
Changes in Photosynthetic Characteristics between Green-Leaf Poplar Linn. “2025” and Its Bud-Sporting Colored-Leaf Cultivars

 ,
,
Abstract
:1. Introduction
2. Results
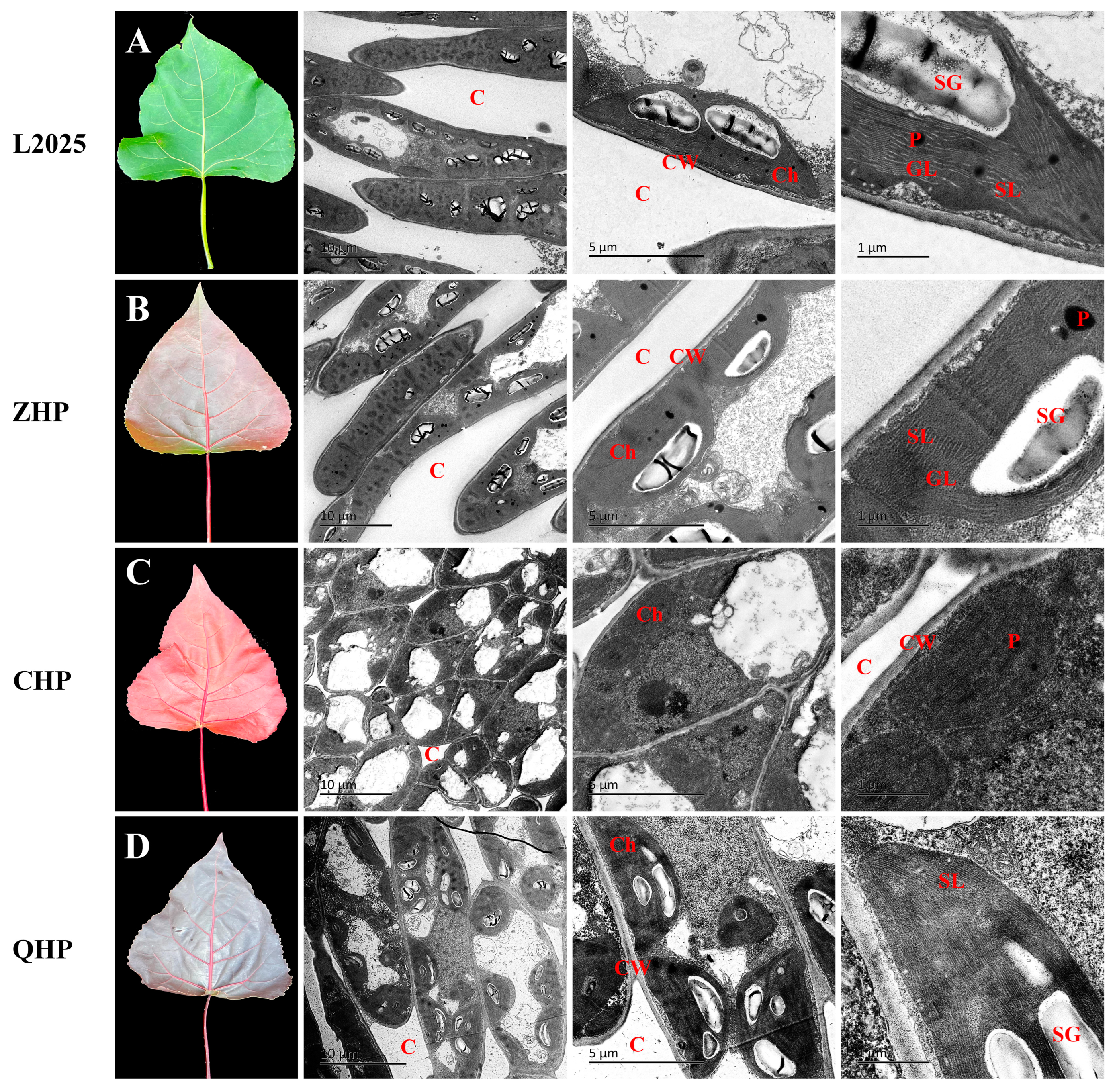
2.1. Leaf Morphology and Chloroplast Ultrastructure
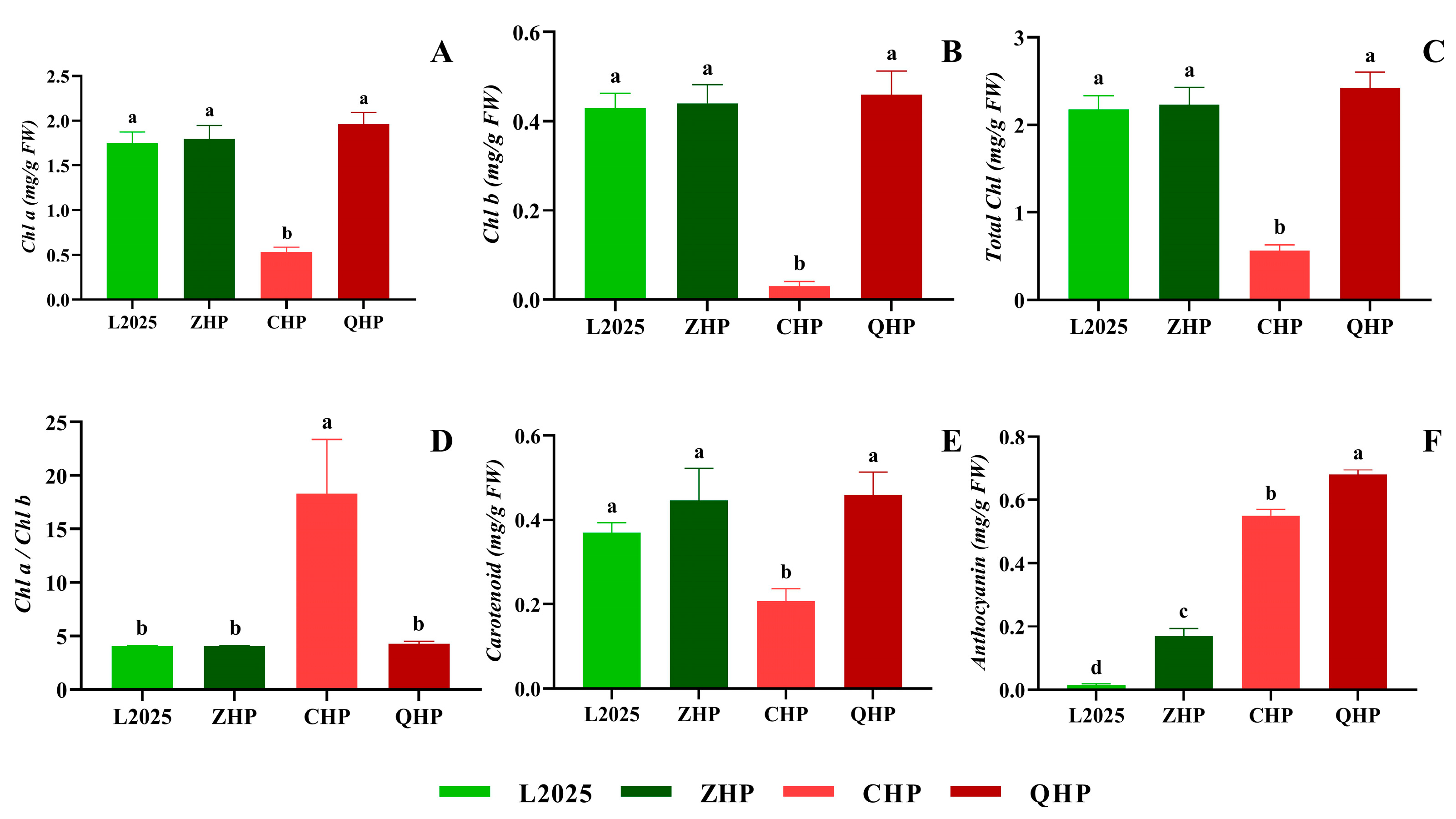
2.2. Contents of Photosynthetic Pigment and Anthocyanin in Different Colored-Leaf Poplars
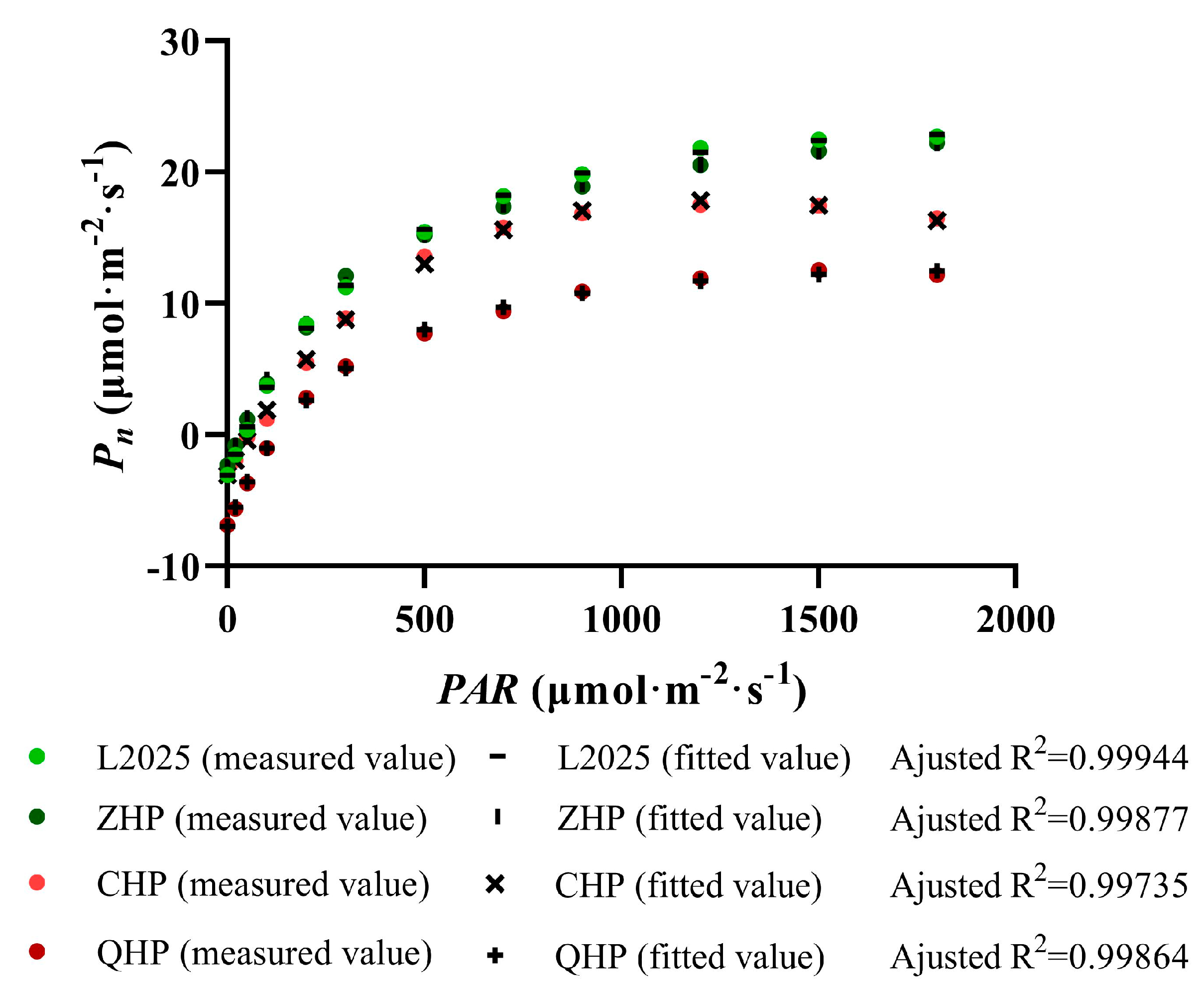
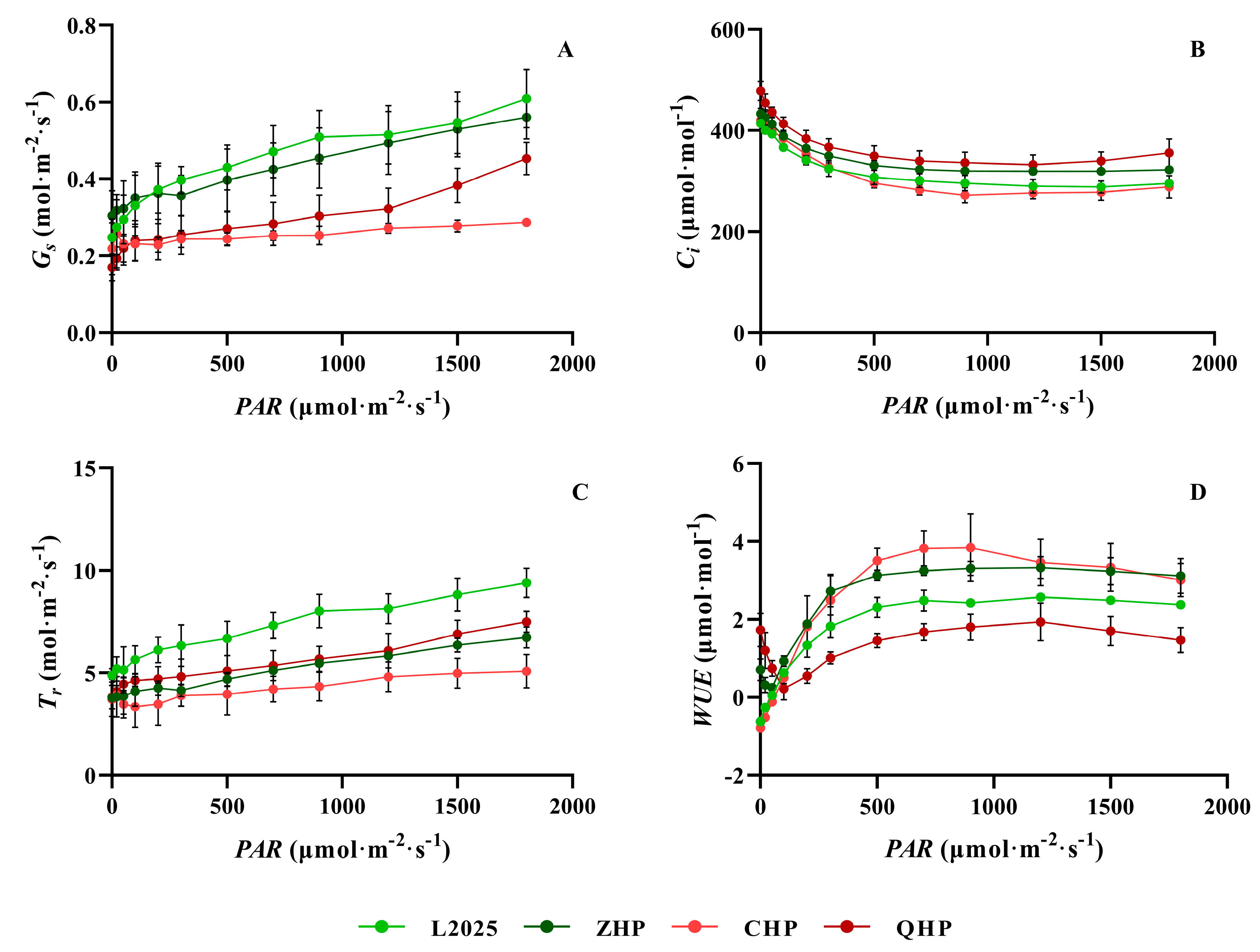
2.3. Measurements of Light Response Curve in Different Colored-Leaf Poplars
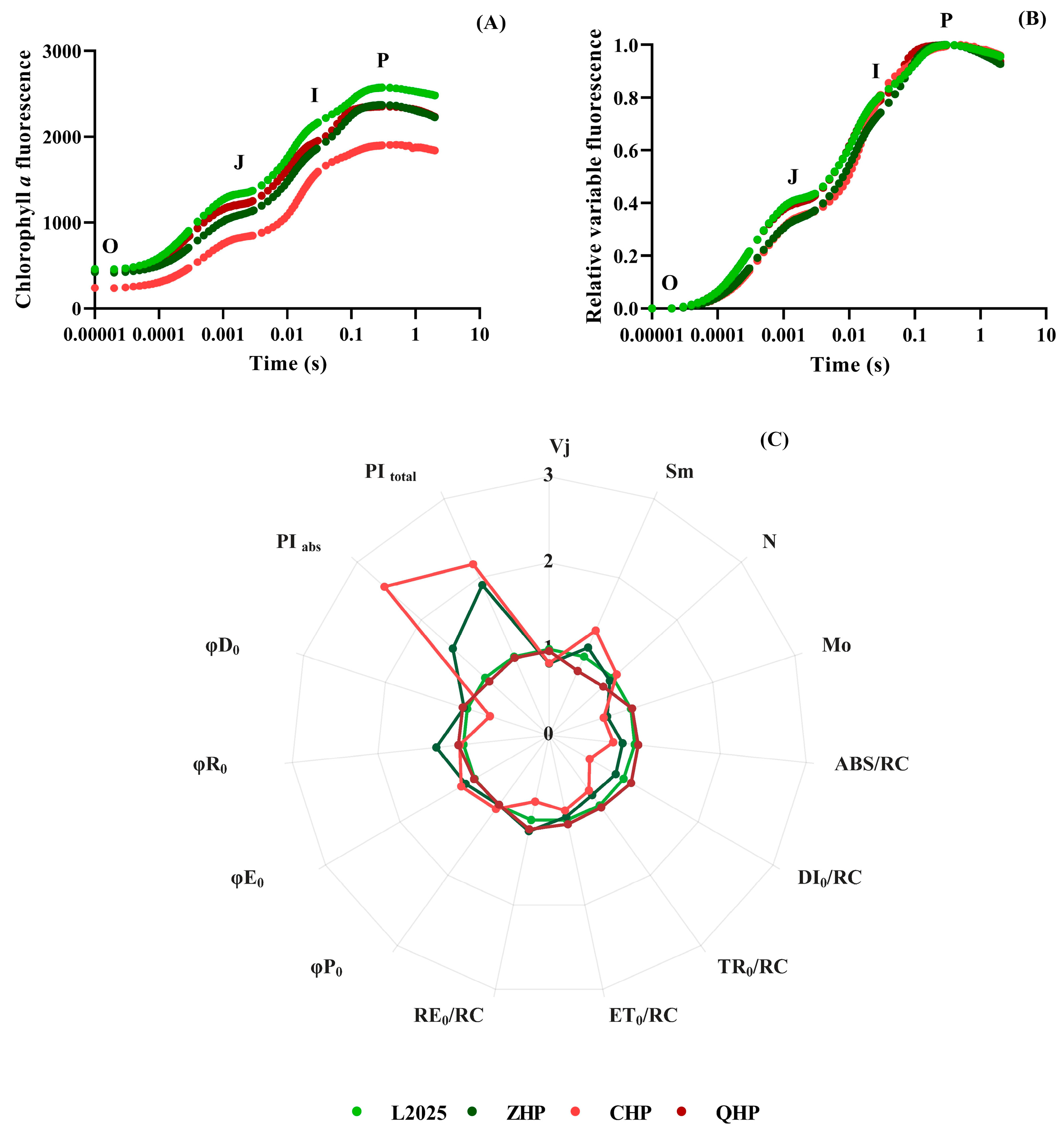
2.4. Fast Chlorophyll Fluorescence Induction Kinetics in Different Colored-Leaf Poplars
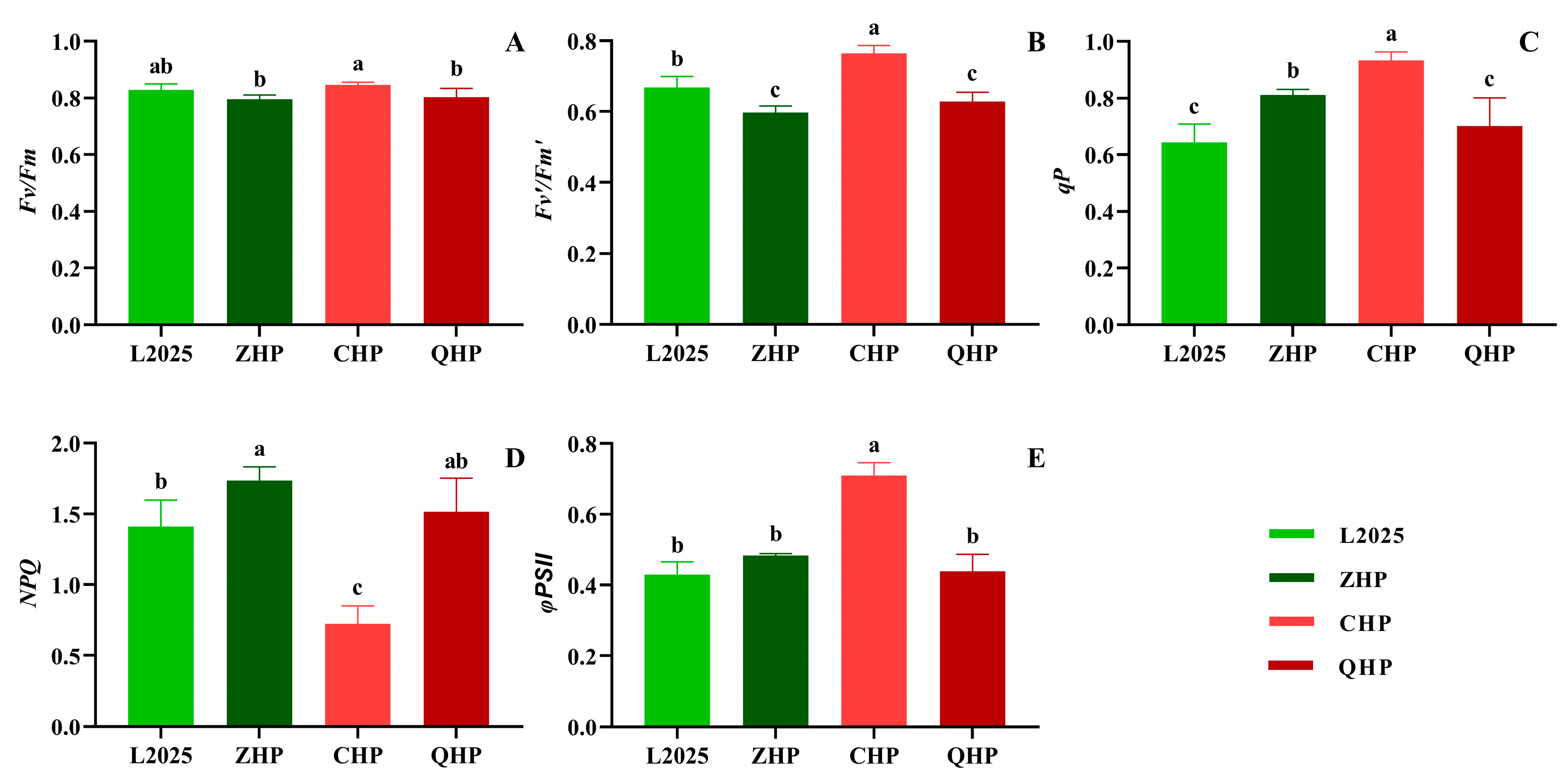
2.5. Chlorophyll Fluorescence Parameters in Different Colored-Leaf Poplars
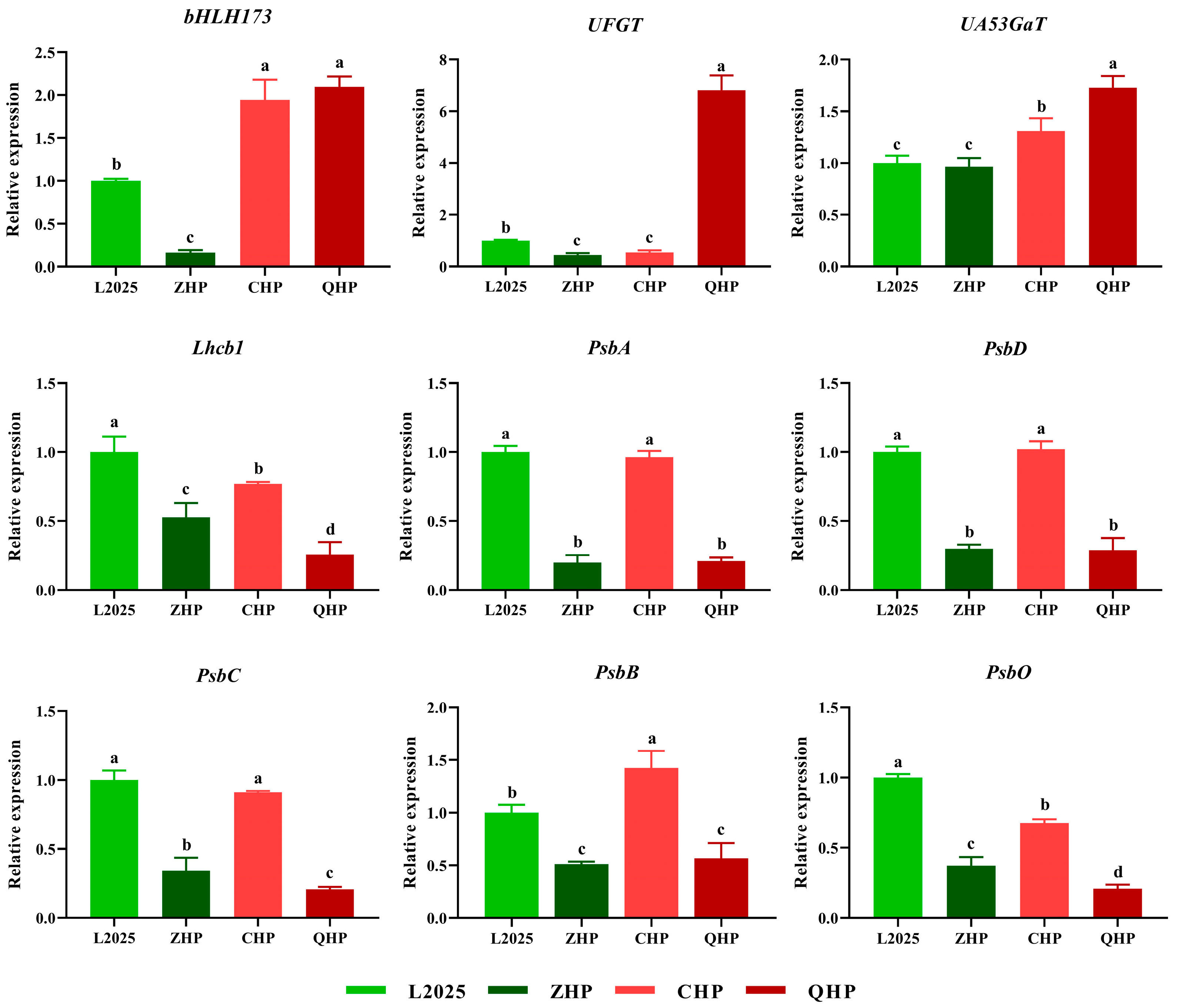
2.6. Expression Levels of Different Genes in Different Colored-Leaf Poplars
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Observation of Chloroplast Ultrastructure
4.3. Measurement of Chlorophyll and Anthocyanin Contents
4.4. Measurements of Photosynthetic-Light Response Curves
4.5. Measurements of Chlorophyll Fluorescence
4.6. Measurements of qRT-PCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, X.-J.; Peng, X.-Q.; Shu, X.-C.; Li, Y.-H.; Wang, Z.; Zhuang, W.-B. Genome-wide identification and characterization of PdbHLH transcription factors related to anthocyanin biosynthesis in colored-leaf poplar (Populus deltoids). BMC Genom. 2022, 23, 244. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, L.; Cheng, G.; Shu, X.; Wang, N.; Zhang, F.; Zhuang, W.; Wang, Z. Physiological and molecular analysis reveals the differences of photosynthesis between colored and green leaf poplars. Int. J. Mol. Sci. 2021, 22, 8982. [Google Scholar] [CrossRef]
- Chen, M.; Chang, C.; Li, H.; Huang, L.; Zhou, Z.; Zhu, J.; Liu, D. Metabolome analysis reveals flavonoid changes during the leaf color transition in Populus× euramericana ‘Zhonghuahongye’. Front. Plant Sci. 2023, 14, 1162893. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wan, X.; Zheng, Y.; Sun, L.; Chen, Q.; Guo, Y.; Zhu, X.; Liu, M. Physiological and related anthocyanin biosynthesis genes responses induced by cadmium stress in a new colored-leaf plant “Quanhong Poplar”. Agrofor. Syst. 2014, 88, 343–355. [Google Scholar] [CrossRef]
- Zhuang, W.; Wang, H.; Liu, T.; Wang, T.; Zhang, F.; Shu, X.; Zhai, H.; Wang, Z. Integrated physiological and genomic analysis reveals structural variations and expression patterns of candidate genes for colored-and green-leaf poplar. Sci. Rep. 2019, 9, 11150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhao, J.; Wan, X.; Luo, X.; Li, W.; Sun, L.; Chen, Q. From green to red: Large-scale transcriptome comparison of a bud sport in poplar (Populus deltoides). Acta Physiol. Plant. 2016, 38, 244. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, X.; Cui, J.; Ogweno, J.; Shi, K.; Zhou, Y.; Yu, J. Characteristics of gas exchange and chlorophyll fluorescence in red and green leaves of Begonia semperflorens. Biol. Plant. 2011, 55, 361–364. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Bornman, J.; Liakoura, V. Different leaf surface characteristics of three grape cultivars affect leaf optical properties as measured with fibre optics: Possible implication in stress tolerance. Funct. Plant Biol. 1999, 26, 47–53. [Google Scholar] [CrossRef]
- Liakopoulos, G.; Nikolopoulos, D.; Klouvatou, A.; Vekkos, K.-A.; Manetas, Y.; Karabourniotis, G. The photoprotective role of epidermal anthocyanins and surface pubescence in young leaves of grapevine (Vitis vinifera). Ann. Bot. 2006, 98, 257–265. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Hanke, G.; Mulo, P. Plant type ferredoxins and ferredoxin-dependent metabolism. Plant Cell Environ. 2013, 36, 1071–1084. [Google Scholar] [CrossRef] [PubMed]
- Pomar, F.; Ros Barceló, A. Are red leaves photosynthetically active? Biol. Plant. 2007, 51, 799–800. [Google Scholar] [CrossRef]
- Zhou, X.-S.; Shen, S.-Q.; Wu, D.-X.; Sun, J.-W.; Shu, Q.-Y. Introduction of a xantha mutation for testing and increasing varietal purity in hybrid rice. Field Crops Res. 2006, 96, 71–79. [Google Scholar] [CrossRef]
- Wen, W.; Alseekh, S.; Fernie, A.R. Conservation and diversification of flavonoid metabolism in the plant kingdom. Curr. Opin. Plant Biol. 2020, 55, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Routaboul, J.-M.; Kerhoas, L.; Debeaujon, I.; Pourcel, L.; Caboche, M.; Einhorn, J.; Lepiniec, L. Flavonoid diversity and biosynthesis in seed of Arabidopsis thaliana. Planta 2006, 224, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Outchkourov, N.S.; Karlova, R.; Hölscher, M.; Schrama, X.; Blilou, I.; Jongedijk, E.; Simon, C.D.; van Dijk, A.D.; Bosch, D.; Hall, R.D. Transcription factor-mediated control of anthocyanin biosynthesis in vegetative tissues. Plant Physiol. 2018, 176, 1862–1878. [Google Scholar] [CrossRef]
- Kós, P.B.; Deák, Z.; Cheregi, O.; Vass, I. Differential regulation of psbA and psbD gene expression, and the role of the different D1 protein copies in the cyanobacterium Thermosynechococcus elongatus BP-1. Biochim. Biophys. Acta (BBA)-Bioenerg. 2008, 1777, 74–83. [Google Scholar] [CrossRef]
- Jain, A.; Cao, A.; Karthikeyan, A.; Baldwin, J.; Raghothama, K. Phosphate deficiency suppresses expression of light-regulated psbO and psbP genes encoding extrinsic proteins of oxygen-evolving complex of PSII. Curr. Sci. 2005, 89, 1592–1596. [Google Scholar]
- Murray, J.W.; Duncan, J.; Barber, J. CP43-like chlorophyll binding proteins: Structural and evolutionary implications. Trends Plant Sci. 2006, 11, 152–158. [Google Scholar] [CrossRef]
- Pan, X.; Cao, P.; Su, X.; Liu, Z.; Li, M. Structural analysis and comparison of light-harvesting complexes I and II. Biochim. Biophys. Acta (BBA)-Bioenerg. 2020, 1861, 148038. [Google Scholar] [CrossRef]
- Ye, Z.P.; Suggett, D.J.; Robakowski, P.; Kang, H.J. A mechanistic model for the photosynthesis–light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, X.; Hao, K.; Yang, Q.; Yang, X.; Zhang, W.; Cong, Y. Light-response curve of photosynthesis and model fitting in leaves of Mangifera indica under different soil water conditions. Photosynthetica 2019, 57, 796–803. [Google Scholar] [CrossRef]
- Vialet-Chabrand, S.R.; Matthews, J.S.; McAusland, L.; Blatt, M.R.; Griffiths, H.; Lawson, T. Temporal dynamics of stomatal behavior: Modeling and implications for photosynthesis and water use. Plant Physiol. 2017, 174, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Qi, H.; Zhang, Z.; Song, Z.; Kou, T.; Zhang, W.; Yu, J. Response of photosynthesis and chlorophyll fluorescence to drought stress in two maize cultivars. Afr. J. Agric. Res. 2012, 7, 4751–4760. [Google Scholar]
- Liu, X.; Fan, Y.; Long, J.; Wei, R.; Kjelgren, R.; Gong, C.; Zhao, J. Effects of soil water and nitrogen availability on photosynthesis and water use efficiency of Robinia pseudoacacia seedlings. J. Environ. Sci. 2013, 25, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, K.; Sun, Y.; Yan, Q. Response of Pinus koraiensis seedling growth to different light conditions based on the assessment of photosynthesis in current and one-year-old needles. J. For. Res. 2014, 25, 53–62. [Google Scholar] [CrossRef]
- Gao, M.; Zhao, Y.; Zong, Y.; Wang, W. Photosynthetic Traits and Ecological Adaptability of Poplar 2025 and Its 3 Bud Sporting Color-leafed Cultivars. J. Northwest For. Univ. 2020, 35, 102–107. [Google Scholar]
- Zhu, X.; Yang, J.; Wen, D.; Han, X.; Ru, G. Photoprotective effects of anthocyanins in leaves of color-leaved poplar. Southwest China J. Agric. Sci. 2020, 33, 958–965. [Google Scholar]
- Zeng, X.Q.; Chow, W.S.; Su, L.J.; Peng, X.X.; Peng, C.L. Protective effect of supplemental anthocyanins on Arabidopsis leaves under high light. Physiol. Plant. 2010, 138, 215–225. [Google Scholar] [CrossRef]
- Gould, K.S.; Jay-Allemand, C.; Logan, B.A.; Baissac, Y.; Bidel, L.P. When are foliar anthocyanins useful to plants? Re-evaluation of the photoprotection hypothesis using Arabidopsis thaliana mutants that differ in anthocyanin accumulation. Environ. Exp. Bot. 2018, 154, 11–22. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, X.; Cui, J.; Zhang, F.; Wan, X.; Liu, Q.; Zhong, Y.; Lin, T. Physiological and transcriptomic analysis of yellow leaf coloration in Populus deltoides Marsh. PLoS ONE 2019, 14, e0216879. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Lai, J.; He, T.; Rong, J.; Tarin, M.W.K.; Zheng, Y. Differences in photosynthesis of variegated temple bamboo leaves with various levels of variegation are related to chlorophyll biosynthesis and chloroplast development. J. Am. Soc. Hortic. Sci. 2018, 143, 144–153. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Springer: Berlin/Heidelberg, Germany, 2004; pp. 321–362. [Google Scholar]
- Alexandrina Stirbet, G. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Wang, T.; Luo, S.; Ma, Y.; Li, L.; Xie, Y.; Zhang, W. Chlorophyll a Fluorescence Transient and 2-Dimensional Electrophoresis Analyses Reveal Response Characteristics of Photosynthesis to Heat Stress in Malus.‘Prairifire’. Plants 2020, 9, 1040. [Google Scholar] [CrossRef]
- Sipka, G.; Magyar, M.; Mezzetti, A.; Akhtar, P.; Zhu, Q.; Xiao, Y.; Han, G.; Santabarbara, S.; Shen, J.-R.; Lambrev, P.H. Light-adapted charge-separated state of photosystem II: Structural and functional dynamics of the closed reaction center. Plant Cell 2021, 33, 1286–1302. [Google Scholar] [CrossRef]
- Garab, G.; Magyar, M.; Sipka, G.; Lambrev, P.H. New foundations for the physical mechanism of variable chlorophyll a fluorescence. Quantum efficiency versus the light-adapted state of photosystem II. J. Exp. Bot. 2023, 74, 5458–5471. [Google Scholar] [CrossRef]
- Gao, J.; Wang, H.; Yuan, Q.; Feng, Y. Structure and function of the photosystem supercomplexes. Front. Plant Sci. 2018, 9, 357. [Google Scholar] [CrossRef]
- Nixon, P.J.; Michoux, F.; Yu, J.; Boehm, M.; Komenda, J. Recent advances in understanding the assembly and repair of photosystem II. Ann. Bot. 2010, 106, 1–16. [Google Scholar] [CrossRef]
- Pospíšil, P. Molecular mechanisms of production and scavenging of reactive oxygen species by photosystem II. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 218–231. [Google Scholar] [CrossRef]
- Jansen, M.A.; Gaba, V.; Greenberg, B.M.; Mattoo, A.K.; Edelman, M. Low threshold levels of ultraviolet-B in a background of photosynthetically active radiation trigger rapid degradation of the D2 protein of photosystem-II. Plant J. 1996, 9, 693–699. [Google Scholar] [CrossRef]
- Bi, A.; Fan, J.; Hu, Z.; Wang, G.; Amombo, E.; Fu, J.; Hu, T. Differential acclimation of enzymatic antioxidant metabolism and photosystem II photochemistry in tall fescue under drought and heat and the combined stresses. Front. Plant Sci. 2016, 7, 453. [Google Scholar] [CrossRef]
- Wang, T.; Li, L.; Qin, Y.; Lu, B.; Xu, D.; Zhuang, W.; Shu, X.; Zhang, F.; Wang, N.; Wang, Z. Effects of seasonal changes on chlorophyll fluorescence and physiological characteristics in the two Taxus species. Plants 2023, 12, 2636. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AQY | Amax (μmol∙m−2∙s−1) | Rd (μmol∙m−2∙s−1) | LSP (μmol∙m−2∙s−1) | LCP (μmol∙m−2∙s−1) | |
|---|---|---|---|---|---|
| L2025 | 0.062 ± 0.004 a | 22.621 ± 3.120 a | 3.124 ± 0.046 b | 2408.087 ± 154.371 a | 40.915 ± 0.436 c |
| ZHP | 0.060 ± 0.002 a | 22.535 ± 1.816 a | 2.640 ± 0.234 c | 2739.033 ± 318.509 a | 34.501 ± 3.677 c |
| CHP | 0.045 ± 0.006 b | 16.425 ± 1.275 b | 3.149 ± 0.274 b | 1220.310 ± 171.868 c | 57.464 ± 3.004 b |
| QHP | 0.048 ± 0.008 b | 11.216 ± 1.210 c | 6.594 ± 0.392 a | 1858.683 ± 265.537 b | 126.374 ± 5.138 a |
| Fluorescence Parameter | Description |
|---|---|
| Derived parameters | |
| Vj | relative variable fluorescence at the J-step |
| Sm | normalized total complementary area above the OJIP transient or total electron carriers per RC |
| N | the times QA was reduced to QA− in the time span from t0 to tFmax |
| Mo | approximated initial slope of the Chl fluorescence transient V = f(t) |
| Specifc fuxes or activities per RC | |
| ABS/RC | average absorbed photon flux per PSII RC |
| TR0/RC | specific energy fluxes per RC for trapping at t = 0 |
| ET0/RC | specific energy fluxes per RC for electron transport at t = 0 |
| DI0/RC | specific energy fluxes per RC for dissipation at t = 0 |
| RE0/RC | specific electron fluxes per RC for reduction of PSI acceptors at t = 0 |
| Yields or fux ratios | |
| φP0 | maximum quantum yield of primary PSII photochemistry |
| φE0 | quantum yield for electron transport from QA− to plastoquinone |
| φD0 | quantum yield (t = 0) of energy dissipation |
| φR0 | quantum yield for reduction in the end electron acceptors at the PSI acceptor side |
| Performance index | |
| PIabs | performance index for energy conservation from photons absorbed by PSII antenna to the reduction of QB |
| PItotal | performance index for energy conservation from photons absorbed by PSII antenna to the reduction of PSI acceptors |
| Gene Name | Accession | Description | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|---|---|
| PsbA | Potri.013G143200 | photosystem II reaction center protein A (D1) | GGGTCGCTTCTGTAATTGG | AGTTGCGGTCAATAAAGTAGG |
| PsbD | Potri.008G208600 | photosystem II reaction center protein D (D2) | TGCAATCGCATTCTCTGG | AACTAGGCGCAAAGAACC |
| PsbC | Potri.010G032700 | photosystem II reaction center protein C (CP43) | GGAAGTCATAGACACCTTTCC | TCGGGTCCTAGAAGTGC |
| PsbB | Potri.011G113900 | photosystem II reaction center protein B (CP47) | GATAAAGAAGGGCGTGAGC | ATTCCGTCTCCGTCTACC |
| PsbO | Potri.005G130400 | photosystem II oxygen-evolving enhancer protein 1 | TGAAGGAGTTCCGAAGAGG | TCAATGGTTGGGCATTGG |
| Lhcb1 | Potri.005G239200 | light-harvesting complex II chlorophyll a/b binding protein 1 (lhcb1) | ATCACTGACCCGATCTACC | CAGTCTACCATTCTTGAGTTCC |
| UFGT | Potri.009G133300 | Flavonoid 3-O-galactosyl transferase family protein | TGGCGTATATCAGCTTTGG | CCTAAGAGACCAAAGGAATGG |
| UA53GaT | Potri.015G027700 | Anthocyanidin 5,3-O-glucosyltransferase | AAACGGCTATTGGGTTGG | TCATTTGGAGTGCTTGACC |
| bHLH173 | bHLH transcription factor | CCTCGAATGTGAGGAAACC | AAACTGAACTTCCTTCCTAGC | |
| Actin2 | Actin | GCCATCTCTCATCGGAATGGAA | AGGGCAGTGATTTCCTTGCTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Xu, D.; Zhang, F.; Yan, T.; Li, Y.; Wang, Z.; Xie, Y.; Zhuang, W. Changes in Photosynthetic Characteristics between Green-Leaf Poplar Linn. “2025” and Its Bud-Sporting Colored-Leaf Cultivars. Int. J. Mol. Sci. 2024, 25, 1225. https://doi.org/10.3390/ijms25021225
Wang T, Xu D, Zhang F, Yan T, Li Y, Wang Z, Xie Y, Zhuang W. Changes in Photosynthetic Characteristics between Green-Leaf Poplar Linn. “2025” and Its Bud-Sporting Colored-Leaf Cultivars. International Journal of Molecular Sciences. 2024; 25(2):1225. https://doi.org/10.3390/ijms25021225
Chicago/Turabian StyleWang, Tao, Donghuan Xu, Fan Zhang, Tengyue Yan, Yuhang Li, Zhong Wang, Yinfeng Xie, and Weibing Zhuang. 2024. "Changes in Photosynthetic Characteristics between Green-Leaf Poplar Linn. “2025” and Its Bud-Sporting Colored-Leaf Cultivars" International Journal of Molecular Sciences 25, no. 2: 1225. https://doi.org/10.3390/ijms25021225
APA StyleWang, T., Xu, D., Zhang, F., Yan, T., Li, Y., Wang, Z., Xie, Y., & Zhuang, W. (2024). Changes in Photosynthetic Characteristics between Green-Leaf Poplar Linn. “2025” and Its Bud-Sporting Colored-Leaf Cultivars. International Journal of Molecular Sciences, 25(2), 1225. https://doi.org/10.3390/ijms25021225