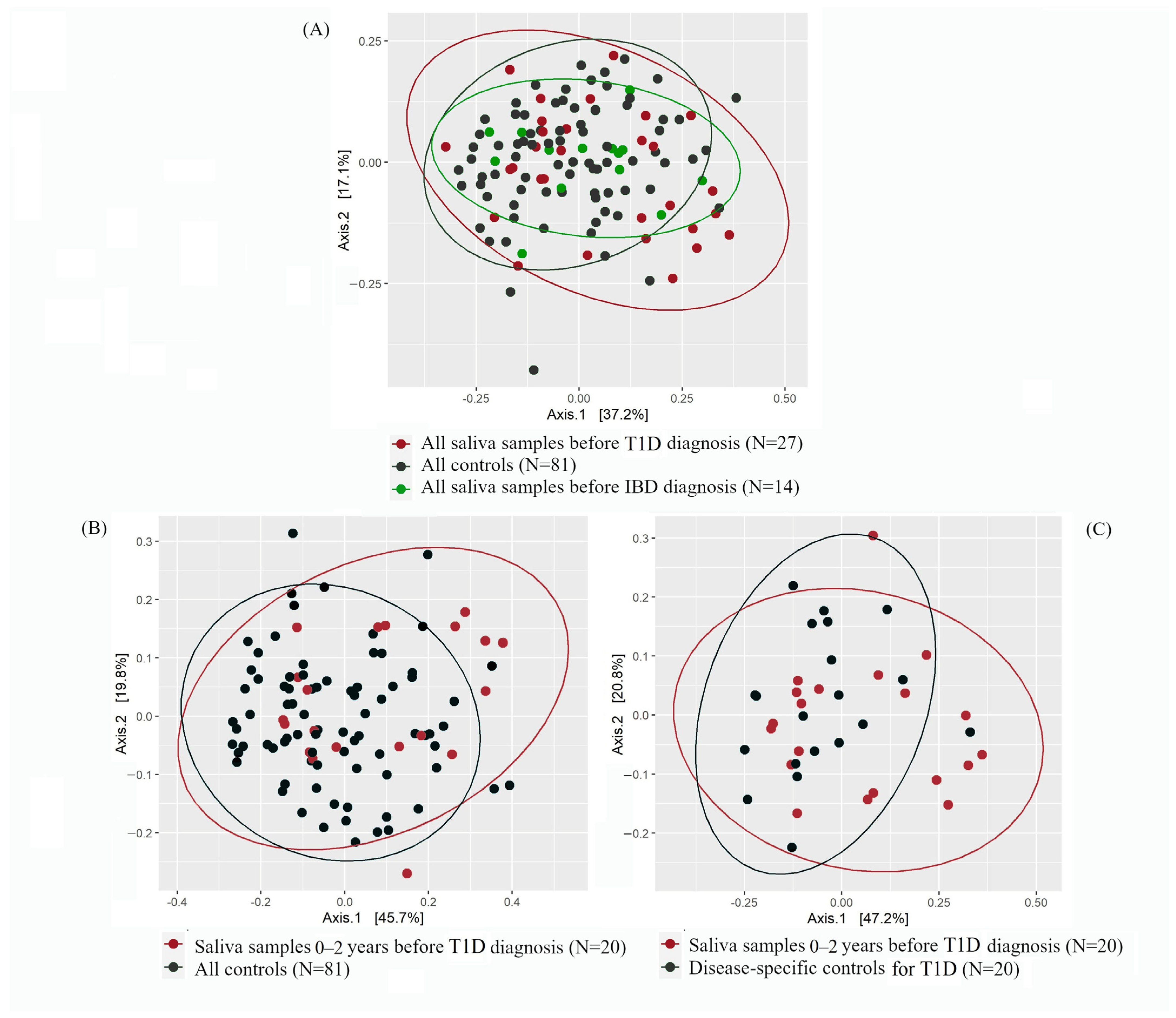
2.2. Alpha- and Beta-Diversity of T1D or IBD Cases vs. Controls
2.2.1. Saliva Microbiota Composition and Differential Abundance
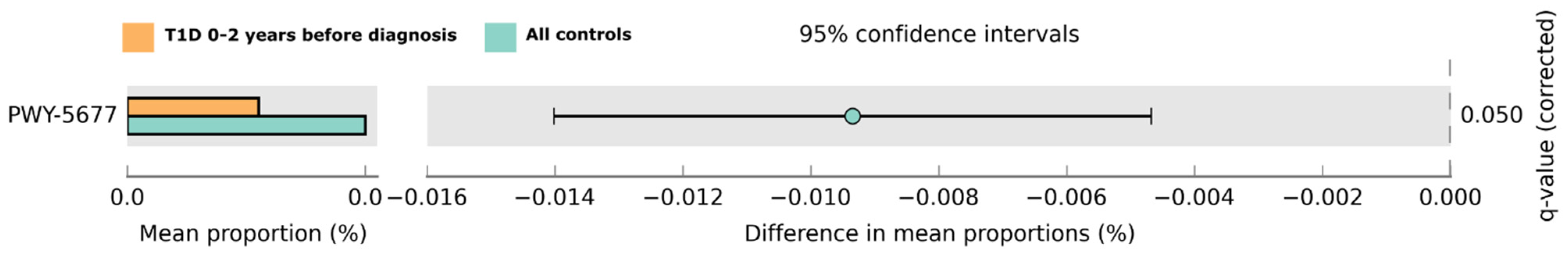
2.2.2. Functional Prediction of Saliva Microbiota
3. Discussion
3.1. Saliva Microbiota and T1D
3.2. Saliva Microbiota and IBD
3.3. Strengths and Limitations of the Study
4. Materials and Methods
4.1. Data Sources for the Study
4.2. DNA Extraction, Amplification, and Sequencing
4.3. Bioinformatics and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| T1D | Type 1 diabetes mellitus |
| DPR | Drug Purchase Register |
| Fin-HIT | Finnish Health in Teens |
| IBD | Inflammatory Bowel Diseases |
| PCoA | Principal Coordinate Analysis |
| SRR | Special Reimbursement Register |
References
- Räisänen, L.; Viljakainen, H.; Sarkkola, C.; Kolho, K.-L. Perinatal risk factors for pediatric onset type 1 diabetes, autoimmune thyroiditis, juvenile idiopathic arthritis, and inflammatory bowel diseases. Eur. J. Pediatr. 2021, 180, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Paschou, S.A.; Papadopoulou-Marketou, N.; Chrousos, G.; Kanaka-Gantenbein, C. On type 1 diabetes mellitus pathogenesis. Endocr. Connect. 2017, 7, R38–R46. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, A. Autoreactive T cells in type 1 diabetes. J. Clin. Investig. 2017, 127, 2881–2891. [Google Scholar] [CrossRef]
- Mitsuyama, K. Antibody markers in the diagnosis of inflammatory bowel disease. World J. Gastroenterol. 2016, 22, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Murri, M.; Leiva, I.; Gomez-Zumaquero, J.M.; Tinahones, F.J.; Cardona, F.; Soriguer, F.; Queipo-Ortuño, M.I. Gut microbiota in children with type 1 diabetes differs from that in healthy children: A case-control study. BMC Med. 2013, 11, 46. [Google Scholar] [CrossRef]
- Kolho, K.-L.; Korpela, K.; Jaakkola, T.; Pichai, M.V.A.; Zoetendal, E.G.; Salonen, A.; de Vos, W.M. Fecal Microbiota in Pediatric Inflammatory Bowel Disease and Its Relation to Inflammation. Am. J. Gastroenterol. 2015, 110, 921–930. [Google Scholar] [CrossRef]
- Knip, M.; Siljander, H. The role of the intestinal microbiota in type 1 diabetes mellitus. Nat. Rev. Endocr. 2016, 12, 154–167. [Google Scholar] [CrossRef]
- Pachoński, M.; Koczor-Rozmus, A.; Mocny-Pachońska, K.; Łanowy, P.; Mertas, A.; Jarosz-Chobot, P. Oral microbiota in children with type 1 diabetes mellitus. Pediatr. Endocrinol. Diabetes Metab. 2021, 27, 100–108. [Google Scholar] [CrossRef]
- Moskovitz, M.; Nassar, M.; Moriel, N.; Cher, A.; Faibis, S.; Ram, D.; Zangen, D.; Yassour, M.; Steinberg, D. Characterization of the Oral Microbiome Among Children With Type 1 Diabetes Compared With Healthy Children. Front. Microbiol. 2021, 12, 756808. [Google Scholar] [CrossRef]
- Docktor, M.J.; Paster, B.J.; Abramowicz, S.; Ingram, J.; Wang, Y.E.; Correll, M.; Jiang, H.; Cotton, S.L.; Kokaras, A.S.; Bousvaros, A. Alterations in diversity of the oral microbiome in pediatric inflammatory bowel disease. Inflamm. Bowel Dis. 2012, 18, 935–942. [Google Scholar] [CrossRef]
- Elmaghrawy, K.; Hussey, S.; Moran, G.P. The Oral Microbiome in Pediatric IBD: A Source of Pathobionts or Biomarkers? Front. Pediatr. 2021, 8, 620254. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Li, J.; Gao, F. New insights into insulin: The anti-inflammatory effect and its clinical relevance. World J. Diabetes. 2014, 5, 89. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Siljander, H.; Vatanen, T.; Hyötyläinen, T.; Hämäläinen, A.-M.; Peet, A.; Tillmann, V.; Pöhö, P.; Mattila, I.; et al. The Dynamics of the Human Infant Gut Microbiome in Development and in Progression toward Type 1 Diabetes. Cell Host Microbe 2015, 17, 260–273. [Google Scholar] [CrossRef]
- Gupta, S.; Nayak, M.T.; Sunitha, J.D.; Dawar, G.; Sinha, N.; Rallan, N.S. Correlation of salivary glucose level with blood glucose level in diabetes mellitus. J. Oral Maxillofac. Pathol. 2017, 21, 334–339. [Google Scholar]
- Insel, R.A.; Dunne, J.L.; Atkinson, M.A.; Chiang, J.L.; Dabelea, D.; Gottlieb, P.A.; Greenbaum, C.J.; Herold, K.C.; Krischer, J.P.; Lernmark, Å.; et al. Staging Presymptomatic Type 1 Diabetes: A Scientific Statement of JDRF, the Endocrine Society, and the American Diabetes Association. Diabetes Care 2015, 38, 1964–1974. [Google Scholar] [CrossRef]
- Wade, W.G. Resilience of the oral microbiome. Periodontology 2000 2021, 86, 113–122. [Google Scholar] [CrossRef]
- Börnigen, D.; Morgan, X.C.; Franzosa, E.A.; Ren, B.; Xavier, R.J.; Garrett, W.S.; Huttenhower, C. Functional profiling of the gut microbiome in disease-associated inflammation. Genome Med. 2013, 5, 65. [Google Scholar] [CrossRef] [PubMed]
- Raju, S.C.; Lagström, S.; Ellonen, P.; De Vos, W.M.; Eriksson, J.G.; Weiderpass, E.; Rounge, T.B. Gender-specific associations between saliva microbiota and body size. Front. Microbiol. 2019, 10, 767. [Google Scholar] [CrossRef]
- Guan, X.; Li, W.; Meng, H. A double-edged sword: Role of butyrate in the oral cavity and the gut. Mol. Oral Microbiol. 2021, 2, 121–131. [Google Scholar] [CrossRef]
- Buckner, J.H. Mechanisms of impaired regulation by CD4 + CD25 + FOXP3 + regulatory T cells in human autoimmune diseases. Nat. Rev. Immunol. Nat. Rev. 2010, 10, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Somineni, H.K.; Weitzner, J.H.; Venkateswaran, S.; Dodd, A.; Prince, J.; Karikaran, A.; Sauer, C.G.; Abramowicz, S.; Zwick, M.E.; Cutler, D.J.; et al. Site- and Taxa-Specific Disease-Associated Oral Microbial Structures Distinguish Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2021, 27, 1889–1900. [Google Scholar] [CrossRef]
- Stahringer, S.S.; Clemente, J.C.; Corley, R.P.; Hewitt, J.; Knights, D.; Walters, W.A.; Knight, R.; Krauter, K.S. Nurture trumps nature in a longitudinal survey of salivary bacterial communities in twins from early adolescence to early adulthood. Genome Res. 2012, 22, 2146–2152. [Google Scholar] [CrossRef]
- Shaw, L.; Ribeiro AL, R.; Levine, A.P.; Pontikos, N.; Balloux, F.; Segal, A.W.; Roberts, A.P.; Smith, A.M. The Human Salivary Microbiome Is Shaped by Shared Environment Rather than Genetics: Evidence from a Large Family of Closely Related Individuals ed C M Fraser. mBio 2017, 8, e01237-17. [Google Scholar] [CrossRef]
- Liu, X.; Tong, X.; Zhu, J.; Tian, L.; Jie, Z.; Zou, Y.; Lin, X.; Liang, H.; Li, W.; Ju, Y.; et al. Metagenome-genome-wide association studies reveal human genetic impact on the oral microbiome. Cell Discov. 2021, 7, 117. [Google Scholar] [CrossRef]
- Lommi, S.; Manzoor, M.; Engberg, E.; Agrawal, N.; Lakka, T.A.; Leinonen, J.; Kolho, K.-L.; Viljakainen, H. The Composition and Functional Capacities of Saliva Microbiota Differ Between Children with Low and High Sweet Treat Consumption. Front. Nutr. 2022, 9, 864687. [Google Scholar] [CrossRef] [PubMed]
- Räisänen, L.; Lommi, S.; Engberg, E.; Kolho, K.-L.; Viljakainen, H. Central obesity in school-aged children increases the likelihood of developing paediatric autoimmune diseases. Pediatr. Obes. 2021, 80, 2115–2123. [Google Scholar] [CrossRef]
- Manzoor, M.; Lommi, S.; Furuholm, J.; Sarkkola, C.; Engberg, E.; Raju, S.; Viljakainen, H. High abundance of sugar metabolisers in saliva of children with caries. Sci. Rep. 2021, 11, 4424. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- de Figueiredo, R.A.O.; Simola-Ström, S.; Rounge, T.B.; Viljakainen, H.; Eriksson, J.G.; Roos, E.; Weiderpass, E. Cohort Profile: The Finnish Health in Teens (Fin-HIT) study: A population-based study. Int. J. Epidemiol. 2019, 48, 23–24h. [Google Scholar] [CrossRef] [PubMed]
- Rylander-Rudqvist, T.; Håkansson, N.; Tybring, G.; Wolk, A. Quality and quantity of saliva DNA obtained from the self-administrated oragene method—A pilot study on the cohort of Swedish men. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1742–1745. [Google Scholar] [CrossRef] [PubMed]
- Iwasiow, R.M.; Desbois, A.; Birnboim, H.C. Long-term stability of DNA from saliva samples stored in the Oragene® self-collection kit. DNA Genotek 2011, 3, PD-WP-005. Available online: https://www.dnagenotek.com/US/pdf/PD-WP-005.pdf (accessed on 4 April 2023).
- Furu, K.; Wettermark, B.; Andersen, M.; Martikainen, J.E.; Almarsdottir, A.B.; Sorensen, H.T. The Nordic Countries as a Cohort for Pharmacoepidemiological Research. Basic Clin. Pharmacol. Toxicol. 2010, 106, 86–94. [Google Scholar] [CrossRef]
- Raju, S.C.; Lagström, S.; Ellonen, P.; de Vos, W.M.; Eriksson, J.G.; Weiderpass, E.; Rounge, T.B. Reproducibility and repeatability of six high-throughput 16S rDNA sequencing protocols for microbiota profiling. J. Microbiol. Methods. 2018, 147, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Timing of Saliva Samples | >2 Years before Diagnosis | 0–2 Years before Diagnosis | 0–2 Years after Diagnosis | >2 Years after Diagnosis | Total |
|---|---|---|---|---|---|
| T1D | 7 | 20 | 18 | 14 | 59 |
| IBD | 5 | 9 | 6 | 3 | 23 |
| Controls | 12 | 28 | 24 | 17 | 81 |
| Children Who Developed T1D (n = 52) | Children Who Developed IBD (n = 21) | Children without Autoimmune Diseases (n = 79) | p-Value a | p-Value b | |
|---|---|---|---|---|---|
| Age at baseline, mean ± SD | 11.0 ± 1.0 | 11.3 ± 0.6 | 11.2 ± 1.0 | 0.457 | 0.521 c |
| Age of diagnosis, mean ± SD | 11.2 ± 2.8 | 12.2 ± 2.8 | |||
| Age at the end of December 2018, mean ± SD | 16.3 ± 1.6 | 16.4 ± 1.1 | 16.3 ± 1.5 | 0.994 | 0.662 c |
| Sex, n (%) | 0.672 | 0.606 d | |||
| Boy | 31 (59.6) | 12 (57.1) | 50 (63.3) | ||
| Girl | 21 (40.4) | 9 (42.9) | 29 (36.7) | ||
| BMI at baseline, n (%) | 0.918 | 0.654 d | |||
| Normal | 35 (67.3) | 16 (76.2) | 67 (84.8) | ||
| Overweight/obese | 5 (9.6) | 3 (14.3) | 9 (11.4) | ||
| Missing | 12 (23.1) | 2 (9.5) | 3 (3.8) | ||
| Waist-to-height ratio (cm/cm) at baseline, n (%) | |||||
| Normal (<0.5) | 35 (67.3) | 18 (85.7) | 69 (87.3) | 0.350 | 0.970 d |
| Central obesity (≥0.5) | 4 (7.7) | 1 (4.8) | 4 (5.1) | ||
| Missing | 13 (25.0) | 2 (9.5) | 6 (7.6) | ||
| Saliva samples available for analyses at baseline/first follow-up, n | |||||
| Baseline | 41 | 18 | 59 | ||
| First follow-up | 18 | 5 | 22 | ||
| Total | 59 | 23 | 81 | ||
| Antibiotic exposure (any type) prior to the collection of saliva samples, n (%) | 0.676 | 0.534 d | |||
| No exposures | 5 (8.5) | 3 (13.0) | 5 (6.2) | ||
| ≤3 months | 2 (3.4) | 1 (4.3) | 5 (6.2) | ||
| >3 months | 52 (88.1) | 19 (82.6) | 71 (87.6) |
| Abundance (%) | All Controls (n = 81) | Before T1D Diagnosis (n = 27) | Before IBD Diagnosis (n = 14) | After T1D Diagnosis (n = 32) | After IBD Diagnosis (n = 9) |
|---|---|---|---|---|---|
| Firmicutes | 57.3 | 55.6 | 55.4 | 54.6 | 51.4 |
| Bacteroidetes | 16.2 | 13.6 | 15.3 | 14.5 | 19.1 |
| Proteobacteria | 12.4 | 16.7 | 15.2 | 17.4 | 16.4 |
| Actinobacteria | 7.6 | 8.0 | 9.3 | 7.5 | 8.7 |
| Fusobacteria | 3.1 | 3.1 | 2.4 | 3.2 | 1.8 |
| Patescibacteria | 2.6 | 2.3 | 1.9 | 2.2 | 2.1 |
| Other | 0.7 | 0.6 | 0.5 | 0.6 | 0.6 |
| TOTAL | 100 | 99.9 | 99.9 | 100 | 100 |
| 0–2 Years before Diagnosis T1D Samples (n = 20) vs. All Control Samples (n = 81) | 0–2 Years before Diagnosis T1D Samples (n = 20) vs. Disease-Specific Controls (n = 20) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Genera | Base Mean | Log2 Fold Change | p-Value | Genera | Base Mean | Log2 Fold Change | p-Value | ||
| Crude Corrected * | Crude Corrected * | ||||||||
| Abiotrophia | 4.06 | 1.27 | 0.02 | 0.42 | uncultured-02 | 10.1 | 1.86 | 0.02 | 0.45 |
| Lautropia | 20.0 | 0.94 | 0.01 | 0.38 | Abiotrophia | 5.88 | 1.55 | 0.02 | 0.45 |
| Aggregatibacter | 38.9 | 0.90 | 0.03 | 0.56 | Gemella | 128 | 1.20 | 0.002 | 0.30 |
| Veillonella | 3133 | −0.64 | 0.003 | 0.22 | Granulicatella | 59.1 | 0.89 | 0.01 | 0.45 |
| Actinomyces | 48.4 | −0.71 | 0.008 | 0.24 | Veillonella | 2819 | −0.51 | 0.04 | 0.53 |
| Prevotella 7 | 785 | −0.72 | 0.007 | 0.24 | Actinomyces | 47.7 | −0.75 | 0.03 | 0.45 |
| Prevotella 6 | 39.5 | −0.95 | 0.01 | 0.30 | Candidatus Saccharimonas | 35.3 | −0.90 | 0.02 | 0.45 |
| Megasphaera | 88.0 | −1.28 | 0.001 | 0.21 | Megasphaera | 70.8 | −1.19 | 0.02 | 0.45 |
| Butyrivibrio 2 | 6.32 | −1.55 | 0.005 | 0.24 | Butyrivibrio 2 | 5.52 | −1.58 | 0.02 | 0.45 |
| Candidatus Ancillula | 1.38 | −1.88 | 0.02 | 0.42 | Candidatus Ancillula | 1.45 | −2.37 | 0.04 | 0.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Räisänen, L.; Agrawal, N.; Mathew, B.; Kääriäinen, S.; Kolho, K.-L.; Viljakainen, H. Pre-Diagnostic Saliva Microbiota of School-Aged Children Who Developed Type 1 Diabetes or Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2023, 24, 8279. https://doi.org/10.3390/ijms24098279
Räisänen L, Agrawal N, Mathew B, Kääriäinen S, Kolho K-L, Viljakainen H. Pre-Diagnostic Saliva Microbiota of School-Aged Children Who Developed Type 1 Diabetes or Inflammatory Bowel Diseases. International Journal of Molecular Sciences. 2023; 24(9):8279. https://doi.org/10.3390/ijms24098279
Chicago/Turabian StyleRäisänen, Laura, Nitin Agrawal, Binu Mathew, Sohvi Kääriäinen, Kaija-Leena Kolho, and Heli Viljakainen. 2023. "Pre-Diagnostic Saliva Microbiota of School-Aged Children Who Developed Type 1 Diabetes or Inflammatory Bowel Diseases" International Journal of Molecular Sciences 24, no. 9: 8279. https://doi.org/10.3390/ijms24098279
APA StyleRäisänen, L., Agrawal, N., Mathew, B., Kääriäinen, S., Kolho, K. -L., & Viljakainen, H. (2023). Pre-Diagnostic Saliva Microbiota of School-Aged Children Who Developed Type 1 Diabetes or Inflammatory Bowel Diseases. International Journal of Molecular Sciences, 24(9), 8279. https://doi.org/10.3390/ijms24098279