
Activity of Potassium Channels in CD8+ T Lymphocytes: Diagnostic and Prognostic Biomarker in Ovarian Cancer?

 ,
,  ,
,  ,
,  ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
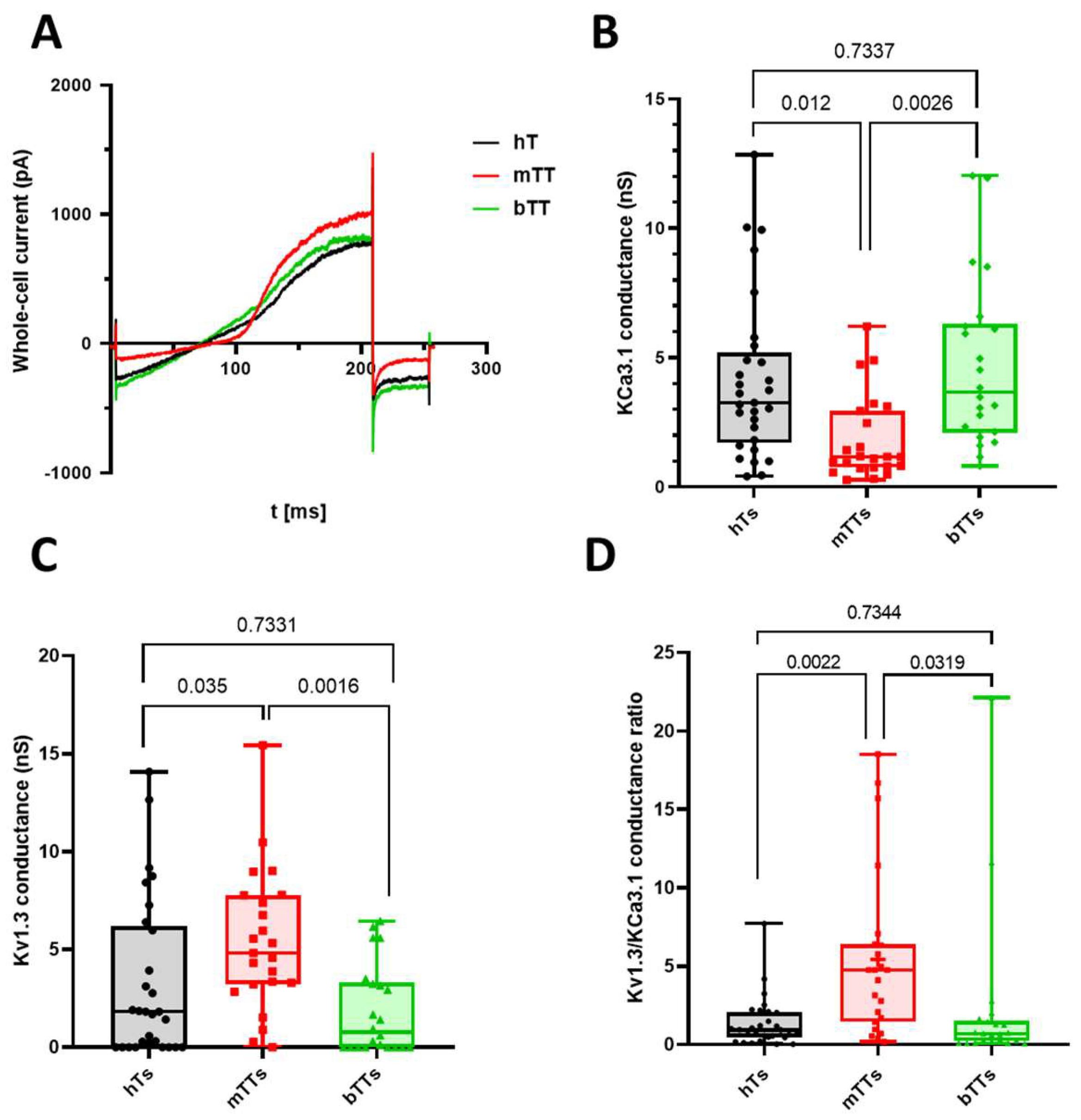
2.1. Malignant Tumor CD8+ Cells Have Low KCa3.1 and High Kv1.3 Conductance
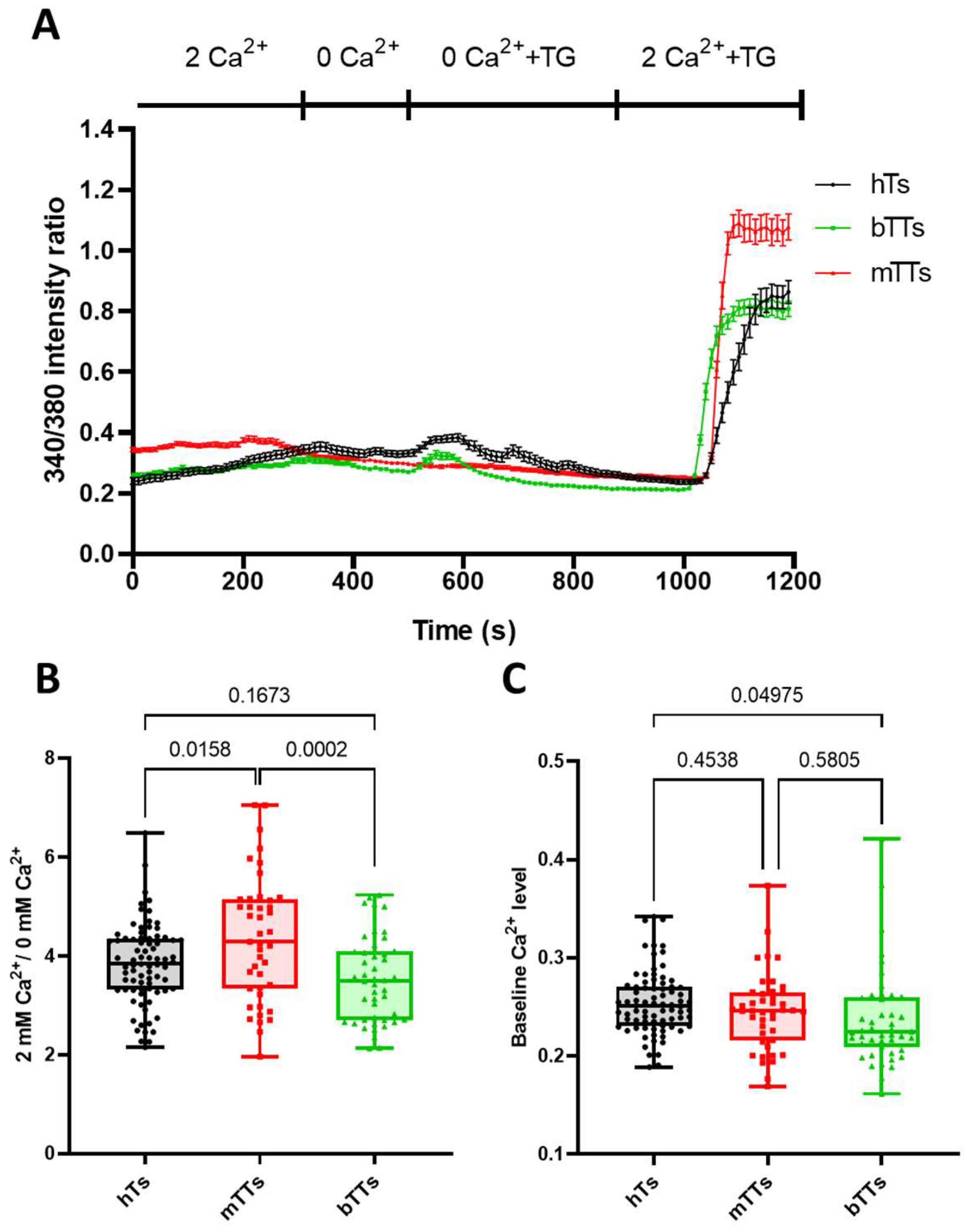
2.2. mTTs Show Higher CRAC-Dependent Ca2+ Response
3. Discussion
4. Materials and Methods
4.1. Human Subject
4.2. PBMC Isolation and Activation
4.3. Monoclonal Antibody Adhesion of CD8+ Cells
4.4. Electrophysiology
4.5. Intracellular Ca2+ Measurements
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feske, S.; Wulff, H.; Skolnik, E.Y. Ion channels in innate and adaptive immunity. Annu. Rev. Immunol. 2015, 33, 291–353. [Google Scholar] [CrossRef] [PubMed]
- Joglekar, A.V.; Li, G. T cell antigen discovery. Nat. Methods 2021, 18, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Svensson, M.; Stockinger, B.; Wick, M.J. Bone marrow-derived dendritic cells can process bacteria for MHC-I and MHC-II presentation to T cells. J. Immunol. 1997, 158, 4229–4236. [Google Scholar] [CrossRef] [PubMed]
- Conforti, L. The ion channel network in T lymphocytes, a target for immunotherapy. Clin. Immunol. 2012, 142, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Tóth, A.; Szilágyi, O.; Krasznai, Z.; Panyi, G.; Hajdú, P. Functional consequences of Kv1.3 ion channel rearrangement into the immunological synapse. Immunol. Lett. 2009, 125, 15–21. [Google Scholar] [CrossRef]
- Prakriya, M.; Lewis, R.S. CRAC channels: Activation, permeation, and the search for a molecular identity. Cell Calcium 2003, 33, 311–321. [Google Scholar] [CrossRef]
- Zweifach, A.; Lewis, R.S. Mitogen-regulated Ca2+ current of T lymphocytes is activated by depletion of intracellular Ca2+ stores. Proc. Natl. Acad. Sci. USA 1993, 90, 6295–6299. [Google Scholar] [CrossRef]
- Hu, L.; Wang, T.; Gocke, A.R.; Nath, A.; Zhang, H.; Margolick, J.B.; Whartenby, K.A.; Calabresi, P.A. Blockade of Kv1.3 potassium channels inhibits differentiation and granzyme B secretion of human CD8+ T effector memory lymphocytes. PLoS ONE 2013, 8, e54267. [Google Scholar] [CrossRef]
- Zhou, X.; Friedmann, K.S.; Lyrmann, H.; Zhou, Y.; Schoppmeyer, R.; Knörck, A.; Mang, S.; Hoxha, C.; Angenendt, A.; Backes, C.S.; et al. A calcium optimum for cytotoxic T lymphocyte and natural killer cell cytotoxicity. J. Physiol. 2018, 596, 2681–2698. [Google Scholar] [CrossRef]
- Eil, R.; Vodnala, S.K.; Clever, D.; Klebanoff, C.A.; Sukumar, M.; Pan, J.H.; Palmer, D.C.; Gros, A.; Yamamoto, T.N.; Patel, S.J.; et al. Ionic immune suppression within the tumour microenvironment limits T cell effector function. Nature 2016, 537, 539–543. [Google Scholar] [CrossRef]
- Olivas-Aguirre, M.; Cruz-Aguilar, L.H.; Pottosin, I.; Dobrovinskaya, O. Reduction of Ca2+ Entry by a Specific Block of KCa3.1 Channels Optimizes Cytotoxic Activity of NK Cells against T-ALL Jurkat Cells. Cells 2023, 12, 2065. [Google Scholar] [CrossRef] [PubMed]
- Koshy, S.; Wu, D.; Hu, X.; Tajhya, R.B.; Huq, R.; Khan, F.S.; Pennington, M.W.; Wulff, H.; Yotnda, P.; Beeton, C. Blocking KCa3.1 channels increases tumor cell killing by a subpopulation of human natural killer lymphocytes. PLoS ONE 2013, 8, e76740. [Google Scholar] [CrossRef] [PubMed]
- Chimote, A.A.; Balajthy, A.; Arnold, M.J.; Newton, H.S.; Hajdu, P.; Qualtieri, J.; Wise-Draper, T.; Conforti, L. A defect in KCa3.1 channel activity limits the ability of CD8+ T cells from cancer patients to infiltrate an adenosine-rich microenvironment. Sci. Signal 2018, 11, eaaq1616. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.K.; Wong, B.H.S.; Poh, Z.S.; Udayakumar, A.; Verma, R.; Goh, R.K.J.; Duggan, S.P.; Shelat, V.G.; Chandy, K.G.; Grigoropoulos, N.F. Obstacles for T-lymphocytes in the tumour microenvironment: Therapeutic challenges, advances and opportunities beyond immune checkpoint. EBioMedicine 2022, 83, 104216. [Google Scholar] [CrossRef] [PubMed]
- Thommen, D.S.; Schumacher, T.N. T Cell Dysfunction in Cancer. Cancer Cell 2018, 33, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Chirra, M.; Newton, H.S.; Gawali, V.S.; Wise-Draper, T.M.; Chimote, A.A.; Conforti, L. How the Potassium Channel Response of T Lymphocytes to the Tumor Microenvironment Shapes Antitumor Immunity. Cancers 2022, 14, 3564. [Google Scholar] [CrossRef] [PubMed]
- Chimote, A.A.; Hajdu, P.; Kottyan, L.C.; Harley, J.B.; Yun, Y.; Conforti, L. Nanovesicle-targeted Kv1.3 knockdown in memory T cells suppresses CD40L expression and memory phenotype. J. Autoimmun. 2016, 69, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Voros, O.; Panyi, G.; Hajdu, P. Immune Synapse Residency of Orai1 Alters Ca2+ Response of T Cells. Int. J. Mol. Sci. 2021, 22, 11514. [Google Scholar] [CrossRef]
- Chimote, A.A.; Hajdu, P.; Kucher, V.; Boiko, N.; Kuras, Z.; Szilagyi, O.; Yun, Y.-H.; Conforti, L. Selective inhibition of KCa3.1 channels mediates adenosine regulation of the motility of human T cells. J. Immunol. 2013, 191, 6273–6280. [Google Scholar] [CrossRef]
- Chimote, A.A.; Hajdu, P.; Sfyris, A.M.; Gleich, B.N.; Wise-Draper, T.; Casper, K.A.; Conforti, L. Kv1.3 Channels Mark Functionally Competent CD8+ Tumor-Infiltrating Lymphocytes in Head and Neck Cancer. Cancer Res. 2017, 77, 53–61. [Google Scholar] [CrossRef]
- Wang, T.; Lee, M.-H.; Choi, E.; Pardo-Villamizar, C.A.; Bin Lee, S.; Yang, I.H.; Calabresi, P.A.; Nath, A. Granzyme B-induced neurotoxicity is mediated via activation of PAR-1 receptor and Kv1.3 channel. PLoS ONE 2012, 7, e43950. [Google Scholar] [CrossRef]
- Wulff, H.; Calabresi, P.A.; Allie, R.; Yun, S.; Pennington, M.; Beeton, C.; Chandy, K.G. The voltage-gated Kv1.3 K+ channel in effector memory T cells as new target for MS. J. Clin. Investig. 2003, 111, 1703–1713. [Google Scholar] [CrossRef]
- Kershaw, M.H.; Westwood, J.A.; Parker, L.L.; Wang, G.; Eshhar, Z.; Mavroukakis, S.A.; White, D.E.; Wunderlich, J.R.; Canevari, S.; Rogers-Freezer, L.; et al. A Phase I study on adoptive immunotherapy using gene-modified T cells for ovarian cancer. Clin. Cancer Res. 2006, 12 Pt 1, 6106–6115. [Google Scholar] [CrossRef]
- Chimote, A.A.; Gawali, V.S.; Newton, H.S.; Wise-Draper, T.M.; Conforti, L. A Compartmentalized Reduction in Membrane-Proximal Calmodulin Reduces the Immune Surveillance Capabilities of CD8+ T Cells in Head and Neck Cancer. Front. Pharmacol. 2020, 11, 143. [Google Scholar] [CrossRef]
- Newton, H.S.; Chimote, A.A.; Arnold, M.J.; Wise-Draper, T.M.; Conforti, L. Targeted knockdown of the adenosine A2A receptor by lipid NPs rescues the chemotaxis of head and neck cancer memory T cells. Mol. Ther. Methods Clin. Dev. 2021, 21, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, R.; Chandy, K.G.; Grissmer, S.; Gutman, G.A.; Kaczmarek, L.K.; Wei, A.D.; Wulff, H. Calcium- and sodium-activated potassium channels (KCa, KNa) in GtoPdb v.2023.1. IUPHAR/BPS Guide Pharmacol. CITE 2023, 2023. [Google Scholar] [CrossRef]
- Matteson, D.R.; Deutsch, C. K channels in T lymphocytes: A patch clamp study using monoclonal antibody adhesion. Nature 1984, 307, 468–471. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jusztus, V.; Medyouni, G.; Bagosi, A.; Lampé, R.; Panyi, G.; Matolay, O.; Maka, E.; Krasznai, Z.T.; Vörös, O.; Hajdu, P. Activity of Potassium Channels in CD8+ T Lymphocytes: Diagnostic and Prognostic Biomarker in Ovarian Cancer? Int. J. Mol. Sci. 2024, 25, 1949. https://doi.org/10.3390/ijms25041949
Jusztus V, Medyouni G, Bagosi A, Lampé R, Panyi G, Matolay O, Maka E, Krasznai ZT, Vörös O, Hajdu P. Activity of Potassium Channels in CD8+ T Lymphocytes: Diagnostic and Prognostic Biomarker in Ovarian Cancer? International Journal of Molecular Sciences. 2024; 25(4):1949. https://doi.org/10.3390/ijms25041949
Chicago/Turabian StyleJusztus, Vivien, Ghofrane Medyouni, Adrienn Bagosi, Rudolf Lampé, György Panyi, Orsolya Matolay, Eszter Maka, Zoárd Tibor Krasznai, Orsolya Vörös, and Péter Hajdu. 2024. "Activity of Potassium Channels in CD8+ T Lymphocytes: Diagnostic and Prognostic Biomarker in Ovarian Cancer?" International Journal of Molecular Sciences 25, no. 4: 1949. https://doi.org/10.3390/ijms25041949
APA StyleJusztus, V., Medyouni, G., Bagosi, A., Lampé, R., Panyi, G., Matolay, O., Maka, E., Krasznai, Z. T., Vörös, O., & Hajdu, P. (2024). Activity of Potassium Channels in CD8+ T Lymphocytes: Diagnostic and Prognostic Biomarker in Ovarian Cancer? International Journal of Molecular Sciences, 25(4), 1949. https://doi.org/10.3390/ijms25041949