1. Introduction
Chlamydia trachomatis is the most common sexually transmitted bacterial pathogen. It causes serious health problems in humans and can cause serious complications, such as pelvic inflammatory disease, ectopic pregnancy and infertility in women [
1]. The medical and socio-economic significance of the search for new-generation drugs using target-specific technologies is due to the lack of effective agents that can help treat chronic bacterial infections and the rapid development of pathogenic resistance to the antibacterial drugs used to treat acute infectious diseases [
2,
3,
4]. In the case of antibacterial drugs, this technology involves selecting as targets the proteins responsible for the microorganism’s display of pathogenic properties, then searching for specific inhibitors using computer software, organic synthesis methods and experimental studies and validating the predicted biological activity on model systems of the infectious process. The secretion of pathogenic factors (proteins responsible for the manifestation of pathogenic properties in bacteria) into macroorganisms’ cells is the key mechanism underlying the development of the infectious process. The type 3 secretion system (T3SS) is the predominant virulence factor of chlamydiae. It is essential for cell invasion and is active at all life stages [
5,
6]. Some T3SS proteins are on the surface and can be targeted by neutralizing antibodies. A T-cell response to T3SS antigens has recently been shown to be associated with protection against
C. trachomatis infection in highly exposed women [
7], and T3SS components have recently attracted attention as vaccine candidates against other pathogenic bacteria [
8,
9,
10,
11]. The
C. trachomatis T3SS filament protein CdsF and its orthologs in other bacteria, such as TC_0037 protein of
Chlamydia muridarum, form the needles of injectisomes and are believed to facilitate the insertion of translocators into the host cell membrane [
5,
6,
12]. CdsF (TC_0037) is highly conserved, showing 95% sequence identity in the genus
Chlamydia. It is abundant on bacterial surfaces, raising the possibility that a CdsF-based vaccine may induce a wide range of protection against all medically significant strains. It was shown that the T3SS needle protein TC_0037 induced specific humoral and T-cell responses, decreased the
Chlamydia loads in the genital tract and abrogated the pathology of the upper genital organs. It could be a good candidate for inclusion in a
Chlamydia vaccine [
13]. In the study just mentioned, the authors immunized mice with a replication-defective adenoviral vector expressing the recombinant target antigen TC_0037. Thus, TC_0037, based on the published data, is a very promising target for the immunotherapeutic suppression of chlamydial infection. The presented work was made possible due to the fact that at the previous stage of research, we were able to obtain highly specific single-domain antibodies (nanobodies) that recognize precisely this promising therapeutic target, TC_0037.
The purpose of our study was to demonstrate the fundamental possibility of using another approach of transmitting information about the target antigen to the host body for targeted immune stimulation. The approach is based on the use of stable molecular structures that mimic the specific pathogenic epitope that is the preferred target of a therapeutic antibody. One of the ways to create such biomimetics is based on the production of anti-idiotypic antibodies (aiAbs). An “idiotype” is an “individual antigenic determinant” for an isolated antibody or for a specific type of T- or B-lymphocyte receptor [
14]. According to the modern terminology, originally proposed in 1960 by N. Yerne [
15], epitopes are determinants of antigens that are recognized by a specific antibody-binding site, the paratope. An idiotype is a structure formed by the association of the variable regions of the heavy and light chains of an antibody. Each idiotype is capable of stimulating the production of complementary “anti-idiotypic” antibodies, the antigen-recognizing site of which may be conformationally similar to the original antigenic determinant (a specific epitope of the antigen). Thus, the active centers of the antibodies formed in response to antigen introduction, in turn, act as antigens. According to the anti-idiotypic principle, the immune system is potentially capable of reproducing or at least modifying the action of any biologically active endogenous or exogenous immunogenic agent, as well as probably regulating a wide range of cellular functions in the body. aiAbs are used when it is difficult to prepare and apply an antigen or when the antigen is toxic, unstable or causes an insufficiently focused immune response. Finally, the native structure of the antigen may be poorly understood, and antibodies (idiotypes) that specifically recognize it in vivo have already been obtained. Due to the potential ability of carefully selected aiAbs to imitate a certain epitope (display the “internal image”) of a given antigen, thus compete with the antigen in binding to the same structures in living systems and competitively change the biological activity of the antigen, aiAbs have become a promising tool in the search for new ways to treat autoimmune diseases, cancer and many other human diseases [
16,
17]. It was demonstrated that aiAbs may very likely have therapeutic potential for use as substitutes (mimetics) of antigens (both of protein, non-protein or mixed nature) in vaccines for the treatment of bacterial, viral and cancer diseases [
18,
19,
20].
In our opinion, single-domain antibodies (nanobodies) are a very promising tool for obtaining such molecules, anti-idiotypic nanobodies (ai-Nbs), structurally mimicking a given epitope [
21,
22,
23,
24,
25]. It was shown that ai-Nbs can be effective mimetics of natural enzymes, an excellent tool for studying and utilizing molecular mimicry techniques. ai-Nbs are easier to generate and better suited to mimicking small ligands and to the subsequent design of peptide mimetics for drug development [
26,
27,
28,
29]. In camelids, an unusual combination of two types of antibodies/receptors of B-cells with markedly different properties was found. The repertoires of possible paratopes of the antigen-binding structures of heavy-chain-only antibodies (HCAbs) and classical antibodies, apparently, may differ markedly. Since these two types of antibodies co-exist in the same organism, it can be assumed that they do not compete but mutually complement each other. Indeed, it has been repeatedly observed that both types can arise in parallel (mutually exclusive or in different ratios) against different epitopes/antigens of multicomponent antigenic material used in the immunization of the same animals. Some authors have expressed doubts about the possibility of adequate antigenic mimicry of the usually convex epitopes of antigens with the help of classical (albeit anti-idiotypic) antibodies, the active centers of which usually have a concave structure [
25]. However, such a protruding structure is quite realistic in the case of non-canonical HCAbs (and their derived nanobodies), in which the more elongated third hypervariable region (CDR3) forms exactly protruding structures [
24,
30,
31,
32]. Thus, the camelid arsenal has the potential to create “more complementary” interactions within the idiotype–anti-idiotype network. In our opinion, it is the structural features of single variable domains (VHH) that make nanobodies a potentially unique tool for creating an anti-idiotype that mimics a specific epitope of the target antigen in terms of specific binding.
In this study, we applied nanobody generation technology to obtain ai-Nbs mimicking the structure of a given model epitope of the pathogen (the epitope of the Chlamydial Type III Secretion System Needle Protein). In a mouse model, we show that the obtained ai-Nbs are able to induce a narrowly specific humoral immune response of the host leading to the generation of intrinsic anti-Chlamydia antibodies, potentially therapeutic, specifically recognizing a given antigenic epitope of Chlamydia. The immune sera derived from mice immunized with ai-Nbs are able to partly suppress chlamydial infection in a neutralization assay in vitro. We hypothesize that the use of ai-Nbs, which mimic and present to the host immune system exactly the desired region of the antigen, creates a fundamentally new universal approach to generating molecular structures as a part of a specific vaccine for the targeted induction of immune response, especially useful in cases where it is difficult to prepare an antigen preserving the desired epitope in its native conformation.
3. Discussion
The main goal of this study was the application of nanobody generation technology to the methodological development and demonstration, as a proof of principle, of an efficient way to create structurally stable polypeptide biomimetics that conformationally reproduce a given epitope of a target pathogen, able to present to the host immune system exactly the desired region of the antigen for the targeted induction of immune response. In our opinion, we managed to achieve this goal. The method of producing the nanobodies described in this article was used for the first time, as far as we know. The main proposed stages for obtaining anti-idiotypic nanobodies are as follows.
(1) With the described method, it is fundamentally important to identify a promising therapeutic target at the beginning of the work and to be able to obtain primary single-domain antibodies that highly specifically recognize it and have therapeutic potential. Most likely, this will be an already known target for which monoclonal antibodies (mAbs) have been obtained and the immunotherapeutic effect of using these antibodies has been demonstrated.
(2) The generation of primary nanobodies against a selected target. Here, we have a classic case of obtaining new nanobodies using a well-established technological platform. From the panel of obtained nanobodies, it is necessary to functionally select those variants that effectively bind to the selected target in vivo and have the desired effect (for example, they can suppress the infectivity of Chlamydia in vitro).
(3) The functionally selected primary nanobody must be formatted in such a way as to most effectively use it as an antigen for the subsequent immunization of a member of the
Camelidae family (using special formatting, carrier proteins and effective adjuvants). From our experience, a parallel trimerized version of the nanobody reproducibly gave a good result in inducing the formation of ai-Nb precursors when immunizing a camel. The use of special formatting to obtain trimerizing nanobodies is a development of our laboratory [
34].
Among some other published examples of the use of anti-idiotypic nanobodies, which are still rare but show great potential, ai-Nbs are usually produced against the idiotypes of classical antibodies [
26,
27,
28,
29,
37,
38,
39]. We consider this an alternative possible route; however, in our experience, the efficiency of the generation and selection of ai-Nbs that mimic the epitope of the original target increases significantly if nanobodies are used as the primary idiotypes instead of mAbs.
(4) The generation of ai-Nbs using a well-established technological nanobody platform. The main desirable criteria for the selection of ai-Nb clones at this stage are: (a) the high specificity and affinity of the binding of selected ai-Nb variants to a unique idiotype (paratope) and not to conservative regions of the primary nanobody and (b) the ai-Nbs should effectively compete with the original target antigen (in this case, TC_0037) in binding to the primary nanobody.
(5) Selected ai-Nbs must be formatted in such a way as to most effectively use them as antigens for the subsequent immunization of a model organism (mouse). For the initial demonstration experiments (in this article), we also used a trimerized version of the ai-Nbs, but this is only one of the possible formats for preparing antigenic material.
(6) Immunization of a model animal (mouse) with antigenic material based on a formatted ai-Nb (or its multi-module derivatives) and functional analysis of the induced host antibodies. The goal is to select an ai-Nb that is able to induce a narrowly specific humoral immune response in the host leading to the generation of intrinsic antibodies against the initially selected therapeutic target.
With the described method, it seems fundamentally important to identify a promising therapeutic target at the beginning of the work and to be able to obtain primary single-domain antibodies that highly specifically recognize it and have therapeutic potential. At the beginning of the presented work, we were able to obtain the desired highly specific single-domain antibody (nanobody) that recognized precisely such a promising therapeutic target, Chlamydial Type III Secretion System Needle Protein TC_0037.
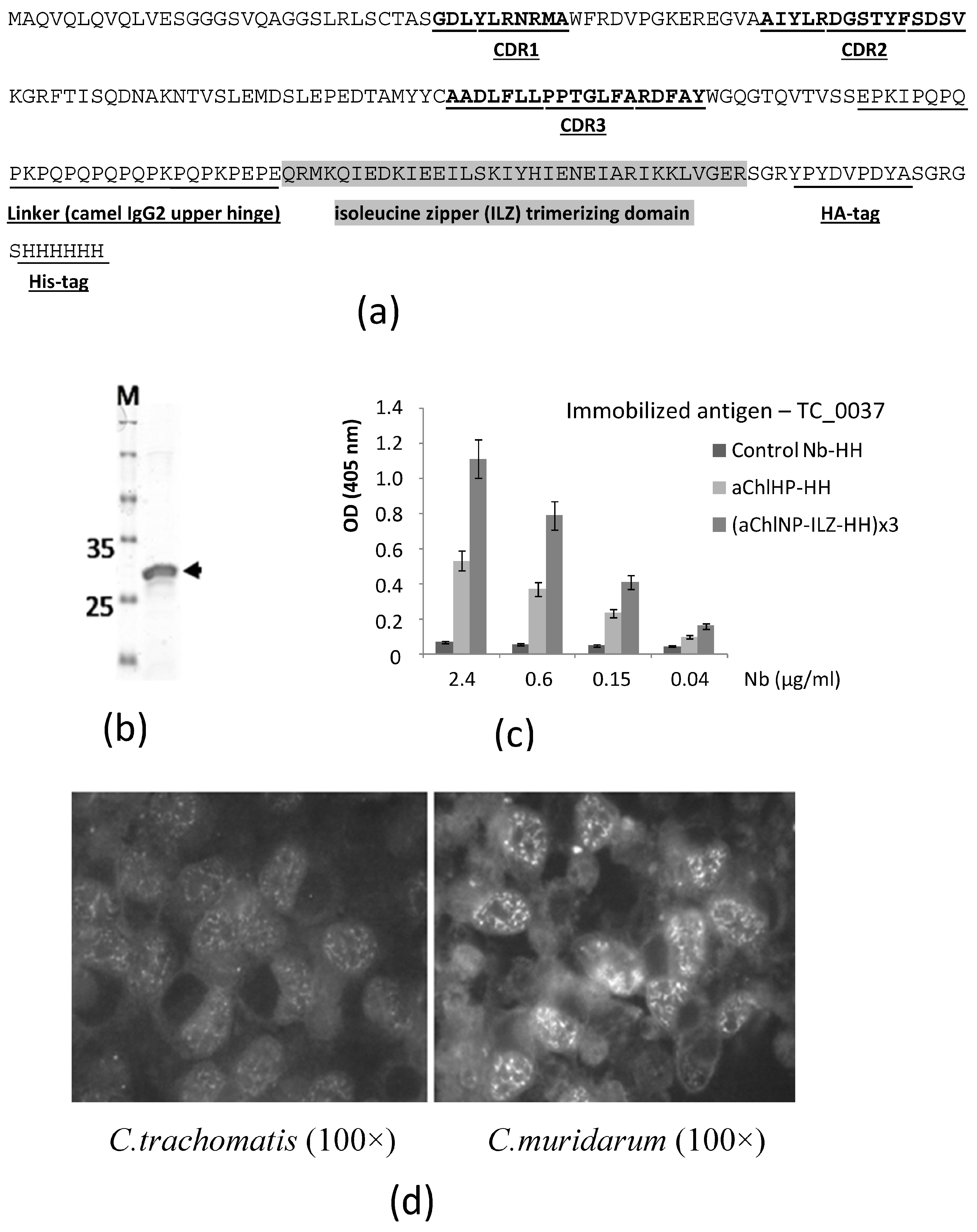
It can be stated that the following sequence of events was triggered: antigen X → nanobody aX recognizing X → anti-idiotypic nanobody (anti-aX) recognizing nanobody aX → induction of the formation of mouse antibodies recognizing antigen X, where the arrow denotes the immunization step. First, a camel (or another member of the Camelidae family) is immunized with biological material enriched with the target antigen in a conformation as close as possible to the native one. Using the RNA of the peripheral blood mononuclear cells of the immunized camel, molecular cloning of the entire repertoire of variable domains (VHH, nanobodies) of the special antibodies (HCAbs) is carried out, and the most promising clone variants are selected using the phage display method. After functional testing, the best variants of the primary nanobodies (aChlNP) are selected for the second cycle of immunization, also using a representative of the Camelidae family. From our previous experience, we hypothesized that the primary nanobody in a trimerizable format would be a more effective inducer of the desired immune response. To this end, during formatting, additional amino acid sequences were added to the C-terminus of the primary sequence of the nanobody (
Figure 1a), including the trimerizing isoleucine zipper domain, as previously described [
34]. The VHH-cDNA library generated after the second cycle of immunization was then used for the selection of clones encoding for ai-Nbs (aiChlNP) that specifically bind to the antigen-recognizing site (idiotype) of the aChlNP nanobody but do not bind to the constant nanobody sites. Four obtained ai-Nbs were formatted to make trimerizing Nb derivatives and then were functionally tested in a mouse model. We have shown that the obtained ai-Nbs are able to induce a narrowly specific humoral immune response in the host leading to the generation of intrinsic anti-
Chlamydia antibodies, potentially therapeutic, specifically recognizing a given antigenic epitope of
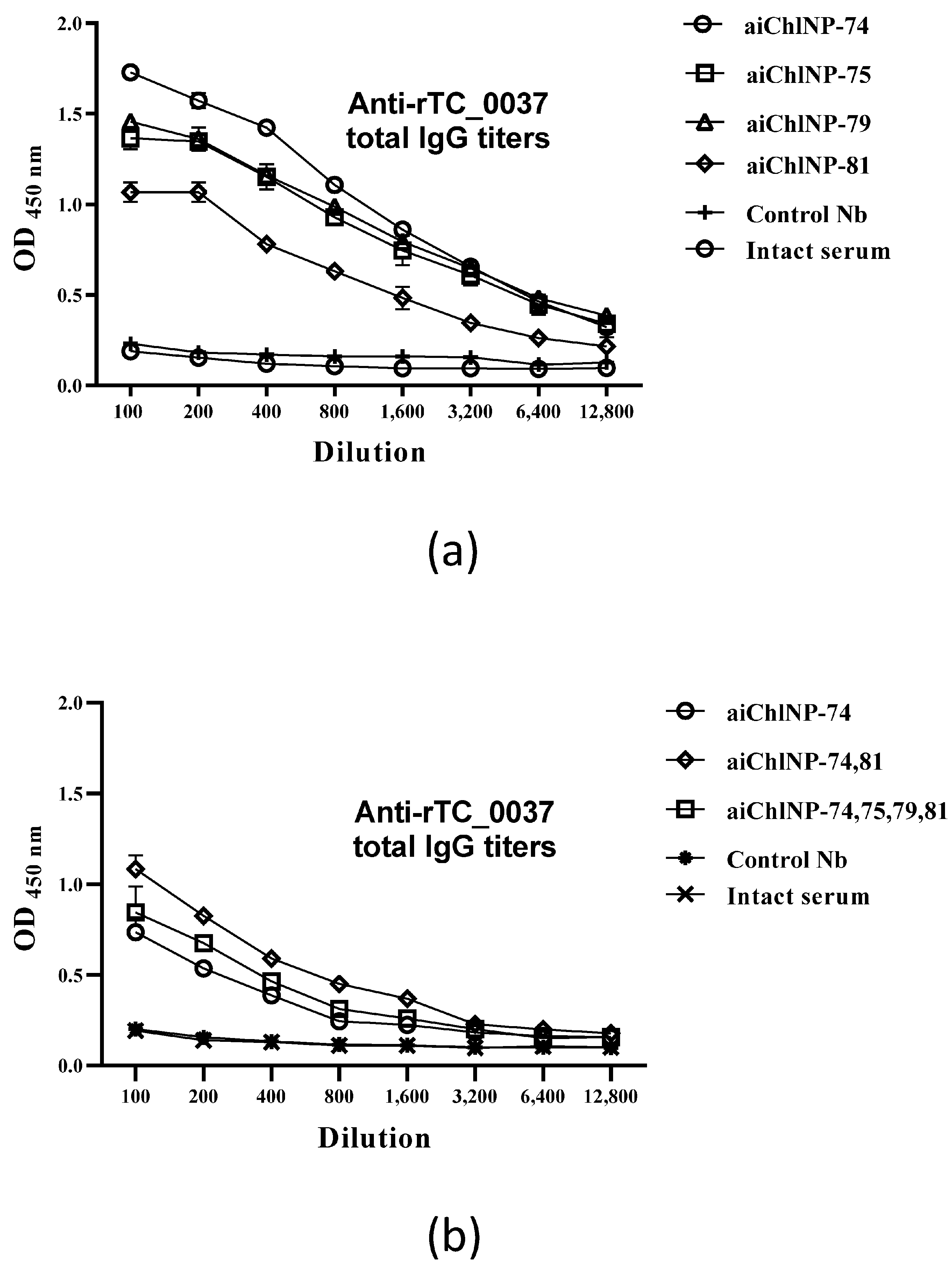
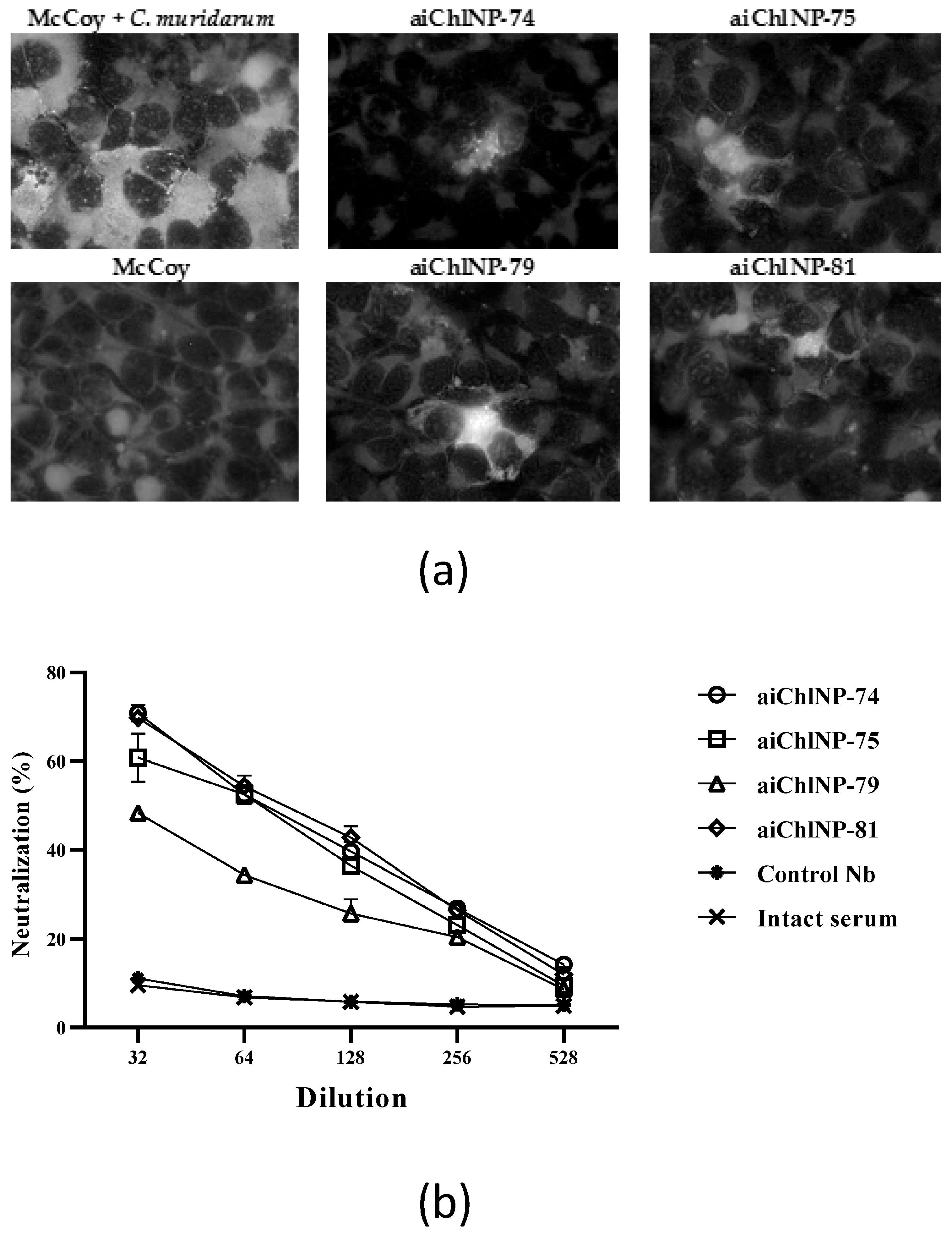
Chlamydia. The immune sera derived from mice immunized with the ai-Nbs were able to suppress chlamydial infection in vitro with slightly varying efficacies. The neutralization results using antibody combinations showed a less than 50% efficacy despite high titers. The most promising ai-Nbs are aiChlNP-74, inducing the highest titer of specific anti-
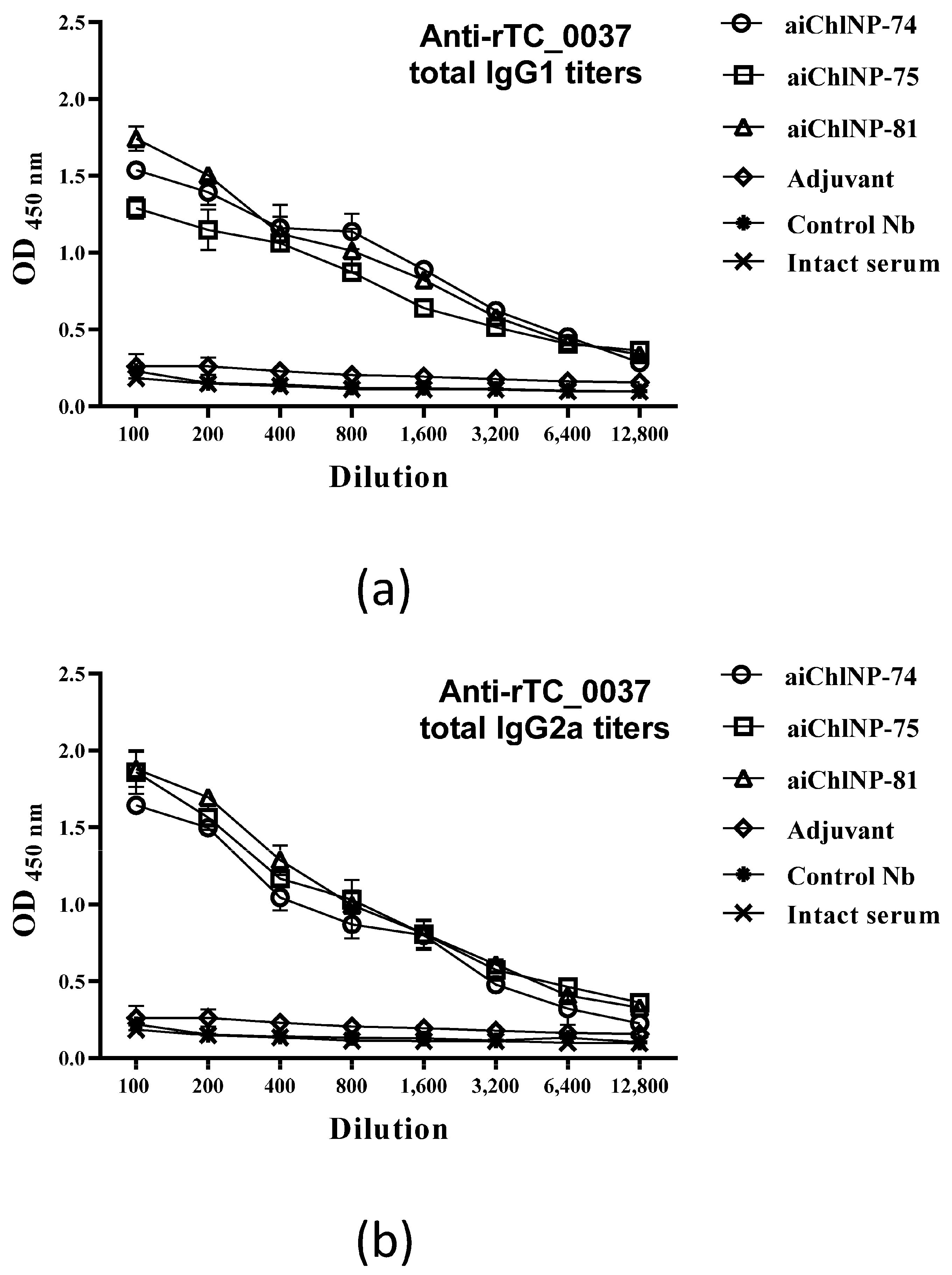
Chlamydia IgG in the mice, and aiChlNP-75 and -81, inducing the formation of IgG with the highest protective effect in the chlamydial infection model in vitro. In addition to the analysis of the humoral immune response, we also obtained preliminary data on a model in vitro indicating also the induction of T-cell-mediated immunity as a result of immunization with the obtained ai-Nbs.
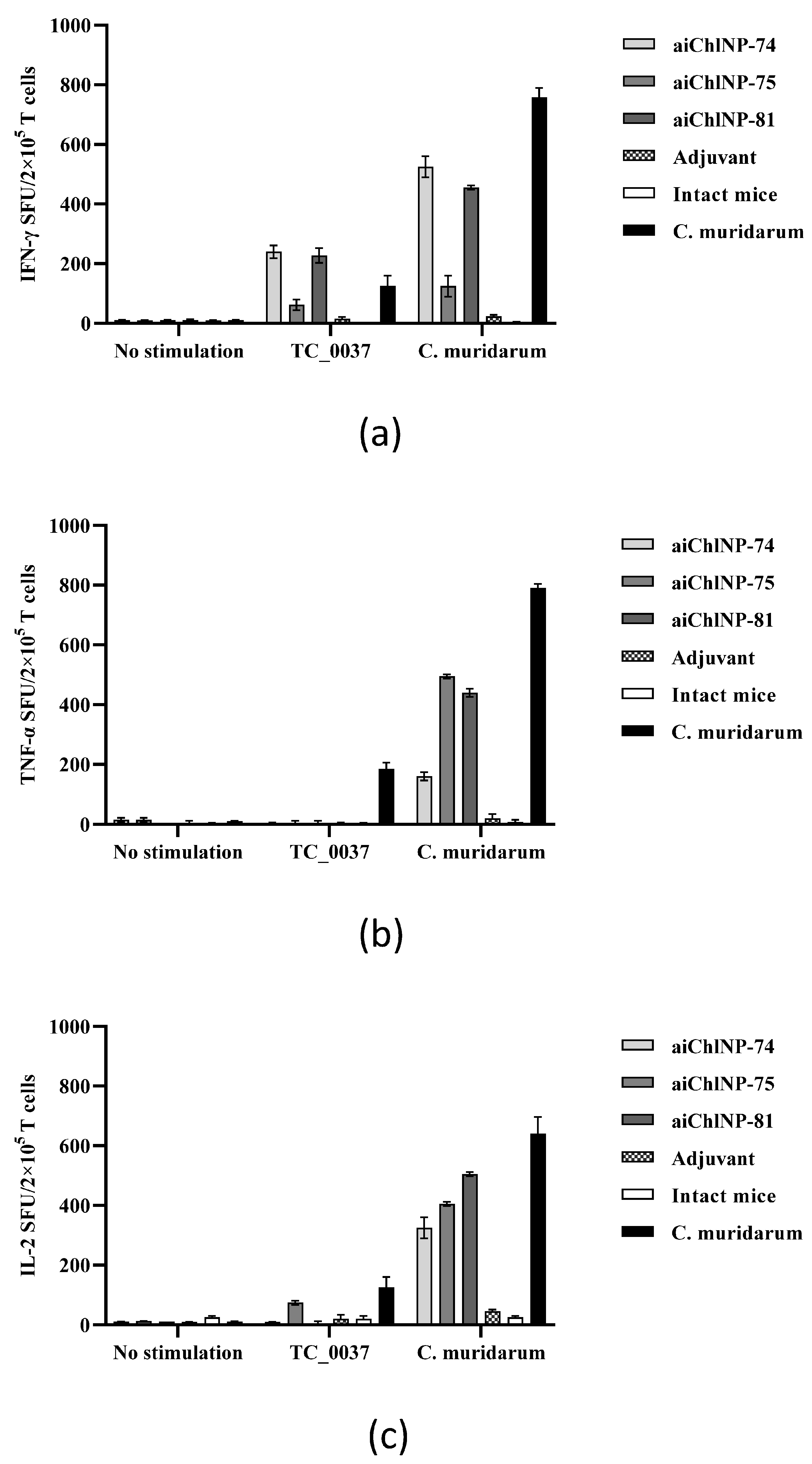
The present study showed that, in response to stimulation with inactivated C. muridarum, aiChlNP-74 and aiChlNP-81 are capable of inducing the specific secretion of proinflammatory cytokines such as IFN-γ and TNF-α. In addition, the correlation of the immune response upon stimulation with C. muridarum with the immune response upon infection with C. muridarum (natural infection) suggests that these ai-Nbs could have therapeutic activity. The T-cells were shown to actively produce TNF-α in response to stimulation with inactivated C. muridarum. As with natural infection, we observed a proinflammatory response in the mice to the inactivated C. muridarum infection. Thus, TNF-α in combination with IFN-γ may act synergistically to mediate pathogen clearance.
We also observed that IL-6 was actively produced in response to
C. muridarum infection, but there was no significant production of chlamydial antigens, which excludes the development of pathological changes as a result of immunization with ai-Nbs. At the same time, the production of the anti-inflammatory cytokines IL-4 and IL-10 by the splenocytes was at the same level as in the intact mice. This is possibly due to the fact that high levels of expression of pro-inflammatory cytokines such as IFN-γ, TNF-α and IL-6 can suppress anti-inflammatory cytokines such as IL-4 and IL-10, and vice versa [
36,
40]. Unlike IFN-γ and TNF-α, IL-2 has no effector function but strongly enhances T-cell expansion [
40,
41,
42]. We have shown that the IL-2 production in response to stimulation with inactivated
C. muridarum is higher than in response to rTC_0037. But at the same time, we observed that immunization with ai-Nbs, especially aiChlNP-75 and -81, leads to an increase in IL-2 expression to the level seen during natural infection. It can be hypothesized that the balance between IL-2 and pro-/anti-inflammatory cytokines is critical for the appropriate initiation and resolution of the immune response during chlamydial infection. Moreover, such an inflammatory reaction, with increased proliferation of the effector cells due to the production of IL-2, will provide more effective protection after immunization with ai-Nbs in response to infection with
C. muridarum. Thus, in our study, we showed the immunogenicity of selected ai-Nbs, which could induce a specific immune response due to the balance of the expression of pro-inflammatory and anti-inflammatory cytokines in response to stimulation with chlamydial antigens, further helping neutralize infections caused by
C. muridarum and its eradication. We suggest further studies on the protective effect of the selected aiChlNP nanobodies be carried out in animal models, such as the already known intravaginal model of chlamydial infection in mice of the DBA/2 line using
C. muridarum [
13].
In this work, we focused our attention on one of the most important virulence factors of
Chlamydia—the type 3 secretion system (T3SS). Recently, T3SS components have very often been used as candidates for vaccination against other pathogenic microorganisms [
8,
9,
10,
11]. The T3SS itself and the injectisome protein TC_0037 (CdsF) are conservative, which is very important when developing vaccine drugs against other bacteria that have the T3SS [
13,
43]. One of the most important treatments for chlamydial infection, as well as many other intracellular pathogens, is to inhibit the pathogen’s invasion and ability to colonize any host organism. We suggest that the mouse antibodies induced by the ai-Nbs obtained in this work bind the injectisome protein TC_0037 and thereby prevent
Chlamydia from infecting other cells. Since the target protein in this case is highly conserved, we assume that the obtained ai-Nbs can be effective in the fight against some other types of
Chlamydia as well. A similar approach can be used to obtain therapeutic nanobodies (ai-Nbs) against T3SS components of other pathogenic microorganisms. In a number of studies, anti-infective nanobodies were obtained that bind to pathogen surface structures, components of secretion and transport systems [
44,
45,
46,
47,
48]. We hope that the ai-Nb-based method described in this paper could be applied to obtaining biomimetics of “sensitive” pathogenic epitopes of these nanobodies, as well to their use in possible therapeutic directed immunomodulation of the host immune system.
We hypothesize that ai-Nbs and their derivatives (for example, when the ai-Nb coding sequences are delivered into the body and expressed as part of an adenoviral or another vector) may be important specific components of a new subunit vaccine. One of the first pieces of evidence of this possibility was an article back in 2009 about an ai-Nb 1HE isolated from a library generated from a Trastuzumab F(ab’)(2)-immunized llama. 1HE has been shown to closely mimic HER2. Serum from BALB/c mice immunized with 1HE contained anti-HER2 antibodies that inhibited the viability of HER2-positive cells [
37]. As a component of a subunit vaccine, ai-Nbs have useful prospects as nanobodies in terms of the very efficient technologies developed for their generation, selection and formatting (modifications and multi-module constructs), for their production and administration, for their safety and for the relatively simplified procedure of their humanization, if required. The lifetime of a nanobody in the bloodstream in case of systemic administration can be adjusted in various ways [
23,
24,
25].
In modern terms, we assume the use of engineered ai-Nbs or their derivatives for the targeted immunomodulation or immunostimulation of the immune cells. Modulating the immune system is a pivotal treatment strategy in modern medicine. Currently, one of the greatest challenges in mAb therapeutics (especially with the systemic use of immunotherapeutic drugs) is their immunogenicity and the formation of anti-drug antibodies which decrease their clinical efficacy [
49,
50]. In the case of ai-Nbs, it is their immunogenicity that is important for the formation of the host’s own therapeutic antibodies. In addition, systemic administration in this case can potentially be replaced by local (subcutaneous) administration.
The subunit vaccines developed thus far have been found to be poorly immunogenic, and thus multiple boosters and suitable adjuvants are necessary to augment their protective potential [
51].
4. Materials and Methods
4.1. Camel Immunization with aChlNP, Construction of a cDNA-VHH Library and the Selection of aiChlNP-Binders
The animal work was approved on 2 February 2018 (registration number 17) by the Commission on Bioethics (formed on 3 May 2017) at the Severtsov Institute of Ecology and Evolution. The camel (
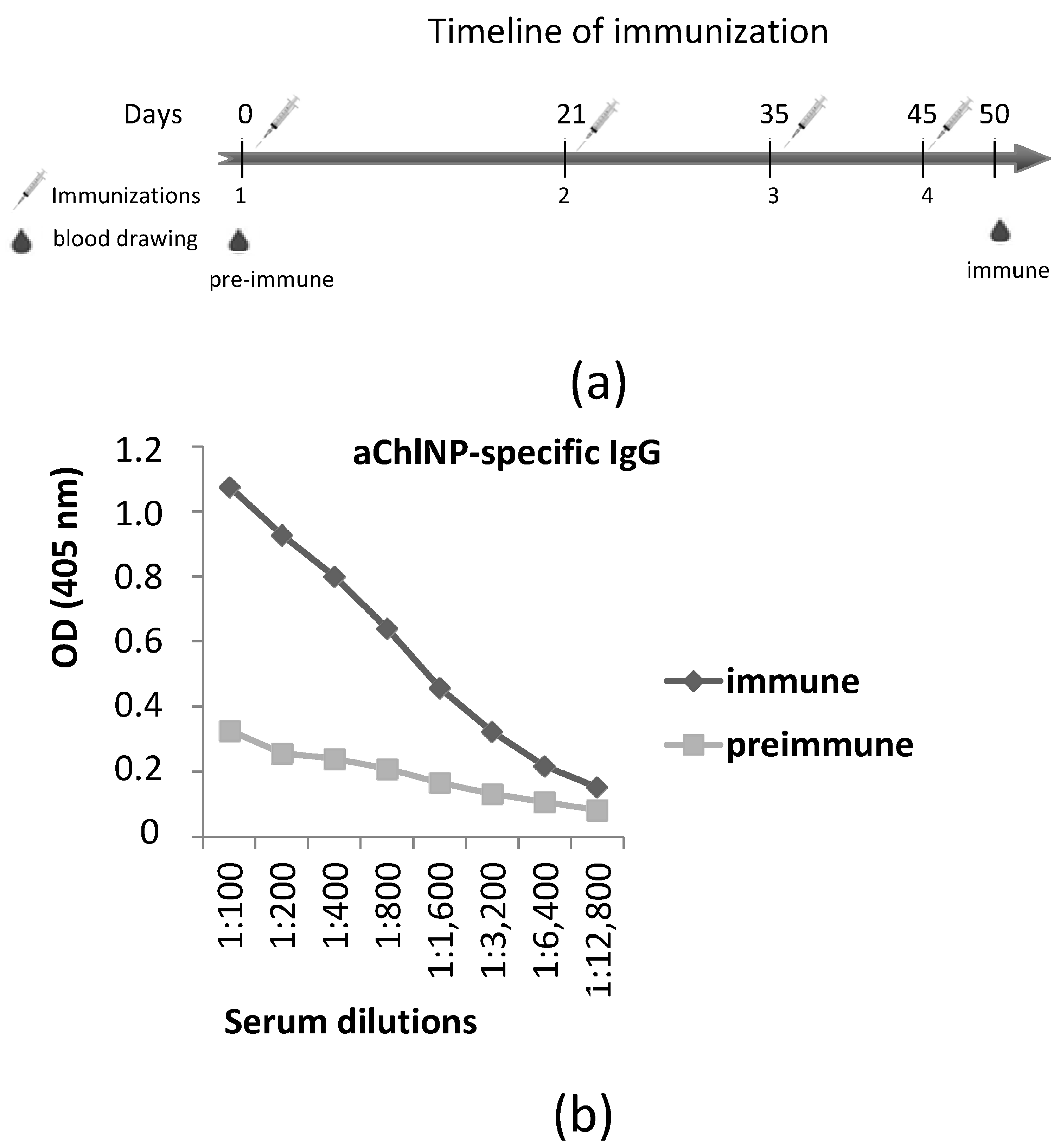
Camelus bactrianus) used for the immunizations was kept at the Center for Collective Use “Live Collection of Wild Mammals”, and the immunizations were performed at the scientific/experimental base “Chernogolovka” of the Severtsov Institute of Ecology and Evolution at the Russian Academy of Sciences (Chernogolovka, Russia). On day 0 before the immunization, blood was taken for later comparisons of the pre-immune serum with the immune serum. The immunization process consisted of 4 subcutaneous injections (each time, the camel was injected at 4–5 sites in the upper body and neck regions) over a time period of 45 days. In each step, 0.9 mg of (aChlNP-ILZ-HH)x3 was used, and equal volumes of the GERBU adjuvant LQ (GERBU Biotechnik, Heidelberg, Germany) were added to the antigen preparations right before injection. Five days after the final immunization (day 50), 150 mL of blood was taken to check for an aChlNP-specific IgG titer and to isolate the peripheral blood mononuclear cells (PBMCs) to construct a cDNA-VHH-library. An equal volume of phosphate-buffered saline (PBS) containing heparin (100 U/mL) and EDTA (3 mM) was added to the blood to prevent clotting. The aChlNP-specific IgG titer was analyzed using ELISA. Briefly, ELISA plate (MaxiSorp, Nunc, Roskilde, Denmark) wells were coated with monomeric aChlNP-HH (2 µg/mL), blocked with 1% BSA and incubated with immune and pre-immune serum diluted 1:100 to 1:12,800 in 1× PBS. Bound IgG was detected using HRP-conjugated rabbit anti-camel IgG serum (previously obtained by Tillib et al. [
34]) and ABTS. The optical density (OD) was measured at 405 nm using a microplate fluorometer.
The preparation of a VHH-cDNA-library and the selection of aChlNP-specific binders using phage display was performed as previously described [
33,
34]. Nanobodies with different (non-Chlamydial) recognition specificity containing the same conserved framework and amino acid sequences added to their C-terminus as in the case of formatted aChlNPs (e.g., trimerized Nbs [
34], available in our laboratory), were used in selection procedures for initial subtraction of nonspecifically bound sticky VHH-phage particles, and then as competitors or blockers of interactions other than anti-idiotype (aiChlNP) with idiotype (aChlNP).
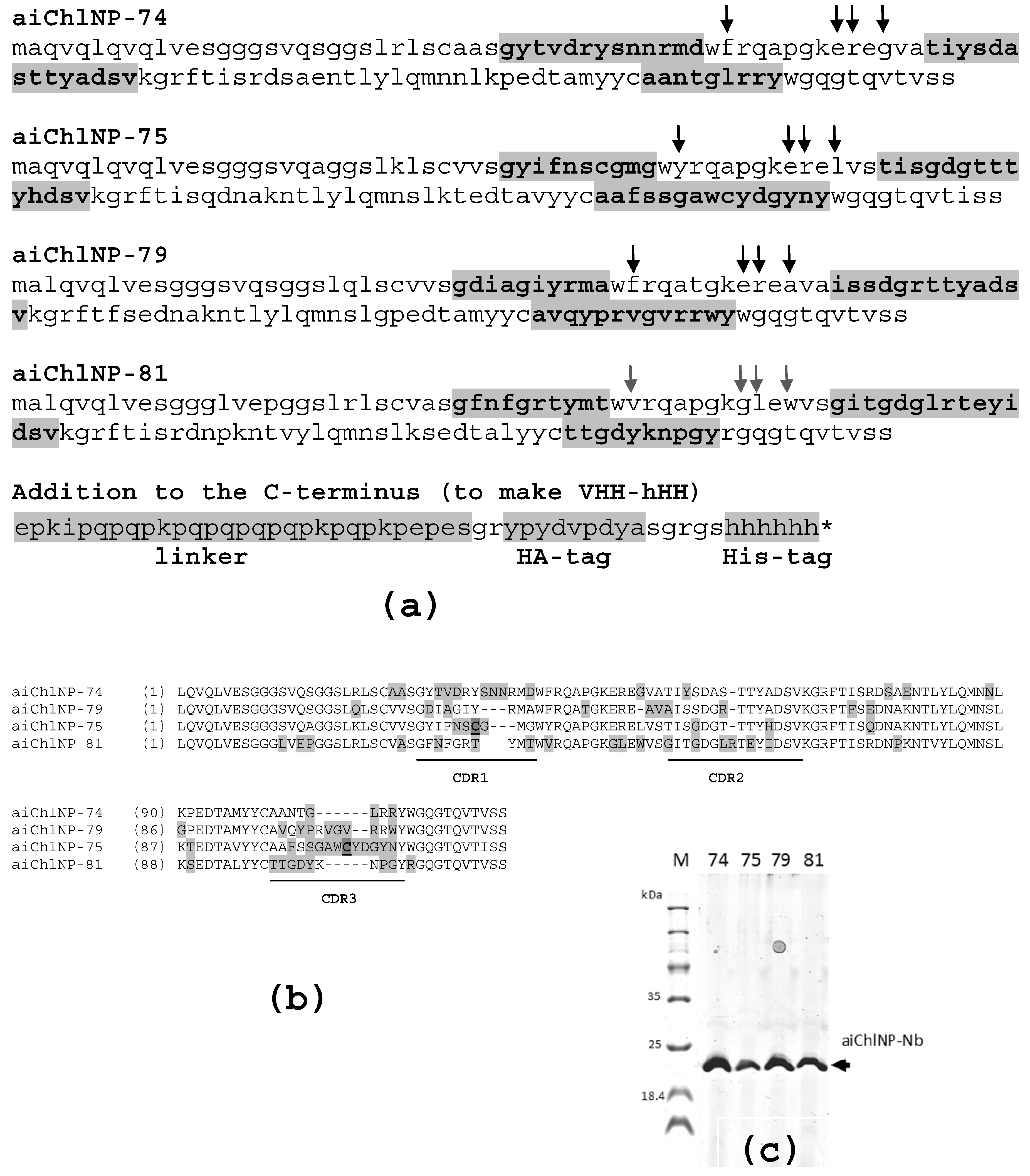
The recombinant protein TC_0037 was used at a high concentration (1 mg/mL) for affinity elution of the VHH phage particles bound to the immobilized aChlNP nanobody at the final stage of the panning procedure. A total of 4 different ai-Nb variants that showed the strongest aChlNP-specific reactivity in ELISA were selected from the original enriched 60 clones. were selected from the original enriched 60 clones.All four selected ai-Nb variants competed with rTC_0037 in binding to the immobilized primary aChlNP nanobody. The selected VHH-DNA sequences (aiChlNP-74, -75, -79 and -81) were subcloned into the expression vector pHEN6 [
52] containing the pelB leader sequence for periplasmic expression. Initially selected nanobodies were subjected to formatting. For this purpose, a long 28-amino acid linker (upper hinge region of camel IgG), an additional trimerizing isoleucine zipper (ILZ) domain, HA-tag and His-tag (for detection and purification purposes) were added after the antigen-binding domains at their C-terminus as described previously [
34].
4.2. Expression and Purification of the Formatted Nanobodies
E. coli XL1-blue competent cells (Agilent Technologies, Santa Clara, CA, USA) were transformed with the pHEN6 plasmid encoding the formatted nanobody sequence and grown on LB agar plates with 100 µg/mL ampicillin and 1% glucose. Single colonies were picked and inoculated in 4 mL 2× YT medium with 100 µg/mL ampicillin and 1% glucose at 37 °C. The overnight cultures were transferred into 250 mL of fresh medium (with 60 µg/mL ampicillin and 0.1% glucose) and grown until an OD600 nm of 0.5–0.7 was reached. Expression was induced with 0.2 mM of IPTG overnight at 28 °C. The cultures were centrifuged (3000× g, 15 min, 4 °C), and the soluble nanobodies were extracted from the periplasm by resuspending and incubating the pellets with 4 mL of TES buffer (50 mM Tris (pH 8.0), 0.5 mM EDTA, 20% saccharose, 10 mM imidazole, 100 µg/mL PMFS, 5 mM β-mercaptoethanol) for 30 min on ice and subsequently adding 6 mL of Solution 2 (10 mM Tris (pH 8.0), 10 mM imidazole, 1 mM MgCl2) for 30 min on ice. The suspension was centrifuged (16,000× g, 30 min, 4 °C) and the supernatant purified using HIS-Select® Nickel Affinity Gel (Sigma-Aldrich, St. Louis, MO, USA). The purified nanobodies were dialyzed against 1× PBS containing 10 mM of imidazole and then analyzed using 14% SDS-PAGE under reducing conditions, after which the proteins in the gel were stained with Coomassie Brilliant Blue.
4.3. Reactivity of aChlNP to Recombinant TC_0037
A total of 5 µg/mL of rTC_0037 and the control protein BSA were coated onto ELISA plates for one hour at 37 °C. The wells were washed with 1× PBS containing 0.05% Tween20 (1× PBST), saturated with 1% BSA in 1× PBST and incubated with the purified formatted nanobodies, monomeric aChlNP-HH or trimerizing aChlNP-ILZ-HH in a series of 4-fold dilutions (from 2.4 to 0.04 µg/mL) in 0.1% BSA/1× PBST for one hour at 37 °C. The wells were washed, and bound nanobodies were detected using a mouse HRP-labeled anti-HA-tag antibody (1:4000 in 0.5% BSA/1× PBST). A color reaction was achieved using the HRP substrate ABTS (1 mg/mL, Sigma-Aldrich). The ODs were measured at 405 nm (with a reference wavelength of 495 nm) using a Tecan Infinite F50 microplate reader (Männedorf, Switzerland) and are shown as means of triplicates ± standard deviation (SD).
4.4. Bacteria, Growth Conditions and Cell Lines
C. muridarum (Nigg strain) ATCC VR-123 was grown in cycloheximide-treated McCoy cells (a hybrid cell line consisting of human synovial cells and mouse fibroblasts) as described previously [
43,
53]. The chlamydial elementary bodies (EBs) were collected, purified, quantified and stored at −70 °C in SPG buffer (sucrose/phosphate/glutamic acid: 0.2 mM sucrose, 20 mM sodium phosphate and 5 mM glutamic acid).
4.5. Production of Recombinant Protein TC_0037 TTSS of C. muridarum
As an antigen for selection, we used the recombinant protein TC_0037 produced in
E. coli, which corresponds to the conservesd protein of the secretion system of the third type (TTSS) in
C. muridarum [
13].
4.6. Mice
Female mice DBA/2, 8–9 weeks old, free from specific pathogens, were bred and kept in the vivarium of N.F. Gamaleya National Research Center for Epidemiology and Microbiology (Moscow, Russia). The animals were used in accordance with the recommendations of the national guidelines, and the methods of the experiments were approved by the N.F. Gamaleya National Research Center Animal Care Committee.
4.7. Immunofluorescence Method for Detecting Intracellular Chlamydia
The eukaryotic cell culture McCoy was infected with
C. trachomatis or
C. muridarum as described earlier [
43]. A 24 h monolayer of cells was infected with the chlamydia strain by adding a chlamydia culture to the culture medium, followed by centrifugation. The cells were incubated at 37 °C for 48 h and then fixed with acetone and incubated with 5% ovalbumin to prevent non-specific binding. The ability of the nanobodies to bind to chlamydia, which forms intracellular inclusions in eukaryotic cells, was tested using an immunofluorescence assay according to standard protocol [
45]. Fixed uninfected cells were used as the control. The detection of bound antibodies was performed using mouse anti-HA antibodies and fluorescein isothiocyanate (FITC)-conjugated secondary antibodies to mouse immunoglobulins.
4.8. Immunization of the Mice with Selected ai-Nbs
The DBA/2 mice (6 mice/group) were subcutaneously injected with a series of trimerized ai-Nbs (aiChlNP-74, -75, -79 and -81), either alone or in combination in total volumes of 200 and 400 μL/mouse with Freund’s adjuvant (1:1). The concentrations of the ai-Nbs before they were mixed with the adjuvant were around 1 mg/mL. The immunization was performed three times and four times with an interval of 14 days. The first immunization was performed with complete Freund’s adjuvant (CFA), and subsequent immunizations were performed with incomplete IFA. A total of 14 days after the last immunization, blood was collected from the mice to obtain sera. The blood was centrifuged, and the sera were obtained and stored at −20 °C. To assess the immunogenicity of the aiChlNP nanobodies, an enzyme-linked immunosorbent assay (ELISA) was performed to detect antibodies specific to the rTC_0037 protein and to analyze the neutralizing activity of the obtained antibodies. Pre-immune sera obtained from intact mice were used as negative controls.
To evaluate specific antibodies of IgG isotypes (IgG1 and IgG2a) and the cellular immune response, immunization with the most immunogenic ai-Nbs (aiChlNP-74, -75 and -81) was performed. Four stages of immunization were carried out with an interval of 2 weeks. The anti-idiotypic nanobodies aiChlNP-74, -75 and -81 were injected subcutaneously into the DBA/2 mice in combination with Freund’s adjuvant (as described previously), while the last immunization was performed using FAMA (GERBU Biotechnik) as the adjuvant to achieve the best effect using small volumes when administered to animals. Intact mice injected with PBS with Freund’s and FAMA adjuvants were used as the negative controls. For the evaluation of the IgG1- and IgG2a-specific antibodies, mouse sera were obtained after 4-fold immunization. Mouse spleens were used to evaluate the IFN-γ-specific T-cell immune response after immunization. The spleen cells (2 × 105 cells/mL) were incubated with UV-inactivated C. muridarum and rTC_0037 for 24 h in complete RPMI-1640 medium (Gibco, Carlsbad, CA, USA) containing 5% fetal bovine serum, 2 mM of L-glutamine and 1% penicillin-streptomycin (Gibco, Carlsbad, CA, USA). Spleens from mice infected intravaginally with 104 IFU/mL C. muridarum were used as the positive controls. Spleens from mice immunized with adjuvants and from the intact mice were used as negative controls.
4.9. Indirect Enzyme-Linked Immunosorbent Assay (ELISA) Method
The ELISA method was used to determine the specific IgG antibodies as well as the IgG1 and IgG2a isotypes in the serum of the mice. For the assay, 96-well microtiter ELISA plates (Nunc, Rochester, NY, USA) were used. The rTC_0037 protein was diluted to a concentration of 10 μg/mL in PBS, applied in a volume of 100 μL into the wells of the plates, after which an incubation was carried out overnight at 4 °C. After incubation, the plates were washed several times with PBS, and non-specific binding was blocked using 0.1% bovine serum albumin (BSA) in PBS for 30 min at room temperature. The mouse sera obtained after immunization were titrated using 0.1% BSA in PBS for dilution and then incubated overnight at 4 °C. The plates were then washed 3 times with 0.1% BSA in PBS. Biotinylated anti-mouse IgG monoclonal antibodies (IgG1 and IgG2a) (BioLegend, San Diego, CA, USA) used at a dilution of 1:1000 were added to the wells of the plates and incubated for 1 h at room temperature (20 °C), after which the streptavidin HRP dye (BioLegend, San Diego, CA, USA) and TMB substrate (BioLegend, San Diego, CA, USA) were added. The reaction was stopped after 20 min according to the addition of 50 μL/well 1 M H2SO4. The absorbance at 450 nm was determined using a Multiskan EX microplate reader (Thermo Fisher Scientific Oy, Vantaa, Finland).
4.10. In Vitro Neutralization Assay
The neutralization assays of
C. muridarum were performed in the McCoy cell culture in 24-well culture plates with glass slides (12 mm diameter), as described in [
13]. The immune mice sera were diluted in Dulbecco’s modified Eagle medium, DMEM (PanEco, Moscow, Russia). To 100 μL of the serial dilutions of the sera (1:32–1:528), 100 μL of the
C. muridarum EB suspension in DMEM medium (1.4 × 10
5 IFU/mL) was added for 30 min at 37 °C on a shaker at 150 rpm. The samples incubated with EB without sera were used as the infection controls. After incubation, suspensions of 100 μL/well (in duplicate) diluted with 900 μL of DMEM containing 10% fetal bovine serum and 1 mg/mL cycloheximide were added to McCoy cells in 24-well culture plates.
The plates were centrifuged at 800× g for 1 h and then incubated at 37 °C for 48 h. Next, the cells adsorbed onto the glass were fixed with ethanol and stained with monoclonal antibodies conjugated with fluorescein isothiocyanate (FITC) against Chlamydia lipopolysaccharides (Nearmedic Plus, Moscow, Russia). The cells containing inclusions were analyzed using a Nikon Eclipse 50i fluorescence microscope (Nikon, Amsterdam, The Netherlands) at 100× magnification and counted to determine the percentage of infected cells in the McCoy monolayer.
4.11. ELISPOT Assay
The C. muridarum- and TC_0037-specific cytokine-producing T-cell immune response was assessed using ELISPOT. The assay was performed using a commercial IFN-γ ELISPOT Ready-SET-Go! kit (eBioscience, Inc., San Diego, CA, USA). For the assay, 96-well MultiScreen-IP filter plates (Millipore, Billerica, MA, USA) with immobilized IFN-γ antibodies plated onto them were used. The plates were incubated overnight at 4 °C. After incubation, the plates were washed with sterile PBS. T-cells were isolated from the mouse spleens using T-cell separation columns (Cederlane, Burlington, ON, Canada) according to the manufacturer’s protocols. Enriched cell suspensions were added to the plates at a concentration of 2 × 105 cells per well (in 100 μL of DMEM culture medium) in the presence of the UV-treated C. muridarum EB at a final concentration of 104 IFU/mL and rTC_0037 at a final concentration of 10 μg/mL. The plates were incubated for 24 h at 4 °C. Next, washing with PBS and incubation with the biotinylated anti-IFN-γ monoclonal antibodies was performed. Stains (spots) were visualized using Avidin HRP dye and a freshly prepared 3-amino-9-ethylcarbazole (AEC) substrate solution. The spots were quantitatively counted using the AID EliSpot Reader (Autoimmun Diagnostika GmbH, Strassberg, Germany). ELISPOT was carried out similarly using kits for TNFα, IL-6 and the inflammatory cytokines IL-4, IL-10 and IL-2 (eBioscience, Inc., San Diego, CA, USA).


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






