

Alterations in Plasma Lipid Profiles Associated with Melanoma and Therapy Resistance

 , , , , ,
, , , , ,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
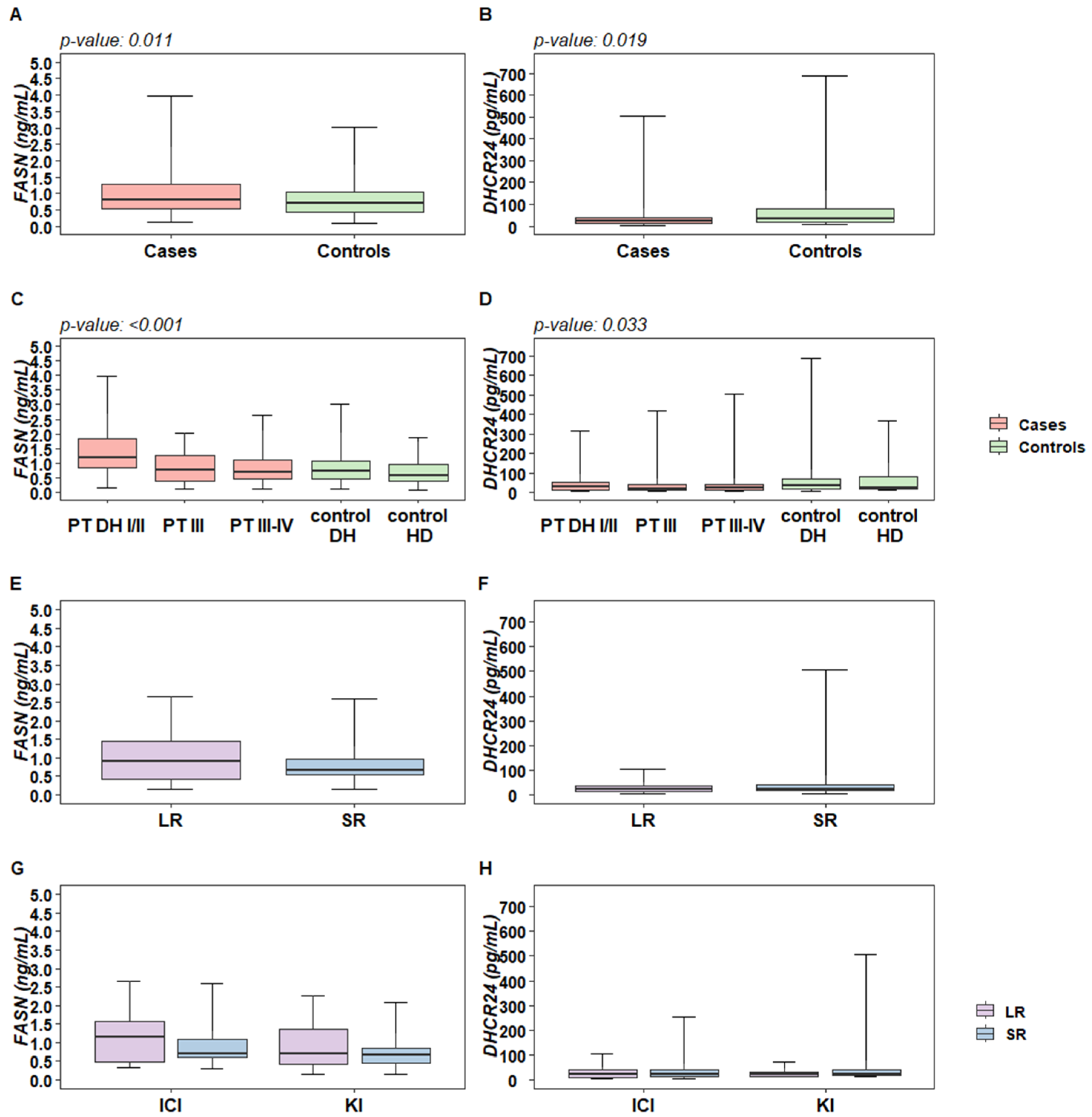
2.1. Plasma Levels of DHCR24 and FASN Enzymes Are Altered in Melanoma Patients Compared to Control Subjects
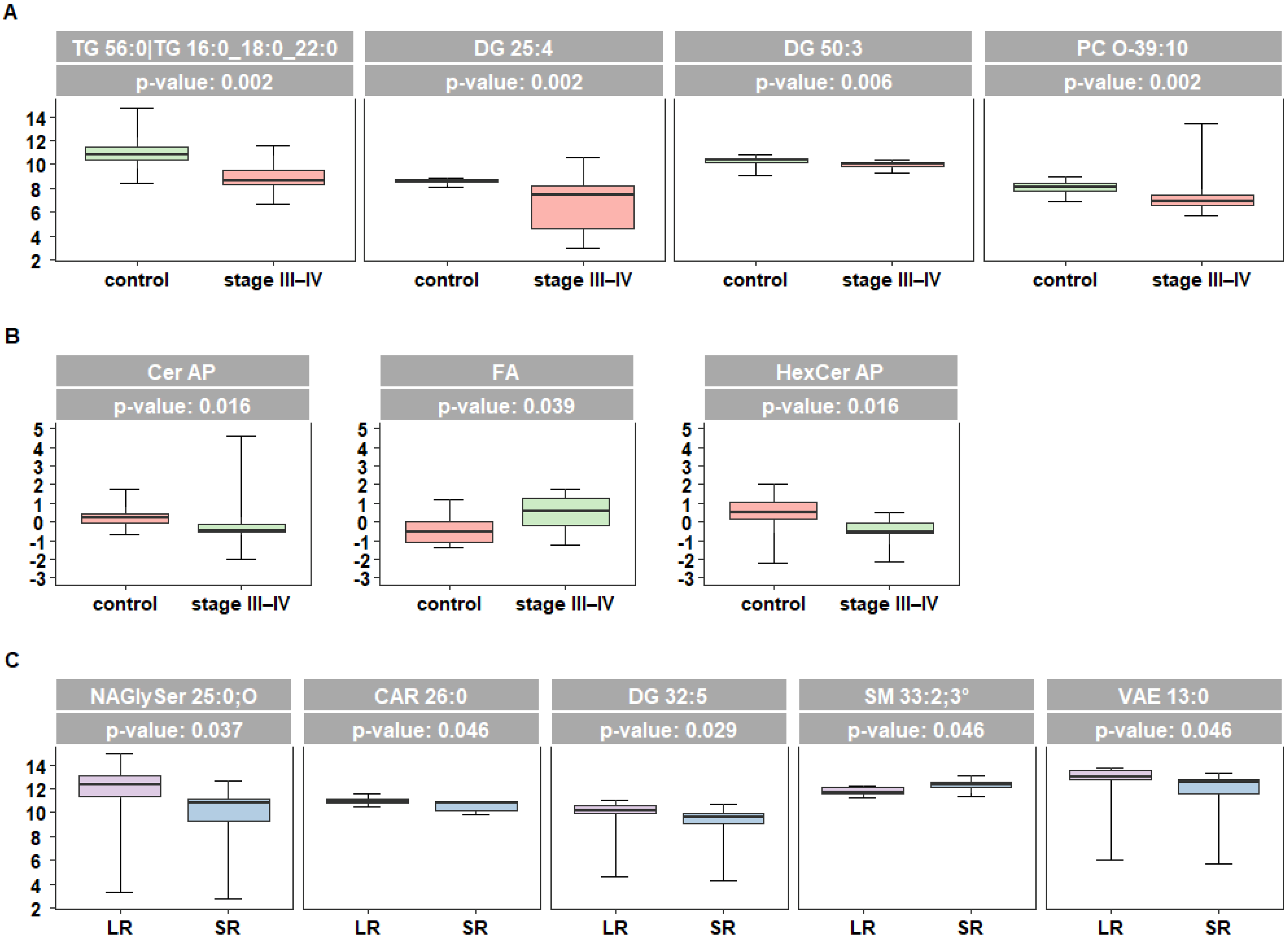
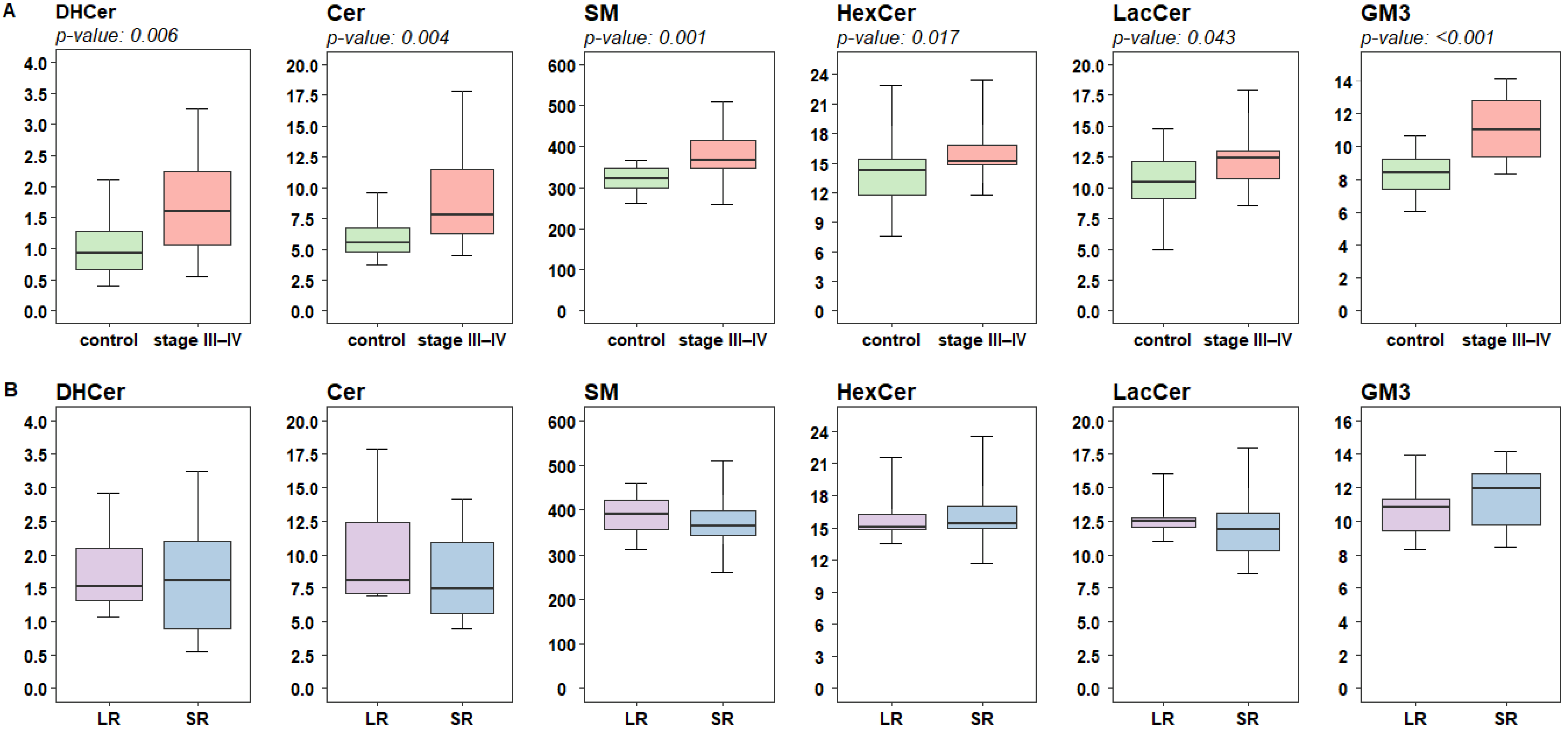
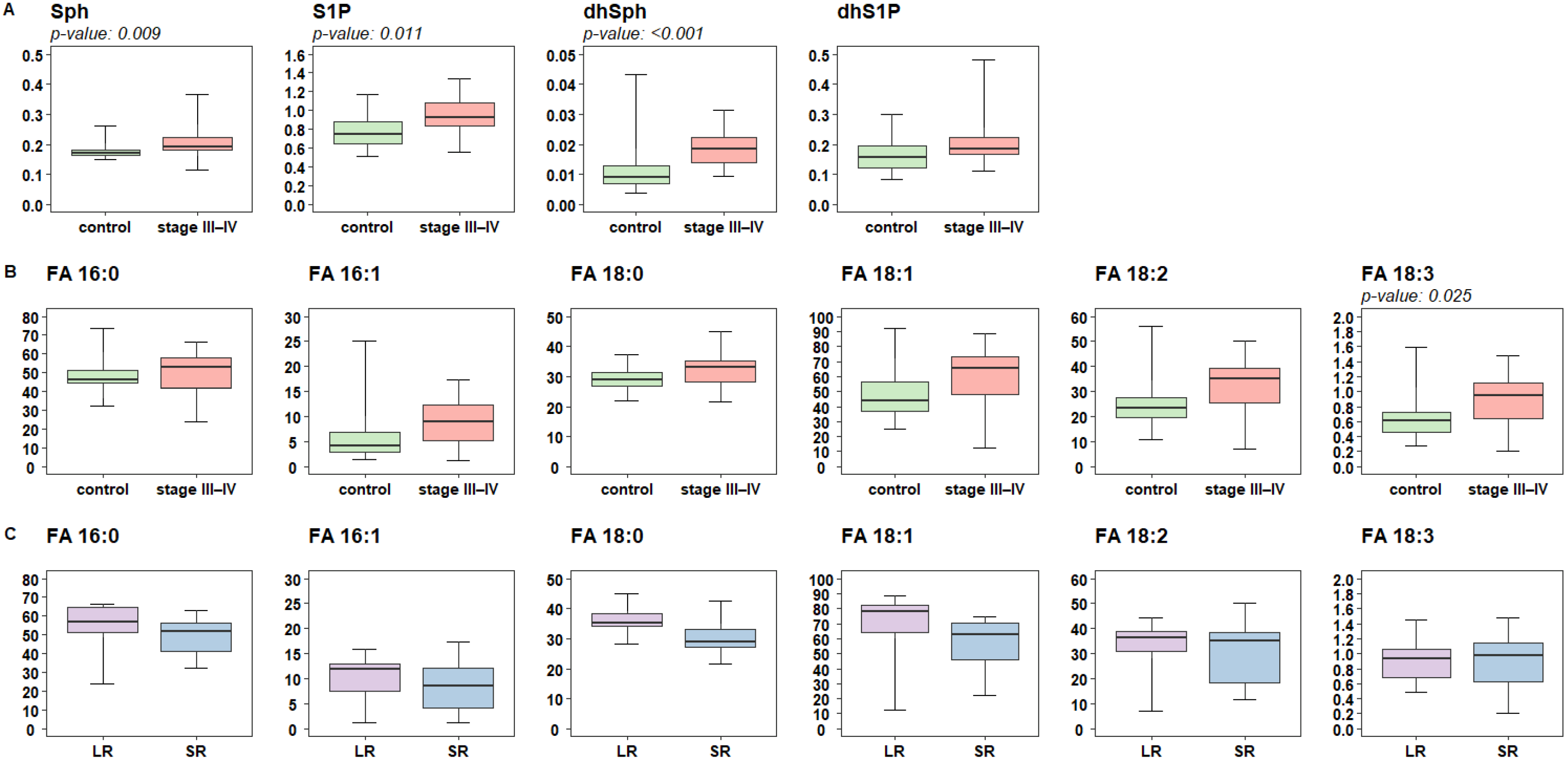
2.2. Lipidomic Profiling of Plasma Reveals Altered Levels of Fatty Acids Derivatives and of Sphingolipids in Melanoma Patients
2.3. Targeted Analysis of Fatty Acid and Sphingolipid Species Identify Quantitative Differences in Patients Responsive to Targeted Therapy
3. Discussion
4. Materials and Methods
4.1. Plasma Samples
4.2. Measurement of FASN and DHCR24 Plasma Levels
4.3. Chemicals and Reagents for Lipidomics
4.4. Untargeted Lipidomics
4.5. Lipidomics Data Processing
4.6. Targeted Lipidomics for Sphingolipids and Free Fatty Acids Quantification
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villani, A.; Scalvenzi, M.; Micali, G.; Lacarrubba, F.; Fornaro, L.; Martora, F.; Potestio, L. Management of Advanced Invasive Melanoma: New Strategies. Adv. Ther. 2023, 40, 3381–3394. [Google Scholar] [CrossRef]
- Seth, R.; Agarwala, S.S.; Messersmith, H.; Alluri, K.C.; Ascierto, P.A.; Atkins, M.B.; Bollin, K.; Chacon, M.; Davis, N.; Faries, M.B.; et al. Systemic Therapy for Melanoma: ASCO Guideline Update. J. Clin. Oncol. 2023, 41, 4794–4820. [Google Scholar] [CrossRef]
- Falletta, P.; Goding, C.R.; Vivas-García, Y. Connecting metabolic rewiring with phenotype switching in melanoma. Front. Cell Dev. Biol. 2022, 10, 930250. [Google Scholar] [CrossRef]
- Pellerin, L.; Carrié, L.; Dufau, C.; Nieto, L.; Ségui, B.; Levade, T.; Riond, J.; Andrieu-Abadie, N. Lipid metabolic Reprogramming: Role in Melanoma Progression and Therapeutic Perspectives. Cancers 2020, 12, 3147. [Google Scholar] [CrossRef]
- Wilcock, D.J.; Badrock, A.P.; Wong, C.W.; Owen, R.; Guerin, M.; Southam, A.D.; Johnston, H.; Telfer, B.A.; Fullwood, P.; Watson, J.; et al. Oxidative stress from DGAT1 oncoprotein inhibition in melanoma suppresses tumor growth when ROS defenses are also breached. Cell Rep. 2022, 39, 110995. [Google Scholar] [CrossRef]
- Shen, S.; Faouzi, S.; Souquere, S.; Roy, S.; Routier, E.; Libenciuc, C.; André, F.; Pierron, G.; Scoazec, J.Y.; Robert, C. Melanoma persister cells are tolerant to BRAF/MEK Inhibitors via ACOX1-mediated Fatty Acid Oxidation. Cell Rep. 2020, 33, 108421. [Google Scholar] [CrossRef]
- Vivas-García, Y.; Falletta, P.; Liebing, J.; Louphrasitthiphol, P.; Feng, Y.; Chauhan, J.; Scott, D.A.; Glodde, N.; Chocarro-Calvo, A.; Bonham, S.; et al. Lineage-Restricted Regulation of SCD and Fatty Acid Saturation by MITF Controls Melanoma Phenotypic Plasticity. Mol. Cell 2020, 77, 120–137. [Google Scholar] [CrossRef]
- Garandeau, D.; Mrad, M.; Levade, T.; Perrotta, C.; Andrieu-Abadie, N.; Diab-Assaf, M. Dysregulation of Sphingolipid Metabolism in Melanoma: Roles in Pigmentation, Cell Survival and Tumor Progression. In Bioactive Sphingolipids in Cancer Biology and Therapy; Hannun, Y., Luberto, C., Mao, C., Obeid, L., Eds.; Springer: Cham, Switzerland, 2015; pp. 123–139. [Google Scholar] [CrossRef]
- Carrié, L.; Virazels, M.; Dufau, C.; Montfort, A.; Levade, T.; Ségui, B.; Andrieu-Abadie, N. New Insights into the Role of Sphingolipid Metabolism in Melanoma. Cells 2020, 9, 1967. [Google Scholar] [CrossRef] [PubMed]
- Vergani, E.; Busico, A.; Dugo, M.; Devecchi, A.; Valeri, B.; Cossa, M.; Di Guardo, L.; De Cecco, L.; Feltrin, E.; Valle, G.; et al. Genetic Layout of Melanoma Lesions Is Associated with BRAF/MEK-Targeted Therapy Resistance and Transcriptional Profiles. J. Investig. Dermatol. 2022, 142, 3030–3040.e5. [Google Scholar] [CrossRef] [PubMed]
- Vergani, E.; Beretta, G.L.; Aloisi, M.; Costantino, M.; Corno, C.; Frigerio, S.; Tinelli, S.; Dugo, M.; Accattatis, F.M.; Granata, A.; et al. Targeting of the Lipid Metabolism Impairs Resistance to BRAF Kinase Inhibitor in Melanoma. Front. Cell Dev. Biol. 2022, 10, 927118. [Google Scholar] [CrossRef] [PubMed]
- Stamatakos, S.; Beretta, G.L.; Vergani, E.; Dugo, M.; Corno, C.; Corna, E.; Tinelli, S.; Frigerio, S.; Ciusani, E.; Rodolfo, M.; et al. Deregulated FASN Expression in BRAF Inhibitor-Resistant Melanoma Cells Unveils New Targets for Drug Combinations. Cancers 2021, 13, 2284. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiù, V.; Leuti, A.; Maccarrone, M. Bioactive Lipids and Chronic Inflammation: Managing the Fire Within. Front. Immunol. 2018, 9, 38. [Google Scholar] [CrossRef]
- Hussain, G.; Anwar, H.; Rasul, A.; Imran, A.; Qasim, M.; Zafar, S.; Imran, M.; Kamran, S.K.S.; Aziz, N.; Razzaq, A.; et al. Lipids as biomarkers of brain disorders. Crit. Rev. Food Sci. Nutr. 2020, 60, 351–374. [Google Scholar] [CrossRef]
- van Kruining, D.; Luo, Q.; van Echten-Deckert, G.; Mielke, M.M.; Bowman, A.; Ellis, S.; Oliveira, T.G.; Martinez-Martinez, P. Sphingolipids as prognostic biomarkers of neurodegeneration, neuroinflammation, and psychiatric diseases and their emerging role in lipidomic investigation methods. Adv. Drug Deliv. Rev. 2020, 159, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Neagu, M. Metabolic Traits in Cutaneous Melanoma. Front. Oncol. 2020, 10, 851. [Google Scholar] [CrossRef]
- Molinelli, E.; Ceccarelli, G.; Fantone, S.; Di Mercurio, E.; Gambini, D.; Maurizi, A.; Perugini, J.; Tossetta, G.; Brisigotti, V.; De Simoni, E.; et al. Melanoma and subcutaneous adipose tissue: Role of peritumoral adipokines in disease characterization and prognosis. Pigment Cell Melanoma Res. 2023, 36, 423–430. [Google Scholar] [CrossRef]
- Lazar, I.; Clement, E.; Dauvillier, S.; Milhas, D.; Ducoux-Petit, M.; LeGonidec, S.; Moro, C.; Soldan, V.; Dalle, S.; Balor, S.; et al. Adipocyte Exosomes Promote Melanoma Aggressiveness through Fatty Acid Oxidation: A Novel Mechanism Linking Obesity and Cancer. Cancer Res. 2016, 76, 4051–4057. [Google Scholar] [CrossRef]
- Zhang, M.; Di Martino, J.S.; Bowman, R.L.; Campbell, N.R.; Baksh, S.C.; Simon-Vermot, T.; Kim, I.S.; Haldeman, P.; Mondal, C.; Yong-Gonzales, V.; et al. Adipocyte-Derived Lipids Mediate Melanoma Progression via FATP Proteins. Cancer Discov. 2018, 8, 1006–1025. [Google Scholar] [CrossRef]
- Alicea, G.M.; Rebecca, V.W.; Goldman, A.R.; Fane, M.E.; Douglass, S.M.; Behera, R.; Webster, M.R.; Kugel, C.H., 3rd; Ecker, B.L.; Caino, M.C.; et al. Changes in Aged Fibroblast Lipid Metabolism Induce Age-Dependent Melanoma Cell Resistance to Targeted Therapy via the Fatty Acid Transporter FATP2. Cancer Discov. 2020, 10, 1282–1295. [Google Scholar] [CrossRef]
- Kuzu, O.F.; Noory, M.A.; Robertson, G.P. The Role of Cholesterol in Cancer. Cancer Res. 2016, 76, 2063–2070. [Google Scholar] [CrossRef]
- Delgado-Goñi, T.; Galobart, T.C.; Wantuch, S.; Normantaite, D.; Leach, M.O.; Whittaker, S.R.; Beloueche-Babari, M. Increased inflammatory lipid metabolism and anaplerotic mitochondrial activation follow acquired resistance to vemurafenib in BRAF-mutant melanoma cells. Br. J. Cancer 2020, 122, 72–81. [Google Scholar] [CrossRef]
- Bataller, M.; Sánchez-García, A.; Garcia-Mayea, Y.; Mir, C.; Rodriguez, I.; LLeonart, M.E. The Role of Sphingolipids Metabolism in Cancer Drug Resistance. Front. Oncol. 2021, 11, 807636. [Google Scholar] [CrossRef]
- Agatonovic-Kustrin, S.; Morton, D.W.; Smirnov, V.; Petukhov, A.; Gegechkori, V.; Kuzina, V.; Gorpinchenko, N.; Ramen-skaya, G. Analytical Strategies in Lipidomics for Discovery of Functional Biomarkers from Human Saliva. Dis. Markers 2019, 2019, 6741518. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lee, H.; Kim, S.H.; Jin, H.; Bae, J.; Choi, H.K. Discovery of potential biomarkers in human melanoma cells with different metastatic potential by metabolic and lipidomic profiling. Sci. Rep. 2017, 7, 8864. [Google Scholar] [CrossRef]
- Sagini, K.; Urbanelli, L.; Buratta, S.; Emiliani, C.; Llorente, A. Lipid Biomarkers in Liquid Biopsies: Novel Opportunities for Cancer Diagnosis. Pharmaceutics 2023, 15, 437. [Google Scholar] [CrossRef]
- Hersey, P. Ganglioside antigens in tissue sections of skin, naevi, and melanoma—Implications for treatment of melanoma. Cancer Treat. Res. 1991, 54, 137–151. [Google Scholar] [CrossRef]
- Garandeau, D.; Noujarède, J.; Leclerc, J.; Imbert, C.; Garcia, V.; Bats, M.L.; Rambow, F.; Gilhodes, J.; Filleron, T.; Meyer, N.; et al. Targeting the Sphingosine 1-Phosphate Axis Exerts Potent Antitumor Activity in BRAFi-Resistant Melanomas. Mol. Cancer Ther. 2019, 18, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Vinceti, M.; Malagoli, C.; Iacuzio, L.; Crespi, C.M.; Sieri, S.; Krogh, V.; Marmiroli, S.; Pellacani, G.; Venturelli, E. Serum Fatty acids and risk of cutaneous melanoma: A population-based case-control study. Dermatol. Res. Pract. 2013, 2013, 659394. [Google Scholar] [CrossRef]
- Liyanage, U.E.; Law, M.H.; Ong, J.S.; Cust, A.E.; Mann, G.J.; Ward, S.V.; Melanoma Meta-Analysis Consortium; Gharahkhani, P.; Iles, M.M.; MacGregor, S. Polyunsaturated fatty acids and risk of melanoma: A Mendelian randomisation analysis. Int. J. Cancer 2018, 143, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Vilbert, M.; Koch, E.C.; Rose, A.A.N.; Laister, R.C.; Gray, D.; Sotov, V.; Penny, S.; Spreafico, A.; Pinto, D.M.; Butler, M.O.; et al. Analysis of the Circulating Metabolome of Patients with Cutaneous, Mucosal and Uveal Melanoma Reveals Distinct Metabolic Profiles with Implications for Response to Immunotherapy. Cancers 2023, 15, 3708. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, D.; Sima, X.; Fu, Y.; Zeng, H.; Hu, Z.; Hou, J.; Pan, Y.; Zhang, Y.; Zhou, Z.; et al. Levels of pretreatment serum lipids predict responses to PD-1 inhibitor treatment in advanced intrahepatic cholangiocarcinoma. Int. Immunopharmacol. 2023, 115, 109687. [Google Scholar] [CrossRef] [PubMed]
- McQuade, J.L.; Daniel, C.R.; Hess, K.R.; Mak, C.; Wang, D.Y.; Rai, R.R.; Park, J.J.; Haydu, L.E.; Spencer, C.; Wongchenko, M.; et al. Association of body-mass index and outcomes in patients with metastatic melanoma treated with targeted therapy, immunotherapy, or chemotherapy: A retrospective, multicohort analysis. Lancet Oncol. 2018, 19, 310–322. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Z.; Isah, A.D.; Chen, S.; Ren, Y.; Cai, H. Lipid metabolism and tumor immunotherapy. Front. Cell Dev. Biol. 2023, 11, 1187989. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, H.; Liu, B.; Wei, J. Fatty acid metabolism and cancer immunotherapy. Curr. Oncol. Rep. 2022, 24, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Dei Cas, M.; Zulueta, A.; Mingione, A.; Caretti, A.; Ghidoni, R.; Signorelli, P.; Paroni, R. An Innovative Lipidomic Workflow to Investigate the Lipid Profile in a Cystic Fibrosis Cell Line. Cells 2020, 9, 1197. [Google Scholar] [CrossRef] [PubMed]
- Morano, C.; Zulueta, A.; Caretti, A.; Roda, G.; Paroni, R.; Dei Cas, M. An Update on Sphingolipidomics: Is Something Still Missing? Some Considerations on the Analysis of Complex Sphingolipids and Free-Sphingoid Bases in Plasma and Red Blood Cells. Metabolites 2022, 12, 450. [Google Scholar] [CrossRef]
- Hosmer, D.W., Jr.; Lemeshow, S. Applied Logistic Regression, 2nd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2000; pp. 1–397. [Google Scholar]
- Artusi, R.; Verderio, P.; Marubini, E. Bravais-Pearson and Spearman correlation coefficients: Meaning, test of hypothesis and confidence interval. Int. J. Biol. Markers 2002, 17, 148–151. [Google Scholar] [CrossRef]
- Hollander, M.; Wolfe, D.A.; Chicken, E. Nonparametric Statistical Methods, 3rd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2014; pp. 1–848. [Google Scholar]
- Joliffe, I. Principal Component Analysis; Springer: New York, NY, USA, 1986. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Frequency | Percent | |
|---|---|---|
| Sex | ||
| F | 107 | 41.31 |
| M | 152 | 58.69 |
| Patient’s Group | ||
| PT DH I/II | 36 | 13.9 |
| PT III | 28 | 10.81 |
| PT III–IV | 80 | 30.89 |
| control DH | 75 | 28.96 |
| control HD | 40 | 15.44 |
| Treatment type (PT III–IV only) | ||
| ICI (IPI/NIVO) | 40 | 50 |
| KI (BRAF/MEK) | 40 | 50 |
| Response class (PT III–IV only) | ||
| SR | 55 | 68.75 |
| LR | 25 | 31.25 |
| Age n; median (range) | 256; 50 (17–85) | |
| Triglycerides (mg/dL) n; median (range) | 258; 114 (40–528) | |
| Total cholesterol (mg/dL) n; median (range) | 258; 194 (69–336) | |
| LDL (mg/dL) n; median (range) | 258; 111.6 (5.8–242.8) | |
| HDL (mg/dL) n; median (range) | 258; 53 (12–227) | |
| DHCR24 (pg/mL) n; median (range) | 259; 24.78 (3.61–687.82) | |
| FASN (ng/mL) n; median (range) | 258; 0.74 (0.08–3.97) | |
| OR (95% CI) * | OR (95% CI) ** | |
|---|---|---|
| DHCR24 (pg/mL) | 0.74 (0.56; 0.96) | 0.84 (0.62; 1.13) |
| FASN (ng/mL) | 1.43 (1.09; 1.88) | 1.24 (0.92;1.67) |
| TG (mg/dL) | 1.26 (0.97; 1.64) | 1.04 (0.79; 1.37) |
| Total CHOL (mg/dL) | 0.97 (0.76; 1.24) | 0.77 (0.58; 1.03) |
| LDL (mg/dL) | 0.99 (0.78; 1.27) | 0.83 (0.62; 1.10) |
| HDL (mg/dL) | 0.77 (0.58; 1.03) | 0.78 (0.56; 1.07) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dei Cas, M.; Ciniselli, C.M.; Vergani, E.; Ciusani, E.; Aloisi, M.; Duroni, V.; Verderio, P.; Ghidoni, R.; Paroni, R.; Perego, P.; et al. Alterations in Plasma Lipid Profiles Associated with Melanoma and Therapy Resistance. Int. J. Mol. Sci. 2024, 25, 1558. https://doi.org/10.3390/ijms25031558
Dei Cas M, Ciniselli CM, Vergani E, Ciusani E, Aloisi M, Duroni V, Verderio P, Ghidoni R, Paroni R, Perego P, et al. Alterations in Plasma Lipid Profiles Associated with Melanoma and Therapy Resistance. International Journal of Molecular Sciences. 2024; 25(3):1558. https://doi.org/10.3390/ijms25031558
Chicago/Turabian StyleDei Cas, Michele, Chiara Maura Ciniselli, Elisabetta Vergani, Emilio Ciusani, Mariachiara Aloisi, Valeria Duroni, Paolo Verderio, Riccardo Ghidoni, Rita Paroni, Paola Perego, and et al. 2024. "Alterations in Plasma Lipid Profiles Associated with Melanoma and Therapy Resistance" International Journal of Molecular Sciences 25, no. 3: 1558. https://doi.org/10.3390/ijms25031558
APA StyleDei Cas, M., Ciniselli, C. M., Vergani, E., Ciusani, E., Aloisi, M., Duroni, V., Verderio, P., Ghidoni, R., Paroni, R., Perego, P., Beretta, G. L., Gatti, L., & Rodolfo, M. (2024). Alterations in Plasma Lipid Profiles Associated with Melanoma and Therapy Resistance. International Journal of Molecular Sciences, 25(3), 1558. https://doi.org/10.3390/ijms25031558