

The Shoot Apical Meristem: An Evolutionary Molding of Higher Plants



Abstract
:1. Introduction
- In the first section, we present essential concepts to understand plant meristems. We introduce the concept of stem cells and compare the stem cell niche of RAM and SAM. We emphasize the importance of plant life cycles, as this concept gives rise to SAM evolution theories.
- Then, the meristem shape is analyzed throughout phylogeny. This section offers a morphological description of SAM across plant evolution. Given the extensive literature on SAM morphology, it provides perspectives on the differences in SAM between clades.
- We provide a comprehensive review of the regulatory and maintenance mechanisms in the SAM. This section focuses on the regulatory loops described for angiosperms and the conserved elements across clades.
- Taking advantage of single-cell transcriptomics to understand SAM, this section delves into the research of single-cell transcriptomics and single-nucleus transcriptomics on SAM. We describe key studies and discuss their findings.
2. Evolutionary Origin of the Meristem
2.1. The Concept of Stem Cells
2.2. SAM and RAM

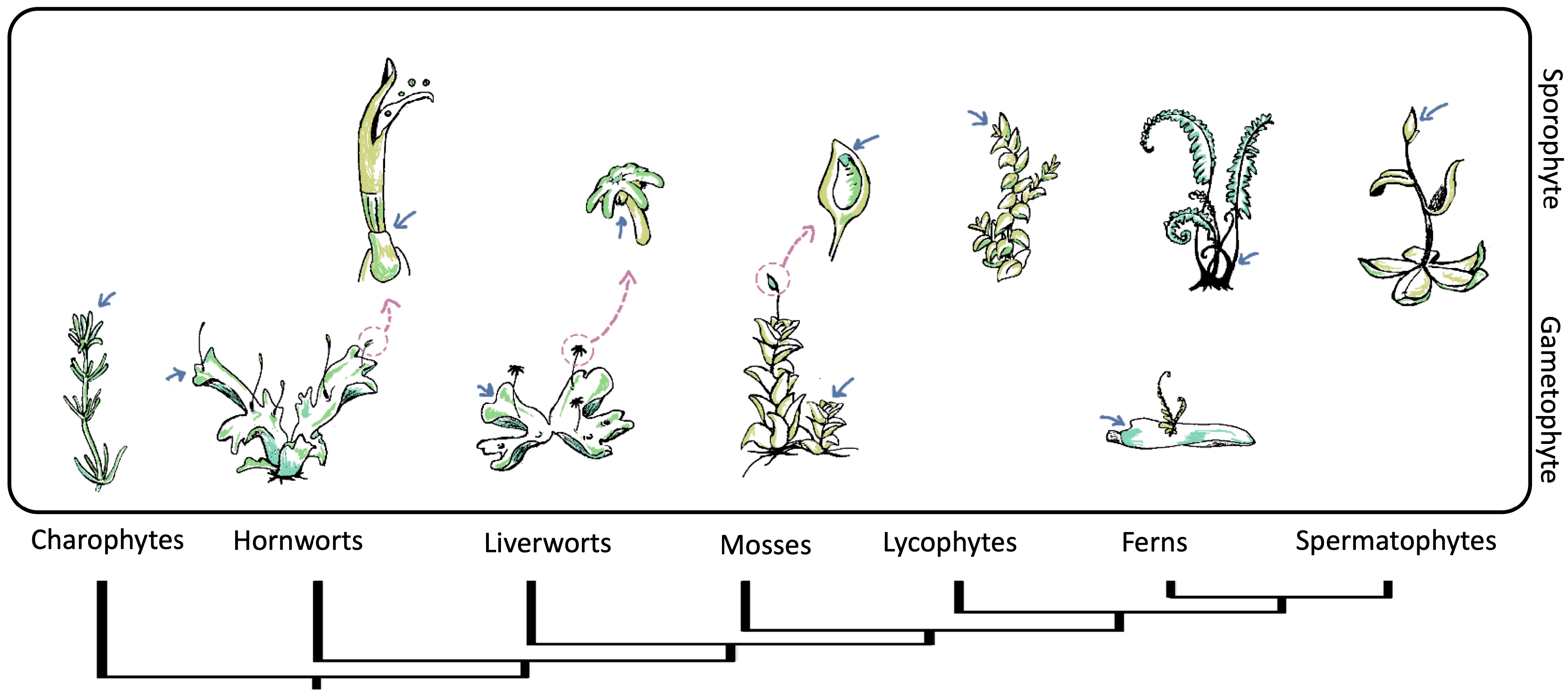
2.3. Apical Meristem of Gametophyte and Sporophyte
3. Meristem Shape throughout Phylogeny
3.1. Algae
3.2. Bryophytes
3.3. Tracheophytes
4. Shoot Apical Meristem Regulation and Maintenance
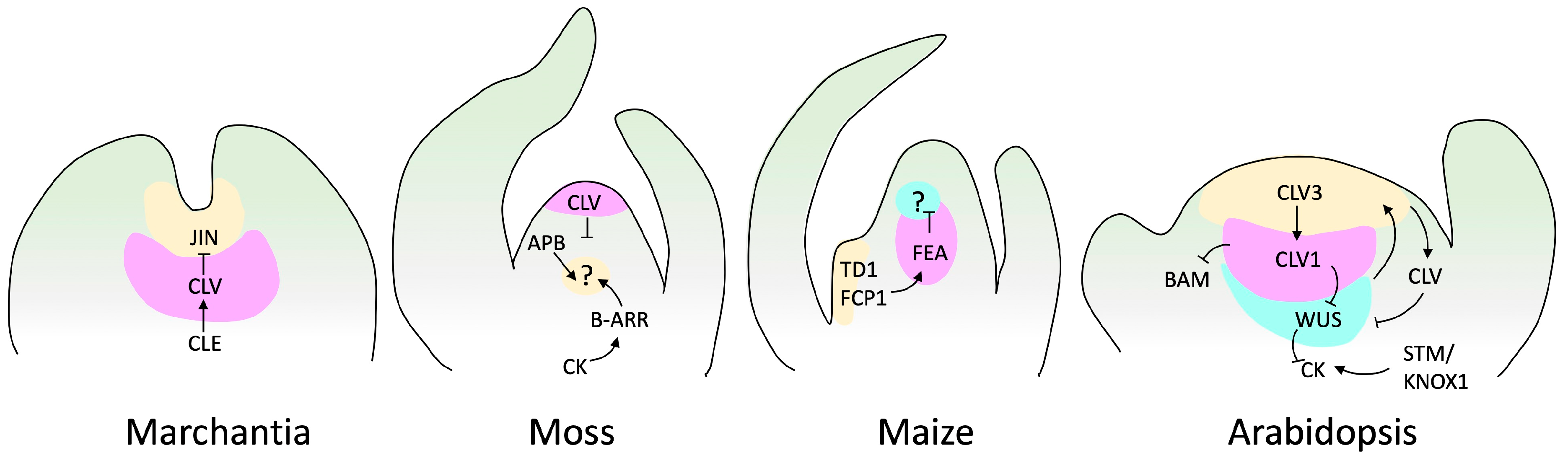
4.1. The Regulatory Model: Angiosperms

4.2. SAM TFs Conserved throughout Evolution
4.2.1. KNOX TFs
4.2.2. MADS TFs
4.2.3. AP2/ERF TFs
5. Taking Advantage of Single-Cell Transcriptomics to Understand SAM
6. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, J.A.H.; Jones, A.; Godin, C.; Traas, J. Systems Analysis of Shoot Apical Meristem Growth and Development: Integrating Hormonal and Mechanical Signaling. Plant Cell 2012, 24, 3907–3919. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Theodore, T. Growth and Development of Trees; Academic Press: New York, NY, USA, 1971; ISBN 9780124242012. [Google Scholar]
- Jacobs, W.P. The Development of the Gynophore of the Peanut Plant, Arachis hypogaea L. I. The Distribution of Mitoses, the Region of Greatest Elongation, and the Maintenance of Vascular Continuity in the Intercalary Meristem. Am. J. Bot. 1947, 34, 361–370. [Google Scholar] [CrossRef]
- Jill, H.C. Development and Genetics in the Evolution of Land Plant Body Plans. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20150490. [Google Scholar]
- Banks, J.A. The Evolution of the Shoot Apical Meristem from a Gene Expression Perspective. New Phytol. 2015, 207, 486–487. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, V.B. The Problem of Stem Cells in Plants. Russ. J. Dev. Biol. 2023, 34, 205–212. [Google Scholar] [CrossRef]
- Barlow, P.W. Stem Cells and Tissue Homeostasis; Cambridge University Press: Cambridge, UK, 1978; ISBN 9780521217996. [Google Scholar]
- Clark, S.E. Organ Formation at the Vegetative Shoot Meristem. Plant Cell 1997, 9, 1067–1076. [Google Scholar] [CrossRef]
- Dubrovsky, J.G.; Ivanov, V.B. The Quiescent Centre of the Root Apical Meristem: Conceptual Developments from Clowes to Modern Times. J. Exp. Bot. 2021, 72, 6687–6707. [Google Scholar] [CrossRef] [PubMed]
- Lev-Yadun, S. Stem cells in plants are differentiated too. Curr. Top. Plant Biol. 2023, 4, 93–102. [Google Scholar]
- Fehér, A. Callus, Dedifferentiation, Totipotency, Somatic Embryogenesis: What These Terms Mean in the Era of Molecular Plant Biology? Front. Plant Sci. 2019, 10, 442509. [Google Scholar] [CrossRef]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.M.; Van Lammeren, A.A.M.; Miki, B.L.A.; et al. Ectopic Expression of BABY BOOM Triggers a Conversion from Vegetative to Embryonic Growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef]
- Xu, M.; Du, Q.; Tian, C.; Wang, Y.; Jiao, Y. Stochastic Gene Expression Drives Mesophyll Protoplast Regeneration. Sci. Adv. 2021, 7, 8466–8477. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.; Kim, H.K.; Bae, S.H.; Lee, H.; Lee, H.J.; Jung, Y.J.; Seo, P.J. Transcriptome Comparison between Pluripotent and Non-Pluripotent Calli Derived from Mature Rice Seeds. Sci. Rep. 2020, 10, 21257. [Google Scholar] [CrossRef] [PubMed]
- Stahl, Y.; Simon, R. Plant Primary Meristems: Shared Functions and Regulatory Mechanisms. Curr. Opin. Plant Biol. 2010, 13, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Seago, J.L.; Fernando, D.D. Anatomical Aspects of Angiosperm Root Evolution. Ann. Bot. 2013, 112, 223–238. [Google Scholar] [CrossRef]
- Haecker, A.; Groß-Hardt, R.; Geiges, B.; Sarkar, A.; Breuninger, H.; Herrmann, M.; Laux, T. Expression Dynamics of WOX Genes Mark Cell Fate Decisions during Early Embryonic Patterning in Arabidopsis Thaliana. Development 2004, 131, 657–668. [Google Scholar] [CrossRef]
- Bennett, T.; Scheres, B. Root Development-Two Meristems for the Price of One? Curr. Top. Dev. Biol. 2010, 91, 67–102. [Google Scholar] [PubMed]
- Sarkar, A.K.; Luijten, M.; Miyashima, S.; Lenhard, M.; Hashimoto, T.; Nakajima, K.; Scheres, B.; Heidstra, R.; Laux, T. Conserved Factors Regulate Signalling in Arabidopsis Thaliana Shoot and Root Stem Cell Organizers. Nature 2007, 446, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Skylar, A.; Wu, X. Regulation of Meristem Size by Cytokinin SignalingF. J. Integr. Plant Biol. 2011, 53, 446–454. [Google Scholar] [CrossRef]
- Greb, T.; Lohmann, J.U. Plant Stem Cells. Curr. Biol. 2016, 26, R816–R821. [Google Scholar] [CrossRef]
- Mähönen, A.P.; ten Tusscher, K.; Siligato, R.; Smetana, O.; Díaz-Triviño, S.; Salojärvi, J.; Wachsman, G.; Prasad, K.; Heidstra, R.; Scheres, B. PLETHORA Gradient Formation Mechanism Separates Auxin Responses. Nature 2014, 515, 125–129. [Google Scholar] [CrossRef]
- Dello Ioio, R.; Galinha, C.; Fletcher, A.G.; Grigg, S.P.; Molnar, A.; Willemsen, V.; Scheres, B.; Sabatini, S.; Baulcombe, D.; Maini, P.K.; et al. A PHABULOSA/Cytokinin Feedback Loop Controls Root Growth in Arabidopsis. Curr. Biol. 2012, 22, 1699–1704. [Google Scholar] [CrossRef] [PubMed]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The Yin-Yang of Hormones: Cytokinin and Auxin Interactions in Plant Development. Plant Cell 2015, 27, 44–63. [Google Scholar] [CrossRef]
- Friedman, W.E.; Moore, R.C.; Purugganan, M.D. The Evolution of Plant Development. Am. J. Bot. 2004, 91, 1726–1741. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, A.J.; Dolan, L. Stepwise and Independent Origins of Roots among Land Plants. Nature 2018, 561, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xu, L. Recruitment of IC-WOX Genes in Root Evolution. Trends Plant Sci. 2018, 23, 490–496. [Google Scholar] [CrossRef]
- Fujinami, R.; Yamada, T.; Imaichi, R. Root Apical Meristem Diversity and the Origin of Roots: Insights from Extant Lycophytes. J. Plant Res. 2020, 133, 291–296. [Google Scholar] [CrossRef]
- Huang, L.; Schiefelbein, J. Conserved Gene Expression Programs in Developing Roots from Diverse Plants. Plant Cell 2015, 27, 2119–2132. [Google Scholar] [CrossRef]
- Augstein, F.; Carlsbecker, A. Getting to the Roots: A Developmental Genetic View of Root Anatomy and Function from Arabidopsis to Lycophytes. Front. Plant Sci. 2018, 9, 411407. [Google Scholar] [CrossRef]
- Leebens-Mack, J.H.; Barker, M.S.; Carpenter, E.J.; Deyholos, M.K.; Gitzendanner, M.A.; Graham, S.W.; Grosse, I.; Li, Z.; Melkonian, M.; Mirarab, S.; et al. One Thousand Plant Transcriptomes and the Phylogenomics of Green Plants. Nature 2019, 574, 679–685. [Google Scholar]
- Ferrari, C.; Shivhare, D.; Hansen, B.O.; Pasha, A.; Esteban, E.; Provart, N.J.; Kragler, F.; Fernie, A.; Tohge, T.; Mutwil, M. Expression Atlas of Selaginella Moellendorffii Provides Insights into the Evolution of Vasculature, Secondary Metabolism, and Roots. Plant Cell 2020, 32, 853–870. [Google Scholar] [CrossRef]
- Su, Y.H.; Liu, Y.B.; Zhang, X.S. Auxin-Cytokinin Interaction Regulates Meristem Development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef]
- Luo, L.; Zeng, J.; Wu, H.; Tian, Z.; Zhao, Z. A Molecular Framework for Auxin-Controlled Homeostasis of Shoot Stem Cells in Arabidopsis. Mol. Plant 2018, 11, 899–913. [Google Scholar] [CrossRef]
- Allen, C.E. The American Society of Naturalists Haploid and Diploid Generations. Am. Nat. 1937, 71, 734. [Google Scholar] [CrossRef]
- Albert, V.A. Shoot Apical Meristems and Floral Patterning: An Evolutionary Perspective. Trends Plant Sci. 1999, 4, 84–86. [Google Scholar] [CrossRef]
- Ligrone, R.; Duckett, J.G.; Renzaglia, K.S. The Origin of the Sporophyte Shoot in Land Plants: A Bryological Perspective. Ann. Bot. 2012, 110, 935. [Google Scholar] [CrossRef] [PubMed]
- Hata, Y.; Kyozuka, J. Fundamental Mechanisms of the Stem Cell Regulation in Land Plants: Lesson from Shoot Apical Cells in Bryophytes. Plant Mol. Biol. 2021, 107, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yan, A.; McAdam, S.A.M.; Banks, J.A.; Zhang, S.; Zhou, Y. Timing of Meristem Initiation and Maintenance Determines the Morphology of Fern Gametophytes. J. Exp. Bot. 2021, 72, 6990–7001. [Google Scholar] [CrossRef]
- Paolillo, D.J.; Bazzaz, F.A. Photosynthesis in Sporophytes of Polytrichum and Funaria. In The Bryologist; American Bryological and Lichenological Society: West Portsmouth, OH, USA, 1968; Volume 71, pp. 335–343. [Google Scholar]
- Haig, D. Homologous Versus Antithetic Alternation of Generations and the Origin of Sporophytes. Bot. Rev. 2008, 74, 395–418. [Google Scholar] [CrossRef]
- Niklas, K.J.; Kutschera, U. The Evolution of the Land Plant Life Cycle. New Phytol. 2010, 185, 27–41. [Google Scholar] [CrossRef]
- Bennici, A. Origin and Early Evolution of Land Plants Problems and Considerations. Commun. Integr. Biol. 2008, 1, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Hemsley, A.R. The origin of the land plant sporophyte: An interpolational scenario. Biol. Rev. 1994, 69, 263–273. [Google Scholar] [CrossRef]
- Kenrick, P. Alternation of generations in land plants: New phylogenetic and palaeobotanical evidence. Biol. Rev. 1994, 69, 293–330. [Google Scholar] [CrossRef]
- Harrison, C.J. Auxin Transport in the Evolution of Branching Forms. New Phytol. 2017, 215, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.H.; Scanlon, M.J. Transcriptomic Evidence for the Evolution of Shoot Meristem Function in Sporophyte-Dominant Land Plants through Concerted Selection of Ancestral Gametophytic and Sporophytic Genetic Programs. Mol. Biol. Evol. 2015, 32, 355–367. [Google Scholar] [CrossRef]
- Youngstrom, C.E.; Geadelmann, L.F.; Irish, E.E.; Cheng, C.L. A Fern WUSCHEL-RELATED HOMEOBOX Gene Functions in Both Gametophyte and Sporophyte Generations. BMC Plant Biol 2019, 19, 416. [Google Scholar] [CrossRef]
- Popham, R.A. Principal types of vegetative shoot apex organization in vascular plants. Ohio J. Sci. 1951, 51, 249–270. [Google Scholar]
- Renzaglia, K.S. A Comparative Developmental Investigation of the Gametophyte Generation in the Metzgeriales (Hepatophyta). Ph.D. Thesis, Southern Illinois University, Edwardsville, IL, USA, 1982. [Google Scholar]
- Korn, R.W. Apical Cells as Meristems; Kluwer Academic Publishers: Amsterdam, The Netherlands, 1993; Volume 41. [Google Scholar]
- Nishihama, R.; Naramoto, S. Apical Stem Cells Sustaining Prosperous Evolution of Land Plants. J. Plant Res. 2020, 133, 279–282. [Google Scholar] [CrossRef]
- Moody, L.A. Unravelling 3D Growth in the Moss Physcomitrium Patens. Essays Biochem. 2022, 66, 769–779. [Google Scholar]
- Shaw, J.; Renzaglia, K. Phylogeny and Diversification of Bryophytes. Am. J. Bot. 2004, 91, 1557–1581. [Google Scholar] [CrossRef]
- McVeigh, I. Vegetative Reproduction of the Fern Sporophyte. Bot. Rev. 1937, 3, 457–497. [Google Scholar] [CrossRef]
- Philipson, W.R. The Significance of Apical Meristems in the Phylogeny of Land Plants. Plant Syst. Evol. 1990, 173, 17–38. [Google Scholar] [CrossRef]
- Imaichi, R.; Kato, M. Developmental Anatomy of the Shoot Apical Cell, Rhizophore and Root of Selaginella uncinate. Bot. Mag. 1989, 102, 369–380. [Google Scholar] [CrossRef]
- Jones, C.S.; Drinnan, A.N. The Developmental Pattern of Shoot Apices in Selaginella kraussiana (Kunze) A. Braun. Int. J. Plant Sci. 2009, 170, 1009–1018. [Google Scholar] [CrossRef]
- Ambrose, B.A. The Morphology and Development of Lycophytes. Evol. Plant 2012, 45, 91–114. [Google Scholar]
- Gifford, E.M.; Wetmore, R.H. Apical meristems of vegetative shoots and strobili in certain gymnosperms. Proc. Natl. Acad. Sci. USA 1957, 43, 571–576. [Google Scholar] [CrossRef]
- Imaichi, R.; Hiratsuka, R. Evolution of Shoot Apical Meristem Structures in Vascular Plants with Respect to Plasmodesmatal Network. Am. J. Bot. 2007, 94, 1911–1921. [Google Scholar] [CrossRef]
- Niklas, K.J.; Wayne, R.; Benítez, M.; Newman, S.A. Polarity, Planes of Cell Division, and the Evolution of Plant Multicellularity. Protoplasma 2019, 256, 585–599. [Google Scholar] [CrossRef]
- Stewart, K.D.; Mattox, K.R. Some Aspects of Mitosis in Primitive Green Algae: Phylogeny and Function. Biosystems 1975, 7, 310–315. [Google Scholar] [CrossRef]
- Graham, L.E.; Cook, M.E.; Busse, J.S. The Origin of Plants: Body Plan Changes Contributing to a Major Evolutionary Radiation. Proc. Natl. Acad. Sci. USA 2000, 97, 4535–4540. [Google Scholar] [CrossRef]
- Umen, J.G. Green Algae and the Origins of Multicellularity in the Plant Kingdom. Cold Spring Harb. Perspect. Biol. 2014, 6, a016170. [Google Scholar] [CrossRef]
- Graham, L.E. Coleochaete and the origin of land plants. Am. J. Bot. 1984, 71, 603–608. [Google Scholar] [CrossRef]
- Puttick, M.N.; Morris, J.L.; Williams, T.A.; Cox, C.J.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Schneider, H.; Pisani, D.; et al. The Interrelationships of Land Plants and the Nature of the Ancestral Embryophyte. Curr. Biol. 2018, 28, 733–745.e2. [Google Scholar] [CrossRef]
- Nishiyama, T.; Wolf, P.G.; Kugita, M.; Sinclair, R.B.; Sugita, M.; Sugiura, C.; Wakasugi, T.; Yamada, K.; Yoshinaga, K.; Yamaguchi, K.; et al. Chloroplast Phylogeny Indicates That Bryophytes Are Monophyletic. Mol. Biol. Evol. 2004, 21, 1813–1819. [Google Scholar] [CrossRef]
- Renzaglia, K.S.; Garbary, D.J. Motile Gametes of Land Plants: Diversity, Development, and Evolution. Crit. Rev. Plant Sci. 2010, 20, 107–213. [Google Scholar] [CrossRef]
- Campbell, D.H. The Structure & Development of the Mosses & Ferns (Archegoniatae); Macmillan: New York, NY, USA, 1895. [Google Scholar]
- Tomescu, A.M.F.; Wyatt, S.E.; Hasebe, M.; Rothwell, G.W. Early Evolution of the Vascular Plant Body Plan—The Missing Mechanisms. Curr. Opin. Plant Biol. 2014, 17, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Ligrone, R.; Duckett, J.G.; Renzaglia, K.S. Conducting Tissues and Phyletic Relationships of Bryophytes. Philos. Trans. R Soc. Lond. B Biol. Sci. 2000, 355, 795–813. [Google Scholar] [CrossRef]
- Sakakibara, K.; Nishiyama, T.; Deguchi, H.; Hasebe, M. Class 1 KNOX Genes Are Not Involved in Shoot Development in the Moss Physcomitrella Patens but Do Function in Sporophyte Development. Evol. Dev. 2008, 10, 555–566. [Google Scholar] [CrossRef]
- Kato, M.; Akiyama, H. Interpolation Hypothesis for Origin of the Vegetative Sporophyte of Land Plants. Taxon 2005, 54, 443–450. [Google Scholar] [CrossRef]
- Renzaglia, K.S.; Schuette, S.; Duff, R.J.; Ligrone, R.; Shaw, A.J.; Mishler, B.D.; Duckett, J.G. Bryophyte Phylogeny: Advancing the Molecular and Morphological Frontiers. BioOne 2007, 110, 179–213. [Google Scholar] [CrossRef]
- Solly, J.E.; Cunniffe, N.J.; Harrison, C.J. Regional Growth Rate Differences Specified by Apical Notch Activities Regulate Liverwort Thallus Shape. Curr. Biol. 2017, 27, 16–26. [Google Scholar] [CrossRef]
- Renzaglia, A.; Wehman, P.; Schutz, R.; Karan, O. Use of Cue Redundancy and Positive Reinforcement to Accelerate Production in Two Profoundly Retarded Workers. Br. J. Soc. Clin. Psychol. 1978, 17, 183–187. [Google Scholar] [CrossRef]
- Esau, K. Plant Anatomy; Wiley: Hoboken, NJ, USA, 1965. [Google Scholar]
- Gifford, E.M.; Corson, G.E. The Shoot Apex in Seed Plants. Bot. Rev. 1971, 37, 143–229. [Google Scholar] [CrossRef]
- Newman, I.V. Pattern in the Meristems of Vascular Plants: III. Pursuing the Patterns in the Apical Meristem Where No Cell Is a Permanent Cell. J. Linn. Soc. Lond. Bot. 1965, 59, 185–214. [Google Scholar] [CrossRef]
- Fouracre, J.P.; Harrison, C.J. How Was Apical Growth Regulated in the Ancestral Land Plant? Insights from the Development of Non-Seed Plants. Plant Physiol. 2022, 190, 100–112. [Google Scholar] [CrossRef]
- Spencer, V.; Nemec Venza, Z.; Harrison, C.J. What Can Lycophytes Teach Us about Plant Evolution and Development? Modern Perspectives on an Ancient Lineage. Evol. Dev. 2021, 23, 174–196. [Google Scholar] [CrossRef]
- Harrison, C.J.; Rezvani, M.; Langdale, J.A. Growth from Two Transient Apical Initials in the Meristem of Selaginella Kraussiana. Development 2007, 134, 881–889. [Google Scholar] [CrossRef]
- Ambrose, B.A.; Vasco, A. Bringing the Multicellular Fern Meristem into Focus. New Phytol. 2016, 210, 790–793. [Google Scholar] [CrossRef] [PubMed]
- White, R.A.; Turner, M.D. Anatomy and Development of the Fern Sporophyte. Bot. Rev. 1995, 61, 281–305. [Google Scholar] [CrossRef]
- Gifford, E.M. The Structure and Development of the Shoot Apex in Certain Woody Ranales. Am. J. Bot. 1950, 37, 595–611. [Google Scholar] [CrossRef]
- Kessler, S.; Townsley, B.; Sinha, N. L1 Division and Differentiation Patterns Influence Shoot Apical Meristem Maintenance. Plant Physiol. 2006, 141, 1349. [Google Scholar] [CrossRef]
- Sinha, N.; Hake, S. Mutant Characters of Knotted Maize Leaves Are Determined in the Innermost Tissue Layers. Dev. Biol. 1990, 141, 203–210. [Google Scholar] [CrossRef]
- Barton, M.K. Twenty Years on: The Inner Workings of the Shoot Apical Meristem, a Developmental Dynamo. Dev. Biol. 2010, 341, 95–113. [Google Scholar] [CrossRef]
- Kebrom, T.H. A Growing Stem Inhibits Bud Outgrowth—The Overlooked Theory of Apical Dominance. Front. Plant Sci. 2017, 8, 309506. [Google Scholar] [CrossRef] [PubMed]
- Thimann, K.V. Auxins and the Inhibition of Plant Growth. Biol. Rev. 1939, 14, 314–337. [Google Scholar] [CrossRef]
- Went, F.W. Auxin, the Plant Growth-Hormone. Review 1935, 1, 162–182. [Google Scholar] [CrossRef]
- Sussex, I.M.; Kerk, N.M. The Evolution of Plant Architecture. Curr. Opin. Plant Biol. 2001, 4, 33–37. [Google Scholar] [CrossRef]
- Cline, M.G. Execution of the Auxin Replacement Apical Dominance Experiment in Temperate Woody Species. Am. J. Bot. 2000, 87, 182–190. [Google Scholar] [CrossRef]
- Norstog, K.; Nicholls, T.J. The Biology of the Cycads, 1st ed.; Cornell University Press: New York, NY, USA, 1998. [Google Scholar] [CrossRef]
- Napoli, C.A.; Beveridge, C.A.; Snowden, K.C. 5 Reevaluating Concepts of Apical Dominance and the Control of Axillary Bud Outgrowth. Curr. Top. Dev. Biol. 1998, 44, 127–169. [Google Scholar]
- Zhao, Y. Auxin Biosynthesis and Its Role in Plant Development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef]
- Azizi, P.; Rafii, M.Y.; Maziah, M.; Abdullah, S.N.A.; Hanafi, M.M.; Latif, M.A.; Rashid, A.A.; Sahebi, M. Understanding the Shoot Apical Meristem Regulation: A Study of the Phytohormones, Auxin and Cytokinin, in Rice. Mech. Dev. 2015, 135, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pagès, V.; Dun, E.A.; Pillot, J.P.; Letisse, F.; Matusova, R.; Danoun, S.; Portais, J.C.; et al. Strigolactone Inhibition of Shoot Branching. Nature 2008, 455, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.; et al. Inhibition of Shoot Branching by New Terpenoid Plant Hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Bertheloot, J.; Barbier, F.; Boudon, F.; Perez-Garcia, M.D.; Péron, T.; Citerne, S.; Dun, E.; Beveridge, C.; Godin, C.; Sakr, S. Sugar Availability Suppresses the Auxin-Induced Strigolactone Pathway to Promote Bud Outgrowth. New Phytol. 2020, 225, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Mazur, E.; Balla, J.; Gallei, M.; Kalousek, P.; Medveďová, Z.; Li, Y.; Wang, Y.; Prát, T.; Vasileva, M.; et al. Strigolactones Inhibit Auxin Feedback on PIN-Dependent Auxin Transport Canalization. Nat. Commun. 2020, 11, 3508. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, Y.; Ma, M.; Zhou, Q.; Zhao, Y.; Zhao, B.; Wang, B.; Wei, H.; Wang, H. Arabidopsis FHY3 and FAR1 Integrate Light and Strigolactone Signaling to Regulate Branching. Nat. Commun. 2020, 11, 1955. [Google Scholar] [CrossRef]
- Wu, F.; Gao, Y.; Yang, W.; Sui, N.; Zhu, J. Biological Functions of Strigolactones and Their Crosstalk With Other Phytohormones. Front. Plant Sci. 2022, 13, 821563. [Google Scholar] [CrossRef] [PubMed]
- López-Ráez, J.A.; Charnikhova, T.; Gómez-Roldán, V.; Matusova, R.; Kohlen, W.; De Vos, R.; Verstappen, F.; Puech-Pages, V.; Bécard, G.; Mulder, P.; et al. Tomato Strigolactones Are Derived from Carotenoids and Their Biosynthesis Is Promoted by Phosphate Starvation. New Phytol. 2008, 178, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, N.; Taylor, C.; Leyser, O. Strigolactone Can Promote or Inhibit Shoot Branching by Triggering Rapid Depletion of the Auxin Efflux Protein PIN1 from the Plasma Membrane. PLoS Biol. 2013, 11, e1001474. [Google Scholar] [CrossRef]
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer, K.F.X.; Jürgens, G.; Laux, T. The Stem Cell Population of Arabidopsis Shoot Meristems Is Maintained by a Regulatory Loop between the CLAVATA and WUSCHEL Genes. Cell 2000, 100, 635–644. [Google Scholar] [CrossRef]
- Laux, T.; Mayer, K.F.X.; Berger, J.; Jürgens, G. The WUSCHEL Gene Is Required for Shoot and Floral Meristem Integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar] [CrossRef]
- Long, J.A.; Moan, E.I.; Medford, J.I.; Barton, M.K. A Member of the KNOTTED Class of Homeodomain Proteins Encoded by the STM Gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Endrizzi, K.; Moussian, B.; Haecker, A.; Levin, J.Z.; Laux, T. The SHOOT MERISTEMLESS Gene Is Required for Maintenance of Undifferentiated Cells in Arabidopsis Shoot and Floral Meristems and Acts at a Different Regulatory Level than the Meristem Genes WUSCHEL and ZWILLE. Plant J. 1996, 10, 967–979. [Google Scholar] [CrossRef]
- Zhou, X.; Guo, Y.; Zhao, P.; Sun, M.X. Comparative Analysis of WUSCHEL-Related Homeobox Genes Revealed Their Parent-of-Origin and Cell Type-Specific Expression Pattern during Early Embryogenesis in Tobacco. Front. Plant Sci. 2018, 9, 337885. [Google Scholar] [CrossRef]
- Aida, M.; Tasaka, M. Genetic Control of Shoot Organ Boundaries. Curr. Opin. Plant Biol. 2006, 9, 72–77. [Google Scholar] [CrossRef]
- Zhang, W. Putting Genes on the Map: Spatial Transcriptomics of the Maize Shoot Apical Meristem. Plant Physiol. 2022, 188, 1931–1932. [Google Scholar] [CrossRef] [PubMed]
- Shimotohno, A.; Scheres, B. Topology of Regulatory Networks That Guide Plant Meristem Activity: Similarities and Differences. Curr. Opin. Plant Biol. 2019, 51, 74–80. [Google Scholar] [CrossRef]
- Takahashi, G.; Kiyosue, T.; Hirakawa, Y. Control of Stem Cell Behavior by CLE-JINGASA Signaling in the Shoot Apical Meristem in Marchantia polymorpha. bioRxiv 2023. [Google Scholar] [CrossRef]
- Fletcher, J.C. The CLV-WUS Stem Cell Signaling Pathway: A Roadmap to Crop Yield Optimization. Plants 2018, 7, 87. [Google Scholar] [CrossRef]
- Floyd, S.K.; Bowman, J.L. The Ancestral Developmental Tool Kit of Land Plants. Int. J. Plant Sci. 2007, 168, 1–35. [Google Scholar] [CrossRef]
- Dierschke, T.; Flores-Sandoval, E.; Rast-Somssich, M.I.; Althoff, F.; Zachgo, S.; Bowman, J.L. Gamete Expression of Tale Class Hd Genes Activates the Diploid Sporophyte Program in Marchantia polymorpha. Elife 2021, 10, e57088. [Google Scholar] [CrossRef]
- Hisanaga, T.; Fujimoto, S.; Cui, Y.; Sato, K.; Sano, R.; Yamaoka, S.; Kohchi, T.; Berger, F.; Nakajima, K. Deep Evolutionary Origin of Gamete-Directed Zygote Activation by KNOX/BELL Transcription Factors in Green Plants. Elife 2021, 10, e57090. [Google Scholar] [CrossRef]
- Lee, J.H.; Lin, H.; Joo, S.; Goodenough, U. Early Sexual Origins of Homeoprotein Heterodimerization and Evolution of the Plant KNOX/BELL Family. Cell 2008, 133, 829–840. [Google Scholar] [CrossRef]
- Harrison, C.J.; Coriey, S.B.; Moylan, E.C.; Alexander, D.L.; Scotland, R.W.; Langdale, J.A. Independent Recruitment of a Conserved Developmental Mechanism during Leaf Evolution. Nature 2005, 434, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Kawai, J.; Tanabe, Y.; Soma, S.; Ito, M. Class 1 KNOX Gene Expression Supports the Selaginella Rhizophore Concept. J. Plant Biol. 2010, 53, 268–274. [Google Scholar] [CrossRef]
- Alvarez-Buylla, E.R.; Liljegren, S.J.; Pelaz, S.; Gold, S.E.; Burgeff, C.; Ditta, G.S.; Vergara-Silva, F.; Yanofsky, M.F. MADS-Box Gene Evolution beyond Flowers: Expression in Pollen, Endosperm, Guard Cells, Roots and Trichomes. Plant J. 2000, 24, 457–466. [Google Scholar]
- Henschel, K.; Kofuji, R.; Hasebe, M.; Saedler, H.; Münster, T.; Theißen, G. Two Ancient Classes of MIKC-Type MADS-Box Genes Are Present in the Moss Physcomitrella Patens. Mol. Biol. Evol. 2002, 19, 801–814. [Google Scholar] [CrossRef]
- Tanabe, Y.; Hasebe, M.; Sekimoto, H.; Nishiyama, T.; Kitani, M.; Henschel, K.; Münster, T.; Theissen, G.; Nozaki, H.; Ito, M. Characterization of MADS-Box Genes in Charophycean Green Algae and Its Implication for the Evolution of MADS-Box Genes. Proc. Natl. Acad. Sci. USA 2005, 102, 2436–2441. [Google Scholar] [CrossRef] [PubMed]
- Singer, S.D.; Krogan, N.T.; Ashton, N.W. Clues about the Ancestral Roles of Plant MADS-Box Genes from a Functional Analysis of Moss Homologues. Plant Cell Rep. 2007, 26, 1155–1169. [Google Scholar] [CrossRef]
- Koshimizu, S.; Kofuji, R.; Sasaki-Sekimoto, Y.; Kikkawa, M.; Shimojima, M.; Ohta, H.; Shigenobu, S.; Kabeya, Y.; Hiwatashi, Y.; Tamada, Y.; et al. Physcomitrella MADS-Box Genes Regulate Water Supply and Sperm Movement for Fertilization. Nat. Plants 2018, 4, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Thangavel, G.; Nayar, S. A Survey of MIKC Type MADS-Box Genes in Non-Seed Plants: Algae, Bryophytes, Lycophytes and Ferns. Front. Plant Sci. 2018, 9, 342764. [Google Scholar] [CrossRef]
- Becker, A.; Theißen, G. The Major Clades of MADS-Box Genes and Their Role in the Development and Evolution of Flowering Plants. Mol. Phylogenet. Evol. 2003, 29, 464–489. [Google Scholar] [CrossRef]
- Ferrario, S.; Shchennikova, A.V.; Franken, J.; Immink, R.G.H.; Angenent, G.C. Control of Floral Meristem Determinacy in Petunia by MADS-Box Transcription Factors. Plant Physiol. 2006, 140, 890–898. [Google Scholar] [CrossRef]
- Zobell, O.; Faigl, W.; Saedler, H.; Münster, T. MIKC* MADS-Box Proteins: Conserved Regulators of the Gametophytic Generation of Land Plants. Mol. Biol. Evol. 2010, 27, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, B.A.; Smalls, T.L.; Zumajo-Cardona, C. All Type II Classic MADS-Box Genes in the Lycophyte Selaginella Moellendorffii Are Broadly yet Discretely Expressed in Vegetative and Reproductive Tissues. Evol. Dev. 2021, 23, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Meyerowitz, E.M. The AP2/EREBP Family of Plant Transcription Factors. Biol. Chem. 1998, 379, 633–654. [Google Scholar] [PubMed]
- Gutterson, N.; Reuber, T.L. Regulation of Disease Resistance Pathways by AP2/ERF Transcription Factors. Curr. Opin. Plant Biol. 2004, 7, 465–471. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, R.; Xu, D.; Bi, H.; Xia, Z.; Peng, H. Genome-Wide Identification and Analysis of the AP2 Transcription Factor Gene Family in Wheat (Triticum Aestivum L.). Front. Plant Sci. 2019, 10, 486684. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Soltis, P.S.; Wall, K.; Soltis, D.E. Phylogeny and Domain Evolution in the APETALA2-like Gene Family. Mol. Biol. Evol. 2006, 23, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Zumajo-Cardona, C.; Vasco, A.; Ambrose, B.A. The Evolution of the KANADI Gene Family and Leaf Development in Lycophytes and Ferns. Plants 2019, 8, 313. [Google Scholar] [CrossRef]
- Dipp-álvarez, M.; Cruz-Ramírez, A. A Phylogenetic Study of the ANT Family Points to a PreANT Gene as the Ancestor of Basal and EuANT Transcription Factors in Land Plants. Front. Plant Sci. 2019, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Bui, L.T.; Pandzic, D.; Youngstrom, C.E.; Wallace, S.; Irish, E.E.; Szövényi, P.; Cheng, C.L. A Fern AINTEGUMENTA Gene Mirrors BABY BOOM in Promoting Apogamy in Ceratopteris richardii. Plant J. 2017, 90, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Krizek, B.A.; Bantle, A.T.; Heflin, J.M.; Han, H.; Freese, N.H.; Loraine, A.E. AINTEGUMENTA and AINTEGUMENTA-LIKE6 Directly Regulate Floral Homeotic, Growth, and Vascular Development Genes in Young Arabidopsis Flowers. J. Exp. Bot. 2021, 72, 5478–5493. [Google Scholar] [CrossRef]
- Farris, S.; Wang, Y.; Ward, J.M.; Dudek, S.M. Optimized Method for Robust Transcriptome Profiling of Minute Tissues Using Laser Capture Microdissection and Low-Input RNA-Seq. Front. Mol. Neurosci. 2017, 10, 272672. [Google Scholar] [CrossRef] [PubMed]
- Rakszewska, A.; Tel, J.; Chokkalingam, V.; Huck, W.T.S. One Drop at a Time: Toward Droplet Microfluidics as a Versatile Tool for Single-Cell Analysis. NPG Asia Mater. 2014, 6, e133. [Google Scholar] [CrossRef]
- Scialdone, A.; Natarajan, K.N.; Saraiva, L.R.; Proserpio, V.; Teichmann, S.A.; Stegle, O.; Marioni, J.C.; Buettner, F. Computational Assignment of Cell-Cycle Stage from Single-Cell Transcriptome Data. Methods 2015, 85, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Bendall, S.C.; Davis, K.L.; Amir, E.A.D.; Tadmor, M.D.; Simonds, E.F.; Chen, T.J.; Shenfeld, D.K.; Nolan, G.P.; Pe’Er, D. Single-Cell Trajectory Detection Uncovers Progression and Regulatory Coordination in Human B Cell Development. Cell 2014, 157, 714–725. [Google Scholar] [CrossRef]
- Zhang, T.Q.; Chen, Y.; Wang, J.W. A Single-Cell Analysis of the Arabidopsis Vegetative Shoot Apex. Dev. Cell 2021, 56, 1056–1074.e8. [Google Scholar] [CrossRef] [PubMed]
- Satterlee, J.W.; Strable, J.; Scanlon, M.J. Plant Stem-Cell Organization and Differentiation at Single-Cell Resolution. Proc. Natl. Acad. Sci. USA 2020, 117, 33689–33699. [Google Scholar] [CrossRef]
- Chen, X.; Ru, Y.; Takahashi, H.; Nakazono, M.; Shabala, S.; Smith, S.M.; Yu, M. Single-Cell Transcriptomic Analysis of Pea Shoot Development and Cell-Type-Specific Responses to Boron Deficiency. bioRxiv 2023. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Du, Q.; Xu, M.; Du, F.; Jiao, Y. Single-Nucleus RNA-Seq Resolves Spatiotemporal Developmental Trajectories in the Tomato Shoot Apex. bioRxiv 2020. [Google Scholar] [CrossRef]
- Conde, D.; Triozzi, P.M.; Pereira, W.J.; Schmidt, H.W.; Balmant, K.M.; Knaack, S.A.; Redondo-López, A.; Roy, S.; Dervinis, C.; Kirst, M. Single-Nuclei Transcriptome Analysis of the Shoot Apex Vascular System Differentiation in Populus. Development 2022, 149, dev200632. [Google Scholar] [CrossRef]
- Bommert, P.; Lunde, C.; Nardmann, J.; Vollbrecht, E.; Running, M.; Jackson, D.; Hake, S.; Werr, W. Thick Tassel Dwarf1 Encodes a Putative Maize Ortholog of the Arabidopsis CLAVATA1 Leucine-Rich Repeat Receptor-like Kinase. Development 2005, 132, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Xu, F.; Jackson, D. All Together Now, a Magical Mystery Tour of the Maize Shoot Meristem. Curr. Opin. Plant Biol. 2018, 45, 26–35. [Google Scholar] [CrossRef]
- Somssich, M.; Je, B.I.; Simon, R.; Jackson, D. CLAVATA-WUSCHEL Signaling in the Shoot Meristem. Development 2016, 143, 3238–3248. [Google Scholar] [CrossRef]
- Nardmann, J.; Werr, W. The Invention of WUS-like Stem Cell-Promoting Functions in Plants Predates Leptosporangiate Ferns. Plant Mol. Biol. 2012, 78, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Laureyns, R.; Joossens, J.; Herwegh, D.; Pevernagie, J.; Pavie, B.; Demuynck, K.; Debray, K.; Coussens, G.; Pauwels, L.; van Hautegem, T.; et al. An in Situ Sequencing Approach Maps PLASTOCHRON1 at the Boundary between Indeterminate and Determinate Cells. Plant Physiol. 2022, 188, 782–794. [Google Scholar] [CrossRef] [PubMed]
- Ståhl, P.L.; Salmén, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and Analysis of Gene Expression in Tissue Sections by Spatial Transcriptomics. Science 2016, 353, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, S.; Lundeberg, J. Preparation of Plant Tissue to Enable Spatial Transcriptomics Profiling Using Barcoded Microarrays. Nat. Protoc. 2018, 13, 2425–2446. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, S.; Salmén, F.; Terebieniec, B.K.; Vickovic, S.; Navarro, J.F.; Alexeyenko, A.; Reimegård, J.; McKee, L.S.; Mannapperuma, C.; Bulone, V.; et al. Spatially Resolved Transcriptome Profiling in Model Plant Species. Nat Plants 2017, 3, 17061. [Google Scholar] [CrossRef]
- Svensson, V.; Natarajan, K.N.; Ly, L.H.; Miragaia, R.J.; Labalette, C.; Macaulay, I.C.; Cvejic, A.; Teichmann, S.A. Power Analysis of Single-Cell RNA-Sequencing Experiments. Nat. Methods 2017, 14, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; Monserrate, F.A.; Ramírez-Villegas, J.; Madriñán, S.; Blair, M.W. Drought Tolerance in Wild Plant Populations: The Case of Common Beans (Phaseolus vulgaris L.). PLoS ONE 2013, 8, e62898. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; This, D.; Chavarro, C.; Madriñán, S.; Blair, M.W. Nucleotide Diversity Patterns at the Drought-Related DREB2 Encoding Genes in Wild and Cultivated Common Bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2012, 125, 1069–1085. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Clade | Technology | Validation | Focus | Reference |
|---|---|---|---|---|---|
| Arabidopsis thaliana | Dicot | scRNA-seq | GFP-reporter lines of identified genes (At2g38300, At4g34970 (ADF9), At4g11290, and At1g13650). | Construction of developmental trajectories of several tissues and Integrative. | [145] |
| Zea mays | Monocot | scRNA-seq | In situ hybridization of KN1-overexpressor lines. | Analysis of maize SAM stem cell function and cell-fate acquisition. | [146] |
| Pisum sativum | Dicot | scRNA-seq | Laser capture microdissection to identify marker genes. | Cell type-specific responses to boron deficiency. | [147] |
| Solanum lycopersicum | Dicot | snRNA-seq | Homologous markers identification. | Construction of developmental trajectories of several tissues. | [148] |
| Populus trichocarpa | Dicot | snRNA-seq | RNA in situ hybridization and GUS-reporter lines. | Comparisons on Phloem and xylem development between a woody plant and annual herb. | [149] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kean-Galeno, T.; Lopez-Arredondo, D.; Herrera-Estrella, L. The Shoot Apical Meristem: An Evolutionary Molding of Higher Plants. Int. J. Mol. Sci. 2024, 25, 1519. https://doi.org/10.3390/ijms25031519
Kean-Galeno T, Lopez-Arredondo D, Herrera-Estrella L. The Shoot Apical Meristem: An Evolutionary Molding of Higher Plants. International Journal of Molecular Sciences. 2024; 25(3):1519. https://doi.org/10.3390/ijms25031519
Chicago/Turabian StyleKean-Galeno, Tania, Damar Lopez-Arredondo, and Luis Herrera-Estrella. 2024. "The Shoot Apical Meristem: An Evolutionary Molding of Higher Plants" International Journal of Molecular Sciences 25, no. 3: 1519. https://doi.org/10.3390/ijms25031519
APA StyleKean-Galeno, T., Lopez-Arredondo, D., & Herrera-Estrella, L. (2024). The Shoot Apical Meristem: An Evolutionary Molding of Higher Plants. International Journal of Molecular Sciences, 25(3), 1519. https://doi.org/10.3390/ijms25031519