
High Stretch Associated with Mechanical Ventilation Promotes Piezo1-Mediated Migration of Airway Smooth Muscle Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
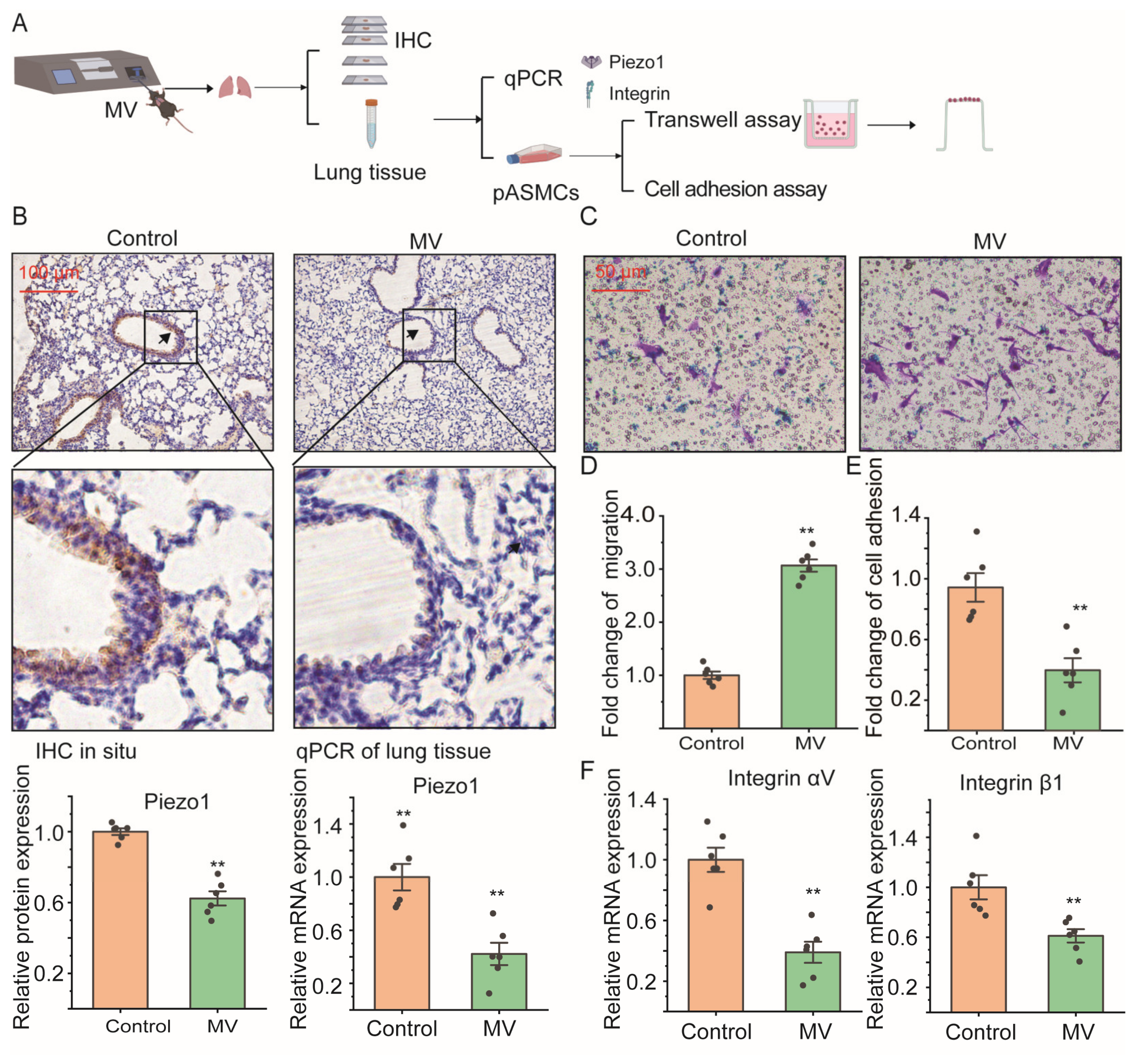
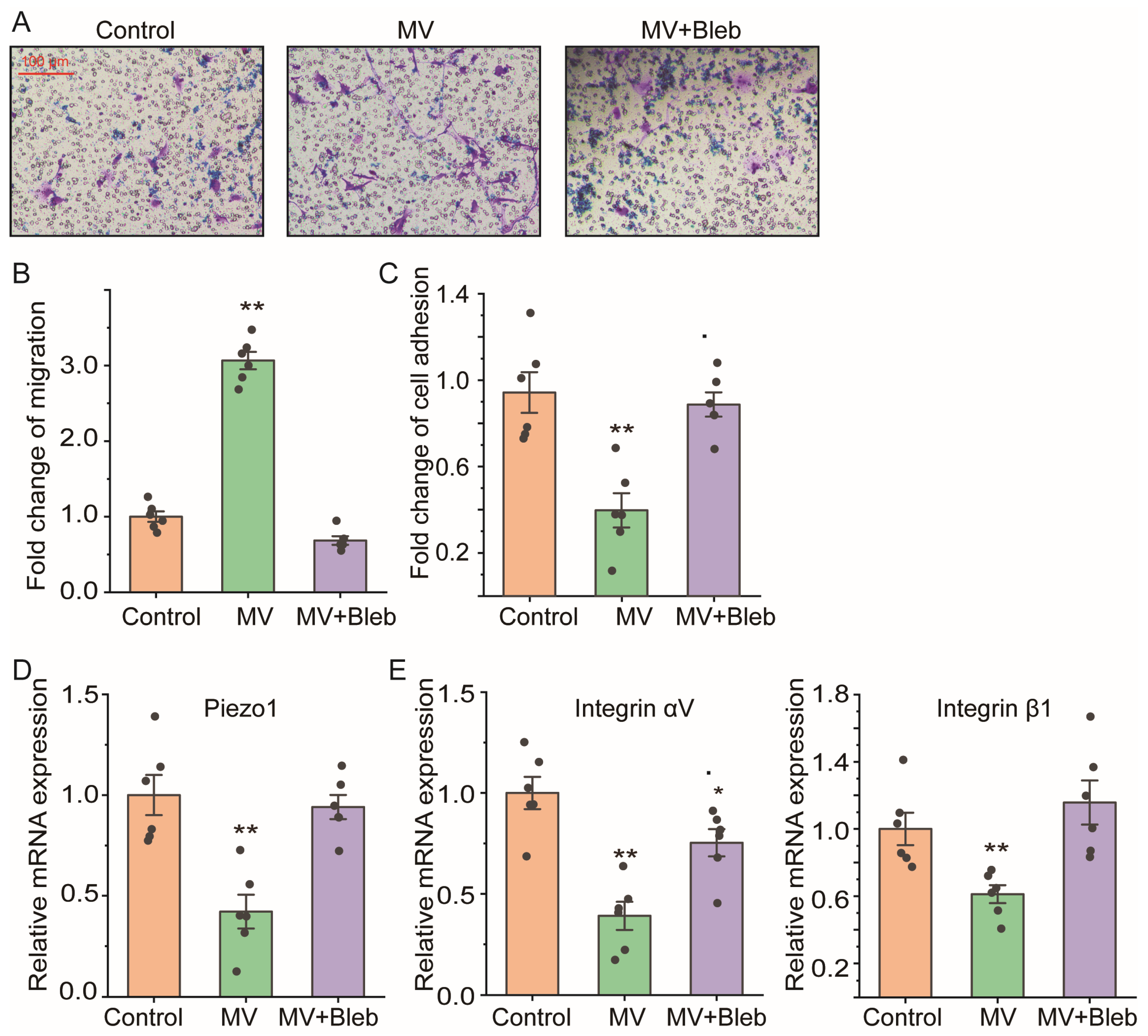
2.1. Mechanical Ventilation Decreased Piezo1 mRNA Expression in Lung Tissue In Vivo with Promoted Cell Migration of ASMCS
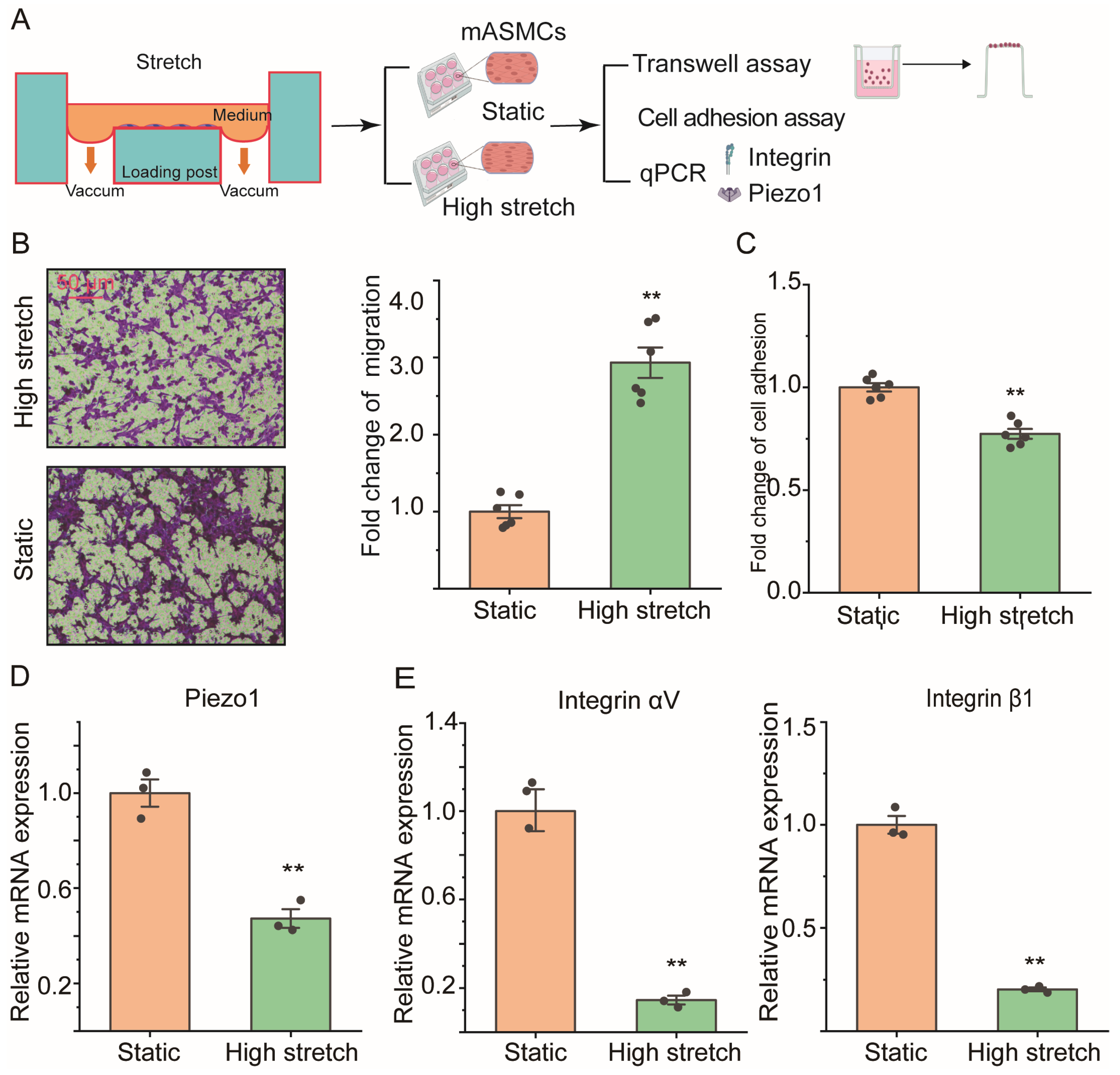
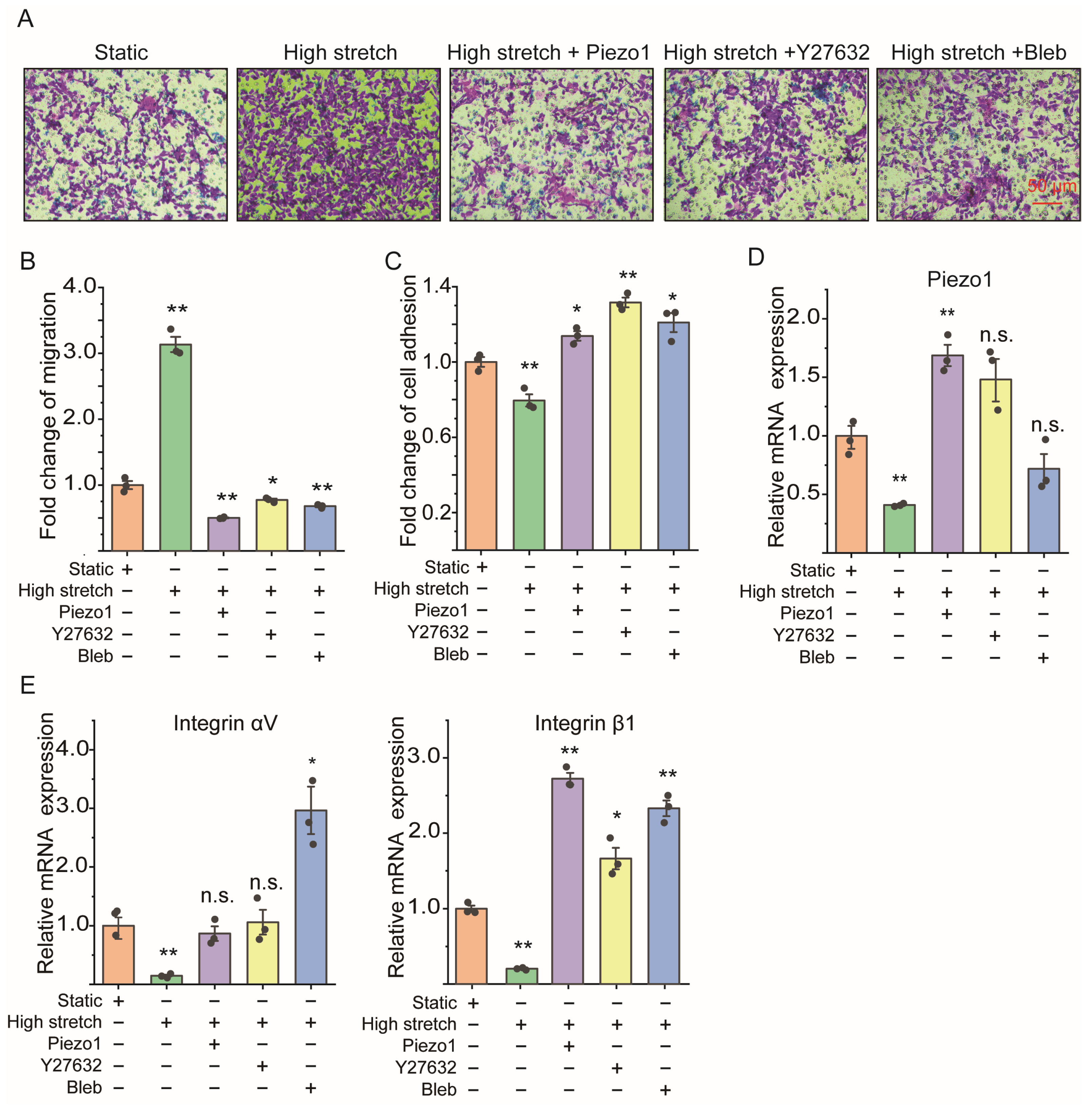
2.2. High Stretch Decreased Piezo1 mRNA Expression and Promoted Migration Transition of ASMCs In Vitro
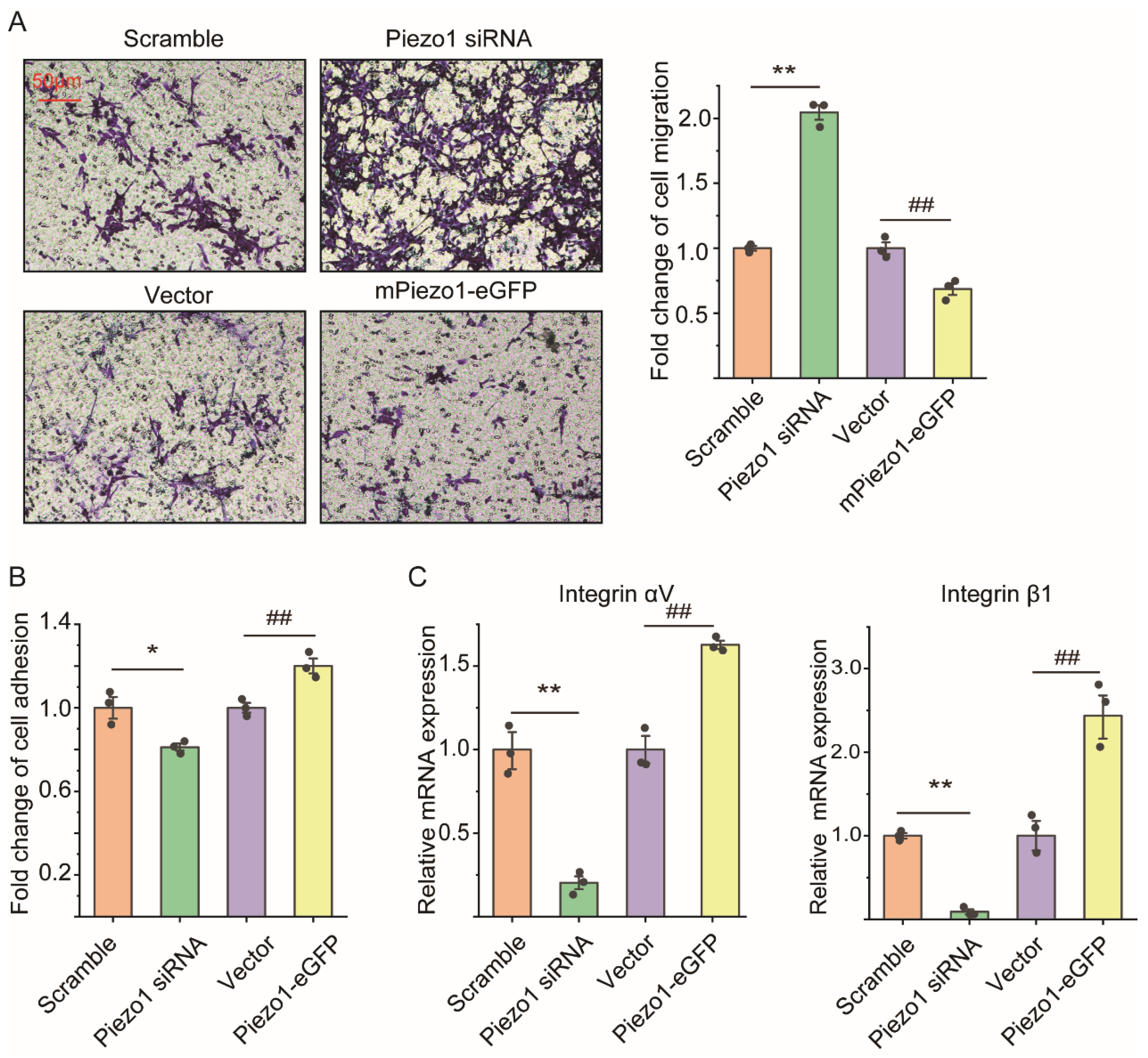
2.3. Loss of Piezo1 Promoted Cell Migration but Reduced Cell Adhesion and Integrin mRNA Expression of ASMCs In Vitro
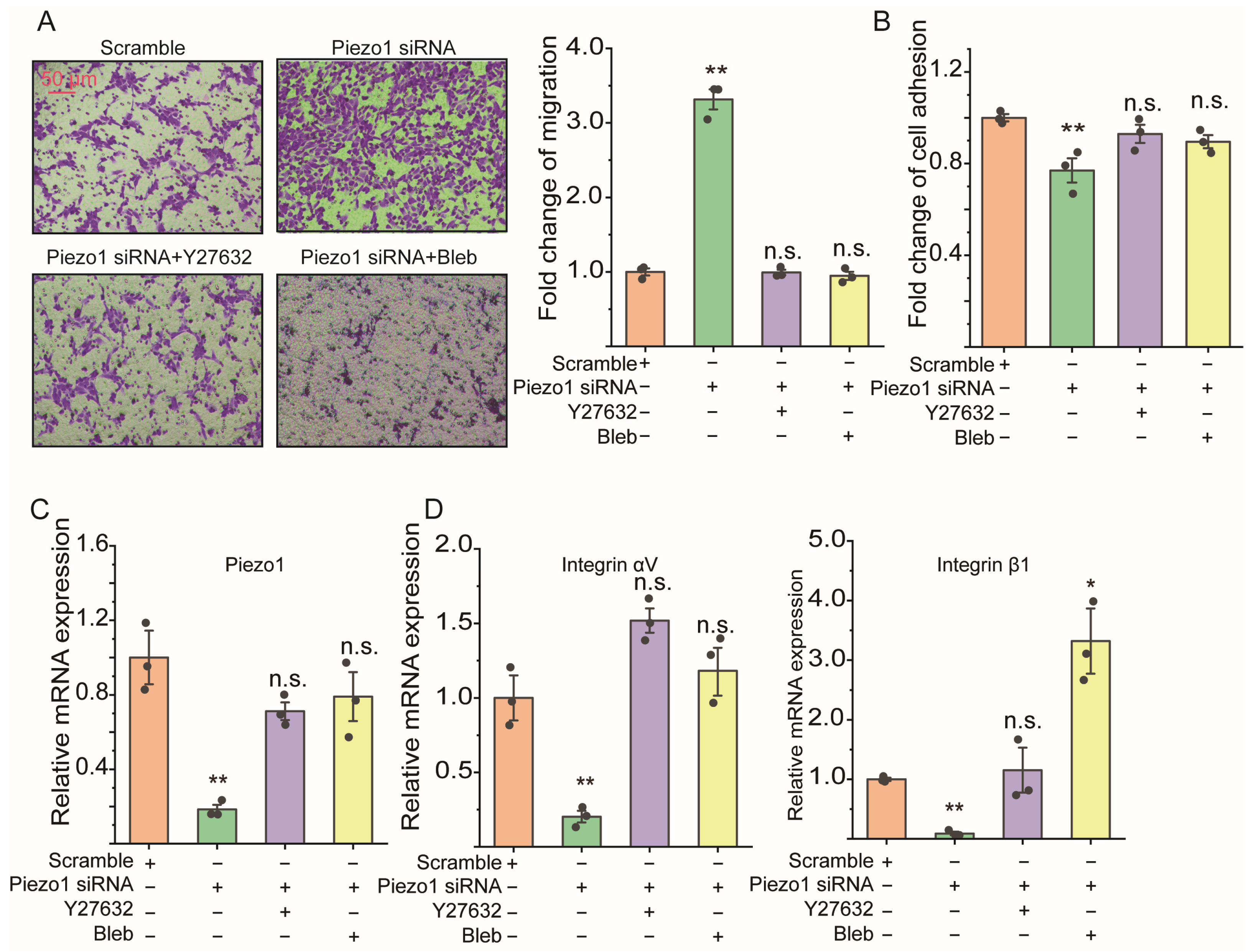
2.4. ROCK Signaling Inhibition Abolished the Cell Migration Due to Loss of Piezo1 mRNA of ASMCs
2.5. Piezo1 Knockin and ROCK Signaling Inhibition Both Reversed the Cell Migration Enhancement of ASMCs Due to High Stretch
2.6. ROCK Signaling Inhibition Abolished the Cell Migration Enhancement and Piezo1 Reduction of ASMCs from Mice after MV
3. Discussion
4. Materials and Methods
4.1. Animals with Mechanical Ventilation
4.2. Histological Evaluation of Piezo1 Expression in Mouse Lung
4.3. Isolation of Primary Mouse ASMCs from Mice after MV
4.4. Assessment of Piezo1 Expression and Distribution in pASMCs
4.5. Culture of Cell Line Mouse ASMCs with High Stretch
4.6. Assessment of Cell Migration of ASMCs
4.7. Assessment of Cell Adhesion of ASMCs
4.8. Assessment of mRNA Expression in ASMCs
4.9. Transfection of Piezo1 siRNA/Piezo1-eGFP into mASMCs
4.10. Assessment of Viability of ASMCs
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plataki, M.; Hubmayr, R.D. The physical basis of ventilator-induced lung injury. Expert Rev. Respir. Med. 2010, 4, 373–385. [Google Scholar] [CrossRef]
- Sinclair, S.E.; Molthen, R.C.; Haworth, S.T.; Dawson, C.A.; Waters, C.M. Airway strain during mechanical ventilation in an intact animal model. Am. J. Respir. Crit. Care Med. 2007, 176, 786–794. [Google Scholar] [CrossRef]
- Ibrahim, I.B.M.; Aghasafari, P.; Pidaparti, R.M. Transient mechanical response of lung airway tissue during mechanical ventilation. Bioengineering 2016, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Retamal, J.; Hurtado, D.; Villarroel, N.; Bruhn, A.; Bugedo, G.; Amato, M.B.P.; Costa, E.L.V.; Hedenstierna, G.; Larsson, A.; Borges, J.B. Does regional lung strain correlate with regional inflammation in acute respiratory distress syndrome during nonprotective ventilation? An experimental porcine study. Crit. Care Med. 2018, 46, e591–e599. [Google Scholar] [CrossRef] [PubMed]
- Villar, J.; Cabrera, N.E.; Valladares, F.; Casula, M.; Flores, C.; Blanch, L.; Quílez, M.E.; Santana-Rodríguez, N.; Kacmarek, R.; Slutsky, A.S. Activation of the wnt/β-catenin signaling pathway by mechanical ventilation is associated with ventilator-induced pulmonary fibrosis in healthy lungs. PLoS ONE 2011, 6, e23914. [Google Scholar] [CrossRef]
- D’Angelo, E.; Koutsoukou, A.; Valle, P.D.; Gentile, G.; Pecchiari, M. Cytokine release, small airway injury, and parenchymal damage during mechanical ventilation in normal open-chest rats. J. Appl. Physiol. 2008, 104, 41–49. [Google Scholar] [CrossRef]
- Xu, H.; Pan, G.; Wang, J. Repairing mechanisms for distal airway injuries and related targeted therapeutics for chronic lung diseases. Cell Transplant. 2023, 32, 09636897231196489. [Google Scholar] [CrossRef] [PubMed]
- Garfield, B.; Handslip, R.; Patel, B.V. Ventilator-associated lung injury. Encycl. Respir. Med. 2021, 361, 406–417. [Google Scholar]
- Cabrera-Benítez, N.E.; Parotto, M.; Post, M.; Han, B.; Spieth, P.M.; Cheng, W.E.; Valladares, F.; Villar, J.; Liu, M.; Sato, M.; et al. Mechanical stress induces lung fibrosis by epithelial-mesenchymal transition. Crit. Care Med. 2012, 40, 510–517. [Google Scholar] [CrossRef]
- Cullen, A.B.; Cooke, P.H.; Driska, S.P.; Wolfson, M.R.; Shaffer, T.H. The impact of mechanical ventilation on immature airway smooth muscle: Functional, structural, histological, and molecular correlates. Biol. Neonate 2006, 90, 17–27. [Google Scholar] [CrossRef]
- Hislop, A.A.; Haworth, S.G. Pulmonary vascular damage and the development of cor pulmonale following hyaline membrane disease. Pediatr. Pulmonol. 1990, 9, 152–161. [Google Scholar] [CrossRef]
- Albertine, K.H.; Jones, G.P.; Starcher, B.C.; Bohnsack, J.F.; Davis, P.L.; Cho, S.C.; Carlton, D.P.; Bland, R.D. Chronic lung injury in preterm lambs. Disordered respiratory tract development. Am. J. Respir. Crit. Care Med. 1999, 159, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Sward-Comunelli, S.L.; Mabry, S.M.; Truog, W.E.; Thibeault, D.W. Airway muscle in preterm infants: Changes during development. J. Pediatr. 1997, 130, 570–576. [Google Scholar] [CrossRef]
- Hasaneen, N.A.; Zucker, S.; Cao, J.; Chiarelli, C.; Panettieri, R.A.; Foda, H.D. Cyclic mechanical strain-induced proliferation and migration of human airway smooth muscle cells: Role of emmprin and mmps. FASEB J. 2005, 19, 1507–1509. [Google Scholar] [CrossRef]
- López-Martínez, C.; Huidobro, C.; Albaiceta, G.M.; López-Alonso, I. Mechanical stretch modulates cell migration in the lungs. Ann. Transl. Med. 2017, 6, 28. [Google Scholar] [CrossRef]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Bavi, N.; Richardson, J.; Heu, C.; Martinac, B.; Poole, K. Piezo1-mediated currents are modulated by substrate mechanics. ACS Nano 2019, 13, 13545–13559. [Google Scholar] [CrossRef]
- Ellefsen, K.L.; Holt, J.R.; Chang, A.C.; Nourse, J.L.; Arulmoli, J.; Mekhdjian, A.H.; Abuwarda, H.; Tombola, F.; Flanagan, L.A.; Dunn, A.R.; et al. Myosin-ii mediated traction forces evoke localized piezo1-dependent Ca2+ flickers. Commun. Biol. 2019, 2, 298. [Google Scholar] [CrossRef]
- Palecek, S.P.; Loftus, J.C.; Ginsberg, M.H.; Lauffenburger, D.A.; Horwitz, A.F. Integrin-ligand binding properties govern cell migration speed through cell-substratum adhesiveness. Nature 1997, 385, 537–540. [Google Scholar] [CrossRef]
- Wen, K.; Ni, K.; Guo, J.; Bu, B.; Liu, L.; Pan, Y.; Li, J.; Luo, M.; Deng, L. MircroRNA let-7a-5p in airway smooth muscle cells is most responsive to high stretch in association with cell mechanics modulation. Front. Physiol. 2022, 13, 830406. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Guo, J.; Ni, K.; Wen, K.; Qin, Y.; Gu, R.; Wang, C.; Liu, L.; Pan, Y.; Li, J.; et al. Mechanical ventilation-related high stretch mainly induces endoplasmic reticulum stress and thus mediates inflammation response in cultured human primary airway smooth muscle cells. Int. J. Mol. Sci. 2023, 24, 3811. [Google Scholar] [CrossRef]
- Carragher, N.O.; Walker, S.M.; Scott Carragher, L.A.; Harris, F.; Sawyer, T.K.; Brunton, V.G.; Ozanne, B.W.; Frame, M.C. Calpain 2 and src dependence distinguishes mesenchymal and amoeboid modes of tumour cell invasion: A link to integrin function. Oncogene 2006, 25, 5726–5740. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Le Berre, M.; Lautenschlaeger, F.; Maiuri, P.; Callan-Jones, A.; Heuzé, M.; Takaki, T.; Voituriez, R.; Piel, M. Confinement and low adhesion induce fast amoeboid migration of slow mesenchymal cells. Cell 2015, 160, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Gudipaty, S.A.; Lindblom, J.; Loftus, P.D.; Redd, M.J.; Edes, K.; Davey, C.F.; Krishnegowda, V.; Rosenblatt, J. Mechanical stretch triggers rapid epithelial cell division through piezo1. Nature 2017, 543, 118–121. [Google Scholar] [CrossRef]
- Solis, A.G.; Bielecki, P.; Steach, H.R.; Sharma, L.; Harman, C.C.D.; Yun, S.; de Zoete, M.R.; Warnock, J.N.; To, S.D.F.; York, A.G.; et al. Mechanosensation of cyclical force by piezo1 is essential for innate immunity. Nature 2019, 573, 69–74. [Google Scholar] [CrossRef]
- McHugh, B.J.; Buttery, R.; Lad, Y.; Banks, S.; Haslett, C.; Sethi, T. Integrin activation by fam38a uses a novel mechanism of r-ras targeting to the endoplasmic reticulum. J. Cell Sci. 2010, 123, 51–61. [Google Scholar] [CrossRef]
- Yao, M.; Tijore, A.; Cheng, D.; Li, J.V.; Hariharan, A.; Martinac, B.; Tran Van Nhieu, G.; Cox, C.D.; Sheetz, M. Force- and cell state-dependent recruitment of piezo1 drives focal adhesion dynamics and calcium entry. Sci. Adv. 2022, 8, eabo1461. [Google Scholar] [CrossRef]
- Cheng, D.; Wang, J.; Yao, M.; Cox, C.D. Joining forces: Crosstalk between mechanosensitive piezo1 ion channels and integrin-mediated focal adhesions. Biochem. Soc. Trans. 2023, 51, 1897–1906. [Google Scholar] [CrossRef]
- McHugh, B.J.; Murdoch, A.; Haslett, C.; Sethi, T. Loss of the integrin-activating transmembrane protein fam38a (piezo1) promotes a switch to a reduced integrin-dependent mode of cell migration. PLoS ONE 2012, 7, e40346. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.N.; Lu, Y.P.; Liu, J.J.; Huang, J.K.; Liu, Y.P.; Xiao, C.X.; Jazag, A.; Ren, J.L.; Guleng, B. Piezo1 is as a novel trefoil factor family 1 binding protein that promotes gastric cancer cell mobility in vitro. Dig. Dis. Sci. 2014, 59, 1428–1435. [Google Scholar] [CrossRef]
- Ridley, A.J. Rho gtpases and cell migration. J. Cell Sci. 2001, 114, 2713–2722. [Google Scholar] [CrossRef]
- Xie, N.; Xiao, C.; Shu, Q.; Cheng, B.; Wang, Z.; Xue, R.; Wen, Z.; Wang, J.; Shi, H.; Fan, D.; et al. Cell response to mechanical microenvironment cues via rho signaling: From mechanobiology to mechanomedicine. Acta Biomater. 2023, 159, 1–20. [Google Scholar] [CrossRef]
- Tu, P.C.; Pan, Y.L.; Liang, Z.Q.; Yang, G.L.; Wu, C.J.; Zeng, L.; Wang, L.N.; Sun, J.; Liu, M.M.; Yuan, Y.F.; et al. Mechanical stretch promotes macrophage polarization and inflammation via the rhoa-rock/nf-κb pathway. BioMed Res. Int. 2022, 2022, 6871269. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, L.; Huang, T.; Lu, D.; Song, Y.; Wang, L.; Gao, J. Mechanosensitive cation channel piezo1 contributes to ventilator-induced lung injury by activating rhoa/rock1 in rats. Respir. Res. 2021, 22, 250. [Google Scholar] [CrossRef]
- Luo, M.; Ni, K.; Gu, R.; Qin, Y.; Guo, J.; Che, B.; Pan, Y.; Li, J.; Liu, L.; Deng, L. Chemical activation of piezo1 alters biomechanical behaviors toward relaxation of cultured airway smooth muscle cells. Biol. Pharm. Bull. 2023, 46, 1–11. [Google Scholar] [CrossRef]
- Cabrera-Benitez, N.E.; Laffey, J.G.; Parotto, M.; Spieth, P.M.; Villar, J.; Zhang, H.; Slutsky, A.S. Mechanical ventilation-associated lung fibrosis in acute respiratory distress syndrome: A significant contributor to poor outcome. Anesthesiology 2014, 121, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Todd, N.W.; Luzina, I.G.; Atamas, S.P. Molecular and cellular mechanisms of pulmonary fibrosis. Fibrogenesis Tissue Repair 2012, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Zou, F.; Li, Y.; Zhang, S.; Zhang, J. Dp1 (prostaglandin d(2) receptor 1) activation protects against vascular remodeling and vascular smooth muscle cell transition to myofibroblasts in angiotensin ii-induced hypertension in mice. Hypertension 2022, 79, 1203–1215. [Google Scholar] [CrossRef]
- Takemoto, F.; Uchida-Fukuhara, Y.; Kamioka, H.; Okamura, H.; Ikegame, M. Mechanical stretching determines the orientation of osteoblast migration and cell division. Anat. Sci. Int. 2023, 98, 521–528. [Google Scholar] [CrossRef]
- Luo, M.; Cai, G.; Ho, K.K.Y.; Wen, K.; Tong, Z.; Deng, L.; Liu, A.P. Compression enhances invasive phenotype and matrix degradation of breast cancer cells via piezo1 activation. BMC Mol. Cell Biol. 2022, 23, 1. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, B.; Tumova, S.; Muraki, K.; Bruns, A.; Ludlow, M.J.; Sedo, A.; Hyman, A.J.; McKeown, L.; Young, R.S.; et al. Piezo1 integration of vascular architecture with physiological force. Nature 2014, 515, 279–282. [Google Scholar] [CrossRef]
- Pathak, M.M.; Nourse, J.L.; Tran, T.; Hwe, J.; Arulmoli, J.; Le, D.T.; Bernardis, E.; Flanagan, L.A.; Tombola, F. Stretch-activated ion channel piezo1 directs lineage choice in human neural stem cells. Proc. Natl. Acad. Sci. USA 2014, 111, 16148–16153. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Marshall, C.J. Differing modes of tumour cell invasion have distinct requirements for rho/rock signalling and extracellular proteolysis. Nat. Cell. Biol. 2003, 5, 711–719. [Google Scholar] [CrossRef]
- Wolf, K.; Mazo, I.; Leung, H.; Engelke, K.; von Andrian, U.H.; Deryugina, E.I.; Strongin, A.Y.; Brocker, E.B.; Friedl, P. Compensation mechanism in tumor cell migration: Mesenchymal-amoeboid transition after blocking of pericellular proteolysis. J. Cell. Biol. 2003, 160, 267–277. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, Y.; Lu, D.; Huang, T.; Yan, K.; Yang, W.; Gao, J. Mechanosensitive piezo1 channel activation promotes ventilator-induced lung injury via disruption of endothelial junctions in ARDS rats. Biochem. Biophys. Res. Commun. 2021, 556, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Hübner, R.H.; Gitter, W.; El Mokhtari, N.E.; Mathiak, M.; Both, M.; Bolte, H.; Freitag-Wolf, S.; Bewig, B. Standardized quantification of pulmonary fibrosis in histological samples. BioTechniques 2008, 44, 507–511, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Lifshitz, L.M.; Uy, K.F.; Ikebe, M.; Fogarty, K.E.; ZhuGe, R. The cellular and molecular basis of bitter tastant-induced bronchodilation. PLoS Biol. 2013, 11, e1001501. [Google Scholar] [CrossRef]
- Bagchi, A.; Vidal Melo, M.F. Follow the voxel-a new method for the analysis of regional strain in lung injury. Crit. Care Med. 2018, 46, 1033–1035. [Google Scholar] [CrossRef]
- Humphries, M.J. Cell adhesion assays. In Extracellular Matrix Protocols, 2nd ed.; Even-Ram, S., Artym, V., Eds.; Humana Press: Totowa, NJ, USA, 2009; pp. 203–210. [Google Scholar]
- Jin, Y.; Liu, L.; Yu, P.; Lin, F.; Shi, X.; Guo, J.; Che, B.; Duan, Y.; Li, J.; Pan, Y.; et al. Emergent differential organization of airway smooth muscle cells on concave and convex tubular surface. Front. Mol. Biosci. 2021, 8, 717771. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, M.; Gu, R.; Wang, C.; Guo, J.; Zhang, X.; Ni, K.; Liu, L.; Pan, Y.; Li, J.; Deng, L. High Stretch Associated with Mechanical Ventilation Promotes Piezo1-Mediated Migration of Airway Smooth Muscle Cells. Int. J. Mol. Sci. 2024, 25, 1748. https://doi.org/10.3390/ijms25031748
Luo M, Gu R, Wang C, Guo J, Zhang X, Ni K, Liu L, Pan Y, Li J, Deng L. High Stretch Associated with Mechanical Ventilation Promotes Piezo1-Mediated Migration of Airway Smooth Muscle Cells. International Journal of Molecular Sciences. 2024; 25(3):1748. https://doi.org/10.3390/ijms25031748
Chicago/Turabian StyleLuo, Mingzhi, Rong Gu, Chunhong Wang, Jia Guo, Xiangrong Zhang, Kai Ni, Lei Liu, Yan Pan, Jingjing Li, and Linhong Deng. 2024. "High Stretch Associated with Mechanical Ventilation Promotes Piezo1-Mediated Migration of Airway Smooth Muscle Cells" International Journal of Molecular Sciences 25, no. 3: 1748. https://doi.org/10.3390/ijms25031748
APA StyleLuo, M., Gu, R., Wang, C., Guo, J., Zhang, X., Ni, K., Liu, L., Pan, Y., Li, J., & Deng, L. (2024). High Stretch Associated with Mechanical Ventilation Promotes Piezo1-Mediated Migration of Airway Smooth Muscle Cells. International Journal of Molecular Sciences, 25(3), 1748. https://doi.org/10.3390/ijms25031748